Introduction
Hepatocellular carcinoma (HCC) is the fifth leading
cause of cancer-related mortality in the world (1,2). The
high recurrence rate and poor prognosis of HCC are responsible for
its high mortality. Even though surgery, locoregional therapy,
transcatheter arterial chemoembolization (3) and chemotherapy are available for the
treatment of HCC, they have offered limited success in reducing
cancer-related mortality. Target therapy with the multiple tyrosine
kinase inhibitor sorafenib has demonstrated improved therapeutic
potential for HCC patients and has prolonged their survival
(4), yet more therapeutic targets
are awaiting identification for HCC.
Among the multiple hallmarks of tumor development,
dependence on glycolysis is one of the major characteristics in
various types of cancers including HCC (5–7), and
it is not only a critical issue in cancer research but also serves
as a therapeutic target for cancer treatment (8–10). In
addition to glucose metabolism, cancer cells also remodel the
metabolism of many macromolecules, including amino acids (6,11) and
fatty acids (12–14), to support the formation of
neoplasia. We previously demonstrated that dysregulation of
argininosuccinate lyase (ASL) (15)
and long-chain acyl-CoA synthetase (ACSL) (16) contributes to the development of HCC.
Fatty acid serves as one of the major energy providers for cellular
metabolism, and its involvement in tumor development has been
demonstrated by the upregulation of fatty acid synthase (FASN)
during cancer formation, and the expression of FASN even correlates
with increased disease progression and decreased patient survival
(14). However, recent studies also
revealed that lipolysis plays a critical role in cancer
pathogenesis. In human multiple myeloma cancer cells, inhibition of
fatty acid degradation was found to reduce proliferation (13). In various types of cancers (17) including colorectal cancer (18), silencing of the lipolysis-related
enzyme monoacylglycerol lipase (MAGL) was found to reduce tumor
formation. In lipolysis-related enzyme acyl-CoA thioesterase
superfamily member 1 (ACOT11) knockout mice, the animals were
protected against disease-prone factors, such as obesity and
insulin resistance (19).
Therefore, production of fatty acid by either de novo
synthesis or by reduction in its degradation is important in cancer
development (20–22).
Among the lipolysis-related enzymes, members of the
acyl-CoA thioesterase (ACOT) superfamily control an important step
in lipid utilization by catalyzing the breakdown of fatty acyl-CoA
into free fatty acid (FFA) and coenzyme A (CoA) molecule. To date,
15 ACOTs have been identified and are divided according to
structure into two superfamilies; the first one is the
α/β-hydrolase superfamily which includes ACOT1–6, and the second
one is hotdog-fold protein superfamily which includes ACOT7–15.
Different ACOTs display different specificity toward fatty acyl-CoA
of different chain length and reside in different cellular
compartments (23–25). Studies addressing the functions of
distinct ACOTs by means of transgenic mice including
ACOT7−/− (26),
ACOT11−/− (19,27) and ACOT13−/− (28,29)
revealed that different ACOTs may have diverse functions in
regulating lipid metabolism. Among these ACOTs, ACOT8 reacts with a
wide range of substrates, including short-, medium- and long-chain
fatty acids (25), implying its
essential role in lipid utilization and regulation of lipid
metabolism for fatty acids of nearly all chain lengths. Therefore,
dysregulation of ACOT8 is expected to have a strong association
with human disease. Indeed, ACOT8 is associated with ovarian cancer
and lung adenocarcinoma. Increased ACOT8 gene copy number has been
observed in clinical specimens from patients with ovarian cancer
(30), and high ACOT8 expression is
associated with metastasis as well as poor outcome in patients with
lung adenocarcinoma (31). However,
the importance of ACOT8 in HCC, which is one of the cancer types
seriously impacted by dysregulated lipid metabolism, has not yet
been investigated. In the present study, we first identified the
importance of ACOT8 during HCC formation by performing
bioinformatic analysis of published microarrays with HCC clinical
specimens in the GEO database (32), and found that ACOT8 gene copy number
and mRNA expression were increased in HCC tissues when compared to
these variables in non-tumor tissues. We then knocked down ACOT8
expression in two HCC cell lines, and observed a decrease in
anchorage-dependent and -independent growth in the cancer cells.
The reduction in cell growth was compensated by reintroduction of
ACOT8 or partially recovered by the addition of FFA myristic acid.
Collectively, ACOT8 plays an important role in the formation of
HCC, and downregulation of ACOT8 inhibits the growth of tumor
cells.
Materials and methods
Data collection of microarrays regarding
human HCC clinical specimens
Microarrays regarding human HCC clinical specimens
published from 2004 to 2013 were collected from the public resource
NCBI Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo). Microarrays
regarding HCC cell lines were not included. Four microarray
datasets GSE14322 (33), GSE1898
(34), GE25097 (35) and GSE10140-GSE10141 (36) were collected, and the change in
ACOT8 gene copy number and mRNA expression in these microarrays was
significantly different in the HCC tissues when compared to these
variables in the non-tumor tissues. We downloaded the normalized
signals for each microarray directly from NCBI Gene Expression
Omnibus, and these normalized signals were further analyzed to
identify the tendency of the change in ACOT8 gene copy number or
mRNA expression in HCC tissues. The result of this analysis was
assessed for statistical significance and diagrammed using GraphPad
Prism, version 5 (GraphPad Software, San Diego, CA, USA). The
detailed information of these microarrays is listed in Table I.
 | Table ICharacteristics of the collected GEO
Datasets. |
Table I
Characteristics of the collected GEO
Datasets.
| GEO accession
No. | References | Type of
microarray | No. of tumors | No. of normal
DNA |
|---|
| GSE14322 | Roessler, et
al (33) | Genome
variation | 76 | -a |
| GSE1898 | Lee, et al
(34) | Expression | 91 | 18 |
| GSE25097 | Tung, et al
(35) | Expression | 268 | 243 |
| GSE10140,
GSE10141 | Hoshida, et
al (36) | Expression | 80 | 82 |
Cell lines
Human HCC cell line Huh7 was kindly provided by I.J.
Su of the National Health Research Institute. Huh7 and another
human HCC cell line Hep3B as well as their shACOT8 stable
transfectant clones were cultured in DMEM containing 10% FBS
(Biological Industries, Beit Haemek, Israel) and 1%
penicillin-streptomycin. Cells were kept in an incubator at 37°C
with 5% CO2.
