Introduction
In recent years, the incidence of breast cancer in
China has been exhibiting an increasing trend on an annual basis
(1). Hematogenous micrometastasis
at initial diagnosis is considered to be one of the causes of
recurrence that affects the overall survival of breast cancer
patients. Therefore, detection of hematogenous micrometastases
based on tissue-specific markers may provide valuable information
and guidance for the early screening of high-risk breast cancer
patients (2). Small breast
epithelial mucin (SBEM) (also known as MUCL1) has
been identified as a putative breast-specific gene and has been
considered to be a promising breast-specific marker (3). In our previous study, we detected
SBEM expression in tissues and peripheral blood specimens of
breast cancer patients to analyze its correlation with prognostic
parameters. SBEM was proposed as a marker for predicting
hematogenous micrometastasis and response to neoadjuvant
chemotherapy in breast cancer (4).
However, although published studies have suggested that SBEM
may represent a suitable marker for molecular detection of isolated
tumor cells in the bone marrow and targeting bone marrow
micrometastasis in breast cancer patients (5,6),
studies that have been conducted to date in order to observe the
effect of the SBEM gene on breast cancer cells and explore
the underlying possible mechanism are sparse.
In the present study, MCF-7 and MDA-MB-231 cells
with stable SBEM knockdown or overexpression were first
generated. After detecting the effect of SBEM on the
migration and invasion abilities of MCF-7 and MDA-MB-231 cells, the
expression of EMT-related markers and regulators in
SBEM-overexpressing MCF-7 cells was monitored. Among the
EMT-related markers and regulators, E-cadherin and claudin-1 are
considered to be two important suppressors of invasion. E-cadherin
plays a crucial role in the maintenance of epithelial cell
polarization, and deficiency of this molecule causes cancer
metastasis due to the loss of cell-cell adhesion, with concomitant
increased cell motility (7). Breast
cancer patients with lower expression of E-cadherin are at a higher
risk of recurrence and metastasis, and have a worse prognosis
(8). Claudin-1 plays a key role in
the formation of tight junctions (9). In several cancers, the loss of
claudin-1 expression has been associated with cancer progression,
invasiveness, and acquisition of the metastatic phenotype (10,11).
Claudin-1 is frequently downregulated in cancer, and its
downregulation has been shown to be associated with poor clinical
outcome in human invasive breast cancer (12). The aim of the present study was to
elucidate the possible mechanisms of action of SBEM by
observing its effects on EMT-related markers and regulators,
including E-cadherin and claudin-1, in the hope of the results
laying an experimental foundation for further exploring the role of
SBEM in breast cancer.
Materials and methods
Cell characteristics and culture
MCF-7 is a widely studied epithelial cancer cell
line derived from breast adenocarcinoma, with the characteristics
of differentiated mammary epithelium. MCF-7 cells express estrogen
receptor-α (ER-α), as well as androgen, progesterone and
glucocorticoid receptors, which make them valuable tools in medical
research. Although MCF-7 cells are easy to propagate, they are
generally a slow-growing population, with a doubling time of 30–40
h. MDA-MB-231 is a highly aggressive, invasive and poorly
differentiated triple-negative breast cancer (TNBC) cell line.
Similar to other invasive cancer cell lines, the invasiveness of
the MDA-MB-231 cells is mediated by proteolytic degradation of the
extracellular matrix. In a 3D culture, this cell line displays
endothelial-like morphology and is distinguished by the stellate
projections that often bridge multiple cell colonies.
MCF-7 and MDA-MB-231 cells were purchased from the
Cell Bank of the Chinese Academy of Sciences (Shanghai, China). The
cells were grown in 25-cm2 cell culture flasks with
RPMI-1640 medium (HyClone; GE Healthcare Life Sciences) or L15
(HyClone; GE Healthcare Life Sciences) supplemented with 10% fetal
bovine serum (FBS; HyClone; GE Healthcare Life Sciences), 2 mM
L-glutamate, 100 U/ml penicillin G, and 100 U/ml streptomycin at
37°C in 5% CO2 and 95% air. The cells were then seeded
into 6- or 24-well culture plates (Corning, Inc.) with the cell
confluence of ~70% prior to transfection.
