Introduction
Nemo-like kinase (NLK) is a conserved
serine/threonine kinase and is involved in a variety of biological
responses and is regulated by a variety of different transcription
factors (1,2). Decreased expression of NLK was
found in various types of human cancers. In human melanoma,
decreased expression of NLK is associated with poor
prognosis and increased vascularity, and metastasis (3). Decreased expression of NLK in
human ovarian carcinomas is associated with poor patient survival
(4,5). In addition, NLK is
significantly downregulated in breast cancer tissues and non-small
cell lung cancer (NSCLC) compared with that in corresponding normal
tissues (6,7). NLK has also been identified as
a tumor-suppressor gene in glioblastoma using an in vivo
RNAi screen (8).
Epithelial-mesenchymal transition (EMT) is a
biological process, involving the functional transition of
polarized epithelial cells into mesenchymal cells, and is involved
in cancer metastasis, migration, invasion, and progression
(9–11). E-cadherin is typically repressed
during EMT. Thus, E-cadherin is considered to be a suppressor of
migration and invasion of malignant epithelial cancers (12,13).
14-3-3ζ (also known as YWHAZ) is a member of the
evolutionally conserved regulatory family that mediates signal
transduction by binding to phosphoserine-containing proteins
(14). 14-3-3ζ was found to be
responsible for silencing of E-cadherin during EMT (15).
In the present study, overexpression of NLK
was found to inhibit the growth and migration of the NSCLC A549
cell line. Moreover, NLK interacts with 14-3-3ζ and prevents
its dimerization. In addition, NLK overexpression was able
to restore the expression of E-cadherin inhibited by 14-3-3ζ but
not with the fused dimer of 14-3-3ζ. The results from the present
study demonstrate that NLK is a negative regulator of
14-3-3ζ and plays a tumor-suppressive role by inhibiting the
migration of cancer cells.
Materials and methods
Cell culture
The human A549, H358, H322, H23, H1437, H1650,
Calu-3 and H441 lung cancer epithelial cell lines and human 293T
cells were obtained from the Chinese Academy of Sciences cell bank,
and were cultured in DMEM (cat no. 10-013-CV; Corning, Inc.)
containing 10% FBS (cat no. 35-010-CV; Corning, Inc.), 100 U/ml
penicillin and 100 mg/ml streptomycin at 37°C in a 95% humidified
incubator with 5% CO2.
Antibodies
Rabbit antibodies to 14-3-3ζ (cat. no. ab155037;
dilution 1:1,000) and β-actin (cat. no. ab227387; dilution 1:2,000)
were obtained from Abcam. Rabbit antibodies to NLK (cat. no.
94350; dilution 1:500), E-cadherin (cat. no. 3195; dilution 1:500)
and HA tag (cat. no. 3724; dilution 1:2,000) were purchased from
Cell Signaling Technology, Inc. Mouse antibodies to FLAG-M2 tag
(cat. no. F1804; dilution 1:2,000), FLAG-M2 affinity gel (cat. no.
A2220) and HA affinity gel (cat. no. A2095) were purchased from
Sigma-Aldrich (Merck KGaA). Horseradish peroxidase (HRP)-linked
anti-mouse IgG (cat. no. 7076; dilution 1:5,000) and anti-rabbit
IgG (cat. no. 7074; dilution 1:5,000) were purchased from Cell
Signaling Technology, Inc. HRP-linked light chain specific goat
anti-mouse IgG (cat. no. 115-005-174) and mouse anti-rabbit IgG
(cat. no. 211-002-171) for immunoprecipitation (IP) were purchased
from Jackson ImmunoResearch Laboratories, Inc.
Lentivirus packaging and
transduction
The lentivirus pCDH vector containing NLK,
C-terminal FLAG tagged NLK, 14-3-3ζ, C-terminal FLAG/HA
tagged 14-3-3ζ, 14-3-3ζ-Fusion [by a 10-amino acid Glycine
(Gly)-rich linker] and FLAG tagged 14-3-3ζ-Fusion were purchased
from Genesent Biological Technology Co., Ltd. To prepare the
lentivirus, 293T cells were transfected with the aforementioned
lentiviral vectors and packaging plasmids using
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Virus-containing
medium was collected 48 h after transfection and filtered through
0.45 µM low protein-binding filters (Nalgene; Thermo Fisher
Scientific, Inc.). For transduction, cells were infected with
lentiviral vectors and selected with puromycin. Subsequently, the
expression levels of the target genes were detected using western
blot analysis.
Colony formation assay
For colony formation, A549 cells were seeded in
6-well plates at 500 cells per well, and cultured for 2 weeks. When
clones were visible, the colonies were fixed with 4%
paraformaldehyde for 30 min and stained with 0.1% crystal violet
(Sigma-Aldrich; Merck KGaA) for 5 min. Subsequently images of the
cells were obtained and the number of cells were counted under an
Olympus BX51 fluorescence microscope (magnification ×100; Olympus
Corp.).
Wound healing assay
Total 5×105 A549 cells infected with
different lentivirus were cultured using 6-well plates. After cells
had grown to produce a 90% confluent monolayer, a pipette tip (200
µl) was used to create a wound. The cells were washed twice with
PBS to remove non-adherent cells and cultured in serum-free medium
for 18 or 24 h. Subsequently, the images of wound healing were
obtained an Olympus BX51 fluorescence microscope (magnification
×100; Olympus Corp.) and the migration rate was calculated. The
migration rate was calculated using the following formula:
Migration rate=(Area of original wound - Area of actual wound)/Area
of original wound ×100%.