Chemicals, reagents, plasmids and
antibodies
Ethidium bromide, sodium dodecyl sulfate (SDS),
G418, myristic acid, myristoyl-CoA lithium salt, and
5,5-dithiobis-(2-nitrobenzoic acid) (DTNB) were purchased from
Sigma (St. Louis, MO, USA). The Micro BCA™ protein assay reagent
kit was purchased from Pierce (Woburn, MA, USA). DMEM and an
antibiotic mixture were purchased from Invitrogen (Carlsbad, CA,
USA). Turbofect transfection reagent was purchased from Fermentas
(Glen Burnie, MD, USA). Antibodies against ACOT8 (NBP1-66562; Novus
Biologicals, Littleton, CO, USA) and tubulin (GTX628802; GeneTex,
Taiwan) were used in the western blot analysis.
RNA interference
The shACOT8 plasmid-carrying bacterial clone was
obtained from the RNAi core facility (Academia Sinica, Taiwan), and
the target sequence used was 5′-CAT TGGCGCTCAACCGAATTG-3′. Cells
were transfected with shRNA against luciferase as vector control or
against ACOT8 and selected for stable transfectant clones with
puromycin.
Plasmid construction
Human ACOT8 (NM_005469.3) was constructed into the
HA6L vector as previously described (37). Cells were transfected with the
ACOT8-overexpression plasmid and selected for stable transfectant
clones with G418. The expression of ACOT8 in these clones was
confirmed at the protein level.
Western blot analysis
Cells were lysed in modified RIPA buffer with
protease inhibitors and let stand on ice for 20 min. After
centrifugation at 12,000 rpm, at 4°C for 10 min, the supernatant
was harvested and the protein concentration was assayed using the
Micro BCA protein assay reagent kit. Samples with the same amount
of protein and 4X sample buffer were mixed, heated at 95°C for 5
min, and subjected to electrophoresis. The protein was then
transferred onto a PVDF membrane (Millipore, Bedford, MA, USA)
using the Hoefer Semiphor Semi-Dry transfer unit (Amersham
Pharmacia, San Francisco, CA, USA), and blocked in 5% non-fat milk
at room temperature for 1 h. Specific antibodies were added to the
probe targets overnight at 4°C. Following washing with 0.1% TBS-T
for 10 min for 3 times, the membrane was probed with the specific
secondary antibody at room temperature for 1 h and washed with 0.1%
TBS-T. The quantity of target proteins and internal control were
identified by adding chemiluminescence reagent ECL (Millipore) onto
the membrane and the luminescent intensity was recorded using the
BioSpectrum AC imaging system (UVP, Upland, CA, USA).
Thioesterase activity assay
The thioesterase activity assay was performed
according to a previous report (19) with some modifications. Total protein
was harvested from cells in homogenization buffer (1% Tergitol
NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 150 mM NaCl, 50 mM
Tris-HCl and 2 mM EDTA) and was allowed to remain in ice for 20
min. After centrifugation at 12,000 rpm at 4°C for 10 min, the
supernatant was harvested and the protein concentration was assayed
using the Micro BCA protein assay reagent kit. Protein at the
indicated amount was added, and the reaction volume was adjusted
with the reaction buffer [50 mM KCl, 10 mM HEPES (pH 7.5) and 0.3
mM DTNB] to a final volume of 200 μl. Myristoyl-CoA at the final
concentration of 200 nM was then added to start the thioesterase
activity assay. Thioesterase activity was analyzed at 5-min
intervals for a total of 1 h with the ELISA reader at a wavelength
of 405 nm.
Colony formation assay
A total of 5×102 Huh7 cells,
1×103 Hep3B cells and same amount of cells for their
stable transfectant clones of ACOT8 knockdown or compensation were
seeded onto 6-well plates and cultured for 9 days to assay the
anchorage-dependent growth ability of the cancer cells. The number
of colonies was identified by methyl blue staining, counted under a
microscope and analyzed.
Soft agar assay
Cells (5×103) were seeded into 1 ml 0.3%
agar-containing medium and then onto 0.6% agar-covered 6-well
plates and cultured for 14 days to assay the anchorage-independent
growth ability of the cancer cells. The number of colonies was
identified by crystal violet staining, counted under a microscope
and analyzed.
Statistical analysis
All statistical analyses were performed by the
GraphPad Prism, version 5 (GraphPad Software, San Diego, CA, USA).
All error bars in the figures represent SEM. Student’s t-test and
two-way ANOVA followed by Bonferroni post-test were used for
analysis of difference between each experimental group, and a
P-value of <0.05 was considered to indicate a statistically
significant difference.
