Introduction
Breast cancer is a major health issue worldwide. It
was estimated that approximately 234,580 individuals were diagnosed
with breast cancer, and more than 40,030 died from this disease in
the US in 2013 (1). In China,
breast cancer is currently the most common malignancy and the sixth
leading cause of cancer-related mortality in women (2). Epidermal growth factor receptor (EGFR)
and ErbB2 (HER2) are both receptor tyrosine kinases (RTKs), which
play important roles in the regulation of cell proliferation,
motility and apoptosis. Accumulating evidence has confirmed that
they are major and attractive targets for cancer therapy. Previous
studies have reported that EGFR and ErbB2 are overexpressed in
~40–50% of breast cancer cases (3),
and increased expression of EGFR or ErbB2 is associated with breast
cancer growth and the poor survival and prognosis of breast cancer
patients (3). To date, a number of
agents have been developed to target EGFR or ErbB2 that result in
tumor cell growth arrest and apoptosis (4–6), most
of which have focused on the inhibition of ligand binding or
tyrosine phosphorylation and have achieved success. However, EGFR
and ErbB2 mutations in cancers make the development of novel agents
targeting the EGFR or ErbB2 signaling pathway urgent for cancer
therapy (7–9).
Growth factor receptor bound protein 2 (Grb2) is a
key adaptor binding to EGFR and ErbB2, which in turn binds to SOS
and activates transduction of cellular responses downstream of Ras
RTKs through the Ras-ERK signaling cascade (10). It consists of a central Src homology
2 (SH2) domain and two Src homology 3 (SH3) domains. The N-terminal
SH3 domain recognizes proline-rich sequences adopting a type II
polyproline (PPII) helix, while the C-terminal SH3 domain
recognizes sequences conforming P-X-I/L/V/-D/N-R-X-X-K-P motif
(11). The SH2 domains recognize
tyrosine phosphorylated sequences expediting
phosphorylation-dependent interactions of proteins resulting in
signal propagation (12,13). Obvious overexpression of Grb2 was
observed in breast cancer cell lines and tissue samples (14,15).
Grb2 links RTKs to the Ras signaling pathway, which controls major
signaling routes of cell proliferation, differentiation and
survival. It has been reported that downregulation of Grb2
suppresses the differentiation of mouse embryonic stem cells
(16) and silencing Grb2 expression
reduces cell growth in vitro (17). It has been suggested that Grb2 is a
potential target for the development of therapeutic agents.
SH2 domains are found in many intercellular
signal-transduction proteins. SH2 is involved in the development of
a wide range of human diseases including cancers, diabetes and
immunodeficiencies (18). SH2 is a
relatively small protein module of ~100 amino acids, which
facilitates activation of intracellular signaling by interacting
with phosphorylated tyrosine containing peptide sequences (19,20).
The binding selectivity of SH2 domains is critical for the fidelity
and specificity of cellular signal transduction pathways (21–24).
Considerable effort has been devoted to study the pathogenic
function of SH2 domains. The fusion Grb2-SH2 protein was found to
inhibit EGFR-Grb2 signaling in squamous cells and may have the
potential to treat EGFR-activated cancer (7). The prokaryotic recombined fusion
Grb2-SH2 protein was confirmed to inhibit the proliferation of
breast cancer cells in our previous study (25).
To investigate the function of the Grb2-SH2 domain,
we generated two fusion proteins. One contained one SH2 domain of
Grb2, together with a signal peptide sequence, the PTD region, and
FLAG-tag sequence. The other one consisted of a mutant SH2 domain,
signal peptide sequence, PTD region and a FLAG-tag sequence. The
PTD sequence was reported in a previous study (25). In the present study, we constructed
eukaryotic expression vectors containing the recombinant gene
sequences and they were expressed in breast cancer cell lines, for
which the biological behavior was also investigated.
Materials and methods
Cell culture
E. coli JM109 (Invitrogen, Carlsbad, CA, USA)
were stored at −70°C and cultured in Luria-Bertani broth medium
(LB). The pUC-57 and pIRES2-EGFP were commercial products (BD
Biosciences, Clontech, CA, USA) and stored at −20°C. The breast
cancer cell lines HER2-positive MCF-7 and HER2-negative MDA-MB-231
were preserved in the Department of Molecular Biology of the Fourth
Military Medical University. The cells were cultured in Dulbecco’s
modified Eagle’s medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA)
supplemented with 10% fetal bovine serum (FBS), 50 units/ml
penicillin and 50 μg/ml streptomycin (FBS; Gibco Invitrogen Corp.,
Carlsbad, CA, USA). Cell cultures were maintained in a humidified
5% CO2 atmosphere at 37°C.
