Introduction
Previous reports have revealed that chronic
inflammation can promote the occurrence, growth and metastasis of
cancer. It is now widely accepted that acute inflammation will
develop into chronic inflammation if not controlled in time, and
chronic inflammation may increase the risk of cancer.
Epidemiological studies have confirmed that 25% of cancers develop
from inflammation. For example, patients with ulcerative colitis
are 10 times more likely to develop colon cancer than normal
individuals, and data show that 20% of patients with ulcerative
colitis develop colon cancer 30 years later (1). The accumulation of inflammatory cells
and inflammatory factors in the tumor microenvironment can promote
angiogenesis, proliferation and metastasis of malignant cells,
promote epithelial-mesenchymal transition (EMT) and reverse
acquired immune response (2). It also
alters the sensitivity of tumor cells to hormones and
chemotherapeutic drugs.
Furthermore, cytokines serve an important role in
the development of inflammation. Cytokines can be divided into
lymphokines produced by lymphocytes and monocytes produced by
monocytes and macrophages. Interleukin (IL), interferon, colony
stimulating factor, tumor necrosis factor and transforming growth
factor are known cytokines produced by immune cells. Cytokines in
the tumor microenvironment usually serve an antitumor role, but in
chronic inflammation, abnormal expression of cytokines can promote
the transformation of surrounding normal cells into cancer cells in
the early stage of cancer, and promote the growth and development
of cancer cells (3).
Among them, IL-17F is located on chromosome
6p12 with a total length of 7,742 bp. It consists of 3 exons and 2
introns. IL-17F is produced by T helper (Th)17 cells (4). Th17 cells associate innate immunity with
adaptive immunity via chemokines and induce other Th cell subsets
to aggregate at the site of infection in the late stage of
inflammation. As a prototype cytokine of the IL-17 family, IL-17F
induces different tissues and cells to produce inflammatory
cytokines, chemokines and metalloproteinases, and collects
centralized granulocytes from tissues. Current research results
have confirmed that Th17 cytokines such as IL-17F can promote the
synthesis of inflammatory cells, increase the levels of chemokines
and promote the expression of tissue degradation matrix
metalloproteinases, thus causing tissue damage (5). However, there are few studies on the
association between IL-17F and cancer, particularly colorectal
cancer (CRC). In the present study, we found that IL-17F
gene is abnormally expressed in CRC tissues. However, the mechanism
by which IL-17F is activated and has an effect on CRC tissues
remains unclear. Therefore, it is of great significance to study
the differential expression of the IL-17F gene in CRC, as well as
the molecular mechanism and signaling pathway in the development of
CRC.
Materials and methods
Patients and specimens
CRC tumor specimens and paired non-tumor mucosa were
collected from July 2012 to July 2018 in Shanghai Pudong Hospital.
Patients with the following criteria were excluded from
participation: Patients that had received adjuvant chemotherapy or
radiotherapy prior to surgery or had been diagnosed with additional
cancer types. All patients were classified according to the 7th
edition of the Tumor-Node-Metastasis (TNM) staging system
(www.cancerstaging.org). Postoperative
adjuvant therapies were performed according to standard schedules
and doses. All participating patients provided their written
informed consent. The present study was approved by the Ethics
Committee of Shanghai Pudong Hospital. Patients' data was shown in
Table I.
 | Table I.Clinical characteristics of the
colorectal cancer patients (N=67). |
Table I.
Clinical characteristics of the
colorectal cancer patients (N=67).
| UICC (TNM) stage |
|
|---|
| I | 7 |
| II | 27 |
| III | 25 |
| IV | 8 |
| Tumor (T) stage |
|
|
pTis-1 | 2 |
| pT2 | 17 |
| pT3 | 43 |
| pT4 | 5 |
| N stage |
|
| N0 | 33 |
| N1 | 30 |
| N2 | 4 |
| M stage |
|
| M0 | 60 |
| M1 | 7 |
| Age (years) |
|
|
<65 | 30 |
|
65-75 | 20 |
|
>75-85 | 16 |
|
>85 | 1 |
| Sex |
|
| Male | 40 |
|
Female | 27 |
| Tumor location |
|
| Right
colon | 15 |
| Left
colon | 3 |
|
Transverse colon | 5 |
| Sigmoid
colon | 15 |
|
Rectum | 29 |
| Histological
grade |
|
| Well
differentiated | 66 |
| Poorly
differentiated | 1 |
| Mucinous colloid
type |
|
| No | 53 |
| Yes | 14 |
Immunohistochemical (IHC)
staining
IHC staining was carried out according to the
manufacturer's instructions (IHC assay kit; ProteinTech Group,
Inc.). Briefly, formalin-fixed and paraffin-embedded tissue
sections were deparaffinized in xylene and hydrated with decreasing
concentrations of ethanol (100, 95, 80 and 75%). The slices were
then soaked in 10% BSA to inhibit endogenous peroxidase activity
and incubated with the anti-IL-17F rabbit polyclonal antibody
(dilution 1:100; AB_354739, R&D Systems, Inc.) at 4°C
overnight. A horseradish peroxidase (HRP)-conjugated rabbit
secondary antibody (dilution 1:4,000; ProteinTech Group, Inc.) was
added for 60 min at room temperature; then, 3,3′-diaminobenzidine
development (DAB Substrate Chromogen System; Dako; Agilent
Technologies, Inc.) and hematoxylin staining were performed
according to standard protocols. Slides were fixed and images were
obtained with an Olympus IX71 inverted microscope and a DP2-BSW
Olympus image acquisition software system. IHC staining was scored
based on the proportion of cell staining and the staining
intensity. The percentage of positive cells was divided into 5
grades (percentage scores): <10% (0), 10–25% (1), >25-50% (2), >50-75% (3) and >75% (4). The intensity of staining was divided
into 4 grades (intensity scores): No staining, 0; weak staining, 1;
moderate staining, 2; strong staining, 3. IL-17F staining
positivity was determined by the following formula: Overall
score=percentage score × intensity score. An overall score ≤3 was
defined as negative and >3 was defined as positive. IL-17F
staining was scored by two independent researchers who were blinded
to the clinical characteristics of the patients.