Results
Bioinformatic analysis of the ACOT8 gene
copy number and mRNA expression in the HCC clinical specimens
To investigate the importance of ACOT8 dysregulation
in HCC tumorigenesis in terms of human cancer patients, we
performed bioinformatic analysis of the published microarray data
with HCC tissues and non-tumor tissues from the GEO database
(32). The analysis revealed that
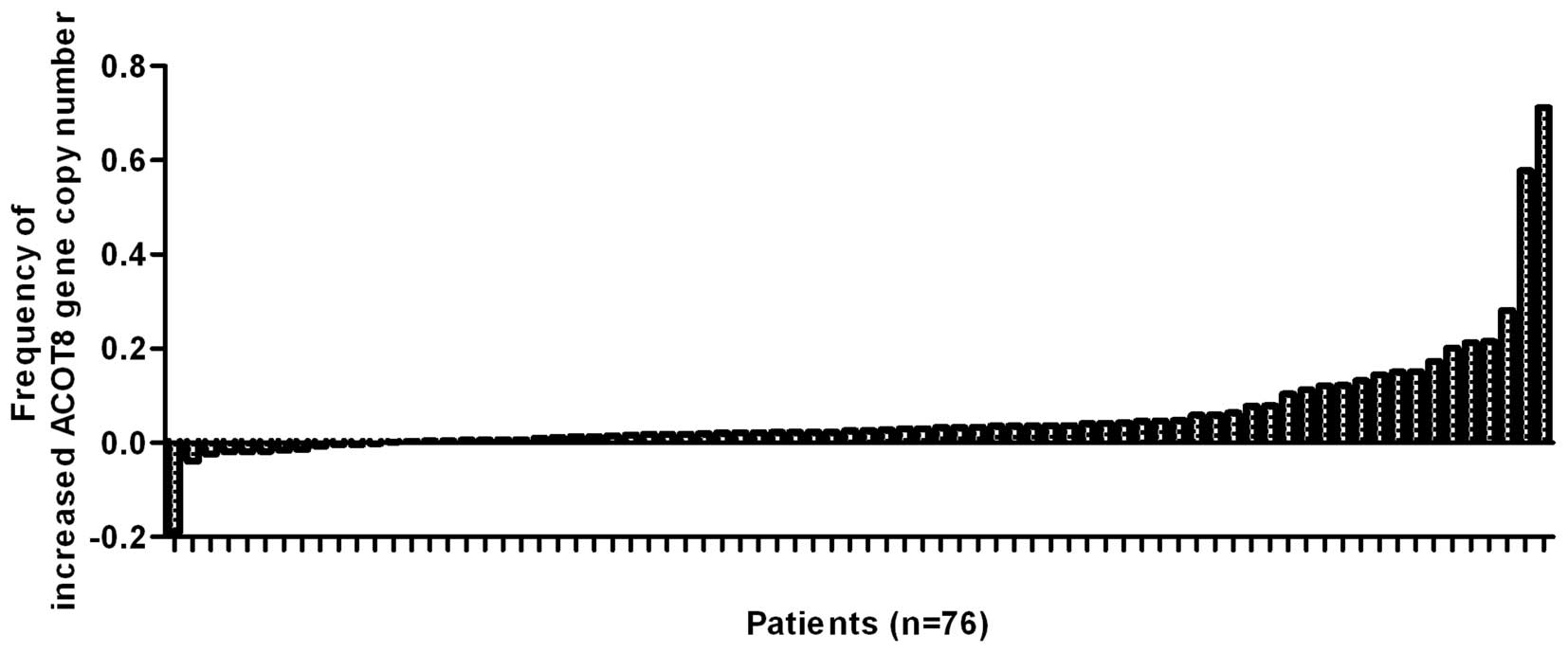
ACOT8 gene copy number was increased in the HCC tissues when
compared to that in the normal DNA in microarray GSE14322 (Fig. 1) from the US, since 64 of the 76 HCC
patients in GSE14322 (33)
displayed increased ACOT8 gene copy number than that of normal DNA.
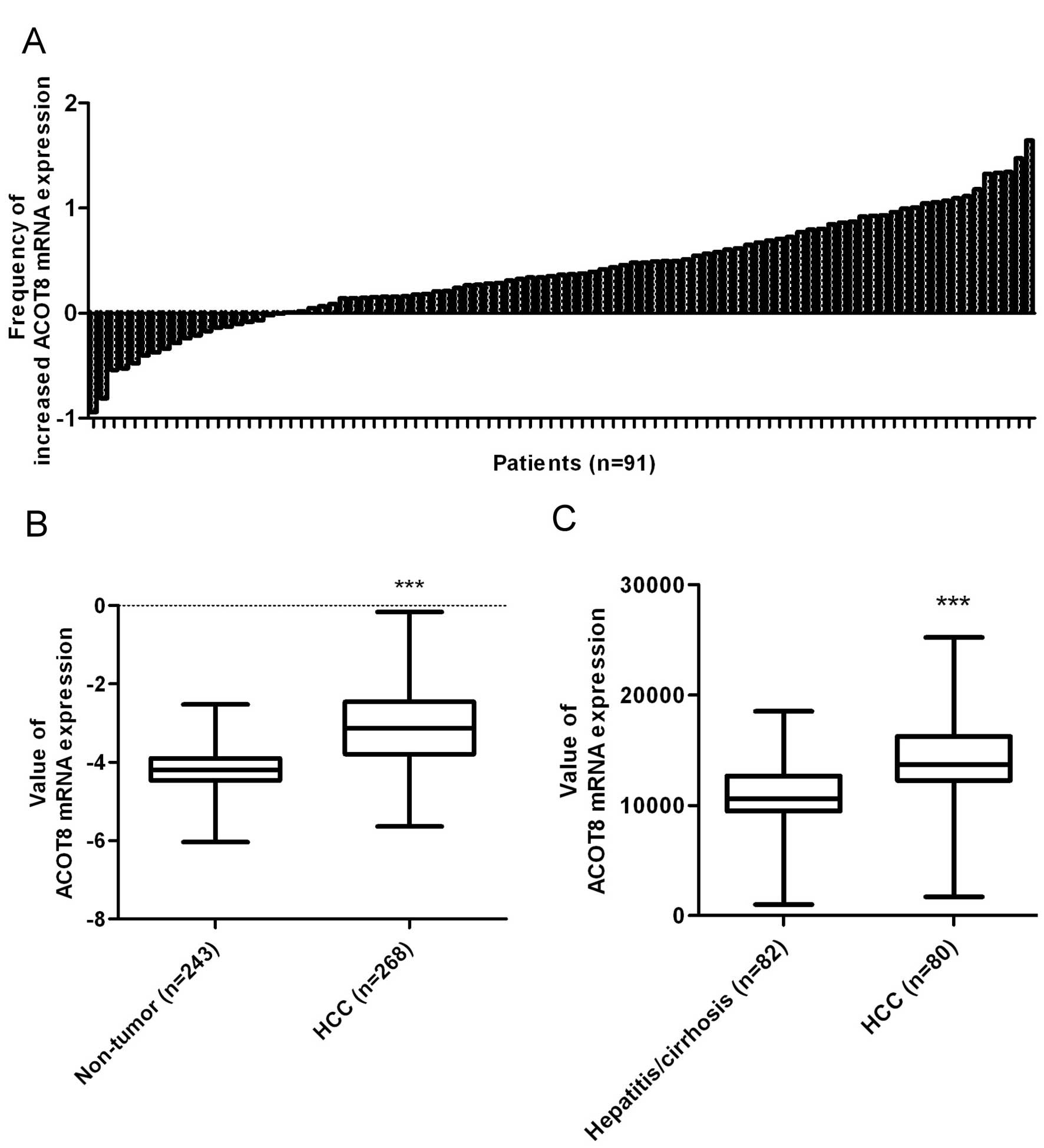
In addition, ACOT8 mRNA expression was significantly increased in
HCC tissues compared to the expression in the non-tumor tissues in
microarrays GSE1898 (Fig. 2A),
GSE25097 (Fig. 2B) and
GSE10140-GSE10141 (Fig. 2C). The
above phenomena were observed in 73 of 91 HCC patients in GSE1898
(34), 268 HCC tissues vs. 243
non-tumor tissues in GSE25097 (35)
and 80 HCC tissues vs. 82 non-tumor tissues in GSE10140-GSE10141
(36). This bioinformatic analysis
revealed that the dysregulation in ACOT8, particularly upregulation
in both gene copy number and mRNA expression, is associated with
HCC development in respect to human cancer patients.