Expression vector construction
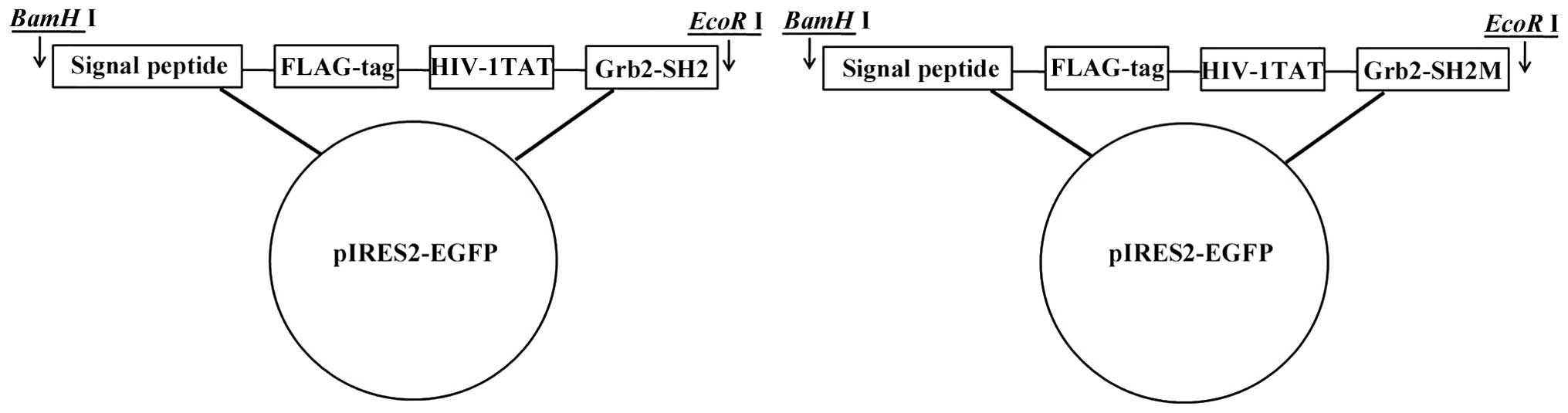
The fusion FPTD-Grb2-SH2 DNA sequence was designed
based on the native Grb2-SH2 coding DNA sequence as previously
described (25). It contained one
signal peptide DNA sequence (5′-ATGAAGCA
CCTTTGGTTCTTCCTTCTTCTTGTAGCAGCACCAAGG TGGGTACTTTCG-3′); one
FLAG-tag DNA sequence (5′-GA CTACAAGGACGATGATGACAAG-3′); one
PTD-transduction domain DNA sequence (5′-TATGGTAGGAAG
AAACGTCGACAGCGTCGTCGG-3′) and one Grb2-SH2 domain DNA sequence,
which was synthesized by Sangon Biotech Co. The artificial DNA
fragment was digested with BamHI and EcoRI into the
pUC-57 plasmid and subsequently subcloned into digested pIRES2-EGFP
to construct the expression vector, which was named
pIRES-FPTD-Grb2-SH2. Meanwhile, a mutant DNA fragment was designed
as a contrast. It was amplified by polymerase chain reaction using
a forward primer (5′-TGGTTTTTTGGCAAAATCCCCT CTTTGAATGAGC-3′) and a
reverse primer (5′-ATCCACC AGCTCATTCAAAGAGGGGATTTTGCC-3′). This DNA
fragment was subsequently inserted into digested pUC-57 and
pIRES2-EGFP. The recombinant expression vector was named
pIRES-FPTD-Grb2-SH2M. All insertions were confirmed by DNA
sequencing and BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/). The recombinant
plasmids were kept in JM109 and stored at −70°C.
Transfection
Cells were transfected with the recombinant plasmids
pIRES-FPTD-Grb2-SH2 and pIRES-FPTD-Grb2-SH2M. The breast cancer
cells were seeded at 2×105/well in 6-well plates for ~24
h until 80% confluency was achieved. Cell transfections were
performed using X-tremeGENE HP DNA transfection reagent (Roche,
Germany) according to the manufacturer’s instructions. Plasmid DNA
(2 μg) was mixed with the transfection reagent, and then the
complex was added to the cell cultured in fresh medium. Transfected
cells were incubated for 36 and 48 h. The untransfected cells
cultured in the same condition were set as controls. The cells
incubated with the transfection reagent and plasmid pIRES2-EGFP
were also included in the experiments. At the end of each
transfection period, cells were collected for the following
measurement [immunofluorescence assay, western blot analysis,
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT)
assay, cell migration assay and flow cytometric detection].
Western blot analysis
Cell lysates containing 30 μg protein were subjected
to SDS-PAGE on a 15% gel and transferred onto a PVDF membrane
(Bio-Rad). They were probed by a primary antibody against FLAG-tag
(Qiagen; 1:2,000) with 2.5% milk in TBS. HRP-labeled secondary
antibodies were detected using ECL detection kit (GE Healthcare),
and the target bands were visualized using the ChemiDoc™ MP
detection system (Bio-Rad Laboratories, Inc.).
Immunofluorescence assay
Cells on coverslips were fixed in 4%
paraformaldehyde and permeabilized in PBS with 0.1% Triton X-100.
After blocking the non-specific binding in 2% normal sheep serum in
PBS, the coverslips were incubated with a rabbit anti-FLAG-tag
polyclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA)
at a 1:400 dilution overnight at 4°C. The cells were then washed
with PBS and incubated with a fluorescence-labeled secondary
antibody (Dako, Glostrup, Denmark) for 3 h. The specific
immunoreactivity was observed under a reflected light fluorescence
microscope (BH2-RFC; Olympus, Tokyo, Japan).
Cell proliferation assay
The MTT assay was used to evaluate cell
proliferation following transfection. Cells were harvested and
plated in 96-well cell plates (2×104 cells/well) in
sextuplicate. Aliquots (20 μl) of 5 mg/ml MTT (Sigma) in PBS were
added to each well and incubated for 4 h at 37°C; 150 μl of
Me2SO was subsequently added. The absorbance (A) values
were measured on a Sunrise microplate reader (Tecan, Groedig,
Austria) at 490 nm.
