Introduction
Ovarian cancer is one of three major female
malignant tumors of the reproductive system. Its incidence is
ranked third following cervical and uterine cancer (1). Due to its high malignant potential,
the mortality rate of ovarian cancer ranks first among all
gynecologic cancers in China. To date, the pathogeny of ovarian
cancer is unclear. It is believed that the incidence may be related
to environment, hormonal, genetic and other factors. According to
different clinical pathological types and genetic features, ovarian
cancer can be divided into type I and type II (2). The ovarian malignant progression of
type I is low, including low grade serous ovarian cancer, mucinous
ovarian cancer and clear cell ovarian cancer. The mutated BRAF or
PTEN gene mainly exhibits abnormal expression in this type of
ovarian cancer (3). Type II ovarian
cancer has a high degree of malignancy, in which BRCA1 and BRCA2
gene mutations are common and the main pathological type is
malignant serous ovarian cancer (4–6).
At present, the scientific community believes that a
tumor is a disease caused by multiple genomic changes, and tumor
progression is based on oncogene activation and/or tumor-suppressor
gene deactivation with gene function mutation. Continual growth
signal stimulation, evasion of growth inhibition, resistance to the
replication of programmed cell death, induction of tumor
angiogenesis, and activation of invasion and metastasis are
necessary physiological changes leading to different types of
malignant tumor cell growth (7).
During development, most tumors undergo similar changes in cell
physiological characteristics, thus oncogenes and tumor-suppressor
genes that are closely associated with these acquired cell
behaviors have become a ‘hot’ topic in the study of tumor molecular
mechanisms and cancer treatment.
Cyclin-dependent kinase 3 (CDKN3), a member of the
family of protein phosphatase inhibitors, participates in the
management of the cell cycle, where it plays a dual role (8,9). CDKN3
plays the role of a cell cycle protein-dependent kinase inhibitor
and selectively combines with CDK2 (10,11),
reducing Rb protein (retinal cell tumor protein, a tumor-suppressor
gene) phosphorylation (9).
Phosphorylation of Rb protein combined with transcriptional factor
E2F1 inhibits cell cycle protein, which is required for G1/S
transition, thus blocking G1 to S phase shift (12,13).
CDKN3, also a type of Mdm2 protein, combines with p53 and Mdm2,
reduces the sensitivity of p21 and reduces p53 target gene
production, so as to promote the progression of the cell cycle
(14). The CDKN3 gene was
demonstrated to exhibit abnormal expression in many types of tumors
(15–17). However, its relationship with
ovarian cancer and its molecular role has not been reported.
Materials and methods
Patient tissue samples
Ninety-seven ovarian cancer (OC) tissues were
collected from patients who underwent routine ovarian resection at
The First Affiliated Hospital of China Medical University. None of
the specimens were pretreated with preoperative chemotherapy,
radiation therapy, or other treatment. All specimens were
respectively assessed by two pathologists for pathological
diagnosis according to the World Health Organization classification
system. The clinicopathologic factors are documented in Table I. Total RNAs and protein were
collected from the fresh OC tissues after surgical resection. The
study protocol was approved by the Institutional Ethics Committee
of China Medical University, and all patients provided written
informed consent.
 | Table ICorrelation of CDKN3 expression and
the clinicopathological characteristics of the EOC patients. |
Table I
Correlation of CDKN3 expression and
the clinicopathological characteristics of the EOC patients.
| CDKN3 expression | |
|---|
|
| |
|---|
| Characteristics | High n (%) | Low n (%) | P-value |
|---|
| Total cases | 66 (68.0%) | 31 (32.0%) | |
| Age (years) | | | 0.216 |
| ≥58 | 16 | 8 | |
| <58 | 50 | 13 | |
| Histological
type | | | 0.879 |
| Serous | 52 (68.4%) | 24 (31.6%) | |
| Mucinous | 14 (66.7%) | 7 (33.3%) | |
| Residual tumor
size | | | 0.021 |
| <1 cm | 22 (55.0%) | 18 (45.0%) | |
| ≥1 cm | 44 (77.2%) | 13 (22.8%) | |
| FIGO stage | | | 0.020 |
| I/II | 20 (54.1%) | 17 (45.9%) | |
| III/IV | 46 (76.7%) | 14 (23.3%) | |
| Grade | | | 0.126 |
| G1 | 20 (57.1%) | 15 (42.9%) | |
| G2 | 13 (65.0%) | 7 (35.0%) | |
| G3 | 33 (78.6%) | 9 (21.4%) | |
| Recurrence | | | 0.017 |
| Yes | 58 (73.4%) | 21 (26.6%) | |
| No | 8 (44.4%) | 10 (55.6%) | |
| Serum CA-125 | | | 0.199 |
| <35 U/ml | 27 (61.4%) | 17 (38.6%) | |
| ≥35 U/ml | 39 (73.6%) | 14 (26.4%) | |
| Metastasis | | | 0.610 |
| Yes | 27 (71.1%) | 11 (28.9%) | |
| No | 39 (66.1%) | 20 (33.9%) | |
Immunohistochemistry
CDKN3 expression was evaluated by
immunohistochemistry (IHC) on 5-μm paraffin-embedded tissue
sections. Rabbit anti-CDKN3 (1:200; Santa Cruz Biotechnology, Santa
Cruz, CA, USA) was used. Sections were stained with
3,3′-diaminobenzidine. Normal rabbit serum was used as a negative
control. The CDKN3 immunostaining score was analyzed according to a
semi-quantitative scale. Cytoplasmic/nuclear immunostaining was
considered positive staining. The percentage of positive cells was
scored as follows: 0 (0%), 1 (1%–10%), 2 (11%–50%), 3 (51%–80%) or
4 (>80%). The intensity of staining was defined as follows: no,
‘0’; weak, ‘1’; moderate, ‘2’ and strong staining, ‘3’. Each sample
was given a final score ranging from 0 to 12. CDKN3 expression was
divided into four levels: −, score 0; 1+, score 1–4; moderate
expression 2+, score 4–8; and 3+, score 9–12. The
immunohistochemical results of CDKN3 were grouped into low
expression (0 to 1+) and high expression (2+ to 3+).