ACOT8 modulates in vitro tumorigenesis
and thioesterase activity in HCC
To study the contribution of ACOT8 expression during
HCC formation in detail, shRNA against ACOT8 was transfected into
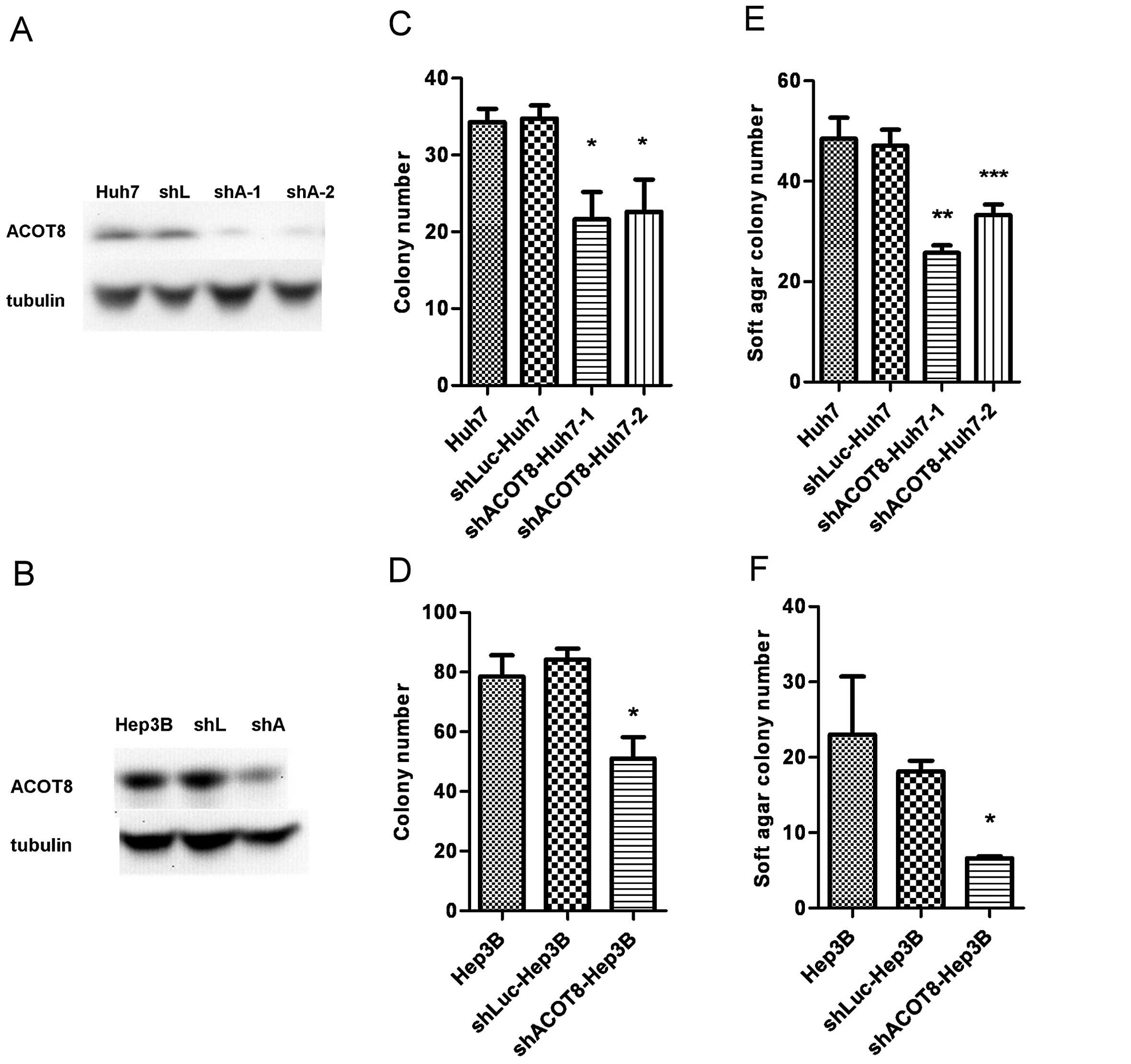
the Huh7 and Hep3B HCC cell lines, and stable transfectant clones
were established. Knockdown efficiency was confirmed at the protein
level (Fig. 3A and B). The ACOT8
shRNA stable transfectants were then subjected to in vitro
tumorigenic and thioesterase activity assays to determine the role
of ACOT8 in HCC development. Knockdown of ACOT8 caused a
significant decrease in anchorage-dependent (Fig. 3C and D) and anchorage-independent
growth (Fig. 3E and F) in both HCC
cell lines, indicating that ACOT8 silencing reduced the in
vitro tumorigenicity of HCC. We also investigated the effect of
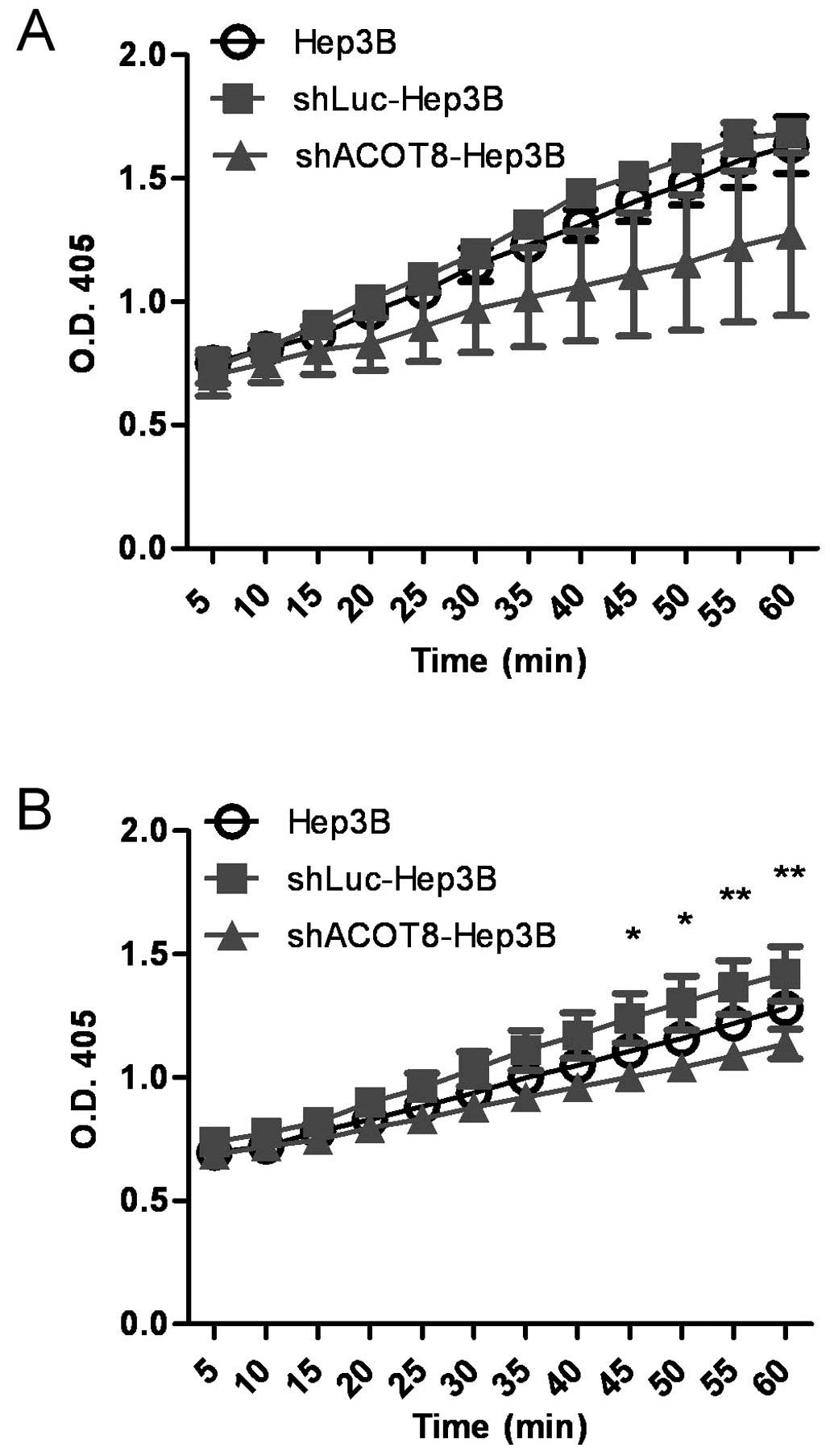
ACOT8 knockdown on thioesterase activity to determine the possible
involvement of the lipolytic role of ACOT8 during HCC formation.