Cell migration assay
The cells (2×105) in 200 μl medium free
of serum were placed in the upper chamber of Transwell cell culture
inserts (8-μm pore size; Corning Life Sciences). The lower chamber
contained 500 μl complete medium (DMEM with 10% FBS). Migration was
carried out for 24 h at 37°C in a 5% CO2 atmosphere.
Cells that had migrated into the lower chamber were quantified by
counting in a Neubauer counting chamber using an inverted cell
culture microscope (Olympus). The migration inhibition rate was
calculated using the formula: Migration inhibition rate = [(1 -
transfected cells/untransfected cells) × 100%].
Analysis of cell apoptosis
Cells were stained with Annexin V-FITC and PI, and
flow cytometric analysis was performed using the Elite ESP flow
cytometer (FACSCalibur; Becton-Dickinson Immunocytometry Systems,
San Jose, CA, USA) for apoptosis analysis according to the
manufacturer’s guidelines. The data were analyzed using the
CellQuest Pro Software (BD Biosciences, San Jose, CA, USA).
Statistical analysis
Statistical analysis was performed using the SPSS
19.0 software package for Windows. P-values <0.05 were
considered to indicate statistically significant results.
Results
Sequence synthesis, expression and
identification
The successful construction of the expression
vectors pIRES-FPTD-Grb2-SH2 and pIRES-FPTD-Grb2-SH2M was confirmed
by restriction mapping (Fig. 1).
The result of the automatic sequencing was consistent with the
expected FPTD-Grb2-SH2 DNA fragment (388 bp) and its mutant (208
bp). The expression vectors were transfected into breast cancer
cells, respectively, with untransfected cells, transfection reagent
and empty vector used as controls. The average transfection
efficiency was ~70% as estimated by fluorescence-positive cells
under a florescence microscope after 36 h. The transfected cells
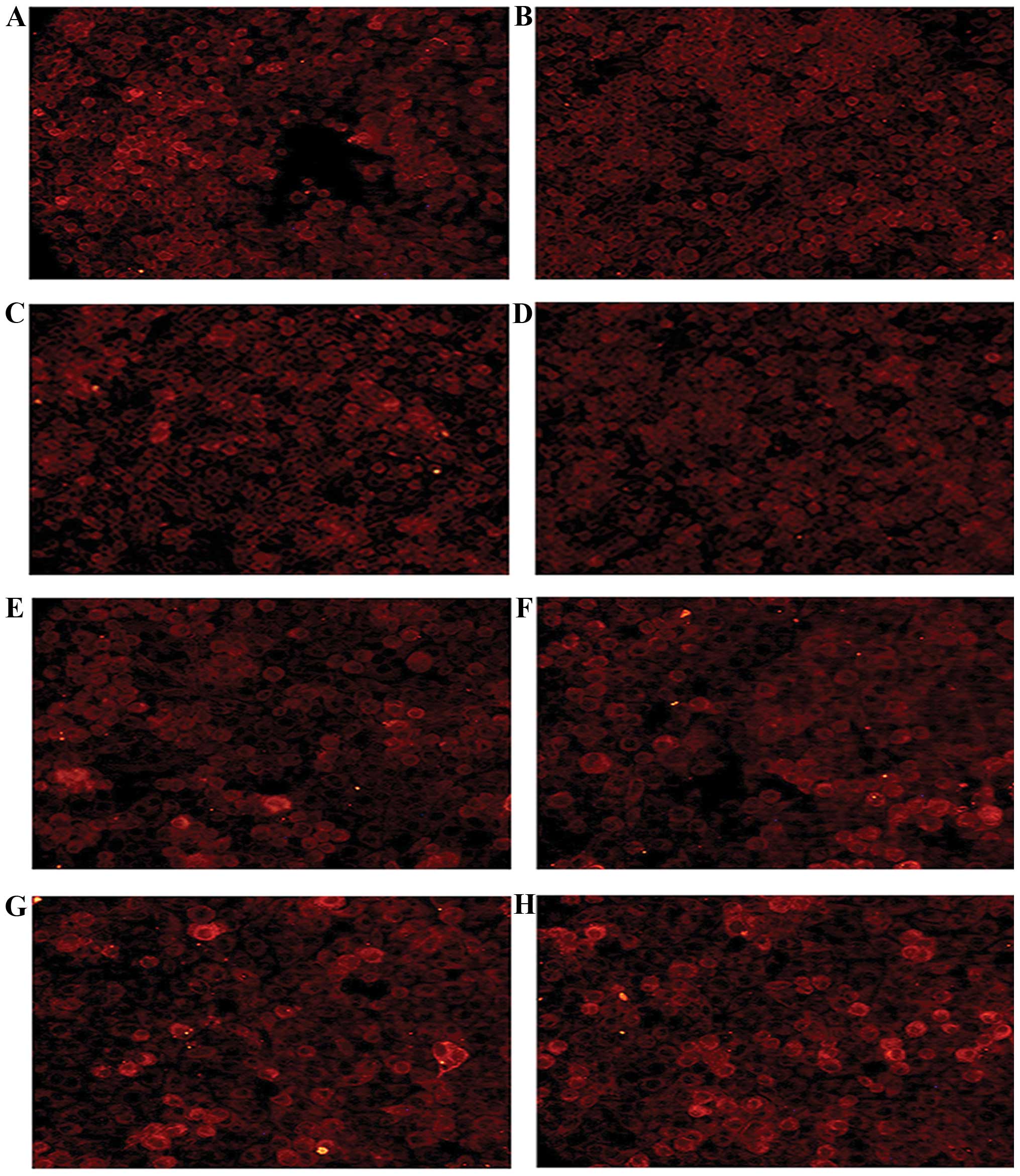
were incubated for 36 and 48 h, respectively. Immunofluorescence
assay with the anti-FLAG-tag antibody revealed that FPTD-Grb2-SH2
and FPTD-Grb2-SH2M proteins were expressed in the MCF-7 and
MDA-MB-231 cells. As shown in Fig.