Sphere formation assay
Monolayer cultured A549 cells (1,000 cells per well)
infected with different lentivirus were suspended in 24-well
ultra-low attachment plates (Corning, Inc.) with serum-free
DMEM/F12 containing 20 ng/ml of epidermal growth factor (Thermo
Fisher Scientific, Inc.), 20 ng/ml of basic fibroblast growth
factor (Sigma-Aldrich; Merck KGaA) and 2% B27 supplement (Thermo
Fisher Scientific, Inc.). After 7 days of incubation, images of the
spheres were obtained under a phase-contrast Olympus BX51
microscope (magnification ×100; Olympus Corp.).
IP and liquid chromatography-mass
spectrometry (LC-MS)/MS analysis
For the IP assay, cells were harvested and lysed
with RIPA buffer [50 mM Tris-HCI (pH7.4), 150 mM NaCl, 1.0% NP-40,
and 0.25% sodium deoxycholate] with protease inhibitor and
phosphatase inhibitor cocktail (Roche Diagnostics, GmbH). Following
brief sonication, the lysates were centrifuged at 15,000 × g for 15
min at 4°C and the supernatants were subsequently incubated with
the indicated primary antibodies together with protein
A/G-sepharose beads (Santa Cruz Biotechnology, Inc.) or direct with
FLAG-M2 affinity gel at 4°C for 2 h. The beads were washed three
times before heating or adding FLAG peptides (Sigma-Aldrich; Merck
KGaA). Isolated protein, including NLK-FLAG and its
associated proteins were forwarded to Taiyuan Rosetta Stone Biotech
Co., Ltd. for LC-MS/MS analysis. The mass of the peptides was
identified using LTQ-XL mass spectrometer (Thermo Fisher
Scientific, Inc.).
SDS-PAGE and western blot
analysis
Total proteins were lysed with RIPA lysis buffer
(cat. no. 9806; Cell Signaling Technology, Inc.). For denaturing
conditions, the cell lysates were mixed with 5X SDS denaturing
sample buffer (cat. no. 39000; Thermo Fisher Scientific, Inc.), and
incubated at 95°C for 5 min. For non-denaturing conditions, the
cell lysates were mixed with 2X SDS non-denaturing sample buffer
(cat. no. LC2673; Thermo Fisher Scientific, Inc.) without
2-mercaptoethanol and incubated at room temperature for 60 min. The
protein was separated using 12% SDS-PAGE or NativePAGE™ (Thermo
Fisher Scientific, Inc.), and subsequently transferred onto a PVDF
membrane (EMD Millipore; Merck KGaA). The PVDF membranes were
blocked in 5% skimmed milk for 1 h at room temperature, and
incubated with aforementioned primary antibodies overnight at 4°C.
Following which, the membranes were washed with TBS-Tween-20 and
incubated with the aforementioned secondary antibodies for 1 h at
room temperature. Proteins were visualized using enhanced
chemiluminescence reagents (Roche Diagnostics GmbH). Densitometric
analysis was performed using ImageJ software (version 1.48;
National Institutes of Health).
Statistical analysis
GraphPad Prism 5 software (GraphPad Software, Inc.)
was used to analyze the data. Data are presented as the mean ±
standard deviation. All experiments were repeated 3 times.
Two-tailed Student's t-test was used when two independent groups
were compared, while one-way analysis of variance (ANOVA) followed
by Tukey's multiple comparison was used for multiple comparison
tests. A P<0.05 was considered to indicate a statistically
significant difference.
Results
NLK inhibits cell growth and
migration
To examine whether overexpression of NLK
affects cell growth, lentivirus transfection was used to upregulate
the gene expression of NLK in A549 cells. From the western
blot analysis, A549 cells showed stable overexpression of
NLK compared with that in the control cells transfected with
the vector only (Fig. 1A). Colony
formation assay (CFA) demonstrated that overexpression of
NLK significantly represses colony formation ability, and
overexpression of NLK significantly repressed sphere
formation capabilities (SFA) in the A549 cells (Fig. 1B). Wound healing assay was used to
determine whether overexpression of NLK affected the
migration ability of A549 cells, and it was found that
overexpression of NLK resulted in significantly decreased
migration of A549 cells compared with that in cells transfected
with the vector only (Fig. 1C). The
results suggest that NLK significantly inhibited cell growth
and migration capabilities.