The alteration in thioesterase activity was measured by the change
in absorbance at 405 nm resulting from the reaction between ACOT
and DTNB. The ACOT-mediated hydrolysis of fatty acyl-CoA and the
release of free CoA molecules lead to a rapid reaction between DTNB
and CoA, giving rise to the production of 5-thio-2-nitrobenzoate,
whose absorbance can be detected at 412 nm (19) or 405 nm (present study). Indeed, the
ACOT8-knockdown clones displayed a reduced absorbance value and
thus reduced thioesterase activity when compared to that of the
vector control clones (Fig. 4A and
B), indicating ACOT8 plays a major role in total lipolysis. To
further exclude the off-target effect of ACOT8 shRNA in the above
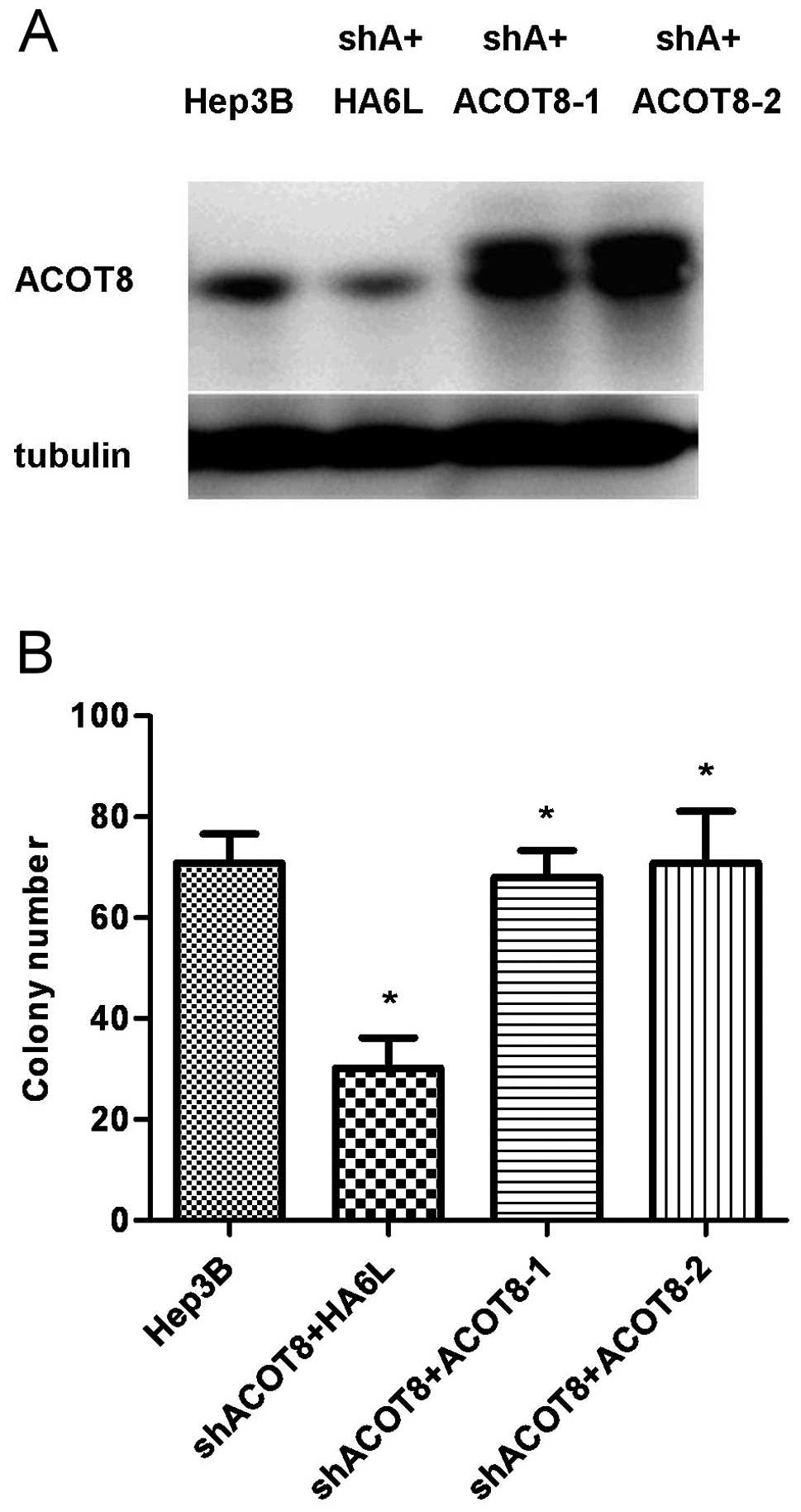
experiments, ACOT8 was reintroduced into the ACOT8-knockdown clones
in Hep3B cells (Fig. 5A), and the
cell growth was restored as determined by a colony formation assay
(Fig. 5B), further supporting the
important role of ACOT8 in HCC formation.