Drug treatment and experimental
design
Recombinant human IL-17F protein (rhIL-17F) and the
anti-IL-17F antibody were purchased from R&D Systems, Inc.
(1335-IL-025, MAB13352). The working concentration of rhIL-17F and
anti-IL-17F antibody were 100 and 500 nM, respectively. The
experiment was divided into 3 groups: Negative control (NC) group,
rhIL-17F group and anti-IL-17F group.
Cell line and cell culture
The human CRC cell line HCT-116 (CVCL_0291) was
purchased from the University of Colorado Cancer Center Cell Bank
and was cultured in RPMI-1640 medium supplemented with 10% FBS
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C in a 5%
CO2 atmosphere. Cells were digested and passaged when
cell fusion reached 80%.
Protein extraction and western blot
analysis
Total protein was extracted using RIPA lysis buffer
with 1% phenylmethanesulfonyl fluoride. Then, equal amounts (20 µg)
of protein, as determined with a bicinchoninic acid protein assay
kit (Thermo Fisher Scientific, Inc.) were separated by 10%
SDS-PAGE. The proteins were then transferred to PVDF membranes
(0.45-mm; Beijing Solarbio Science & Technology Co., Ltd.). The
membranes were blocked with 5% non-fat milk for 1 h at room
temperature and then incubated with primary antibodies at 4°C for
12 h. The following antibodies were employed: Anti-E-cadherin
(AB_10697811), anti-Twist1 (AB_10598006), anti-vimentin
(AB_2273020), anti-Snail1 (13099-1) (dilution 1:1,000; ProteinTech
Group, Inc.) and anti-IL-17F (dilution 1:1,000; R&D Systems,
Inc.) rabbit polyclonal antibodies. Anti-GAPDH rabbit polyclonal
antibodies (dilution 1:4,000; AB_2107436, ProteinTech Group, Inc.)
were used as loading controls for normalization. The secondary
antibodies were anti-rabbit antibodies conjugated to HRP (dilution
1:4,000; AB_2722564, ProteinTech Group, Inc.). The antibodies were
used at 1:4,000 dilution and were incubated for ~1 h at room
temperature. The bands were visualized with ECL reagents (Thermo
Fisher Scientific, Inc.) and developed by Omega Lum™ G (Aplegen
Life Science).
Immunofluorescence (IF)
Coverslips were placed horizontally on the bottom of
a 6-well plate upon cleaning, disinfection and 24-h ultraviolet
irradiation. Then, the cells were seeded on the coverslips at a
density of 1×106 cells per well and cultured in an
incubator. The coverslips were rinsed with PBS for 5 min 3 times
and fixed with 4% paraformaldehyde for 15 min, followed by
permeabilization of the cells with 0.3% Triton X-100 for additional
10 min. Next, the coverslips were rinsed with PBS again for 5 min 3
times and blocked by incubating the cells in 5% BSA for 60 min.
Then, the cells were stained with anti-E-cadherin rabbit polyclonal
antibody (dilution 1:100; ProteinTech Group, Inc.), followed by a
12-h incubation period at 4°C. After washing the unbound antibody
with PBS for 5 min 3 times, the cells were incubated with an Alexa
488-conjugated goat anti-rabbit immunoglobulin G secondary antibody
(1:1,000; Cell Signaling Technology, Inc.) for 1 h at 4°C in the
darkness. Finally, DAPI was used as a counterstain to label the
nuclei. The stained cells were then acquired and photographed with
a fluorescence microscope.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Complementary DNA was
obtained from total RNA using PrimeScript™ RT Reagent kit (Takara
Bio, Inc.). The expression of mRNA was assessed by RT-qPCR, which
was carried out in triplicate using a SYBR Premix Ex Taq™ kit
(Takara Bio, Inc.) and an ABI 7900HT Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The primers used are
shown in Table II. The comparative
quantification cycle threshold values (2−ΔΔCq) were
employed to analyze the results (6).
| Table II.Primers for RT-qPCR. |
Table II.
Primers for RT-qPCR.
| Gene | Forward primer | Reverse primer |
|---|
| IL-17F |
GCTGTCGATATTGGGGCTTG |
GGAAACGCGCTGGTTTTCAT |
| Actin |
GGGACCTGACTGACTACCTC |
TCATACTCCTGCTTGCTGAT |
| E-cadherin |
AGTCACTGACACCAACGATAAT |
ATCGTTGTTCACTGGATTTGTG |
| Vimentin |
AGTCCACTGAGTACCGGAGAC |
CATTTCACGCATCTGGCGTTC |
| Snail1 |
AAGGATCTCCAGGCTCGAAAG |
GCTTCGGATGTGCATCTTGA |
| Twist1 |
GTACATCGACTTCCTCTACCAG |
CATCCTCCAGACCGAGAAG |
Cell proliferation assay
For this assay, 5×103 cells were seeded
into 96-well plates and incubated for the following times: 0, 24,
48 and 72 h. Next, 10 µl Cell Counting Kit-8 (CCK-8; Dojindo
Molecular Technologies, Inc.) solution was added to each well of
the plate, and incubation was continued for 2 h. Finally, the
absorbance of each well at a 450-nm wavelength was measured.