NLK interacts with 14-3-3ζ
To further understand the mechanism of NLK in
the regulation of cell growth and migration, IP and MS analyses
were used to identify NLK binding proteins in A549 cells
stably expressing C-terminally FLAG-tagged NLK
(NLK-FLAG) created using lentivirus transfection. Cell
extracts were subjected to IP with FLAG agarose beads. The specific
bands of NLK-FLAG in isolated proteins were identified using
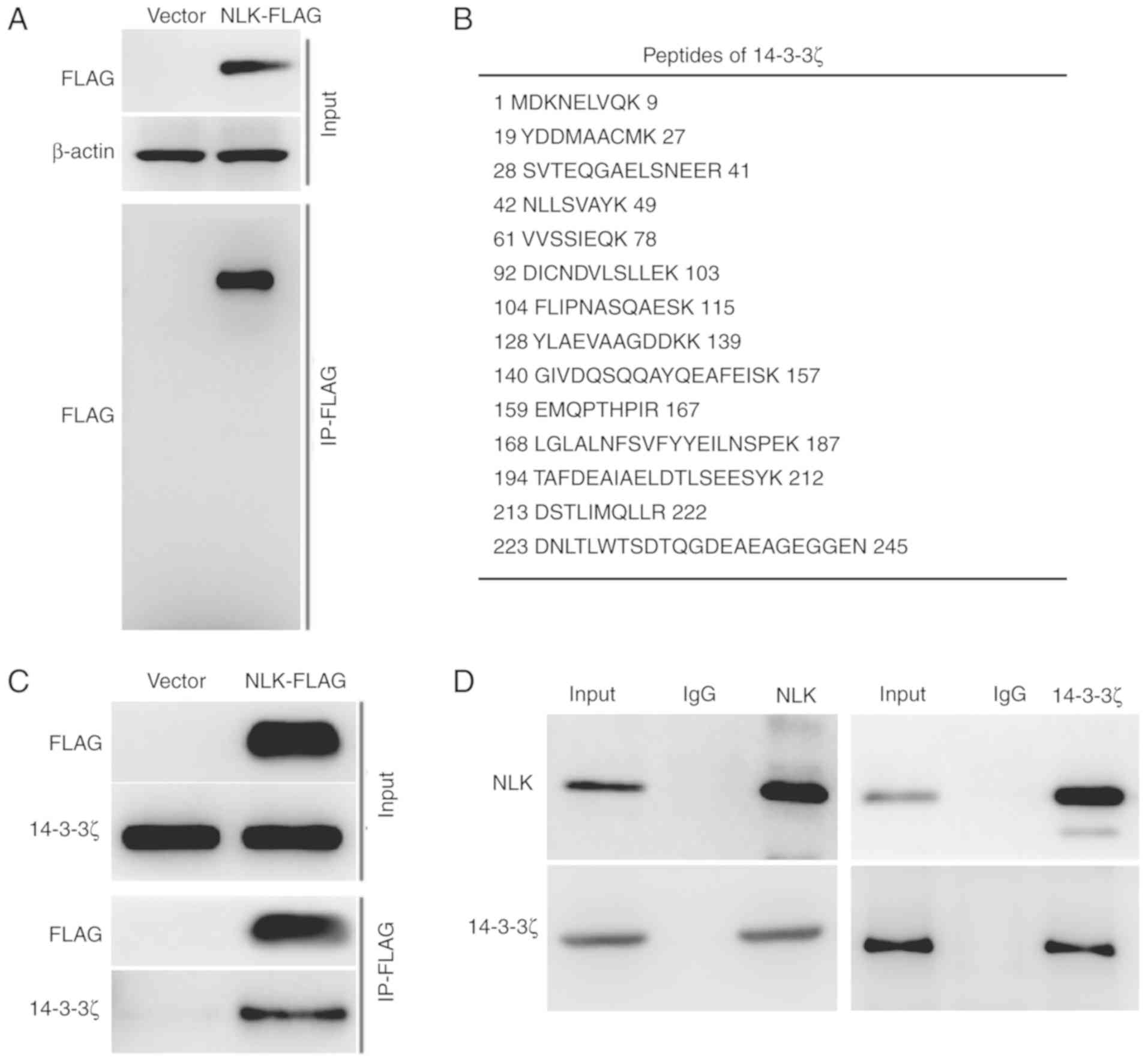
western blot analysis (Fig. 2A).
Subsequently, the isolated proteins, including NLK-FLAG and
its associated proteins were analyzed using LC-MS/MS to identify
NLK-interacting proteins. The tryptic peptides corresponding
to 14-3-3ζ protein with a high sequence coverage were identified
(Fig. 2B). The interaction between
NLK-FLAG and 14-3-3ζ was further verified using western blot
analysis with the precipitated proteins (Fig. 2C). Moreover, endogenous NLK
and 14-3-3ζ also showed an interaction in A549 cells (Fig. 2D). These data indicate that 14-3-3ζ
is a novel NLK-interacting protein.
NLK inhibits the dimerization of
14-3-3ζ and restores 14-3-3ζ-repressed E-cadherin expression
A previous study has shown that 14-3-3ζ is
responsible for silencing of E-cadherin during EMT (15). Lentivirus transfection was used to
upregulate the gene expression of 14-3-3ζ in A549 cells (Fig. 3A). It was subsequently confirmed
that overexpression of 14-3-3ζ induced a more spindle-like cell
shape and a scattered distribution, suggesting decreased epithelial
cell-to-cell contacts in the A549 cells (Fig. 3A). In addition, A549 cells stably
overexpressing 14-3-3ζ were transfected with the NLK
lentivirus to create NLK/14-3-3ζ double-overexpressing
cells. EMT morphological changes induced by 14-3-3ζ overexpression
were inhibited by NLK overexpression (Fig. 3A). Subsequently it was confirmed
that overexpression of 14-3-3ζ inhibited the expression of
E-cadherin (Fig. 3B). Furthermore,
NLK/14-3-3ζ double-overexpressing cells revealed restored
expression of E-cadherin compared with that in cells overexpressing
14-3-3ζ (Fig. 3B). It has been
reported that dimerization is essential for 14-3-3ζ stability and
the ability of 14-3-3ζ to bind to its target proteins (16,17).
Using non-denaturing conditions, which would preserve any disulfide
linkages during western blot analysis, an endogenous 14-3-3ζ dimer
band was found in A549 cells (Fig.
3C). Moreover, the 14-3-3ζ monomer band and 14-3-3ζ dimer band
were found to be increased in A549 cells stably overexpressing
14-3-3ζ. On the other hand, the 14-3-3ζ dimer band was inhibited in
NLK/14-3-3ζ double-overexpressing cells as compared with
that in 14-3-3ζ overexpressing cells (Fig. 3C).