ACOT8-mediated HCC tumorigenesis is
dependent on the FFA level
To elucidate the metabolic mechanisms involved in
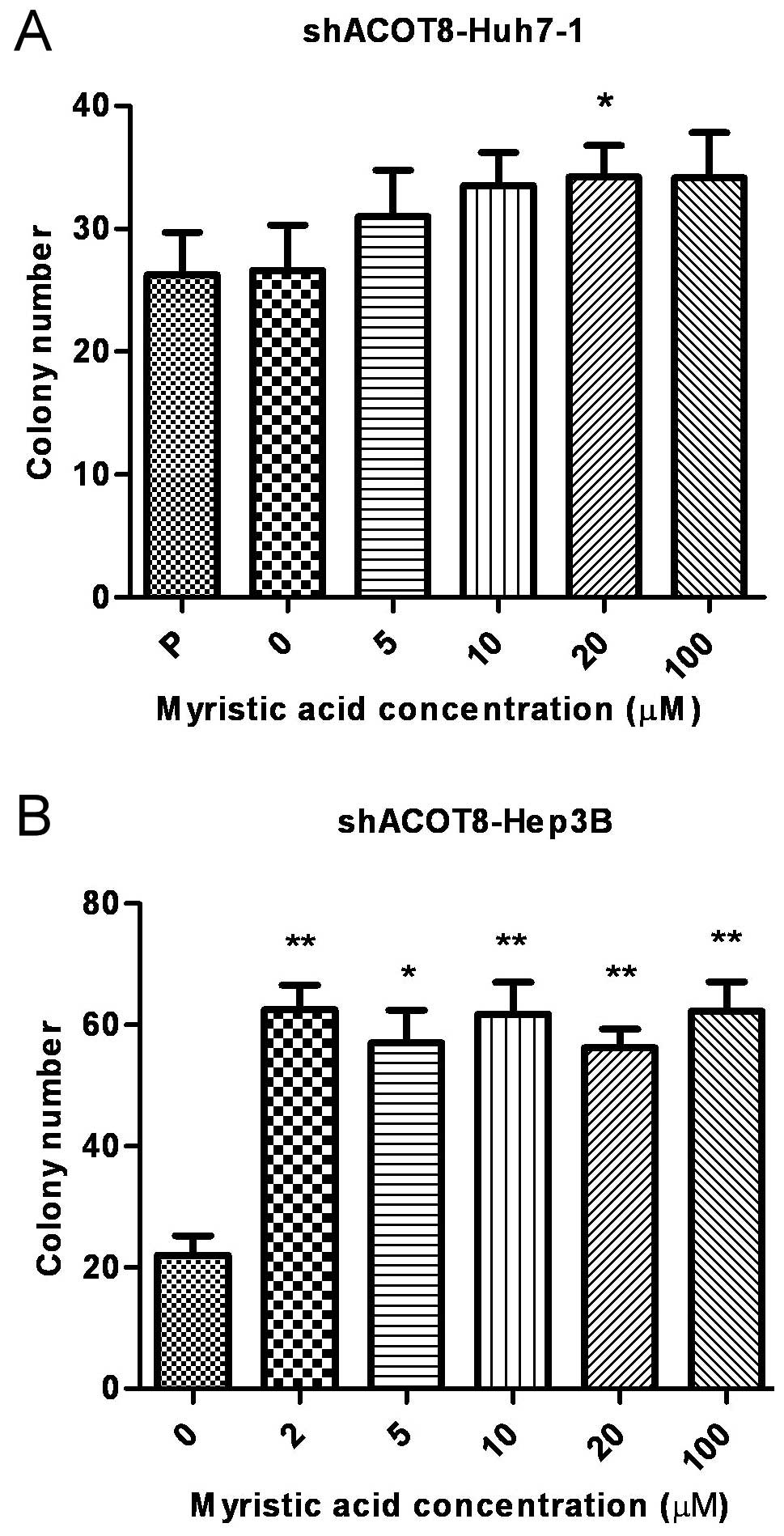
ACOT8-modulated HCC development, we examined the effect of FFA
metabolites on the ACOT8 knockdown-reduced tumorigenesis. One of
the metabolic products downstream of ACOT8-mediated reaction, the
FFA myristic acid, was added to the medium to ascertain whether FFA
affects the growth inhibition induced by ACOT8 knockdown. The
addition of myristic acid indeed led to the increased growth of
cancer cells as determined by a colony formation assay in
ACOT8-knockdown stable transfectants of Huh7 and Hep3B HCC cell
lines (Fig. 6A and B). The effects
of myristic acid on restoration of the cell growth were more
prominent in the Hep3B stable transfectants. This result revealed
that the metabolite downstream of the ACOT8-mediated reaction was
able to rescue ACOT8 knockdown-decreased HCC cell growth, further
supporting the notion that lipolysis enzyme ACOT8 plays an
important role in HCC development.