Western blot analysis
Total protein was separated from tissues and cells
using lysis buffer (Pierce, Rockford, IL, USA) and quantified by
the Bradford method. Total protein (50 μg) was subjected to 10%
SDS-PAGE and electrotransferred to PVDF membranes (Millipore,
Billerica, MA, USA), which were blocked with blocking buffer
(Beyotime, China) for 1 h at room temperature. The primary CDKN3
rabbit polyclonal antibody (1:1000; Santa Cruz Biotechnology, USA)
and mouse anti-β-actin antibody (1:1000; Santa Cruz Biotechnology)
were incubated on the PVDF membranes at 4°C overnight. The
membranes were incubated with secondary antibodies for 2 h at room
temperature (1:5000; Beyotime). The relative protein levels were
visualized using an ECL system (Pierce).
Cell culture and siRNA treatment
OVCAR3 cancer cells were obtained from the Cell
Biology China Academy of Science (Shanghai, China) and cultured in
RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) containing 10%
fetal calf serum. The cells were cultured in a 24-well plate at a
density of 6×104 cells/well and transfected with
siRNA-CDKN3 or Neg. siRNA using the Lipofectamine LTX reagent
(Invitrogen) according to the manufacturer’s protocol. CDKN3 siRNAs
(purchased from GenePharma Co., Ltd., Shanghai, China) were as
follow: CCAUCAAGCAAUACAAUUATT (si-CDKN3#1) and
CUGCUUGUCUCCUACUAUATT (si-CDKN3#2).
Colony formation and MTT assay
For the colony formation assay, 100 cells were
plated into 6-well plate culture dishes and incubated for 10 days.
Plates were stained with 0.1% crystal violet, and colonies with
more than 50 cells were counted. For the MTT assay, cells were
plated at ~2000 cells/well in 96-well plates in medium containing
10% FBS 24 h after transfection. Then, 20 μl of 5 mg/ml MTT (KeyGen
Bio., Nanjing, China) solution was added to each well and incubated
for 4 h at 37°C. The media were removed from each well, and the
resultant MTT formazan product was solubilized in 150 μl of DMSO.
The results were quantitated using a test wavelength of 490 nm.
Cell cycle analysis
OVCAR3 cells were plated in a 6-well plate at a
density of 1×105 cells/well. Cells were collected after
24 h and washed in cold PBS, and fixed in 75% cold ethanol. Then
the cells were centrifuged at 800 rpm and washed with cold PBS.
RNase A and propidium iodide solution were added, and the cells
were incubated at room temperature for 30 min in the dark. Cell