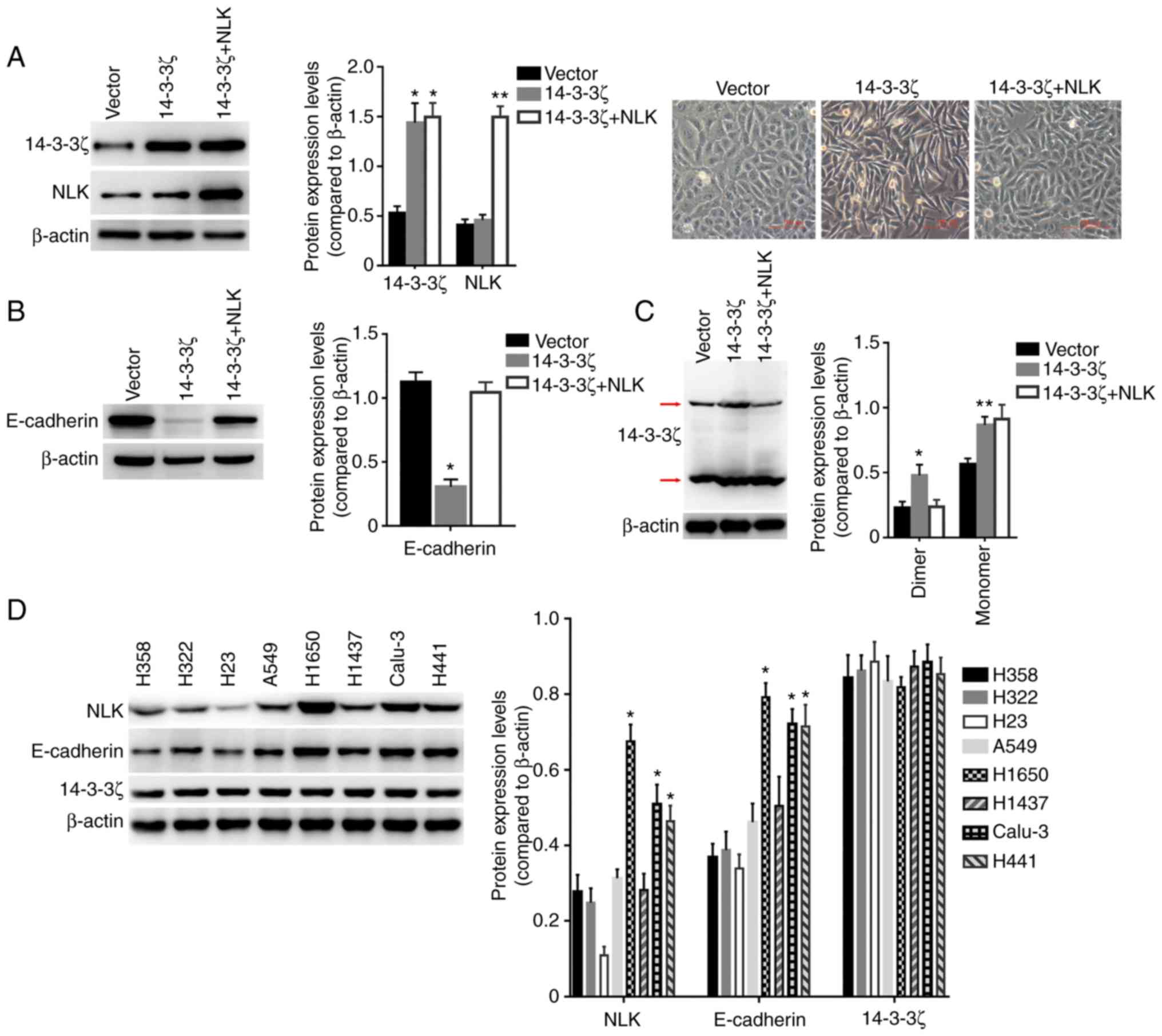 | Figure 3.NLK regulates
14-3-3ζ-repressed E-cadherin expression. (A) Left panels:
Expression of NLK, 14-3-3ζ and β-actin were detected by
western blot in A549 cells transfected with the vector lentivirus,
14-3-3ζ lentivirus or 14-3-3ζ plus NLK lentivirus, and
densitometric analysis of the relative expression levels of 14-3-3ζ
and NLK. *P<0.001 compared to the Vector group;
**P<0.001 compared to the other groups. Right images: The cells
exhibited different morphologies. (B) Expression of E-cadherin and
β-actin were detected by western blot analysis in A549 cells
transfected with the vector lentivirus, 14-3-3ζ lentivirus or
14-3-3ζ plus NLK lentivirus, and densitometric analysis of
the relative expression level of E-cadherin. *P<0.005 compared
to other groups. (C) Under non-denaturing condition, the expression
of 14-3-3ζ and β-actin were detected by western blot analysis in
A549 cells transfected with the vector lentivirus, 14-3-3ζ
lentivirus or 14-3-3ζ plus NLK lentivirus, and densitometric
analysis of the relative expression levels of dimer and monomer of
14-3-3ζ. *P<0.01 compared to other groups; **P<0.05 compared
to the Vector group. (D) Expression of NLK, E-cadherin,
14-3-3ζ and β-actin were detected by western blot analysis in 8
human lung cancer cell lines, and densitometric analysis of the
relative expression levels of NLK, E-cadherin and 14-3-3ζ.