Discussion
In the present study, we identified the importance
of lipolysis in HCC development by determining the contribution of
an acyl-CoA degrading lipolytic enzyme ACOT8 in HCC formation. ACOT
is a family of enzymes controlling one of the critical steps in
lipid utilization by catalyzing the breakdown of fatty acyl-CoA
into FFA and CoA molecules. ACOT8 is one of the family members with
a wide range of substrates, which also indicates its possible
involvement in disease progression or even cancer formation.
Indeed, the participation of ACOT8 in ovarian cancer (30) and lung adenocarcinoma (31) has been reported in a clinical
respect, while the impact of ACOT8 expression on other cancer types
which are seriously influenced by dysregulated lipid metabolism
such as HCC and whether the lipolytic role of ACOT8 participates in
these processes remain to be elucidated. Aiming to identify the
importance of ACOT8 in HCC tumorigenesis and the mechanism
involved, we first investigated the expression pattern of ACOT8 in
HCC patients. By performing bioinformatic analysis on published
microarray data with HCC clinical specimens in the GEO database, we
determined that both gene copy number and mRNA expression of ACOT8
were increased in HCC tissues compared to these variables in the
non-tumor tissues, indicating the association between ACOT8
expression and HCC tumorigenesis in clinical specimens. To further
ascertain the effect of ACOT8 expression on HCC tumorigenesis, we
performed in vitro tumorigenic and thioesterase activity
assays following ACOT8 knockdown in HCC cell lines. ACOT8 knockdown
reduced not only anchorage-dependent and -independent growth but
also thioesterase activity in the cancer cells, suggesting the
importance of ACOT8 in modulating HCC development. ACOT8
compensation in shRNA-knockdown clones increased cancer cell
growth, which further supports the critical role of ACOT8 in HCC
formation. The involvement of lipid metabolism in ACOT8-modulated
HCC tumorigenesis was further demonstrated by the rescue of cell
growth via addition of FFA myristic acid, indicating
ACOT8-modulated HCC formation is partially, if not totally,
regulated by its lipolytic role.
Since lipid metabolism is one of the important
characteristics in the formation of neoplasias, numerous studies
have investigated the role of lipolytic enzymes in cancer
development (13,17,18,22).
The functions of ACOT family members in normal physiology and
diseases have also been studied (23–29,31).
In addition to findings that ACOT8 gene copy number and expression
are increased, respectively, in ovarian cancer and lung
adenocarcinoma (30,31), ACOT2 and ACOT4 have also be
suggested to participate in breast cancer due to their
overexpression (38,39), with the finding that ACOT2 modulates
the production of prostaglandin in breast cancer cells (38). In the present study, myristic acid,
the 14-carbon saturated fatty acid, was able to increase the cell
growth of ACOT8 shRNA stable transfectants to the level comparable
to that of the vector control. Saturated fatty acids have been
reported to have pro-apoptosis effects on normal and cancer cells
due to lipotoxicity, but they have also been shown to enhance
production of the pro-inflammatory cytokine interleukin-8 (IL-8) to
interrupt lipid metabolism (40) in
hepatocytes and HCC cells. Saturated fatty acids have also been
linked to the increased risk of developing prostate cancer in a
Japanese cohort (41). These
findings indicate that the diverse functions of saturated fatty
acids operate in a cell context-dependent fashion, and our study
which supports a possible tumorigenic role of saturated fatty acid
in HCC development is in accordance with these reports.
In conclusion, we reported the importance of
acyl-CoA degrading lipolytic enzyme ACOT8 in the development of HCC
by demonstrating that its gene copy number and mRNA expression are
increased in HCC patients, while its knockdown reduced both in
vitro tumorigenesis and thioesterase activity in HCC cells. To
the best of our knowledge, this is the first report regarding the
contribution of ACOT8 in HCC development, implying the importance
of lipolysis in HCC formation.
Acknowledgements
This study was supported by the grant to M.-D. Lai,
NSC-100-2325-B-006-008 from the National Science Council, Taiwan;
NHRI-EX100-9927B1 from the National Health Research Institute,
Taiwan; to Establish Centers of Excellence for Cancer Research in
Taiwan, DOH101-TD-C-111-003 Department of Health, Executive Yuan,
Taiwan, R.O.C.
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
ACOT
|
acyl-CoA thioesterase
|
|
FFA
|
free fatty acid
|
|
CoA
|
coenzyme A
|
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thorgeirsson SS and Grisham JW: Molecular
pathogenesis of human hepatocellular carcinoma. Nat Genet.
31:339–346. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bruix J, Sala M and Llovet JM:
Chemoembolization for hepatocellular carcinoma. Gastroenterology.
127:S179–S188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhu AX: Development of sorafenib and other
molecularly targeted agents in hepatocellular carcinoma. Cancer.
112:250–259. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kroemer G and Pouyssegur J: Tumor cell
metabolism: cancer’s Achilles’ heel. Cancer Cell. 13:472–482.
2008.
|
|
6
|
Ward PS and Thompson CB: Metabolic
reprogramming: a cancer hallmark even Warburg did not anticipate.
Cancer Cell. 21:297–308. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu RH, Pelicano H, Zhou Y, et al:
Inhibition of glycolysis in cancer cells: a novel strategy to
overcome drug resistance associated with mitochondrial respiratory
defect and hypoxia. Cancer Res. 65:613–621. 2005.
|
|
8
|
Pelicano H, Martin D, Xu RH and Huang P:
Glycolysis inhibition for anticancer treatment. Oncogene.
25:4633–4646. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vander Heiden MG: Targeting cancer
metabolism: a therapeutic window opens. Nat Rev Drug Discov.
10:671–684. 2011.PubMed/NCBI
|
|
10
|
Tennant DA, Durán RV and Gottlieb E:
Targeting metabolic transformation for cancer therapy. Nat Rev
Cancer. 10:267–277. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Locasale JW: Serine, glycine and
one-carbon units: cancer metabolism in full circle. Nat Rev Cancer.
13:572–583. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Menendez JA and Lupu R: Oncogenic
properties of the endogenous fatty acid metabolism: molecular
pathology of fatty acid synthase in cancer cells. Curr Opin Clin
Nutr Metab Care. 9:346–357. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tirado-Vélez JM, Joumady I, Sáez-Benito A,
Cózar-Castellano I and Perdomo G: Inhibition of fatty acid
metabolism reduces human myeloma cells proliferation. PloS One.