Construction of recombinant shRNA
lentivirus vector
According to the design principle of mammalian
eukaryotic cell RNA interference, shRNA sequences were designed for
the different targets of mucin-like protein 1. Each interference
sequence was synthesized into a single chain of sense and
antisense, and a double chain was formed after annealing. The
primers were as follows:
5′-CCGGGTGTGTCCCTGAGATGGAATCCTCGAGGATTCCATCTCAGGGACACACTTTTTG-3′
(sense) and
5′-AATTCAAAAAGTGTGTCCCTGAGATGGAATCCTCGAGGATTCCATCTCAGGGACACAC-3′
(antisense).
The two chains were annealed into double chains,
connected by T4DNA ligase and the AgeI/EcoRI double
enzyme cutting carrier PLKO1 (Fig.
1), converted by DH5a competent cells, and single clones were
selected. The contrast scramble plasmid was provided by Genesent,
and its hairpin structure was as follows:
5′-CCTAAGGTTAAGTCGCCCTCGCTCGAGCGAGGGCGACTTAACCTTAGG-3.
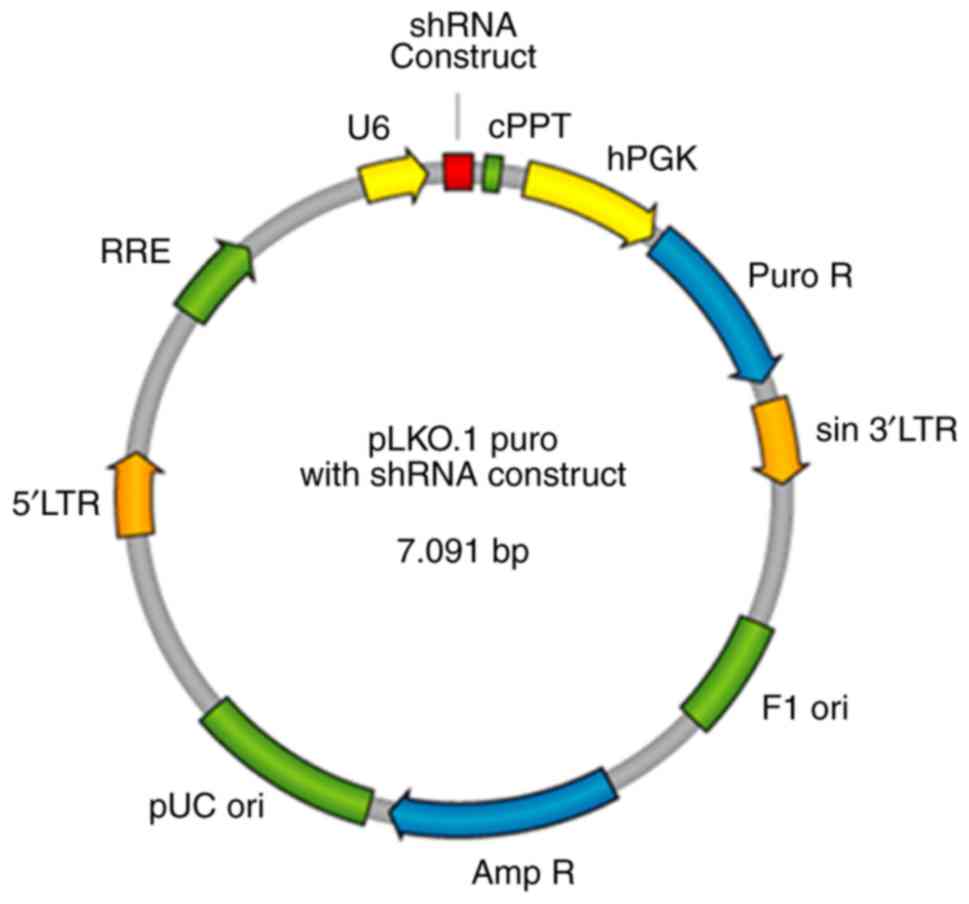 | Figure 1.SBEM shRNA plasmid map.