cycle analysis was performed in a flow cytometer (FACSCalibur; BD
Biosciences, USA), and ModFit LT software was used to analyze the
percentage of cells in each cell cycle phase.
Transwell assay
OVCAR3 cells (5×104), with 200 μl serum-
free RMPI-1640, were added to the upper layer which was coated with
20 μl Matrigel (1:4 dilution; Costar, Corning, NY, USA). RMPI-1640
containing 10% FBS was added to the lower layer, and incubation was
carried out for 24 h in a cell incubator. The cells that penetrated
the membrane of the chamber were stained with 0.1% crystal violet
for 20 min. Cells on the upper membrane were wiped off with a
cotton tip. The number of invasive cells were analyzed in 5 random
fields under a microscope. The experiments were performed in
triplicate.
Statistical analysis
The statistical data were analyzed by SPSS 13.0. The
correlation between CDKN3 expression and EOC patient
clinicopathological features was analyzed using the χ2
test. The t-test was used to compare messenger RNA (mRNA) in
various groups. Kaplan-Meier method was used to evaluate the
patient overall survival. A Cox regression model was used to
demonstrate the univariate and multivariate analyses of prognostic
variables. P<0.05 was considered to indicate a statistically
significant result.
Results
CDKN3 expression in the EOC tissue
samples and cell lines
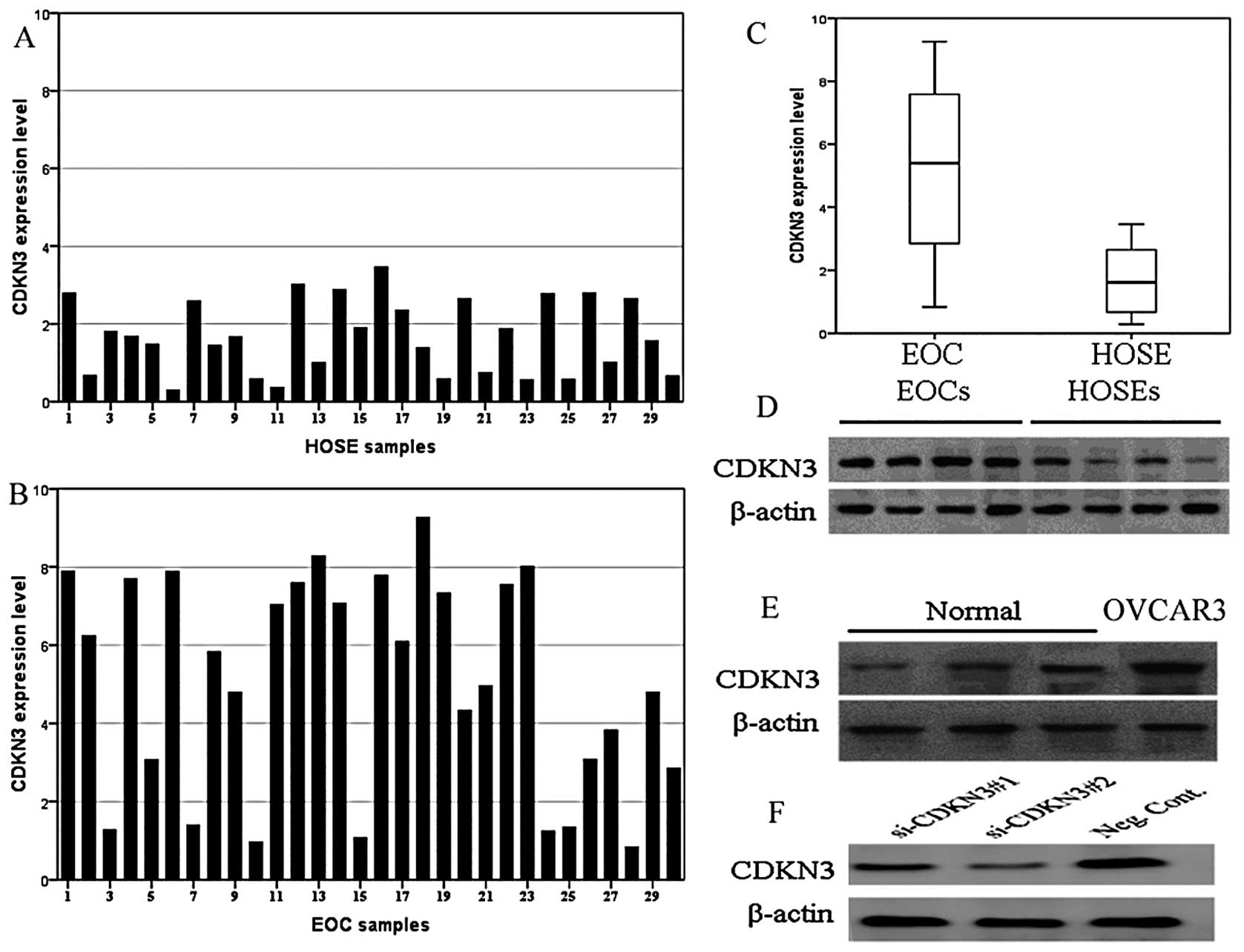
CDKN3 protein levels in 30 EOC and 30 HOSE tissues
were determined and the results showed that CDKN3 was expressed
weakly in the HOSE tissues (20/30), while CDKN3 expression was
upregulated >2-fold in 76.7% (23/30) of the EOC tissues
(EOC/HOSE >2) (Fig. 1A and B).
The mean level of CDKN3 expression in the EOC tissues was 3.35-fold
that in the HOSE tissues (Fig. 1C).
Western blotting analysis revealed that expression of the CDKN3
protein was markedly higher in the EOC tissues and the OVCAR3 cell
line when compared with the levels in the HOSE samples (Fig. 1D and E). In addition, CDKN3 protein
expression in OVCAR3 cells transfected with the CDKN3 siRNA showed
efficient depletion (Fig. 1F). As
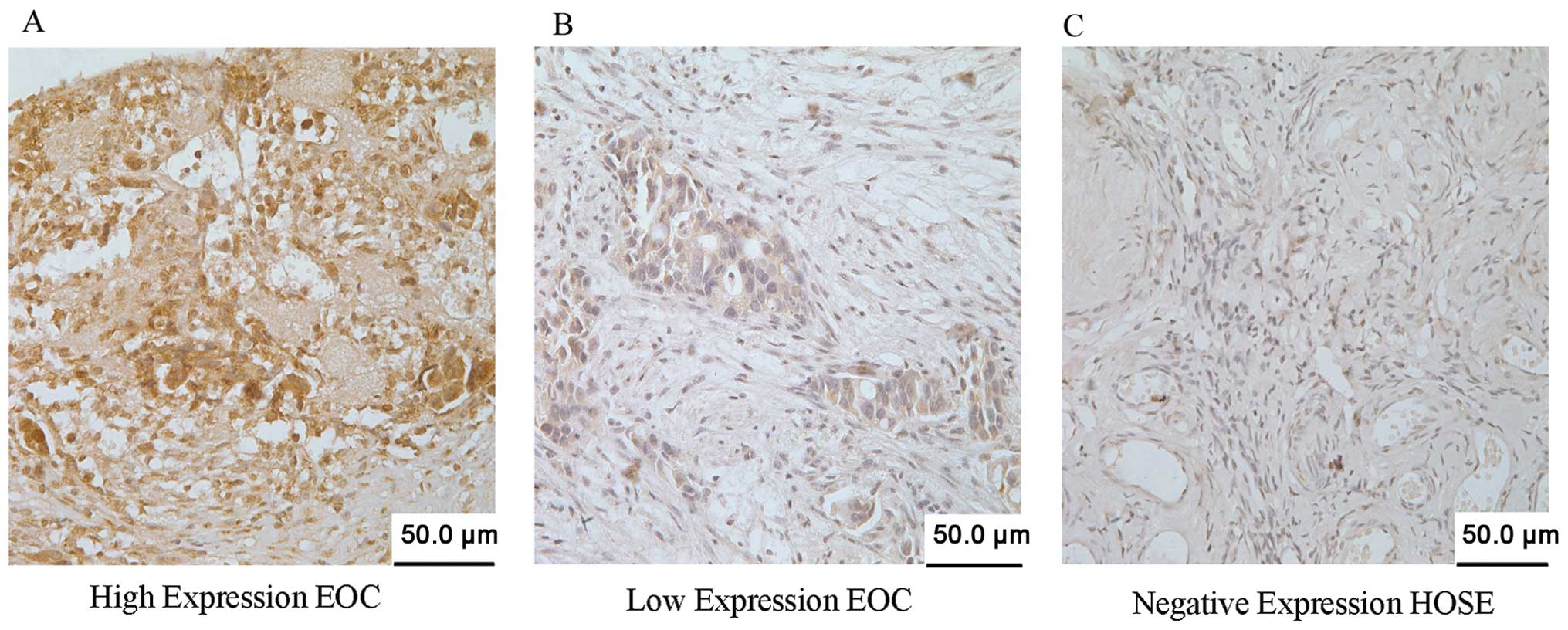
shown in Fig. 2, the IHC staining
intensity and positivity of CDKN3 in the representative tissues
were highly expressed (66/97, 68.0%) and weakly expressed (31/97,
32.0%; Fig. 2A and B), while
negative expression was noted in the HOSE tissues (Fig. 2C).