*P<0.005 compared to the other groups. NLK, nemo-like
kinase. |
At the protein level, expression of E-cadherin was
evaluated in 8 human lung cancer cell lines using western blot
analysis. There were no differences in the expression levels of
14-3-3ζ in these cells, and the expression levels between
NLK and E-cadherin were highly correlated (Fig. 3D). These results suggest that
NLK may be involved in 14-3-3ζ-regulated E-cadherin
expression by the interaction with 14-3-3ζ. Using lentivirus
transfection, A549 cells stably sole-expressing or co-expressing
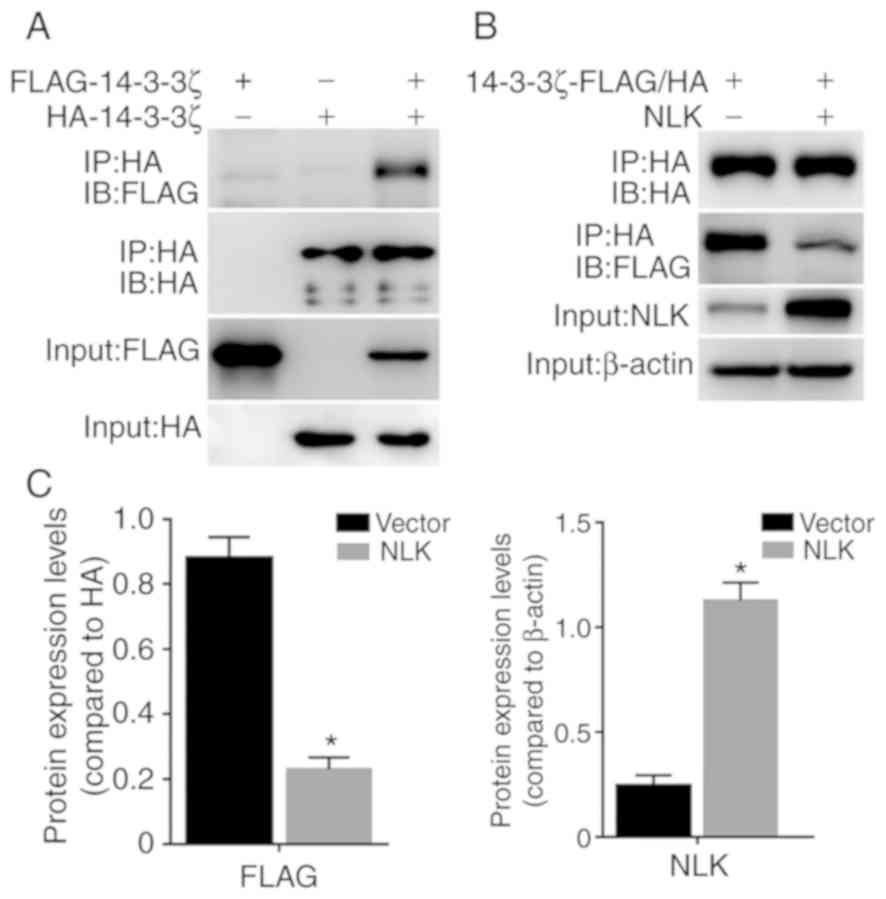
FLAG-14-3-3ζ and HA-14-3-3ζ were generated. As shown in Fig. 4A, FLAG-14-3-3ζ was
co-immunoprecipitated with HA-14-3-3ζ in FLAG-14-3-3ζ and
HA-14-3-3ζ co-expressing A549 cells. Notably, overexpression of
untagged NLK in FLAG-14-3-3ζ and HA-14-3-3ζ co-expressing
cells inhibited the interaction between FLAG-14-3-3ζ and HA-14-3-3ζ
(Fig. 4B and C). These results
suggest that NLK disturbs 14-3-3ζ-14-3-3ζ dimerization.
NLK regulation of E-cadherin
expression depends on the homodimer of 14-3-3ζ
Gly-rich linkers are flexible and naturally exist in
a number of proteins of their Gly residues (18–20).
Thus, Gly-rich linkers have been used to create fusion proteins
with independent sequences, including the connection with the
homodimer, ligand-receptor and co-activators (21–23).
By fusing two copies of the 14-3-3ζ gene via a 10-amino acid
Gly-rich linker, a non-dissociable dimer of 14-3-3ζ
(14-3-3ζ-Fusion) with lentiviral vector (Fig. 5A) was generated. As shown in
Fig. 5A, the expression of the
fused dimer of 14-3-3ζ and endogenous 14-3-3ζ were examined using
western blot analysis, under denaturing and non-denaturing
conditions.