SBEM, small breast epithelial mucin; RRE, Rev responsive
element; 5′LTR, 5′ long terminal repeat; pUC ori, pUC bacterial
origin of replication; Amp R, ampicillin resistance gene for
selection of pLKO.1 plasmid in bacterial cells; Puro R,
puromycin-resistance gene for selection of pLKO.1 plasmid in
mammalian cells; sin 3′LTR, 3′ self-inactivating long terminal
repeat; F1 ori, F1 bacterial origin of replication; hPGK, human
phosphoglycerate kinase promoter drives expression of puromycin;
cPPT, central polypurine tract. |
Construction of SBEM lentivirus
expression plasmid
SBEM cDNA was provided by Genesent, and the
lentivirus overexpression vector was pCDH-CMV-MCS-EF1-Puro. The
plasmid map is shown in the Fig. 2.
The cloning primers were as follows:
5′-TAGAGCTAGCGAATTATGAAGTTCTTAGCAGTCC-3′ (sense) and
5′-AGATCCTTCGCGGCCTCAGGGACACACTCTACCA-3′ (antisense).
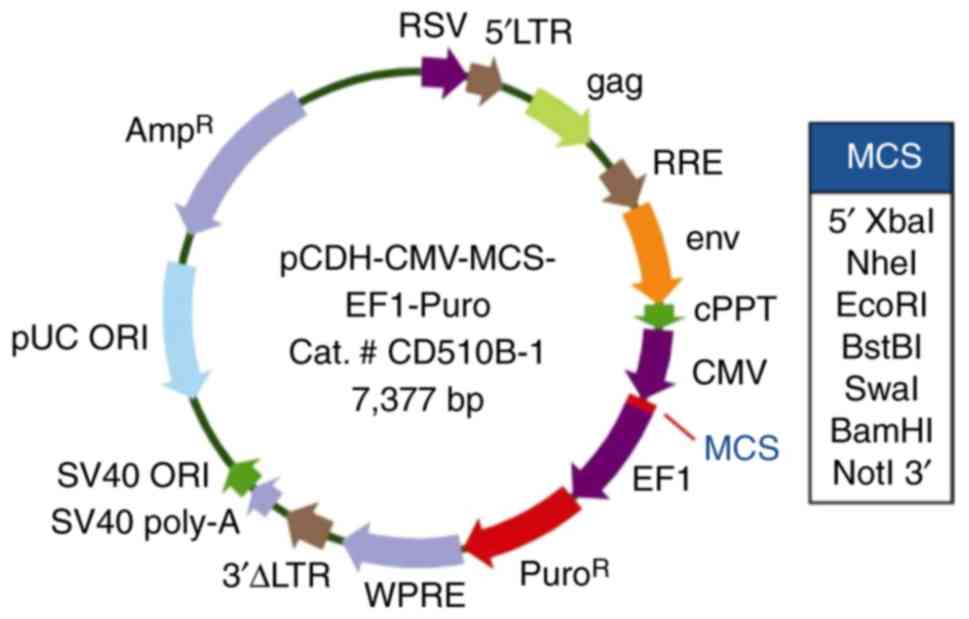 | Figure 2.The SBEM-overexpressing
plasmid map. SBEM, small breast epithelial mucin. SV40 ORI,
SV40 promoter/origin; SV40 poly-A, simian vacuolating virus 40
poly-A; 3′ΔLTR, 3′ self-inactivating long terminal repeat; WPRE, In
cis Woodchuck hepatitis virus post &
hyphentranscriptional regulatory element; EF1, elongation factor 1;
MCS, multiple cloning site; CMV, cytomegalovirus; env, envelope;
RSV, Rous sarcoma virus. |
Generation of stable SBEM knockdown or
overexpression cell lines
MCF-7 and MDA-MB-231 cells with stable SBEM
knockdown or overexpression were generated by lentiviral vectors
carrying either sh-SBEM, a SBEM overexpression
construct, or the respective negative controls (Genesent), in
accordance with the manufacturer's instructions. Cells were placed
into 60-mm dishes at 3×105/dish and allowed to grow
overnight. Lentivirus expression plasmid (8 µg) was mixed with 20
µl Lipofectamine 2000 (Thermo Fisher Scientific, Inc.) and
transfected into the cells according to the manufacturer's
instructions. Twenty-four hours after transfection, the cells were
trypsinized, diluted, and placed into 96-well plates. Transfected
cells were then selected with 2 µg/ml puromycin and 200 µg/ml G418.