Colony formation assay
For this assay, 500 cells were seeded into 6-well
plates and incubated at 37°C. Colony size was observed daily under
a microscope until the number of cells in the majority of colonies
was >50. Then, the medium was removed and the cells were stained
with 0.2% crystal violet for 30 min. The cells were washed 3 times
with PBS, and then photographed and the colonies were counted. The
percentage of colony formation was calculated using the following
equation: Percentage of colony formation (%) = colony number/500 ×
100.
Cell migration and invasion
assays
Cell migration and invasion capacities were analyzed
with Transwell plates (24-well insert, 8-µm pore size; BD
Biosciences). The filters (Corning Inc.) were coated with (invasion
assay) or without (migration assay) 55 µl Matrigel (1:8 dilution;
BD Biosciences). For the migration assays, 5×104 cells
were suspended in 200 µl serum-free medium and seeded into the
Matrigel-uncoated upper chambers. Then, 600 µl medium containing
10% FBS was added to the lower chamber as a chemoattractant. After
incubation at 37°C for 24 h, the membranes were fixed with 4%
formaldehyde for 30 min and stained with 0.1% crystal violet at
room temperature for 30 min. For invasion assays, 1×105
cells suspended in 200 µl serum-free medium were seeded into the
Matrigel-coated upper chambers. The rest of the protocol was
identical to that described above. The cells were counted and
photographed under an inverted microscope (×400) over 5 different
fields for each triplicate filter.
Wound healing assay
For this assay, 5×105 cells were seeded
into 6-well plates and cultured at 37°C for 24 h. A 200-µl sterile
micropipette tip was used to scratch the confluent monolayers in a
straight line when cells were 80–90% confluent. Then, floating
cells were washed with PBS 3 times, and the cell culture was
continued to upon changing the complete medium to serum-free
medium. Images of the same wound position were captured at 0 and 48
h under an inverted microscope (×200). The migration results were
evaluated by ImageJ software (National Institutes of Health,
Bethesda, MD, USA).
Statistical analysis