Correlation of the clinical
characteristics and CDKN3 expression in the EOC cases
CDKN3 expression levels and clinicopathological
characteristics of the EOC patients are summarized in Table I. High expression of CDKN3 was
detected in 68.0% (66/97) of the patients and low expression of
CDKN3 was noted in 32.0% (31/97) of the EOC patients. High levels
of CDKN3 expression were significantly associated with FIGO stage,
recurrence and residual tumor size, but no significant correlation
was noted with patient age, tumor type, serum Ca-125 level and
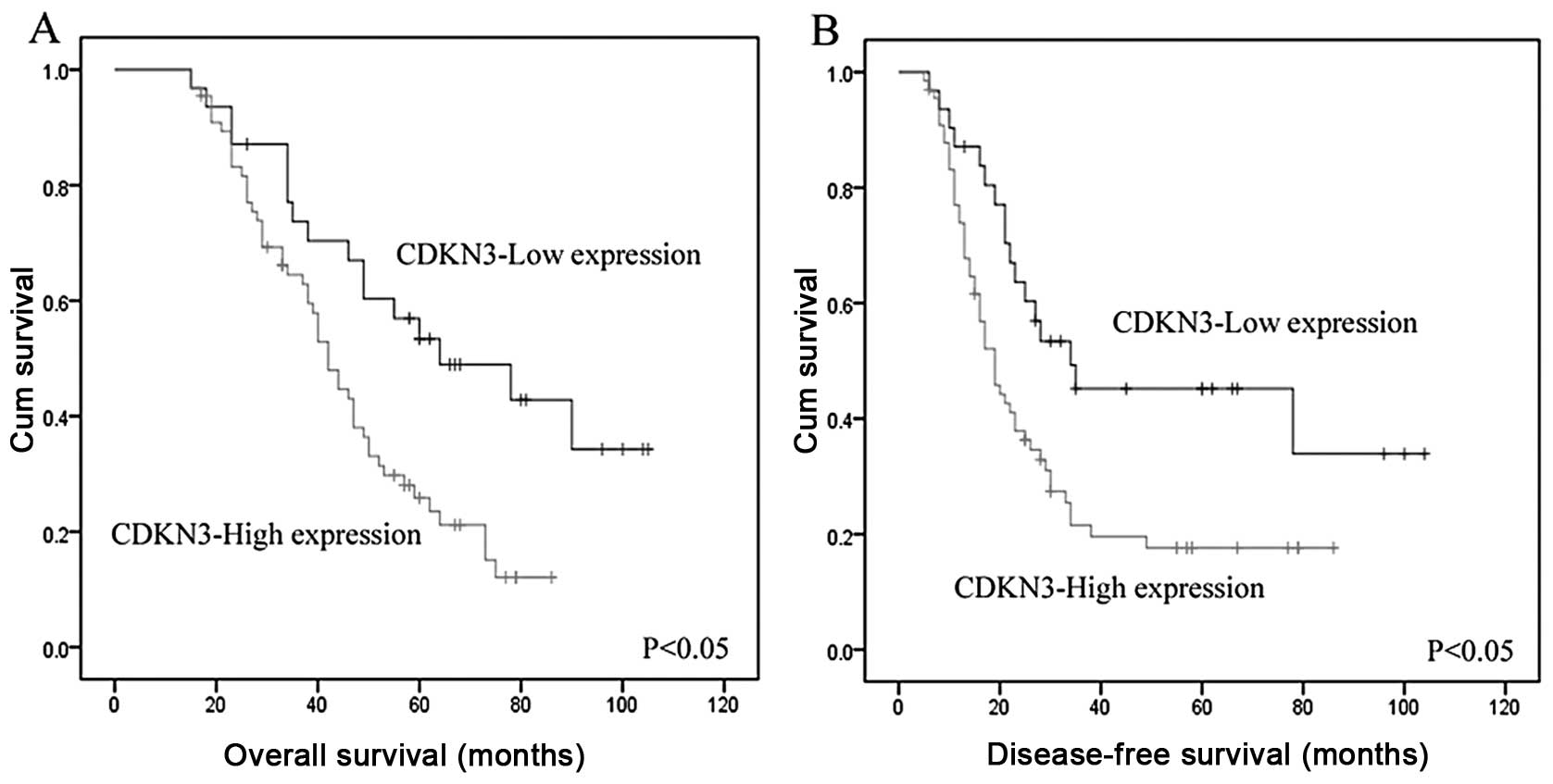
histological type. Overall survival and disease-free survival were
significantly reduced in patients with high CDKN3 expression than
in patients with low CDKN3 expression (P<0.05, Fig. 3A and B). Furthermore, multivariate
analysis demonstrated that a high level of CDKN3 expression was an
independent predictor of prognosis of EOC patients, and was
associated with increased risk of a poor prognosis (hazard ratio,
1.661; P=0.005) (Table II).
| Table IIUnivariate and multivariate analyses
of factors correlated with overall survival of the EOC
patients. |
Table II
Univariate and multivariate analyses
of factors correlated with overall survival of the EOC
patients.