Western blotting was conducted to detect the knockdown and
overexpression effects of SBEM.
Scratch wound-healing assay
To determine the regeneration and repair abilities
of breast cancer cells, 4×105 MCF-7 and MDA-MB-231 cells
were seeded in 6-well plates and incubated at 37°C with 5%
CO2 overnight. Artificial wounds were created using a
10-µl pipette tip (0 h) to generate a gap in the confluent cell
layer with confluence percentage of ~80%. The cells were washed
with PBS twice and incubated with serum-free medium at 37°C with 5%
CO2 as a control. At 0 and 8 h, phase-contrast images
were captured using a microscope at a magnification of ×400
(CX41-32C02PH, Olympus Corporation).
Cell migration and invasion
assays
The detailed procedures of cell migration and
invasion assays were conducted as previously reported (13). Briefly, 3×105 cells were
suspended in serum-free medium and seeded into the upper layer of
Transwell membrane with an 8-µm pore size in a 24-well plate
(Corning, Inc.). The membranes were coated with Matrigel (1:8; BD
Biosciences) for invasion assays, or left uncoated for migration
assays. Medium containing 10% FBS was placed in the bottom chamber
as an attractant. After 24 h, the cells were fixed in 4%
paraformaldehyde for 15 min at room temperature and stained with
0.1% crystal violet solution for 15 min. The invading cells were
then examined and counted in 10 randomly selected fields under a
light microscope at a magnification of ×400. The mean number of
invading cells was then calculated.
Real-time PCR
Total RNA of 5×106 MCF-7 cells was
extracted using the Trizol RNA extraction protocol (cat. no.
10606ES60, Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. Reverse transcription of mRNA to
cDNA was performed in 20 µl reaction volumes with random priming
using an RT-PCR Kit (cat. no. RR047A, Takara Biomedical Technology
Co., Ltd.). The sequences of primers used in this study are listed
in Table I, and the real-time PCR
reaction system is provided in Table
II. The primers were synthesized by Beijing AuGCT Biotech Co.
Ltd. Quantitative PCR was performed using SYBR Green PCR Kit (cat.
no. RR820A, Takara Biomedical Technology Co. Ltd.) and Fast
Real-Time PCR System (ABI 7900HT, Applied Biosystems; Thermo Fisher
Scientific, Inc.). The detailed process was similar to that
previously described (4). The
increase of fluorescence was detected due to the exponential
accumulation of PCR products and the 2−ΔΔCq method was
used to calculate the relative quantity of gene expression in each
sample (14).
 | Table I.Sequence of the primers for real-time
PCR. |
Table I.
Sequence of the primers for real-time
PCR.
| Primer | Sequence (5′ to
3′) |
|---|
| N-cadherin
forward |
GATGTTGAGGTACAGAATCGT |
| N-cadherin
reverse |
GGTCGGTATGGATGGCGA |
| Twist forward |
GGAGTCCGCAGTCTTACGAG |
| Twist reverse |
TCTGGAGGACCTGGTAGAGG |
| Vimentin
forward |
GGACCAGCTAACCAACGACA |
| Vimentin
reverse |
AAGGTCAAGACGTGCCAGAG |
| E-cadherin
forward |
ATTCTGATTCTGCTGCTCTTG |
| E-cadherin
reverse |
AGTAGTCATAGTCCTGGTCTT |
| Claudin-1
forward |
CAGCTGTTGGGCTTCATTCTC |
| Claudin-1
reverse |
ATCACTCCCAGGAGGATGCC |
| Table II.Real-time PCR reaction system (25
µl). |
Table II.