SPSS software (version 19.0; IBM Corp.) was used for
statistical analysis of all the experimental data. GraphPad Prism
(version 7; GraphPad Software, Inc.) was used to determine the
statistical results. All data are expressed as the mean ± standard
deviation. Statistical analysis of the data derived from 2 groups
was performed with the Student's t-test. Comparisons of multiple
groups were performed by one-way analysis of variance (ANOVA)
followed by Least Significant Difference test. For survival
analysis, PROGgeneV2 (http://watson.compbio.iupui.edu/chirayu/proggene)
website tools was used to analyze the overall survival (OS) and
relapse-free survival (RFS) of the patients with CRC based on Gene
Expression Omnibus (GEO) databases (accession nos. GSE14333,
GSE17536, GSE16125 and GSE29621) (7–10).
P<0.05 was considered to indicate a statistically significant
difference.
Results
IL-17F is upregulated in CRC tumor
tissues
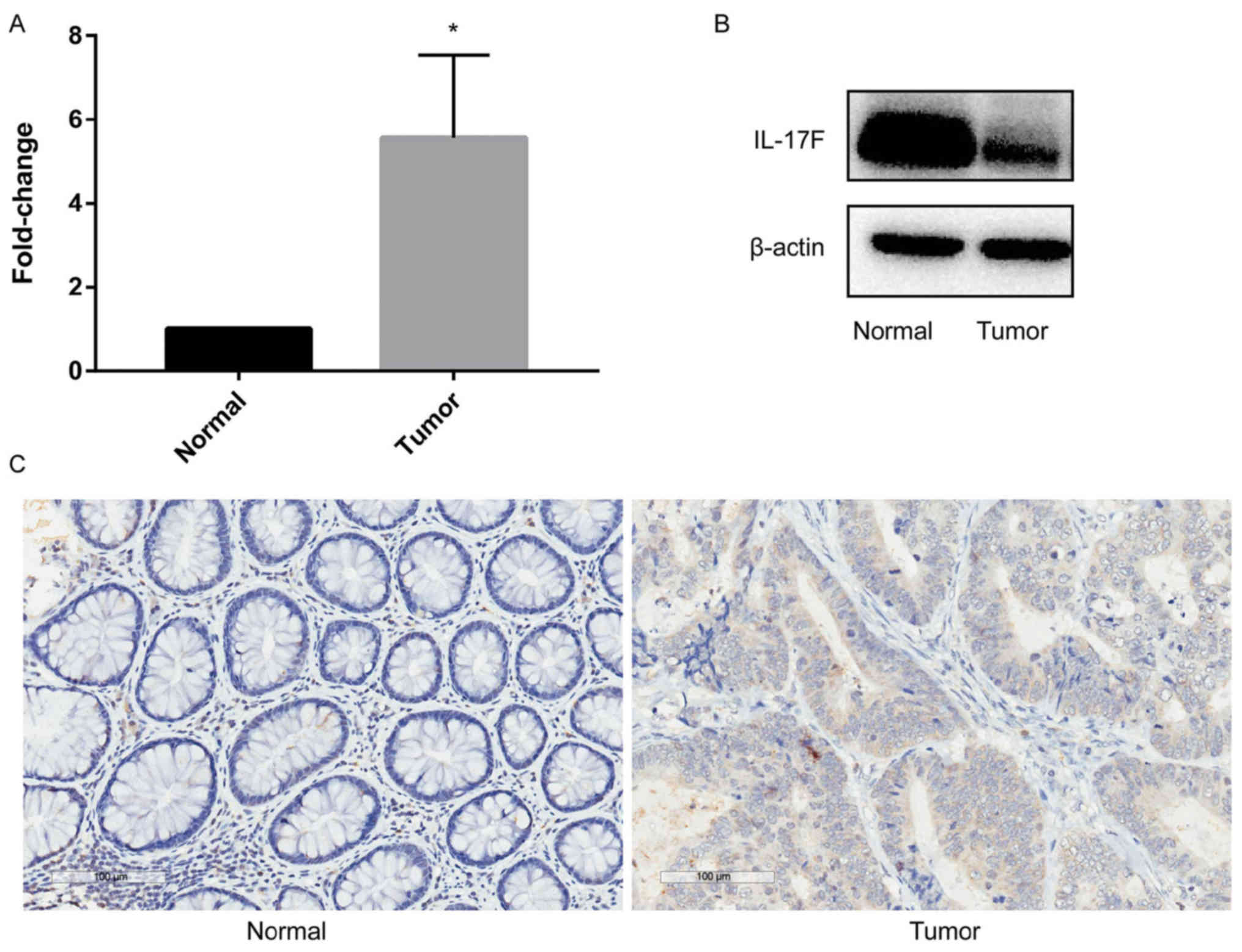
In order to analyze the function of IL-17F in CRC,
RT-qPCR was performed to detect IL-17F (Fig. 1A). Significant overexpression of
IL-17F in CRC tumor tissues compared with that noted in the paired
non-tumor mucosa was observed. Western blotting (Fig. 1B) and IHC (Fig. 1C) were used to detect the protein
expression of IL-17F, and the same trend was observed. Thus,
upregulation of IL-17F was observed in tumor tissues at both the
mRNA and protein levels.
IL-17F influences the overall survival
(OS) and relapse-free survival (RFS) of patients with CRC
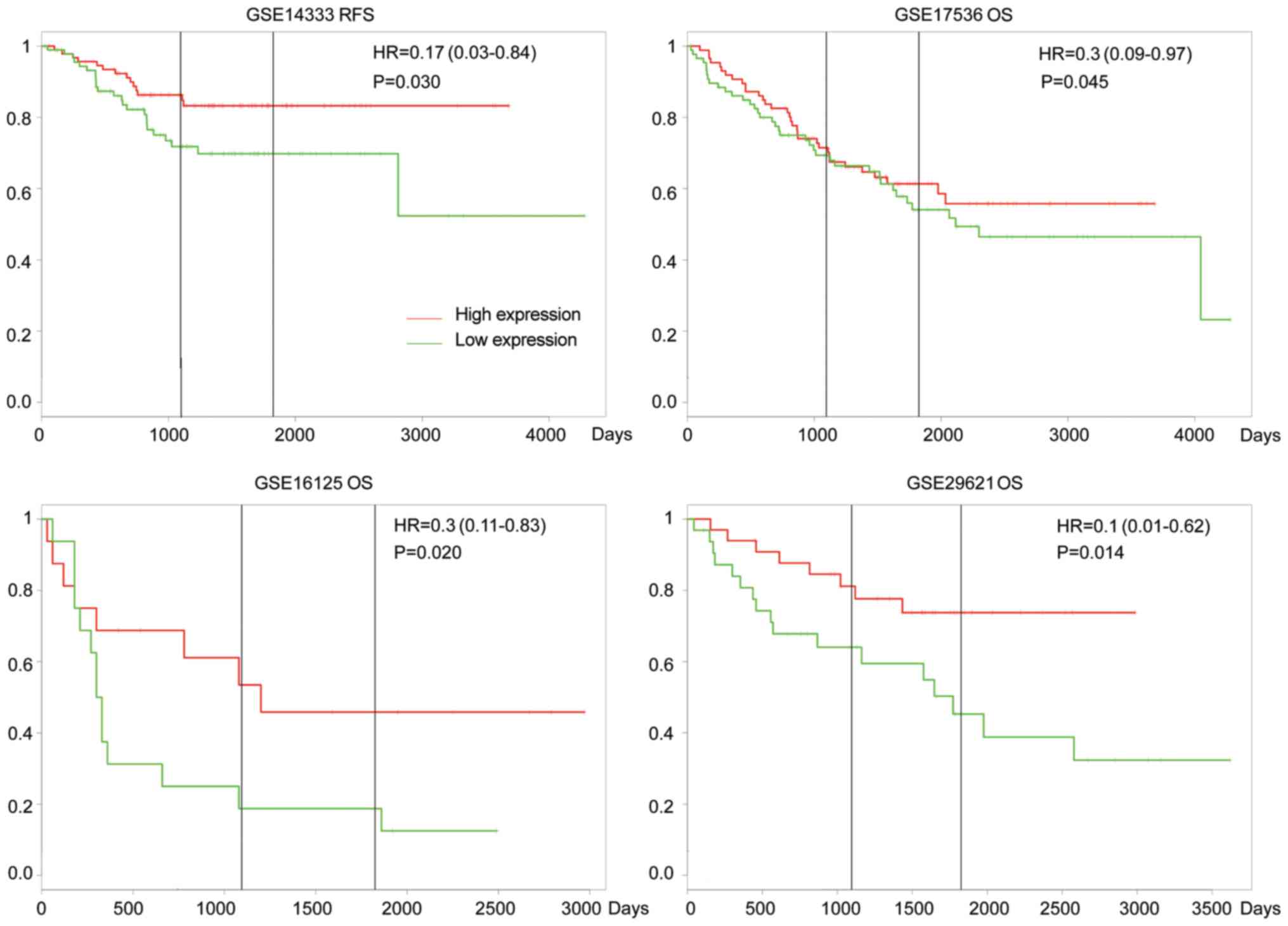
To explore the clinical significance of the
upregulation of IL-17F in CRC, the online tool PROGgeneV2
(http://watson.compbio.iupui.edu/chirayu/proggene)
was used to analyze the OS and RFS of patients with CRC based on
Gene Expression Omnibus (GEO) databases (6) (Fig. 2). In
GSE14333, overexpression of IL-17F was associated with worse RFS
[hazard ratio (HR), 0.17, P=0.03]; in GSE17536, GSE16125 and
GSE29621, overexpression of IL-17F was associated with worse OS
(HR, 0.3, P=0.045; HR, 0.3, P=0.02; and HR, 0.1, P=0.014,
respectively). The data indicated that IL-17F was a prognostic
predictor of OS and RFS in patients with CRC.