2, both the recombinants were mainly located in the cytoplasm,
indicating that signal peptide and HIV-1 TAT48-60 helped the target
peptides to pass through the nuclear membranes in living cells.
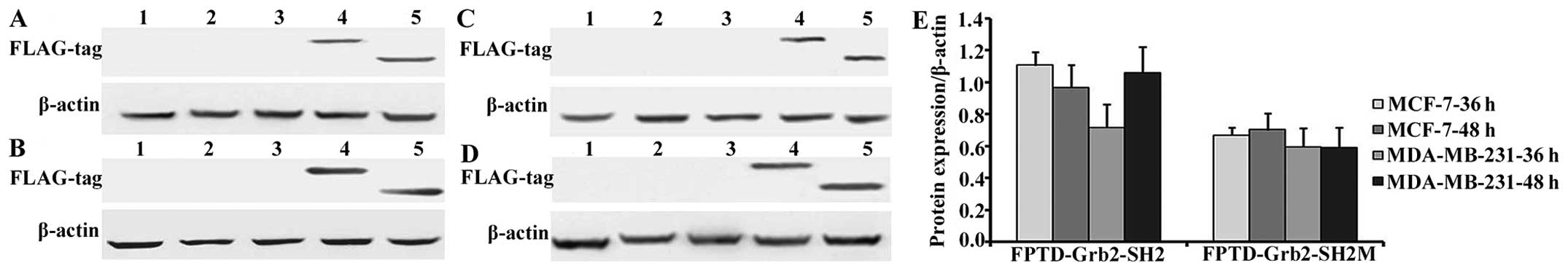
Protein expression levels were further determined by
western blotting (Fig. 3). A
statistically significant stronger expression of FPTD-Grb2-SH2
compared to FPTD-Grb2-SH2M was observed in both breast cancer cell
lines (P<0.05). Expression of FPTD-Grb2-SH2 protein increased
when transfected MDA-MB-231 cells were incubated for 48 h, while it
decreased in MCF-7 cells as shown in Fig. 3E.
Growth inhibition of fusion FPTD-Grb2-SH2
and fusion FPTD-Grb2-SH2M proteins in the breast cancer MCF-7 and
MDA-MB-231 cell lines
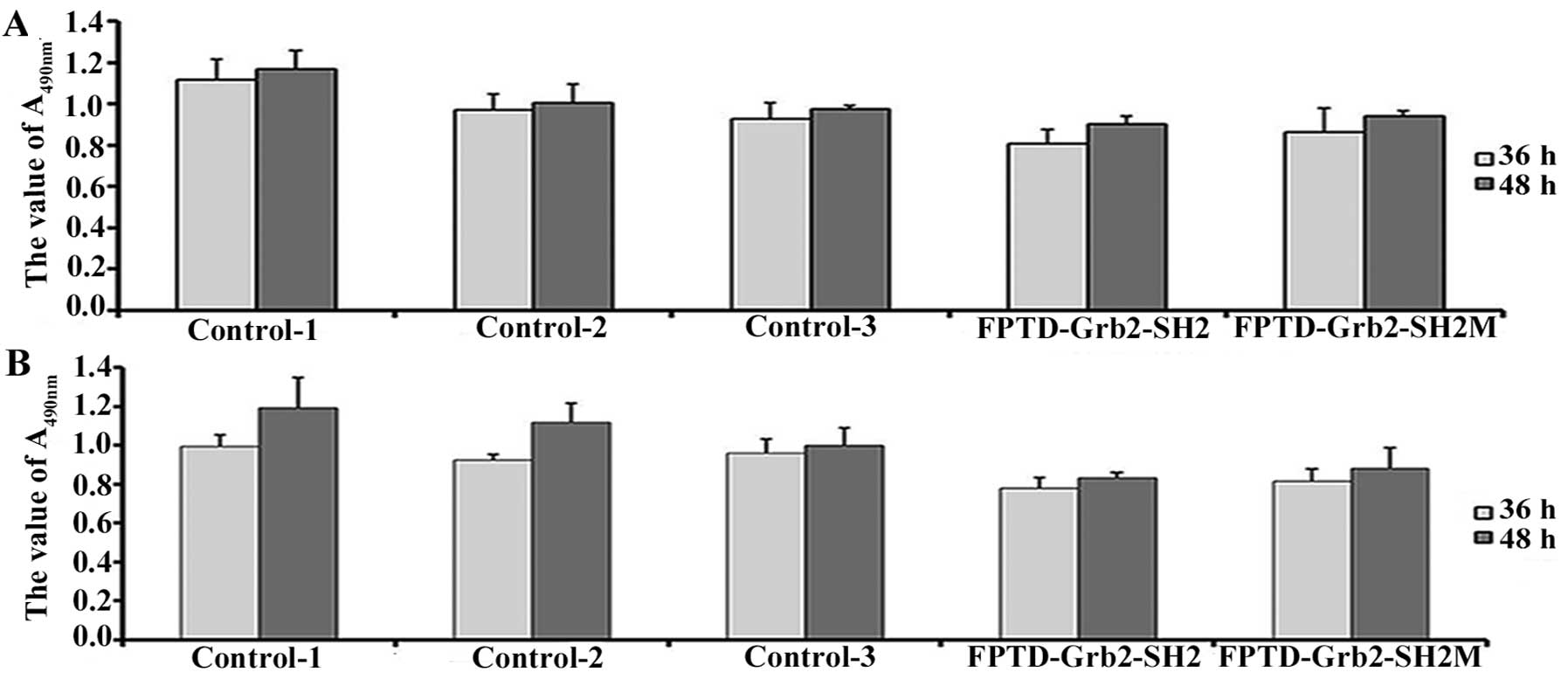
The recombinant plasmids pIRES-FPTD-Grb2-SH2 and
pIRES-FPTD-Grb2-SH2M were transfected into breast cancer cell lines
MCF-7 and MDA-MB-231 to investigate whether the new genes could
affect the proliferation of breast cancer cells in vivo. The
growth inhibitory effects of FPTD-Grb2-SH2 and its mutant were
observed in both breast cancer cells (Fig. 4). FPTD-Grb2-SH2 exhibited a
significantly stronger inhibitory effect on the breast cancer
cells. After incubation for 48 h, a decreased rate of cell
proliferation inhibition was detected in the transfected cells,
when compared to the transfected cells incubated for 36 h
(P<0.05).