7:e464842012.PubMed/NCBI
|
|
14
|
Menendez JA and Lupu R: Fatty acid
synthase and the lipogenic phenotype in cancer pathogenesis. Nat
Rev Cancer. 7:763–777. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang HL, Hsu HP, Shieh SC, et al:
Attenuation of argininosuccinate lyase inhibits cancer growth via
cyclin A2 and nitric oxide. Mol Cancer Ther. 12:2505–2516. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang YS, Tsai CT, Huangfu CA, et al:
ACSL3 and GSK-3β are essential for lipid upregulation induced by
endoplasmic reticulum stress in liver cells. J Cell Biochem.
112:881–893. 2011.PubMed/NCBI
|
|
17
|
Nomura DK, Long JZ, Niessen S, Hoover HS,
Ng SW and Cravatt BF: Monoacylglycerol lipase regulates a fatty
acid network that promotes cancer pathogenesis. Cell. 140:49–61.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye L, Zhang B, Seviour EG, et al:
Monoacylglycerol lipase (MAGL) knockdown inhibits tumor cell growth
in colorectal cancer. Cancer Lett. 307:6–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Li Y, Niepel MW, et al: Targeted
deletion of thioesterase superfamily member 1 promotes energy
expenditure and protects against obesity and insulin resistance.
Proc Natl Acad Sci USA. 109:5417–5422. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nieman KM, Kenny HA, Penicka CV, et al:
Adipocytes promote ovarian cancer metastasis and provide energy for
rapid tumor growth. Nat Med. 17:1498–1503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaini RR, Sillerud LO, Zhaorigetu S and Hu
CA: Autophagy regulates lipolysis and cell survival through lipid
droplet degradation in androgen-sensitive prostate cancer cells.
Prostate. 72:1412–1422. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li J, Zhao S, Zhou X, et al: Inhibition of
lipolysis by mercaptoacetate and etomoxir specifically sensitize
drug-resistant lung adenocarcinoma cell to paclitaxel. PloS One.
8:e746232013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hunt MC and Alexson SE: The role acyl-CoA
thioesterases play in mediating intracellular lipid metabolism.
Prog Lipid Res. 41:99–130. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hunt MC, Yamada J, Maltais LJ, Wright MW,
Podesta EJ and Alexson SE: A revised nomenclature for mammalian
acyl-CoA thioesterases/hydrolases. J Lipid Res. 46:2029–2032. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hunt MC, Siponen MI and Alexson SE: The
emerging role of acyl-CoA thioesterases and acyltransferases in
regulating peroxisomal lipid metabolism. Biochim Biophys Acta.
1822:1397–1410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ellis JM, Wong GW and Wolfgang MJ: Acyl
Coenzyme A thioesterase 7 regulates neuronal fatty acid metabolism
to prevent neurotoxicity. Mol Cell Biol. 33:1869–1882. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han S and Cohen DE: Functional
characterization of thioesterase superfamily member 1/Acyl-CoA
thioesterase 11: implications for metabolic regulation. J Lipid
Res. 53:2620–2631. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kang HW, Ozdemir C, Kawano Y, et al:
Thioesterase superfamily member 2/Acyl-CoA thioesterase 13
(Them2/Acot13) regulates adaptive thermogenesis in mice. J Biol
Chem. 288:33376–33386. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kang HW, Niepel MW, Han S, Kawano Y and
Cohen DE: Thioesterase superfamily member 2/acyl-CoA thioesterase
13 (Them2/Acot13) regulates hepatic lipid and glucose metabolism.
FASEB J. 26:2209–2221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ramakrishna M, Williams LH, Boyle SE, et
al: Identification of candidate growth promoting genes in ovarian
cancer through integrated copy number and expression analysis. PloS
One. 5:e99832010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jung WY, Kim YH, Ryu YJ, et al: Acyl-CoA
thioesterase 8 is a specific protein related to nodal metastasis
and prognosis of lung adenocarcinoma. Pathol Res Pract.
209:276–283. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Edgar R, Domrachev M and Lash AE: Gene
Expression Omnibus: NCBI gene expression and hybridization array
data repository. Nucleic Acids Res. 30:207–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Roessler S, Long EL, Budhu A, et al:
Integrative genomic identification of genes on 8p associated with
hepatocellular carcinoma progression and patient survival.
Gastroenterology. 142:957–966. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee JS, Chu IS, Mikaelyan A, et al:
Application of comparative functional genomics to identify best-fit
mouse models to study human cancer. Nat Genet. 36:1306–1311. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tung EK, Mak CK, Fatima S, et al:
Clinicopathological and prognostic significance of serum and tissue
Dickkopf-1 levels in human hepatocellular carcinoma. Liver Int.
31:1494–1504. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hoshida Y, Villanueva A, Kobayashi M, et
al: Gene expression in fixed tissues and outcome in hepatocellular
carcinoma. N Engl J Med. 359:1995–2004. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lu TJ, Lai WY, Huang CY, et al: Inhibition
of cell migration by autophosphorylated mammalian sterile 20-like
kinase 3 (MST3) involves paxillin and protein-tyrosine
phosphatase-PEST. J Biol Chem. 281:38405–38417. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Maloberti PM, Duarte AB, Orlando UD, et
al: Functional interaction between acyl-CoA synthetase 4,
lipooxygenases and cyclooxygenase-2 in the aggressive phenotype of
breast cancer cells. PloS One. 5:e155402010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sun Z, Asmann YW, Kalari KR, et al:
Integrated analysis of gene expression, CpG island methylation, and
gene copy number in breast cancer cells by deep sequencing. PloS
One. 6:e174902011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Joshi-Barve S, Barve SS, Amancherla K, et
al: Palmitic acid induces production of proinflammatory cytokine
interleukin-8 from hepatocytes. Hepatology. 46:823–830. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kurahashi N, Inoue M, Iwasaki M, Sasazuki
S and Tsugane S: Dairy product, saturated fatty acid, and calcium
intake and prostate cancer in a prospective cohort of Japanese men.
Cancer Epidemiol Biomarkers Prev. 17:930–937. 2008. View Article : Google Scholar : PubMed/NCBI
|