IL-17F promotes CRC invasion and
migration in vitro
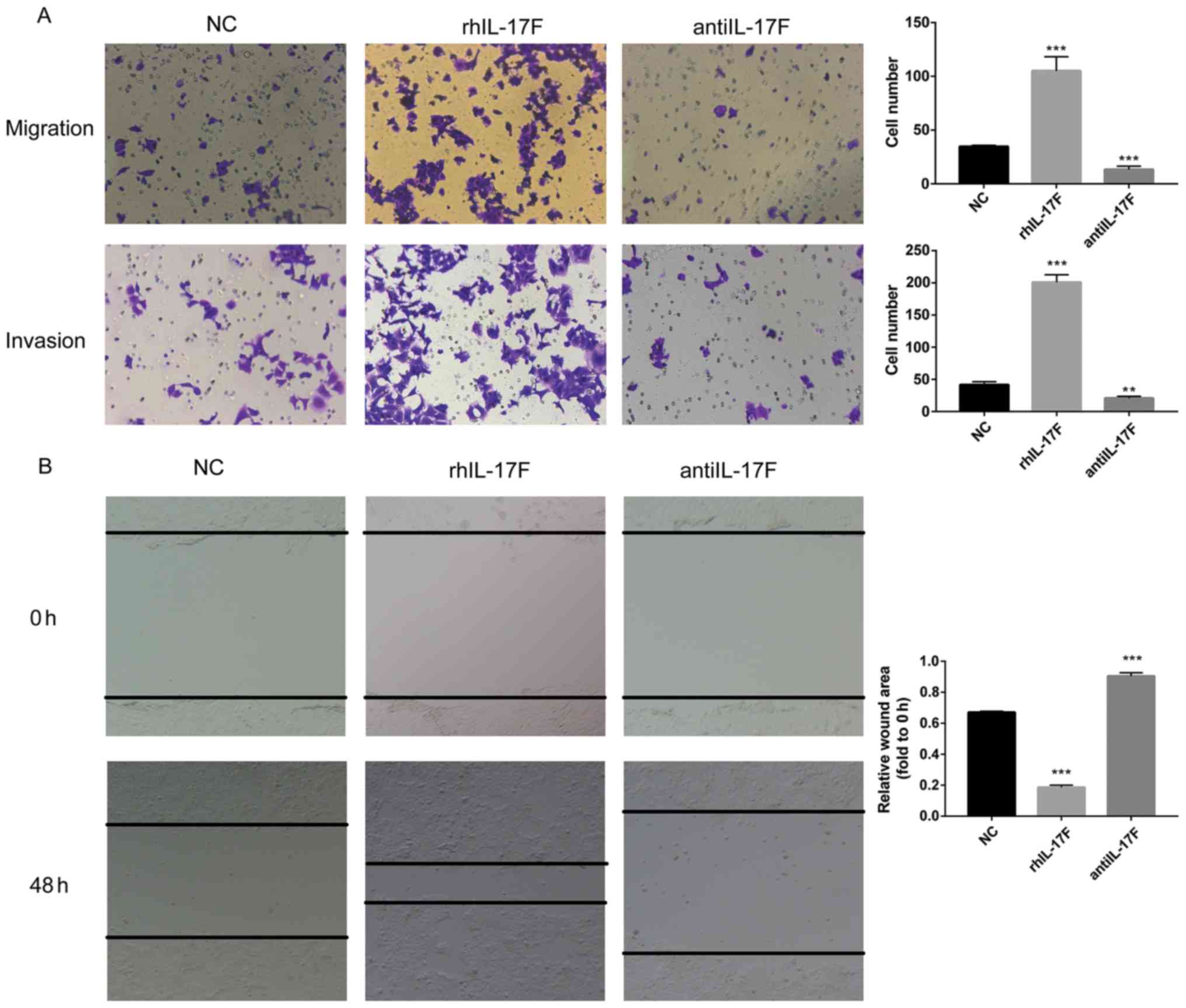
Transwell assays were used to evaluate the migration
and invasion capabilities of HCT-116 cells (Fig. 3A). There were more migrated and
invasive cells in the rhIL-17F group than these numbers in the NC
group in the migration and invasion assays, while the group treated
with anti-IL-17F antibody exhibited fewer cells compared with the
NC group. In the wound healing assay (Fig. 3B), the rhIL-17F group exhibited a
smaller wound area after 48 h, whereas the anti-IL-17F group
exhibited a larger area. Collectively, these results indicated that
IL-17F promotes HCT-116 cell invasion and migration.