Inhibition of migratory ability by fusion
FPTD-Grb2-SH2 and fusion FPTD-Grb2-SH2M proteins in the MCF-7 and
MDA-MB-231 cells
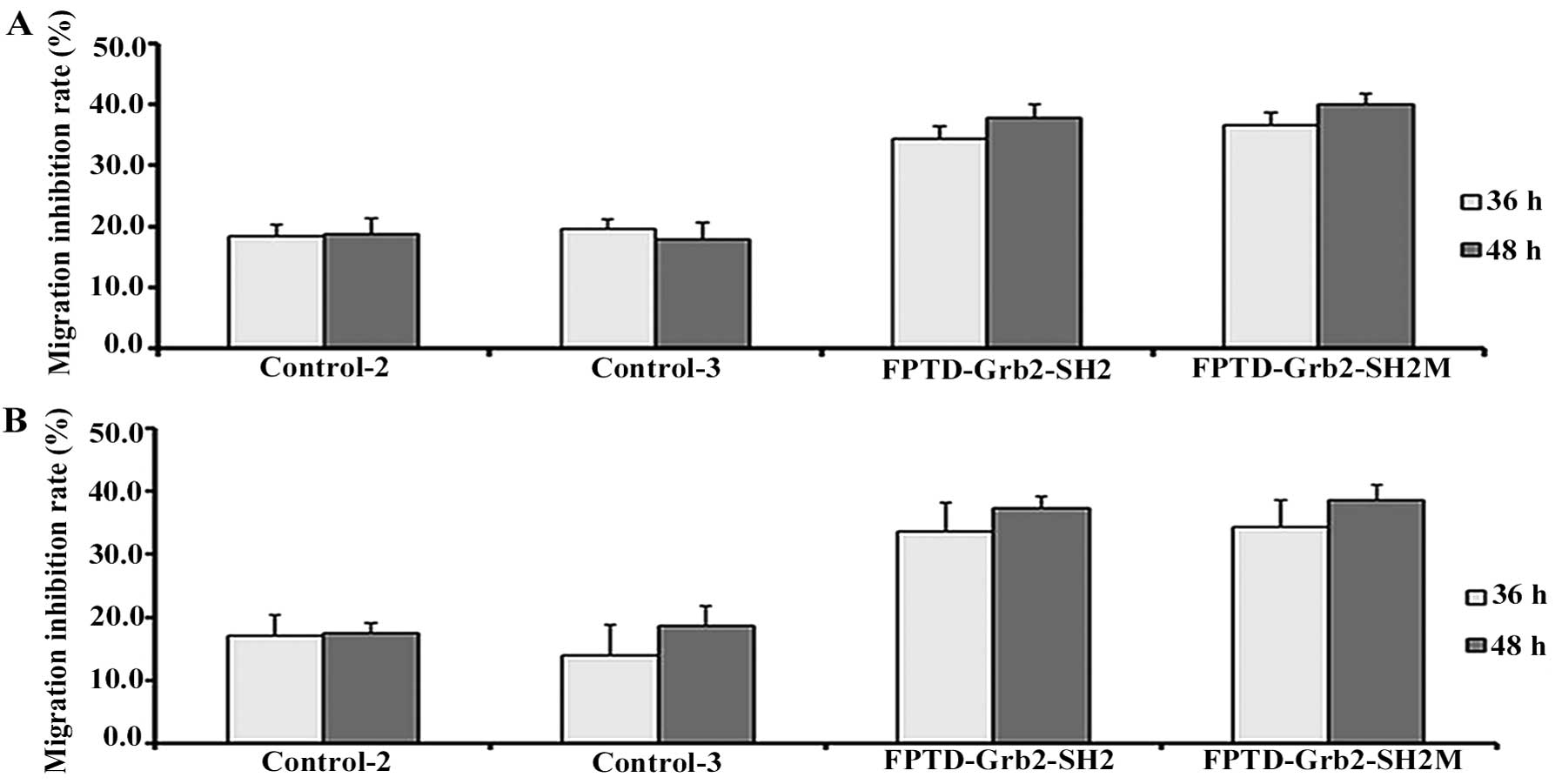
As shown in Fig. 5,
the migratory ability of the MCF-7 and MDA-MB-231 cells was reduced
by recombinant FPTD-Grb2-SH2 and FPTD-Grb2-SH2M when compared to
the empty vector control. Fig. 5A and
B shows that the percentage of viable cells transfected with
the fusion proteins was reduced by at least ~15% as compared with
the cells transfected with the empty vector. The migration
inhibition rate was enhanced when the transfected cells were
incubated for 48 h. No significant difference was noted in the
cells transfected with the empty vector control.