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|---|
| Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| CDKN3 | 3.264 | 1.781–5.983 | <0.001a | 1.661 | 1.324–4.351 | 0.005a |
| Age | 0.883 | 0.634–1.598 | 0.775 | | | |
| Serum Ca-125
level | 1.645 | 0.837–1.933 | 1.259 | | | |
| FIGO stage | 5.234 | 2.871–9.542 | <0.001a | 4.730 | 2.352–7.328 | 0.011a |
| Grade | 1.745 | 1.306–2.332 | 0.001a | | | |
| Histological
type | 1.399 | 0.860–1.805 | 0.345 | | | |
| Residual tumor
size | 4.511 | 2.605–7.814 | <0.001a | 4.231 | 2.219–6.942 | 0.006a |
| Recurrence | 3.486 | 1.504–8.082 | 0.004a | | | |
| Metastasis | 1.462 | 0.895–2.389 | 0.129 | | | |
Depletion of CDKN3 expression suppresses
OVCAR3 cell proliferation
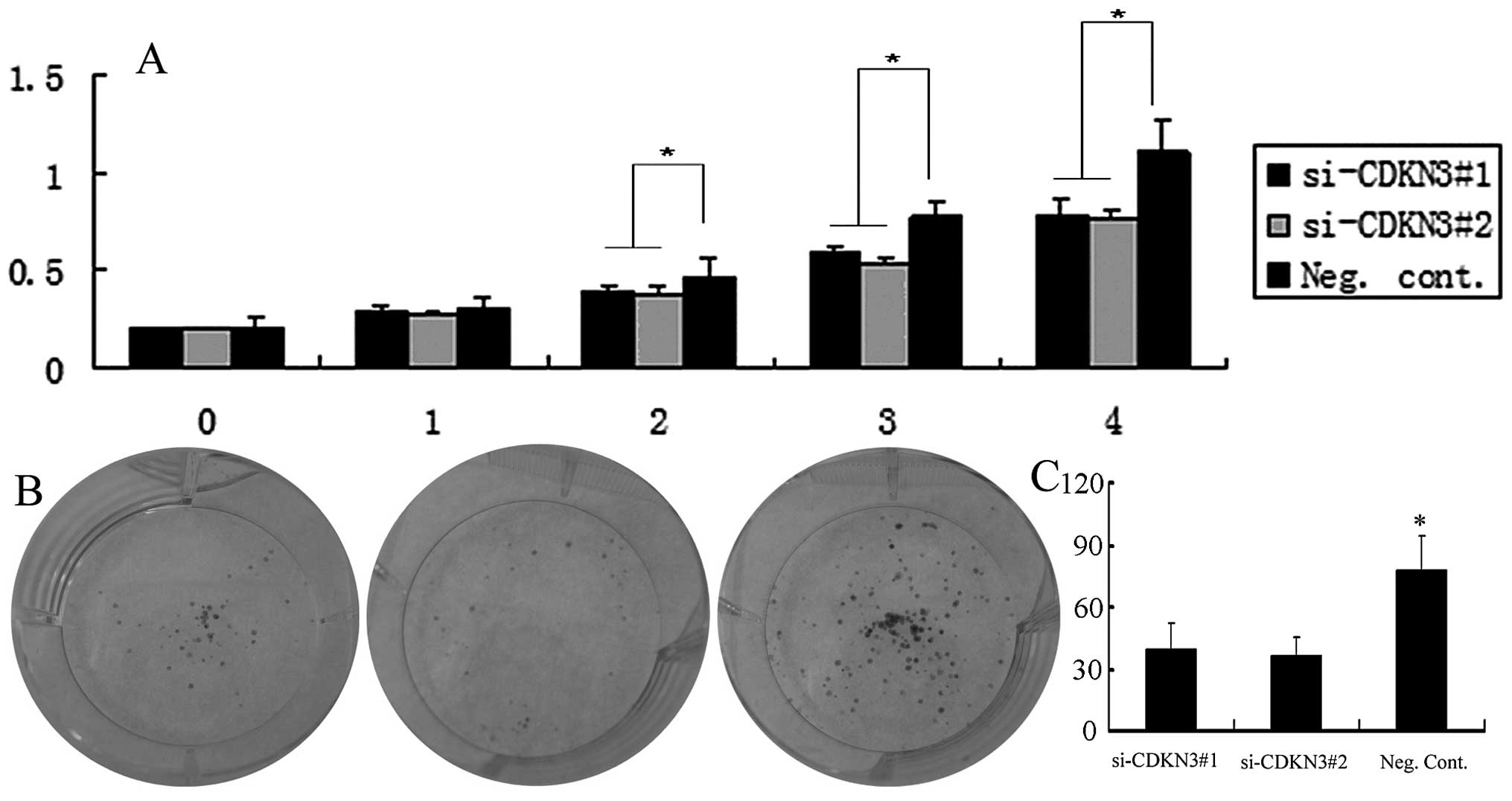
MTT assay showed that downregulation of CDKN3
expression significantly reduced the viability of the OVCAR3 cell
line (Fig. 4A). The depletion of
CDKN3 in the OVCAR3 cells (si-CDKN3#1 and si-CDKN3#2 vs. Neg.