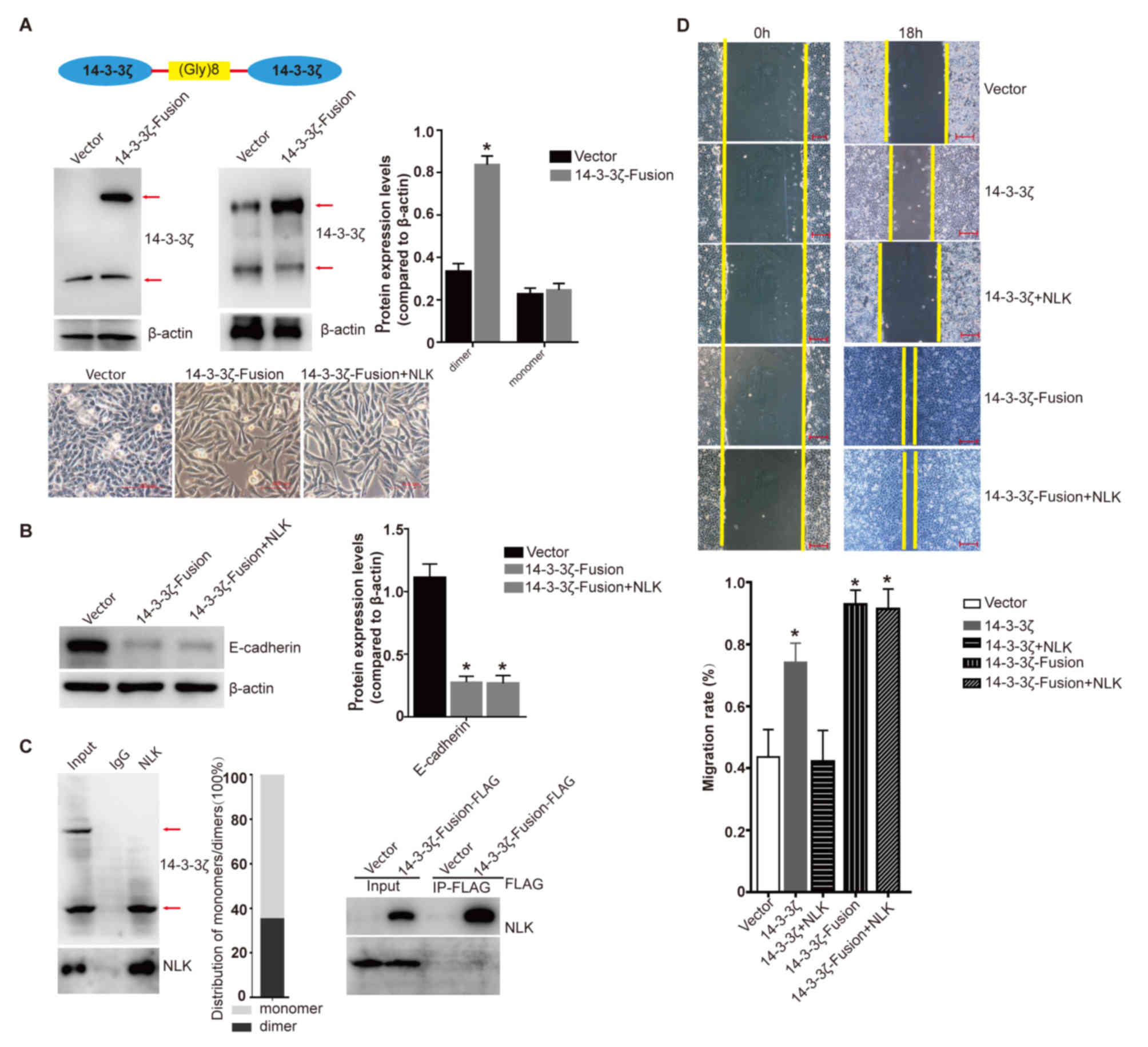 | Figure 5.The function of non-dissociable
homodimer of 14-3-3ζ. (A) Two copies of the 14-3-3ζ gene via a
10-amino acid Gly-rich linker to get a non-dissociable dimer of
14-3-3ζ (14-3-3ζ-Fusion) with lentiviral vector. Expression of
endogenous 14-3-3ζ and 14-3-3ζ-Fusion was detected by western blot
analysis in A549 cells transfected with the vector and
14-3-3ζ-Fusion lentivirus (left: Under denaturing condition, right:
Under non-denaturing condition). Densitometric analysis of the
relative expression level of dimer and monomer of 14-3-3ζ.
*P<0.005 compared to the Vector group. Lower images: The cells
exhibited different morphologies with the transfection of the
vector, 14-3-3ζ-Fusion or 14-3-3ζ-Fusion plus untagged NLK
lentivirus (scale bar, 100 µm). (B) Expression of E-cadherin and
β-actin was detected by western blot analysis in A549 cells
transfected with the vector lentivirus, 14-3-3ζ-Fusion or
14-3-3ζ-Fusion plus untagged NLK lentivirus. Densitometric
analysis of the relative expression level of E-cadherin.
*P<0.001 compared to the Vector group. (C) Left: A549 cells were
lysed and subjected to IP with NLK antibodies, controlled by
corresponding IgG. Immunoprecipitated protein were then analyzed by
western blot analysis under non-denaturing condition. Right:
Immunoprecipitated 14-3-3ζ-Fusion-FLAG-associated protein in A549
cells was subjected to western blot analysis. (D) Wound healing
analysis of A549 cells transfected with the vector, 14-3-3ζ,
14-3-3ζ-Fusion, 14-3-3ζ plus NLK or 14-3-3ζ-Fusion plus
NLK lentivirus. *P<0.005 compared to the Vector or
14-3-3ζ+NLK groups (scale bar, 200 µm). NLK,
nemo-like kinase; IP, immunoprecipitation. |
Moreover, spindle-like cell shape and a scattered
distribution were also found in cells stably expressing
14-3-3ζ-Fusion. The EMT morphological changes were not inhibited by
overexpression of NLK (Fig.
5A). Meanwhile, 14-3-3ζ-repressed E-cadherin expression could
not be restored by NLK overexpression (Fig. 5B).
To further determine whether NLK binds to the
monomer or dimer form of 14-3-3ζ, the endogenous interaction
between NLK and 14-3-3ζ under non-denaturing conditions was
investigated. As shown in Fig. 5C,
NLK was found to be associated with the monomer form but not
with the dimer form of 14-3-3ζ under non-denaturing conditions.