Recombinant FPTD-Grb2-SH2 and
FPTD-Grb2-SH2M proteins induce the apotosis of MCF-7 and MDA-MB-231
cells
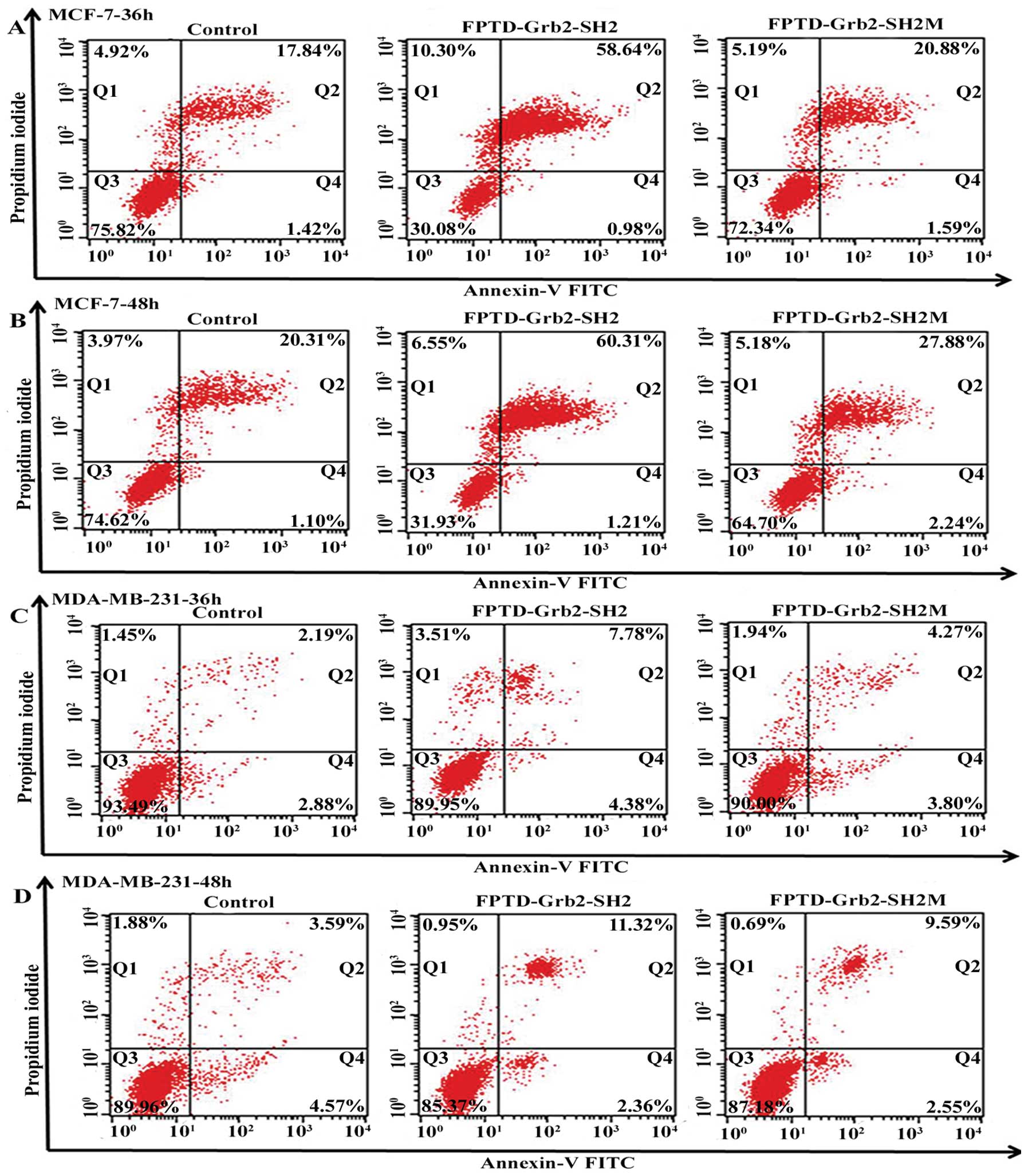
MCF-7 and MDA-MB-231 cells were stained with Annexin
V-FITC and PI, and then subjected to flow cytometry. Fig. 6 reveals that both FPTD-Grb2-SH2 and
its mutant induced apoptosis in the MCF-7 and MDA-MB-231 cells,
when compared to the empty vector controls. However, marked
induction of apoptosis of the MCF-7 cells was induced by
FPTD-Grb2-SH2. Apoptosis proceeded in 59.62% of the infected MCF-7
cells at 36 h and 61.52% of infected MCF-7 cells at 48 h, which was
much higher than the percentage in the MDA-MB-231 cells (12.06 and
13.68%, respectively) (Fig. 6). In
addition, the ability to induce apoptosis exerted by FPTD-Grb2-SH2
in both breast cell lines was much more obvious than that by
FPTD-Grb2-SH2M.