Cont.: 45±11 and 34±9 vs. 81±13; P<0.05) also led to a
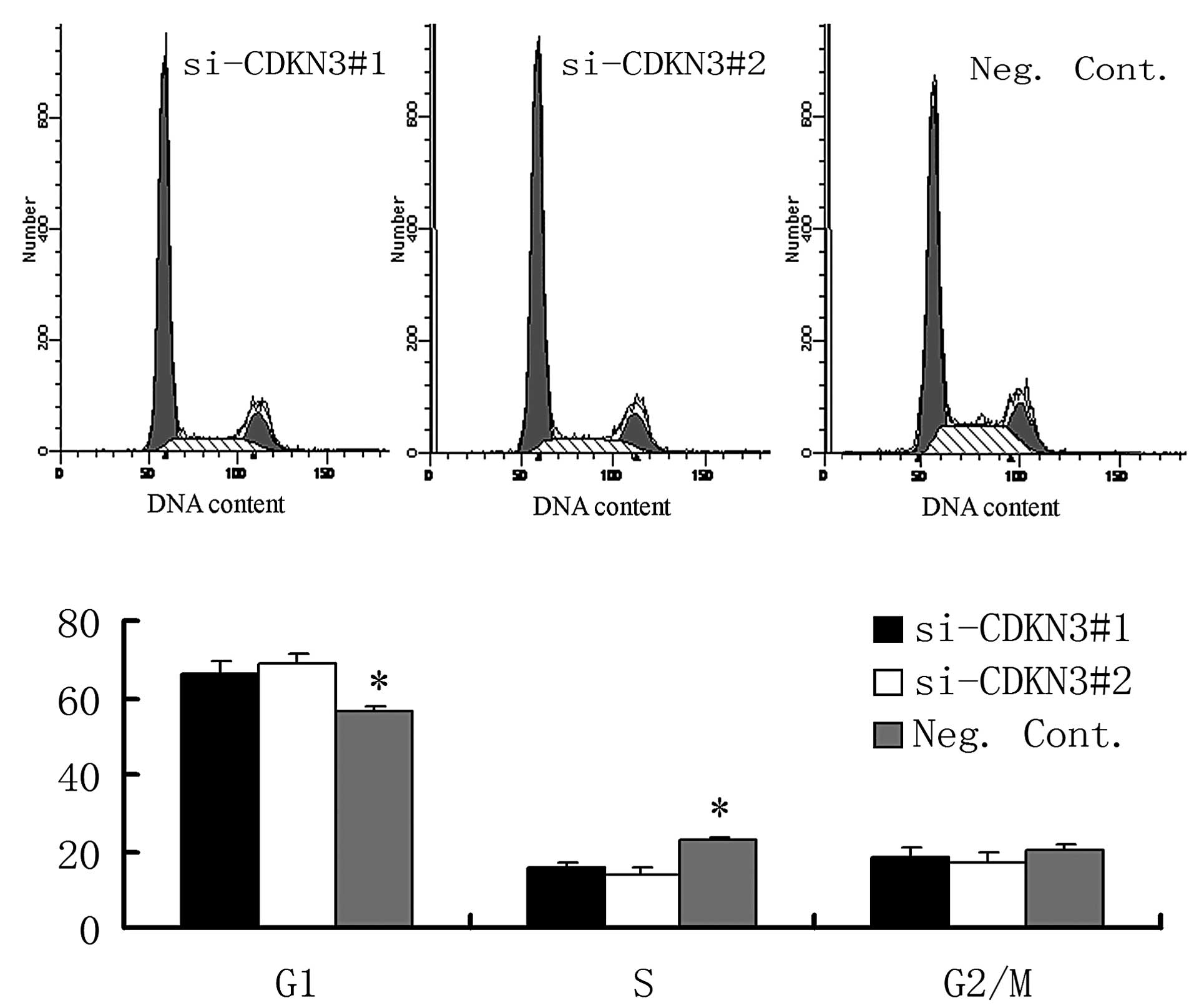
significant reduction in the number and size of foci (Fig. 4B). The DNA content using flow
cytometry demonstrated that CDKN3 siRNA transfection increased the
percentage of cells in the G1 phase and decreased those in the S
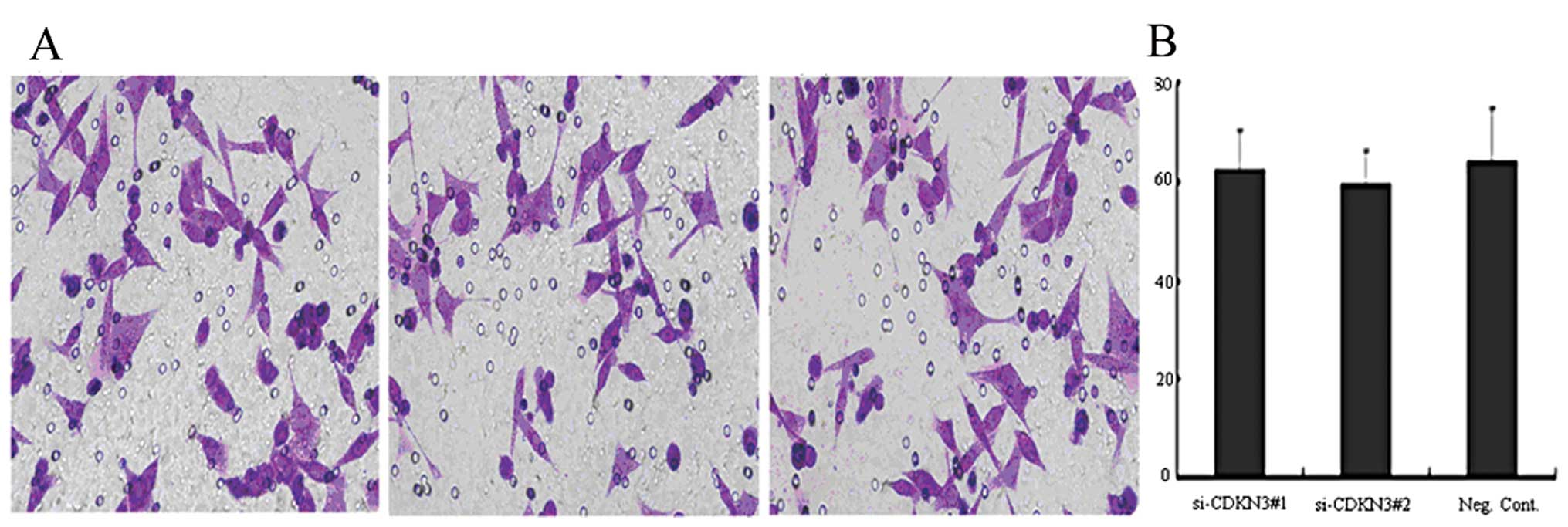
phase in the OVCAR3 cell line (P<0.05, Fig. 5). As shown in Fig. 6, the depletion of CDKN3 in OVCAR3
cells did not have a measurable blocking effect on cell
invasion.