Moreover, exogenous FLAG-tagged 14-3-3ζ dimer fusion protein
(14-3-3ζ-Fusion-FLAG) could not bind to endogenous NLK in
14-3-3ζ-Fusion-FLAG expressing cells (Fig. 5C). In addition, wound healing assay
revealed that 14-3-3ζ overexpression enhanced the migration of A549
cells. NLK was able to inhibit the increased migration from
14-3-3ζ overexpression. On the other hand, 14-3-3ζ-Fusion
overexpression revealed a higher migration ability compared with
that in 14-3-3ζ-overexpressing cells and this effect was not
regulated by NLK (Fig. 5D).
Together, these results suggest that NLK regulation of
E-cadherin expression depends on the homodimer of 14-3-3ζ.
Discussion
EMT is involved in cancer metastasis, migration,
invasion, and progression. E-cadherin is essential for the
maintenance of intercellular adhesion as a key component of the
adherens junctions (24). Thus, the
loss of E-cadherin is considered to be a major hallmark of EMT and
has been reported in various types of cancers (25). 14-3-3ζ, a member of the
evolutionally conserved family, mediates signal transduction by
binding to phosphoserine-containing proteins. Dimerization is
essential for 14-3-3ζ stability and the ability of 14-3-3ζ to bind
its target proteins (16). 14-3-3ζ
was found to be responsible for silencing of E-cadherin during EMT
(15). In the present study, a
novel NLK-interacting protein was identified. NLK was
found to inhibit cell growth capabilities using colony formation
assay. The number or size of spheres partly reveals the
proliferation of cancer stem/progenitor cells (26). The sphere formation assay revealed
that NLK markedly repressed the sphere formation capacities
in A549 cells, in the present study. Furthermore, wound healing
assay was performed to examine the effect of NLK on
migration ability. Overexpression of NLK markedly inhibited
cell migration. As an NLK-interacting protein, 14-3-3ζ
overexpressing cells revealed enhanced migration capacity.
NLK and 14-3-3ζ co-expressing cells revealed decreased
migration compared with that in cells only expressing 14-3-3ζ.
These findings suggest that NLK inhibition of migration may
be dependent on the regulation of 14-3-3ζ function.
14-3-3ζ has been previously found to promote EMT and
inhibit the expression of E-cadherin (15). In the present study, 14-3-3ζ was
found to induce EMT and repress E-cadherin in A549 cells.
Co-immunoprecipitation assay was performed with exogenous tagged
protein or endogenous proteins to verify the interaction between
NLK and 14-3-3ζ identified by LC-MS/MS analysis of
NLK-interacting proteins. In light of the key role of
NLK in the regulation of 14-3-3ζ-induced E-cadherin
repression, A549 cells stably co-expressing FLAG-14-3-3ζ and
HA-14-3-3ζ were constructed. FLAG-14-3-3ζ was found to be
co-immunoprecipitated with HA-14-3-3ζ. Notably, overexpression of
untagged NLK in FLAG-14-3-3ζ and HA-14-3-3ζ co-expressing
cells inhibits the interaction between FLAG-14-3-3ζ and HA-14-3-3ζ.
These results suggest that NLK disturbs the 14-3-3ζ-14-3-3ζ
dimerization, and hence the function of NLK to regulate
E-cadherin.
Gly-rich linkers naturally exist in a number of
proteins and have been used to create fusion proteins with
independent sequences, such as the connection with homodimer,
ligand-receptor and co-activators (18). Overexpression of 14-3-3ζ-Fusion
created by Gly-rich linker successfully repressed the expression of
E-cadherin, promoted EMT morphological changes and cell migration.
Notably, NLK lost its ability to regulate the expression of
E-cadherin, EMT morphological changes and cell migration induced by
14-3-3ζ-Fusion overexpression. In conclusion, the results from the
present study reveals that NLK interacts with 14-3-3ζ to
represses the expression of E-cadherin by interrupting 14-3-3ζ
dimer formation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Scientific
Research Youth Project of Wuxi Health and Family Planning
Commission (Q201751).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
JC, QL, TN, DS and WM conceived and designed the
experiments. JC, QL and TN performed the majority of the
experiments and assembled the data. JZ, FL, XL, YL, SR, ZL, TZ and
SH participated in the other experiments and article revision. All
authors read and approved the final manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Huang Y, Yang Y, He Y and Li J: The
emerging role of Nemo-like kinase (NLK) in the regulation of
cancers. Tumour Biol. 36:9147–9152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li SZ, Zhang HH, Liang JB, Song Y, Jin BX,
Xing NN, Fan GC, Du RL and Zhang XD: Nemo-like kinase (NLK)
negatively regulates NF-kappa B activity through disrupting the
interaction of TAK1 with IKKβ. Biochim Biophys Acta.
1843:1365–1372. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang Y, Zhe H, Massoumi R and Ke H:
Decreased expression of nemo-like kinase in melanoma is correlated
with increased vascularity and metastasis. Melanoma Res.