Real-time PCR reaction system (25
µl).
| Reaction | Volume (µl) |
|---|
| cDNA | 2 |
| SYBR Green I |
12.5 |
| Primer F (0.2
µmol/l) |
0.5 |
| Primer R (0.2
µmol/l) |
0.5 |
| Sterile deionized
water |
9.5 |
Western blot assay
The proteins were quantified using the BCA method.
An amount of 10 µl protein plus 10 µl PBS were loaded and 200 µl
working liquid was added in 96-well plates. The absorbance value at
560 nm was measured by enzyme-labeling instrument. The
concentration of proteins was calculated on the basis of the
standard curve. For western blotting, 1×106 cells were
plated in 100-mm Petri dishes for 24 h. The cells were then washed
with cold PBS and lysed with 200 µl of cold lysis buffer [150
mmol/l NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 50
mmol/l Tris-HCl (pH 7.2), 0.2 mmol/l sodium vanadate, 1%
phenylmethylsulfonyl fluoride and 0.2% aprotinin].
The samples were kept on ice for 20 min and then
spun at 12,000 × g at 4°C for 20 min, and the protein concentration
of the supernatant was determined. Cell lysates were fractionated
on 10% SDS-PAGE, and protein was transferred onto nitrocellulose
membranes (Pall Life Sciences). The membranes were blocked with 5%
skimmed milk powder dissolved in TBST at room temperature for 2 h.
The membranes were then probed with primary antibodies against
SBEM (dilution 1:1,000, cat. no. HPA-039093; Sigma-Aldrich;
Merck KGaA), N-cadherin (dilution 1:500, cat. no. 13116S; Cell
Signaling Technology, Inc.), E-cadherin (dilution 1:500, cat. no.
3195S; Cell Signaling Technology, Inc.), Twist (dilution 1:500,
cat. no. 69366S; Cell Signaling Technology, Inc.), vimentin
(dilution 1:500, cat. no. 5741S; Cell Signaling Technology, Inc.)
and claudin-1 (dilution 1:500, cat. no. 13995S; Cell Signaling
Technology, Inc.). The expression of GAPDH as control was
determined using anti-GAPDH (dilution 1:500, cat. no. 5174S; Cell
Signaling Technology, Inc.). After hybridization at 4°C overnight,
HRP-labeled IgG (dilution 1:500, cat. no. 074S; Cell Signaling
Technology, Inc.) was added and incubated at 37°C for 2 h. The
membrane was washed with TBST (0.1% Tween-20) for 3 times, and ECL
chemiluminescence system (Applygen Technologies Inc.) was used for
coloration. The immunoreactive bands were detected using an Odyssey
Infrared Imaging System (Gene Company Ltd.). The intensity of each
band was measured with Odyssey 3.0 software (Li Cor Inc.).
Statistical analysis
All data are presented as mean ± standard deviation.
Statistical analysis was performed using Statistical Package for
the Social Sciences version 14.0 (SPSS, Inc.). All statistical
tests were two-sided. The comparison between two groups of samples
adopted the t-test, and P-value ≤0.05 was considered to indicate a
statistically significant difference.
Results
Observation of cell morphology after
transfection
In accordance with the manufacturer's instructions,
MCF-7 and MDA-MB-231 cells with stable SBEM knockdown or
overexpression were generated by lentiviral vectors carrying
sh-SBEM, a SBEM overexpression construct, or the
negative controls, respectively. The microscopic observations of
cell morphology are presented in Figs.