Anti-IL-17F antibody inhibits the
colony formation of CRC cells
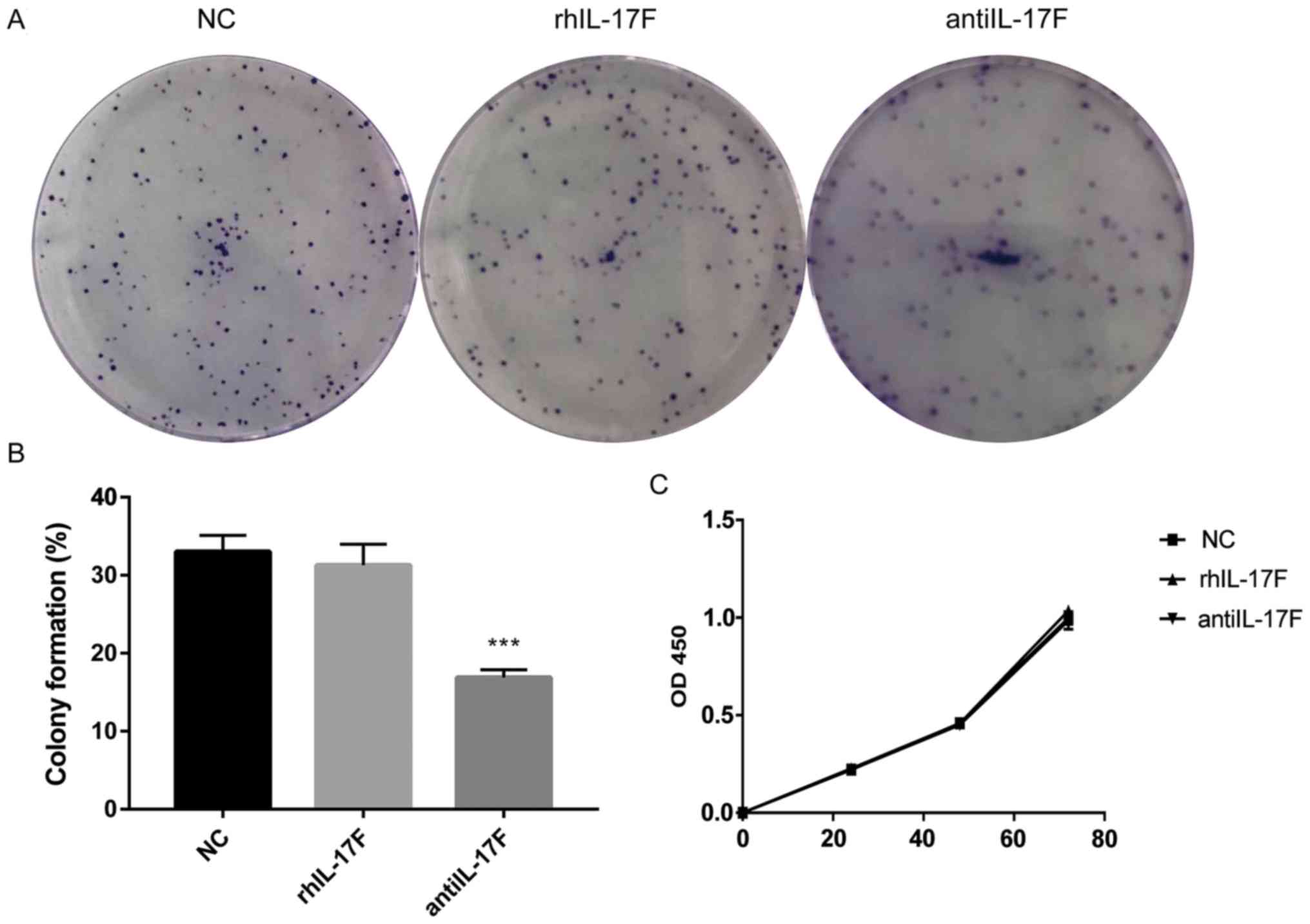
The colony formation assay was performed to assess
the influence of IL-17F on HCT-116 colony formation ability
(Fig. 4A and B). There was no
difference in the percentage of colony formation between the NC
group and the rhIL-17F group, whereas the anti-IL-17F group
exhibited a significantly inhibited colony formation ability
compared with the NC group. In addition, CCK-8 assay was further
performed to detect cell proliferation (Fig. 4C), but no difference was identified
between each group. These results indicated that rhIL-17F had no
effect on the colony formation ability or proliferation of HCT-116
cells.