29:376–381. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stevens KN, Kelemen LE, Wang X, Fridley
BL, Vierkant RA, Fredericksen Z, Armasu SM, Tsai YY, Berchuck A,
Narod SA, et al: Ovarian cancer association consortium: Common
variation in Nemo-like kinase is associated with risk of ovarian
cancer. Cancer Epidemiol Biomarkers Prev. 21:523–528. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang Y, Peng C, Wu G, Wang Y, Liu R, Yang
S, He S, He F, Yuan Q, Huang Y, et al: Expression of NLK and its
potential effect in ovarian cancer chemotherapy. Int J Gynecol
Cancer. 21:1380–1387. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang Y, Jiang Y, Lu W and Zhang Y:
Nemo-like kinase associated with proliferation and apoptosis by
c-Myb degradation in breast cancer. PLoS One. 8:e691482013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lv L, Wan C, Chen B, Li M, Liu Y, Ni T,
Yang Y, Liu Y, Cong X, Mao G and Xue Q: Nemo-like kinase (NLK)
inhibits the progression of NSCLC via negatively modulating WNT
signaling pathway. J Cell Biochem. 115:81–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sa JK, Yoon Y, Kim M, Kim Y, Cho HJ, Lee
JK, Kim GS, Han S, Kim WJ, Shin YJ, et al: In vivo RNAi screen
identifies NLK as a negative regulator of mesenchymal activity in
glioblastoma. Oncotarget. 6:20145–20159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Campbell K, Rossi F, Adams J, Pitsidianaki
I, Barriga FM, Garcia-Gerique L, Batlle E, Casanova J and Casali A:
Collective cell migration and metastases induced by an
epithelial-to-mesenchymal transition in Drosophila intestinal
tumors. Nat Commun. 10:23112019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sjöberg E, Meyrath M, Milde L, Herrera M,
Lövrot J, Hägerstrand D, Frings O, Bartish M, Rolny C, Sonnhammer
E, et al: A novel ACKR2-dependent role of fibroblast-derived CXCL14
in epithelial-to-mesenchymal transition and metastasis of breast
cancer. Clin Cancer Res. 25:3702–3717. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang J, Shao S, Han D, Xu Y, Jiao D, Wu
J, Yang F, Ge Y, Shi S, Li Y, et al: High mobility group box 1
promotes the epithelial-to-mesenchymal transition in prostate
cancer PC3 cells via the RAGE/NF-κB signaling pathway. Int J Oncol.
53:659–671. 2018.PubMed/NCBI
|
|
12
|
Dong C, Yuan T, Wu Y, Wang Y, Fan TW,
Miriyala S, Lin Y, Yao J, Shi J, Kang T, et al: Loss of FBP1 by
Snail-mediated repression provides metabolic advantages in
basal-like breast cancer. Cancer Cell. 23:316–331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qi M, Zhou Y, Liu J, Ou X, Li M, Long X,
Ye J and Yu G: AngII induces HepG2 cells to activate
epithelial-mesenchymal transition. Exp Ther Med. 16:3471–3477.
2018.PubMed/NCBI
|
|
14
|
Guo F, Gao Y, Sui G, Jiao D, Sun L, Fu Q
and Jin C: miR-375-3p/YWHAZ/β-catenin axis regulates migration,
invasion, EMT in gastric cancer cells. Clin Exp Pharmacol Physiol.
46:144–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu J, Guo H, Treekitkarnmongkol W, Li P,
Zhang J, Shi B, Ling C, Zhou X, Chen T, Chiao PJ, et al: 14-3-3zeta
Cooperates with ErbB2 to promote ductal carcinoma in situ
progression to invasive breast cancer by inducing
epithelial-mesenchymal transition. Cancer Cell. 16:195–207. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Messaritou G, Grammenoudi S and Skoulakis
EM: Dimerization is essential for 14-3-3zeta stability and function
in vivo. J Biol Chem. 285:1692–1700. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tzivion G, Luo Z and Avruch J: A dimeric
14-3-3 protein is an essential cofactor for Raf kinase activity.
Nature. 394:88–92. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Reddy Chichili VP, Kumar V and Sivaraman
J: Linkers in the structural biology of protein-protein
interactions. Protein Sci. 22:153–167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mishra R, Gorlov IP, Chao LY, Singh S and
Saunders GF: PAX6, paired domain influences sequence recognition by
the homeodomain. J Biol Chem. 277:49488–49494. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wilson KA, Bär S, Maerz AL, Alizon M and
Poumbourios P: The conserved glycine-rich segment linking the
N-terminal fusion peptide to the coiled coil of human T-cell
leukemia virus type 1 transmembrane glycoprotein gp21 is a
determinant of membrane fusion function. J Virol. 79:4533–4539.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagi AD and Regan L: An inverse
correlation between loop length and stability in a
four-helix-bundle protein. Fold Des. 2:67–75. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuusinen A, Arvola M and Keinänen K:
Molecular dissection of the agonist binding site of an AMPA
receptor. EMBO J. 14:6327–6332. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang W, Prosise WW, Chen J, Taremi SS, Le
HV, Madison V, Cui X, Thomas A, Cheng KC and Lesburg CA:
Construction and characterization of a fully active PXR/SRC-1
tethered protein with increased stability. Protein Eng Des Sel.
21:425–433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mir N, Jayachandran A, Dhungel B, Shrestha
R and Steel JC: Epithelial-to-mesenchymal transition: A mediator of
sorafenib resistance in advanced hepatocellular carcinoma. Curr
Cancer Drug Targets. 17:698–706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ahn HM, Yoo JW, Lee S, Lee HJ, Lee HS and
Lee DS: Peroxiredoxin 5 promotes the epithelial-mesenchymal
transition in colon cancer. Biochem Biophys Res Commun.
487:580–586. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Castro NP, Rangel MC, Merchant AS,
MacKinnon G, Cuttitta F, Salomon DS and Kim YS: Sulforaphane
suppresses the growth of triple-negative breast cancer stem-like
cells in vitro and in vivo. Cancer Prev Res (Phila). 12:147–158.
2019. View Article : Google Scholar : PubMed/NCBI
|