Discussion
The mechanisms of the cell cycle and cancer is one
of the ‘hot’ subjects in the field of oncology research in recent
years. The cell cycle is the basic process of life activity. Under
normal circumstance, cells enter into the physiological state of
proliferation, differentiation, aging and death through normal cell
cycle phases. If abnormal cell cycle regulation occurs, the cell
enters into a cancerous state. Thus, cell cycle regulation is the
core event of cell proliferation regulation, which has a close
relationship with cellular carcinogenesis. Involved in cell cycle
regulation are: cell cycle proteins (cyclins), cell cycle
protein-dependent kinases (cyclin-dependent kinases, CDKs) and cell
cycle protein kinase inhibitors (cyclin-dependent kinase
inhibitors, CKI/CDKN). Cyclins consist of 8 members, respectively
named cyclin A to H. CDK consist of 7 members, respectively named
CDK1–7.
CKI/CDKN, a newly discovered protein, inhibits CDK
kinase activity combined with CDK, cyclin or cyclin-CDK complex. In
the present study, 97 EOC samples were analyzed by χ2
test, indicating that CDKN3 expression had a close relationship
with EOC proliferation and tumor recurrence, and it may play a
significant role in tumor carcinogenesis and EOC progression. Based
on Kaplan-Meier analysis, we discovered that high expression of
CDKN3 indicates the poor prognosis of EOC patients, suggesting that
it can represent a novel prognostic factor for EOC. This was the
first time we evaluated the relationship among CDKN3,
clinicopathological features and prognosis in EOC. Furthermore, we
recognized that CDKN3 may be used as a novel independent prognostic
biomarker to predict the overall survival rate of EOC patients
based on the univariate and multivariate Cox hazards analysis. This
function of CDKN3 corroborated the results of studies concerning
renal cancers and hepatocellular carcinomas (18,19).
In previous research, several predictors similar to CDKN3, such as
p19INK4d (20), CD163 (21), KPNA2 (22) and p21Waf1/Cip1 (23), were recognized as being related to
the prognosis of EOC. To date, however, it is still not definitely
confirmed whether indices of this type could be a substitute for
predicting the prognosis of EOC. Therefore, further research must
be carried out to determine whether CDKN3 together with other
molecules of this category may be valuable for determining the
prognosis of EOC.
To better understand the biological function of
CDKN3, we investigated whether depletion of CDKN3 reduces the
malignant phenotypes in EOC cell lines. CDKN3 was previously found
to be relatively highly expressed in various cancer cell lines
(18,23). Antisense KAP (CDKN3) in HeLa and
LNCaP cells exhibited S-phase reduction and also suppression of the
cell growth rate in vivo and in vitro (24). Accordingly, we reasoned that CDKN3
may play a role in the regulation of cell growth in ovarian cancer
cell lines. We examined the colony formation potential and cell
growth rate in OVCAR3 cells following siRNA treatment. We found
that OVCAR3 knockdown caused a significant reduction in the
proliferation rate and colony formation ability. Therefore, CDKN3
overexpression stimulated malignant cell proliferation. Moreover,
we analyzed the role of CDKN3 on cell cycle progression, similar to
previous studies (16,25,26),
CDKN3 inhibited cell cycle progression in OVCAR3 cells. These data
together demonstrated that CDKN3 had a measurable effect on ovarian
cancer cell proliferation.
Invasion and metastasis are biological
characteristics of malignant tumors, and pose the most problems for
clinical treatments. Recent studies indicate that CDKN3 is
positively correlated with metastasis of neuroendocrine tumors when
compared with their non-metastatic counterparts (27). However, in our study, we did not
find such an association. Transwell cell migration assays showed
that CDKN3 knockdown did not have a measurable effect on OVCAR3
cell invasion. This finding is coincident with the clinical data of
Xing et al (16) in liver
cancer. A possible explanation is possibly that tumor cells which
express CDKN3 are not able to migrate from the primary tumor site,
leading to reduction in metastatic potential, yet CDKN3 expression
could support tumor cell survival and proliferation. In addition,
CDKN3 may have different influence on cancer cells in various
cancer types.
Another issue involves the mechanism of how CDKN3
promotes ovarian cancer progression. A previous study reported that
p21 interacts with cyclin/CDK complexes and PCNA to inhibit their
kinase activities (28), and CDKN3
could promote the proliferation of liver cells through a p53-p21
manner to induce G1/S phase transition (16). Other studies have shown that CDKN3
through its protein product KAP regulates the cell cycle. It
regulates retinoblastoma protein (pRb) activation via a
phosphorylation mechanism that is responsible for G1 to S
transition (29,30). Currently, one hypothesis of this
mechanism is the hypomethylation of its promoter region (9). This hypothesis warrants further
study.
In conclusion, this study identified CDKN3 as an
oncoprotein overexpressed in EOC which is important for the
maintenance of the malignant phenotype and recognized CDKN3 as a
candidate target protein for future cancer therapeutics.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (no. 81171649 to Yi Guo).
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2
|
Dundr P: Ovarian carcinoma: current
diagnostic principles. Cesk Patol. 46:53–61. 2010.(In Czech).
|
|
3
|
Seidman JD, Ronnett BM and Kurman RJ:
Pathology of borderline (low malignant potential) ovarian tumours.
Best Pract Res Clin Obstet Gynaecol. 16:499–512. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Seidman JD and Kurman RJ: Pathology of
ovarian carcinoma. Hematol Oncol Clin North Am. 17:909–925.
vii2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Westfall D, Roma AA and Silva EG:
High-grade serous carcinoma of the ovary. Ann Diagn Pathol.