Discussion
Grb2 is a dynamic downstream intermediary in the
HER2 signaling pathways, which has been implicated directly in the
pathogenesis of several specific human malignancies. In addition,
overexpression of Grb2 is found in breast and bladder cancer cells
(26,27), which suggests that Grb2 can be a
valid therapeutic target for cancer treatment. The Grb2-SH2 domain
is a relatively conserved 100 amino acid modular unit, and
disruption of Grb2-SH2 domain interactions can significantly
inhibit the Grb2-mediated signaling pathway (28). Therefore, the Grb2-SH2 domain has
been recognized as an ideal pharmaceutical target.
In our previous study, we expressed and purified a
novel prokaryotic fusion SH2 protein and its mutant, which were
successfully transduced into living breast cancer cells resulting
in the inhibition of growth of these cells (25). In the present study, the eukaryotic
fusion proteins containing the SH2 domain and the mutant with a
signal peptide sequence, PTD domain sequence and FLAG-tag sequence
were directly expressed in breast cancer cells, respectively. (The
proteins were named FPTD-Grb2-SH2 and FPTD-Grb2-SH2M,
respectively). The fusion proteins were small proteins with a
molecular weight <20 kDa. Automatic sequencing examined the
target sequences to prevent mis-construction and expression. Unlike
the prokaryotic mutant protein, FPTD-Grb2-SH2M also exerted growth
inhibition in the breast cancer cell lines MCF-7 and MDA-MB-231,
suggesting that the absence of protein modification in the
eukaryotic expression system would result in dysfunction of the
target protein.
The recombinant proteins FPTD-Grb2-SH2 and
FPTD-Grb2-SH2M were expressed in both HER2-negative MDA-MB-231 and
HER2-positive MCF-7 cells. Data showed that both these two novel
fusion proteins inhibited the growth and migration of the breast
cancer cells. The effects of FPTD-Grb2-SH2 and FPTD-Grb2-SH2M on
cell migration, were similar while a slight stronger proliferation
inhibition by FPTD-Grb2-SH2 was observed. This implies that the
reserved sequence FPTD-Grb2-SH2M may still maintain the basic
function of the SH2 domain. Despite that both SH2 and SH3 domains
recognize and specifically bind to their ligands, SH2 domains are
major known binding modules for tyrosine-phosphorylated proteins
and are essential for breast cancer cell survival (29). Furthermore, compared with
FPTD-Grb2-SH2M, FPTD-Grb2-SH2 showed enhanced ability to induce
apoptosis in MCF-7 cells. The discrepancy may be explained by the
possibility that the pathway structure affected by administration
of the proteins resulted in abnormal cell viability. However, we
did not estimate the downstream molecules or the pathways of the
Grb2-SH2 protein domain involved in the intricate intracellular
networks (30), and these aspects
need elucidation in further studies.
It has been reported that the function of Grb2 is
closely associated with HER2 and the signaling pathway induced by
the HER2 receptor in human cancer cells (31,32).
In the cell apoptosis analysis, unconspicuous sensitivity to
FPTD-Grb2-SH2 and FPTD-Grb2-SH2M was shown in the HER2-negative
MDA-MB-231 cells, compared with the HER2-positive MCF-7 cells. This
indicates that the SH2 domain may function with more specificity to
the HER2 signal pathway during cell apoptosis. This is opposite to
previous results (25), which
confirmed that the eukaryotic expression system may be helpful for
the function of protein in the present study.
In conclusion, the recombinant proteins
FPTD-Grb2-SH2 and FPTD-Grb2-SH2M were successfully expressed in the
breast cancer cell lines regardless of HER2 phenotype. The SH2
domain may maintain its biological activity alone. Moreover, both
the wild-type and the mutant one exhibited the ability to inhibit
growth and migration and induce apoptosis of breast cancer cells.
This indicated that FPTD-Grb2-SH2 and FPTD-Grb2-SH2M exhibited
significant toxicity to breast cancer cells. The proteins expressed
in the eukaryocyte may be used for anticancer drug development.
Acknowledgements
This study was supported by grant no. 30901457 from
the National Natural Science Foundation of China.
References
|
1
|
Theriault RL, Carlson RW, Allred C, et al:
Breast cancer, version 3.2013: featured updates to the NCCN
guidelines. J Natl Compr Canc Netw. 11:753–761. 2013.PubMed/NCBI
|
|
2
|
Zheng S, Bai JQ, Li J, et al: The
pathologic characteristics of breast cancer in China and its shift
during 1999–2008: a national-wide multicenter cross-sectional image
over 10 years. Int J Cancer. 131:2622–2631. 2012.PubMed/NCBI
|
|
3
|
Tari AM, Hung MC, Li K and Lopez-Berestein
G: Growth inhibition of breast cancer cells by Grb2 downregulation
is correlated with inactivation of mitogen-activated protein kinase
in EGFR, but not in ErbB2, cells. Oncogene. 18:1325–1332. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Roskoski R Jr: The ErbB/HER receptor
protein-tyrosine kinases and cancer. Biochem Biophys Res Commun.
319:1–11. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Normanno N, De Luca A, Bianco C, et al:
Epidermal growth factor receptor (EGFR) signaling in cancer. Gene.
366:2–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang X, Diaz MR and Yee D: Fulvestrant
regulates epidermal growth factor (EGF) family ligands to activate
EGF receptor (EGFR) signaling in breast cancer cells. Breast Cancer
Res Treat. 139:351–360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saito Y, Furukawa T, Arano Y, Fujibayashi
Y and Saga T: Fusion protein based on Grb2-SH2 domain for cancer
therapy. Biochem Biophys Res Commun. 399:262–267. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rexer BN, Ghosh R, Narasanna A, et al:
Human breast cancer cells harboring a gatekeeper T798M mutation in
HER2 overexpress EGFR ligands and are sensitive to dual inhibition
of EGFR and HER2. Clin Cancer Res. 19:5390–5401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Piechocki MP, Yoo GH, Dibbley SK and
Lonardo F: Breast cancer expressing the activated HER2/neu is
sensitive to gefitinib in vitro and in vivo and acquires resistance
through a novel point mutation in the HER2/neu. Cancer Res.