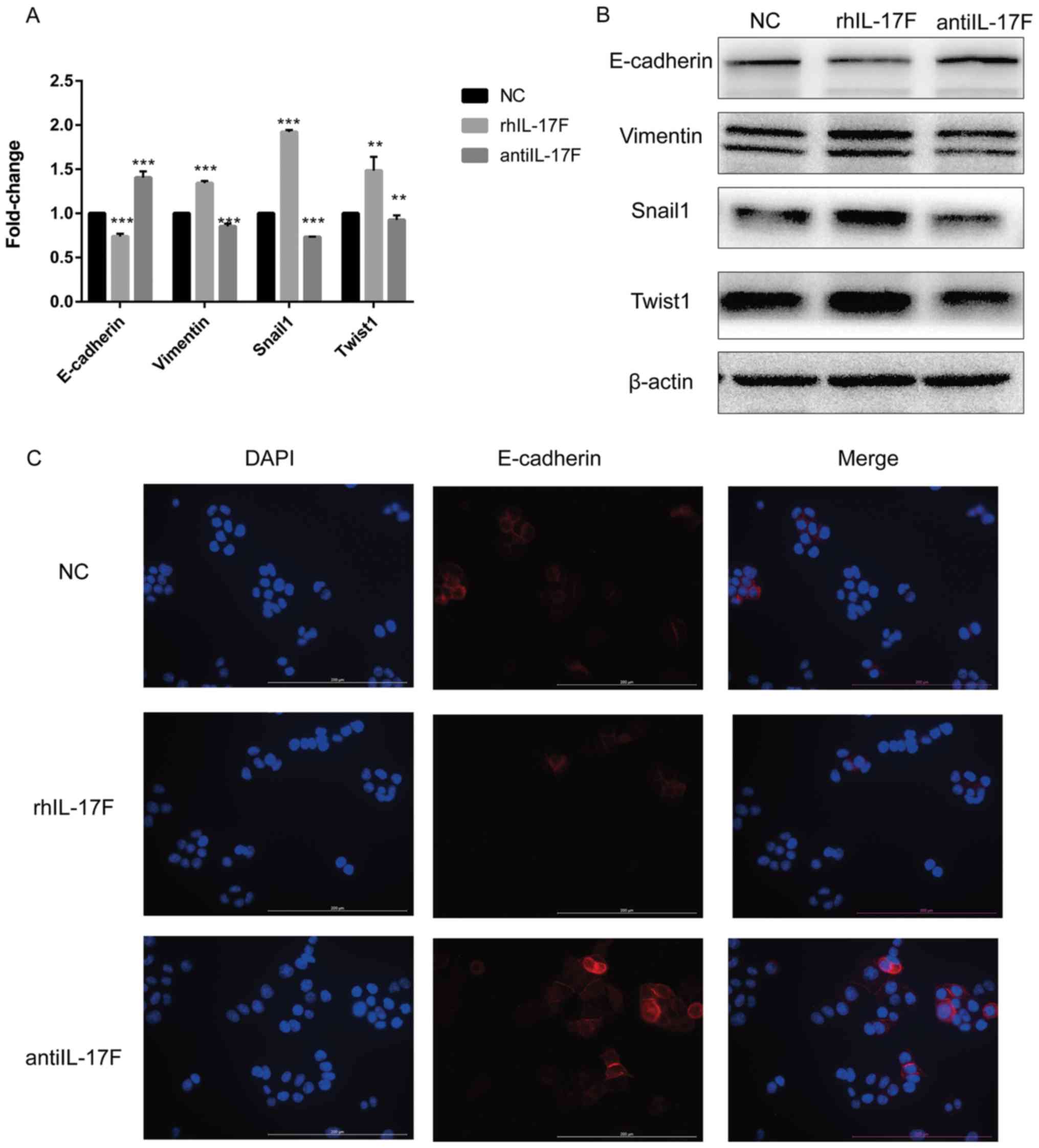
IL-17F induces EMT in CRC cells
RT-qPCR was performed to detect the mRNA expression
of several main EMT-associated markers (Fig. 5A). E-cadherin was significantly
decreased in the rhIL-17F group compared with the NC group, whereas
vimentin, Snail1 and Twist1 were significantly upregulated.
Notably, the anti-IL-17F group exhibited the opposite changes.
Furthermore, western blotting was performed to detect the protein
expression of EMT-associated markers (Fig. 5B). The changing trend was in agreement
with the results of RT-qPCR. Next, the location and expression of
E-cadherin were assessed by IF (Fig.
5C). E-cadherin was located in the cell membrane and cytoplasm,
and the changing trend in expression was in agreement with the
results of western blotting and RT-qPCR.
Discussion
IL-17F is member of the IL-17 family. Previous
research concerning IL-17 focused on inflammation and immunity
(11). Inflammation is closely
associated with cancer, as recent studies have demonstrated the
influence of IL-17 on different tumors. In non-small cell lung
cancer (NSCLC), IL-17 was found to increase the net angiogenic
activity and in vivo growth of NSCLC by promoting
CXCR-2-dependent angiogenesis (12).
In early-stage breast cancer (13),
IL-17BR was found to serve as a prognostic factor. However, the
number of studies which have focused on the association between
IL-17F and cancer, particularly colorectal cancer (CRC), is
limited.
The present study observed a significant
upregulation of IL-17F in tumor tissue compared with that noted in
paired non-tumor mucosa. Furthermore, overexpression of IL-17F was
associated with worse overall survival (OS) and relapse-free
survival (RFS) according to GEO databases. IL is a secretory
protein. Therefore, rhIL-17F was used to study its effect in
vitro. rhIL-17F obviously promoted HCT-116 cell invasion and
migration in vitro according to the results of Transwell and
wound healing assays.