13:285–290. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vang R, Shih Ie M and Kurman RJ: Ovarian
low-grade and high-grade serous carcinoma: pathogenesis,
clinicopathologic and molecular biologic features, and diagnostic
problems. Adv Anat Pathol. 16:267–282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tarin D: Inappropriate gene expression in
human cancer and its far-reaching biological and clinical
significance. Cancer Metastasis Rev. 31:21–39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Patterson KI, Brummer T, O’Brien PM and
Daly RJ: Dual-specificity phosphatases: critical regulators with
diverse cellular targets. Biochem J. 418:475–489. 2009.PubMed/NCBI
|
|
9
|
Niculescu MD, Yamamuro Y and Zeisel SH:
Choline availability modulates human neuroblastoma cell
proliferation and alters the methylation of the promoter region of
the cyclin-dependent kinase inhibitor 3 gene. J Neurochem.
89:1252–1259. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yeh CT, Lu SC, Chen TC, Peng CY and Liaw
YF: Aberrant transcripts of the cyclin-dependent kinase-associated
protein phosphatase in hepatocellular carcinoma. Cancer Res.
60:4697–4700. 2000.PubMed/NCBI
|
|
11
|
Yeh CT, Lu SC, Chao CH and Chao ML:
Abolishment of the interaction between cyclin-dependent kinase 2
and Cdk-associated protein phosphatase by a truncated KAP mutant.
Biochem Biophys Res Commun. 305:311–314. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blum R, Nakdimon I, Goldberg L, et al:
E2F1 identified by promoter and biochemical analysis as a central
target of glioblastoma cell-cycle arrest in response to Ras
inhibition. Int J Cancer. 119:527–538. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu L, Timmers C, Maiti B, et al: The
E2F1-3 transcription factors are essential for cellular
proliferation. Nature. 414:457–462. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Demetrick DJ, Matsumoto S, Hannon GJ, et
al: Chromosomal mapping of the genes for the human cell cycle
proteins cyclin C (CCNC), cyclin E (CCNE), p21 (CDKN1) and KAP
(CDKN3). Cytogenet Cell Genet. 69:190–192. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
MacDermed DM, Khodarev NN, Pitroda SP, et
al: MUC1-associated proliferation signature predicts outcomes in
lung adenocarcinoma patients. BMC Med Genomics. 3:162010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xing C, Xie H, Zhou L, et al:
Cyclin-dependent kinase inhibitor 3 is overexpressed in
hepatocellular carcinoma and promotes tumor cell proliferation.
Biochem Biophys Res Commun. 420:29–35. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taylor KJ, Sims AH, Liang L, et al:
Dynamic changes in gene expression in vivo predict prognosis of
tamoxifen-treated patients with breast cancer. Breast Cancer Res.
12:R392010. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lai MW, Chen TC, Pang ST and Yeh CT:
Overexpression of cyclin-dependent kinase-associated protein
phosphatase enhances cell proliferation in renal cancer cells. Urol
Oncol. 30:871–878. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang L, Sun L, Huang J and Jiang M:
Cyclin-dependent kinase inhibitor 3 (CDKN3) novel cell cycle
computational network between human non-malignancy associated
hepatitis/cirrhosis and hepatocellular carcinoma (HCC)
transformation. Cell Prolif. 44:291–299. 2011. View Article : Google Scholar
|
|
20
|
Felisiak-Golabek A, Dansonka-Mieszkowska
A, Rzepecka I, et al: p19 mRNA and protein expression as new
prognostic factor in ovarian cancer patients. Cancer Biol Ther.
14:973–981. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
No JH, Moon JM, Kim K and Kim YB:
Prognostic significance of serum soluble CD163 level in patients
with epithelial ovarian cancer. Gynecol Obstet Invest. 75:263–267.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang L, Wang HY, Li JD, et al: KPNA2
promotes cell proliferation and tumorigenicity in epithelial
ovarian carcinoma through upregulation of c-Myc and downregulation
of FOXO3a. Cell Death Dis. 4:e7452013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bali A, O’Brien PM, Edwards LS, Sutherland
RL, Hacker NF and Henshall SM: Cyclin D1, p53, and p21Waf1/Cip1
expression is predictive of poor clinical outcome in serous
epithelial ovarian cancer. Clin Cancer Res. 10:5168–5177. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee SW, Reimer CL, Fang L, Iruela-Arispe
ML and Aaronson SA: Overexpression of kinase-associated phosphatase
(KAP) in breast and prostate cancer and inhibition of the
transformed phenotype by antisense KAP expression. Mol Cell Biol.
20:1723–1732. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chinami M, Yano Y, Yang X, et al: Binding
of HTm4 to cyclin-dependent kinase (Cdk)-associated phosphatase
(KAP). Cdk2cyclin A complex enhances the phosphatase activity of
KAP, dissociates cyclin A, and facilitates KAP dephosphorylation of
Cdk2. J Biol Chem. 280:17235–17242. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu Y, Jiang X, Schoch BS, Carroll RS,
Black PM and Johnson MD: Aberrant splicing of cyclin-dependent
kinase-associated protein phosphatase KAP increases proliferation
and migration in glioblastoma. Cancer Res. 67:130–138. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee J, Sung CO, Lee EJ, et al: Metastasis
of neuroendocrine tumors are characterized by increased cell
proliferation and reduced expression of the ATM gene. PLoS One.
7:e344562012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen J, Jackson PK, Kirschner MW and Dutta
A: Separate domains of p21 involved in the inhibition of Cdk kinase
and PCNA. Nature. 374:386–388. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barford D: The mechanism of protein kinase
regulation by protein phosphatases. Biochem Soc Trans. 29:385–391.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Johnson LN, De Moliner E, Brown NR, et al:
Structural studies with inhibitors of the cell cycle regulatory
kinase cyclin-dependent protein kinase 2. Pharmacol Ther.
93:113–124. 2002. View Article : Google Scholar : PubMed/NCBI
|