67:6825–6843. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bianco R, Melisi D, Ciardiello F and
Tortora G: Key cancer cell signal transduction pathways as
therapeutic targets. Eur J Cancer. 42:290–294. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Berry DM, Nash P, Liu SK, Pawson T and
McGlade CJ: A high-affinity Arg-X-X-Lys SH3 binding motif confers
specificity for the interaction between Gads and SLP-76 in T cell
signaling. Curr Biol. 12:1336–1341. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Groveman BR, Xue S, Marin V, et al: Roles
of the SH2 and SH3 domains in the regulation of neuronal Src kinase
functions. FEBS J. 278:643–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mayer BJ and Gupta R: Functions of SH2 and
SH3 domains. Curr Top Microbiol Immunol. 228:1–22. 1998.PubMed/NCBI
|
|
14
|
Daly RJ, Binder MD and Sutherland RL:
Overexpression of the Grb2 gene in human breast cancer cell lines.
Oncogene. 9:2723–2727. 1994.PubMed/NCBI
|
|
15
|
Verbeek BS, Adriaansen-Slot SS, Rijksen G
and Vroom TM: Grb2 overexpression in nuclei and cytoplasm of human
breast cells: a histochemical and biochemical study of normal and
neoplastic mammary tissue specimens. J Pathol. 183:195–203. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Liu Q, Jia W, et al: MicroRNA-200a
regulates Grb2 and suppresses differentiation of mouse embryonic
stem cells into endoderm and mesoderm. PLoS One. 8:e689902013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Di Fulvio M, Henkels KM and
Gomez-Cambronero J: Short-hairpin RNA-mediated stable silencing of
Grb2 impairs cell growth and DNA synthesis. Biochem Biophys Res
Commun. 357:737–742. 2007.PubMed/NCBI
|
|
18
|
Liu BA, Jablonowski K, Shah EE, Engelmann
BW, Jones RB and Nash PD: SH2 domains recognize contextual peptide
sequence information to determine selectivity. Mol Cell Proteomics.
9:2391–2404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Songyang Z, Shoelson SE, Chaudhuri M, et
al: SH2 domains recognize specific phosphopeptide sequences. Cell.
72:767–778. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Broadbridge RJ and Sharma RP: The Src
homology-2 domains (SH2 domains) of the protein tyrosine kinase
p56lck: structure, mechanism and drug design. Curr Drug Targets.
1:365–386. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Machida K and Mayer BJ: The SH2 domain:
versatile signaling module and pharmaceutical target. Biochim
Biophys Acta. 1747:1–25. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pawson T: Protein modules and signalling
networks. Nature. 373:573–580. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mayer BJ, Jackson PK and Baltimore D: The
noncatalytic src homology region 2 segment of abl
tyrosine kinase binds to tyrosine-phosphorylated cellular proteins
with high affinity. Proc Natl Acad Sci USA. 88:627–631.
1991.PubMed/NCBI
|
|
24
|
Mayer BJ, Jackson PK, Van Etten RA and
Baltimore D: Point mutations in the abl SH2 domain coordinately
impair phosphotyrosine binding in vitro and transforming activity
in vivo. Mol Cell Biol. 12:609–618. 1992.PubMed/NCBI
|
|
25
|
Yin J, Cai Z, Zhang L, et al: A recombined
fusion protein PTD-Grb2-SH2 inhibits the proliferation of breast
cancer cells in vitro. Int J Oncol. 42:1061–1069.
2013.PubMed/NCBI
|
|
26
|
Watanabe T, Shinohara N, Moriya K, et al:
Significance of the Grb2 and son of sevenless (Sos) proteins in
human bladder cancer cell lines. IUBMB Life. 49:317–320. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Giubellino A, Burke TR Jr and Bottaro DP:
Grb2 signaling in cell motility and cancer. Expert Opin Ther
Targets. 12:1021–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Smithgall TE: SH2 and SH3 domains:
potential targets for anti-cancer drug design. J Pharmacol Toxicol
Methods. 34:125–132. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ursini-Siegel J, Hardy WR, Zheng Y, et al:
The ShcA SH2 domain engages a 14-3-3/PI3’K signaling complex and
promotes breast cancer cell survival. Oncogene. 31:5038–5044.
2012.PubMed/NCBI
|
|
30
|
Dierck K, Machida K, Mayer BJ and Nollau
P: Profiling the tyrosine phosphorylation state using SH2 domains.
Methods Mol Biol. 527:131–155. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gril B, Vidal M, Assayag F, Poupon MF, Liu
WQ and Garbay C: Grb2-SH3 ligand inhibits the growth of
HER2+ cancer cells and has antitumor effects in human
cancer xenografts alone and in combination with docetaxel. Int J
Cancer. 121:407–415. 2007.PubMed/NCBI
|
|
32
|
Yu GZ, Chen Y and Wang JJ: Overexpression
of Grb2/HER2 signaling in Chinese gastric cancer: their
relationship with clinicopathological parameters and prognostic
significance. J Cancer Res Clin Oncol. 135:1331–1339. 2009.
View Article : Google Scholar : PubMed/NCBI
|