3 and 4. MCF-7 cells displayed
typical epithelioid characteristics. They were small and polygonal
in shape. MDA-MB-231 cells are mostly spindle-like with narrow
strip in shape, and some of the cells adopted a more rounded
morphology.
SBEM protein expression in MCF-7 and
MDA-MB-231 cells after transfection
In order to test and verify the knockdown and
overexpression of the SBEM gene, western blotting was used
to detect SBEM protein expression in MCF-7 and MDA-MB-231
cells. Compared with the scramble group, SBEM protein
expression decreased significantly after cells were transfected
with sh-SBEM. Compared with the vector group, SBEM
protein expression was increased significantly after the cells were
transfected with the overexpression plasmid. The difference was
statistically significant in both MCF-7 and MDA-MB-231 cells
(P<0.05). Therefore, MCF-7 and MDA-MB-231 cell lines with stable
SBEM knockdown or overexpression were successfully
established (Fig. 5).
SBEM promotes scratch wound healing of
breast cancer cells
Wound healing was observed after 8 h. The results of
the MCF-7 cells demonstrated that the cell repair rate in
sh-SBEM group was (6.8%) much lower than that in the
scramble group (39.6%). The cell repair rate in the SBEM
overexpression group was higher (48.1%) than that in the vector
group (27.0%) (Fig. 6 and Table III). The results of the MDA-MB-231
cells demonstrated that the cell repair rate in the sh-SBEM
group (4.8%) was markedly lower compared with that in the scramble
group (31.0%). The cell repair rate in the SBEM
overexpression group was higher (69.9%) compared with that in the
vector group (26.2%) (Fig. 7,
Table III). These results
indicated that SBEM knockdown obviously inhibited and
SBEM overexpression obviously promoted the migration ability
of the breast cancer cells.
| Table III.Cell repair rate (%) of each group in
the scratch wound-healing assay. |
Table III.
Cell repair rate (%) of each group in
the scratch wound-healing assay.
|
| Cell line |
|---|
|
|
|
|---|
| Group | MCF-7 | MDA-MB-231 |
|---|
| sh-SBEM | 6.8 | 4.8 |
| Scramble | 39.6 | 31.0 |
| SBEM
overexpression | 48.1 | 69.9 |
| Vector | 27.0 | 26.2 |
SBEM promotes the invasion of breast
cancer cells
The numbers of cells crossing the basement membrane
were recorded for 24 h. In the MCF-7 cells, the cell numbers were
20±1.15 in the sh-SBEM group, and 75.6±2.01 in the scramble
group (P<0.05); the cell numbers were 81.2±1.47 in the
SBEM-overexpressing group, and 25.5±1.08 in the vector
group. The difference between the two groups was statistically
significant (P<0.05; Fig. 8). In
MDA-MB-231 cells, the cell numbers were 48.5±1.35 in sh-SBEM
group, and 161.7±1.25 in the scramble group; the cell numbers were
241.7±1.15 in the SBEM-overexpressing group, and 118±1.63 in
the vector group. The difference between the two groups was
statistically significant (P<0.05; Fig. 9). The results demonstrated that
SBEM knockdown obviously inhibited and SBEM
overexpression obviously promoted the invasion ability of breast
cancer cells.