It is known that epithelial-mesenchymal transition
(EMT) is a key mechanism associated with cancer invasion. EMT
refers to the transformation of epithelial cells into mesenchymal
cells under specific physiological and pathological conditions
(14). The concept of EMT was first
proposed in the field of embryonic development. Since then,
numerous studies on EMT have been carried out, involving different
life phenomena and pathological processes (15). During EMT, intercellular junctions and
cell polarity disappear, epithelial markers are downregulated,
mesenchymal phenotypes and associated markers are gradually
upregulated, and the biological behavior of cells also changes,
which is characterized by enhanced migration and invasion abilities
(16). This process is accompanied by
decreased E-cadherin expression and concurrent increases in the
expression of vimentin, N-cadherin and the transcription regulators
Twist and Snail. These key EMT-associated markers were further
detected in our study. rhIL-17F significantly decreased the
expression of E-cadherin and increased the expression of vimentin,
Snail1, Twist1; whereas the anti-IL-17F antibody produced the
opposite effect. This result indicates that IL-17F promotes HCT-116
cell invasion and migration by inducing EMT. There are some
shortcomings in our study, as we only researched the effect of
IL-17F on EMT in vivo. In fact, IL-17F polymorphism is also
closely related to the occurrence and development of various
tumors, which will be our next research focus.
Taken together, the present study demonstrated that
IL-17F was upregulated in CRC tumor tissues and was associated
worse OS and RFS of CRC patients. In vitro, IL-17F promoted
CRC cell invasion and migration by inducing EMT.
Acknowledgements
Not applicable.
Funding
The present study was funded by Puxiu Medical
Talents Training Program of Pudong Hospital (grant no.
PX201702).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC and ZY performed the experiments and wrote the
manuscript. DW contributed to the statistical analysis of the data
and collected tissues. YQ and ZM conceived and designed the study.
All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All procedures involving human participants were
performed in accordance with the Ethics Committee of Shanghai
Pudong Hospital and with the 1964 Declaration of Helsinki and its
later amendments or comparable ethical standards. All patients
provided their written informed consent. The study protocol was
approved by the Pudong Hospital Committee on human research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Long AG, Lundsmith ET and Hamilton KE:
Inflammation and colorectal cancer. Curr Colorectal Cancer Rep.
13:341–351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song J, Feng L, Zhong R, Xia Z, Zhang L,
Cui L, Yan H, Jia X and Zhang Z: Icariside II inhibits the EMT of
NSCLC cells in inflammatory microenvironment via down-regulation of
Akt/NF-κB signaling pathway. Mol Carcinog. 56:36–48. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Taniguchi K and Karin M: IL-6 and related
cytokines as the critical lynchpins between inflammation and
cancer. Semin Immunol. 26:54–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Conti HR, Shen F, Nayyar N, Stocum E, Sun
JN, Lindemann MJ, Ho AW, Hai JH, Yu JJ, Jung WJ, et al: Th17/IL-17
receptor signaling and not Th1 cells are essential for mucosal host
defense against oral candidiasis. Cytokine. 206:299–311. 2009.
|
|
5
|
Hymowitz SG, Filvaroff EH, Yin JP, Lee J,
Cai L, Risser P, Maruoka M, Mao W, Foster J, Kelley RF, et al:
IL-17s adopt a cystine knot fold: Structure and activity of a novel
cytokine, IL-17F, and implications for receptor binding. EMBO J.
20:5332–5341. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schmittgen TD: Real-time quantitative PCR.
Methods. 25:383–385. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jorissen RN, Gibbs P, Christie M, Prakash
S, Lipton L, Desai J, Kerr D, Aaltonen LA, Arango D, Kruhoffer M,
et al: Metastasis-associated gene expression changes predict poor
outcomes in patients with dukes stage B and C colorectal cancer.
Clin Cancer Res. 15:7642–7651. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Smith JJ, Deane NG, Fei WU, Merchant NB,
Zhang B, Jiang A, Pengcheng LU, Johnson JC, Schmidt C, Bailey CE,
et al: Experimentally derived metastasis gene expression profile
predicts recurrence and death in patients with colon cancer.
Gastroenterology. 138:958–968. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Reid JF, Gariboldi M, Sokolova V,
Capobianco P, Lampis A, Perrone F, Signoroni S, Costa A, Leo E,
Pilotti S and Pierotti MA: Integrative approach for prioritizing
cancer genes in sporadic colon cancer. Genes Chromosomes Cancer.
48:953–962. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen DT, Hernandez JM, Shibata D, McCarthy
SM, Humphries LA, Clark W, Elahi A, Gruidl M, Coppola D and Yeatman
T: Complementary strand microRNAs mediate acquisition of metastatic
potential in colonic adenocarcinoma. J Gastrointest Surg.
16:905–913. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
van der Waart AB, van der Velden WJ,
Blijlevens NM and Dolstra H: Targeting the IL17 pathway for the
prevention of graft-versus-host disease. Biol Blood Marrow
Transplant. 20:752–759. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Numasaki M, Watanabe M, Suzuki T,
Takahashi H, Nakamura A, McAllister F, Hishinuma T, Goto J, Lotze
MT, Kolls JK and Sasaki H: IL-17 enhances the net angiogenic
activity and in vivo growth of human non-small cell lung cancer in
SCID mice through promoting CXCR-2-dependent angiogenesis. J
Immunol. 175:6177–6189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma XJ, Hilsenbeck SG, Wang W, Ding L,
Sgroi DC, Bender RA, Osborne CK, Allred DC and Erlander MG: The
HOXB13:IL17BR expression index is a prognostic factor in
early-stage breast cancer. J Clin Oncol. 24:4611–4619. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nieto MA: Epithelial-mesenchymal
transitions in development and disease: Old views and new
perspectives. Int J Dev Biol. 53:1541–1547. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lim J and Thiery JP:
Epithelial-mesenchymal transitions: Insights from development.
Development. 139:3471–3486. 2012. View Article : Google Scholar : PubMed/NCBI
|