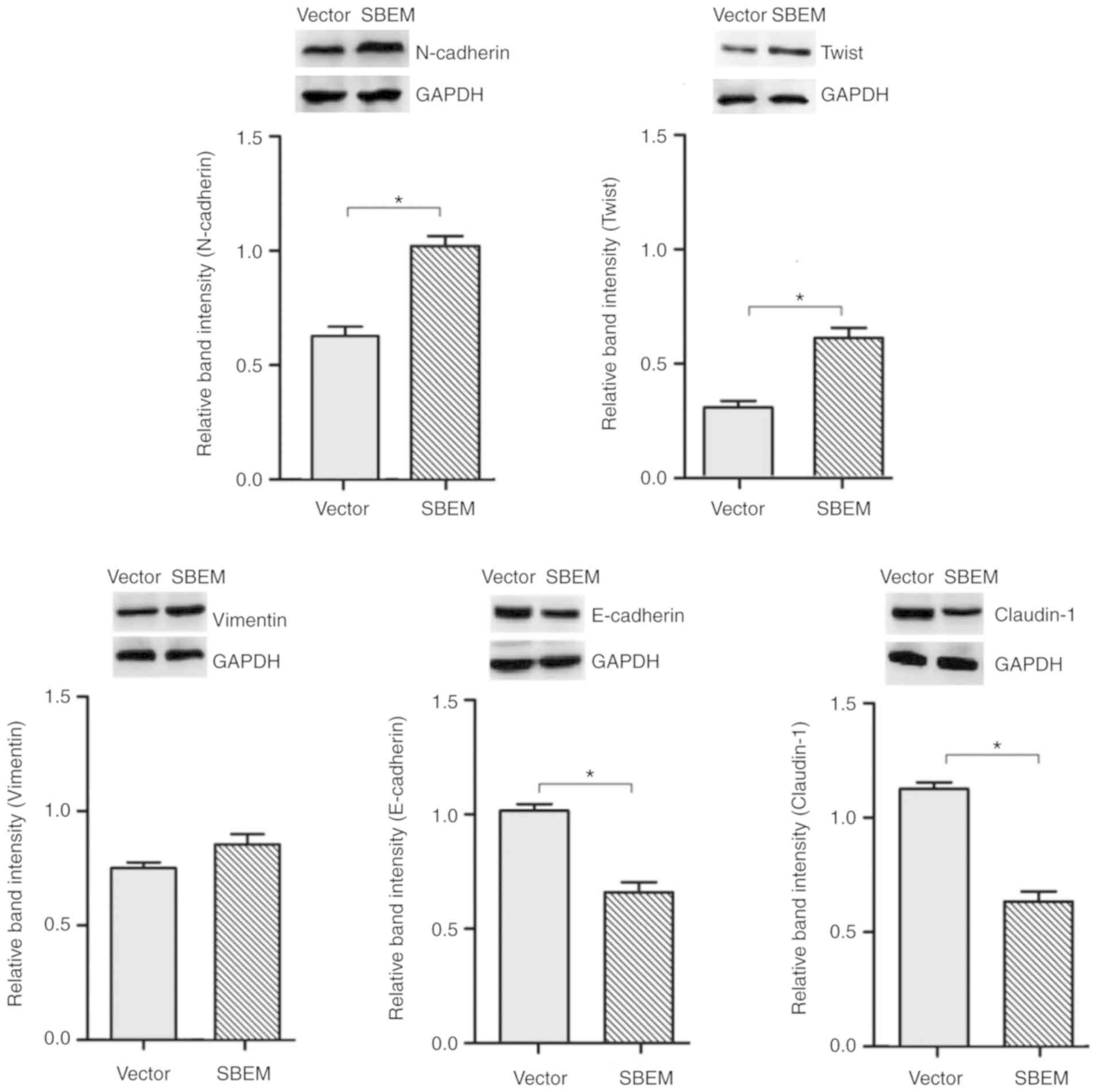
SBEM affects the expression of
EMT-related markers and regulators
In order to confirm the association of the
SBEM gene with the process of EMT, the real-time PCR and
western blotting were used to detect the expression of EMT-related
markers and regulators in SBEM-overexpressing MCF-7 cells.
The results revealed that the levels of N-cadherin, Twist and
vimentin were elevated, while those of E-cadherin and claudin-1
were decreased following SBEM overexpression. The relative
mRNA expression levels, protein bands and relative band intensities
are presented in Figs. 10 and
11.
Discussion
In recent years, breast cancer has become the
primary cause of cancer-related death among women worldwide
(15). Patients with isolated tumor
cells or micrometastases have a comparably poor 5-year disease-free
survival rate (16). Detection of
breast cancer micrometastases based on specific molecular markers
and exploration of the potential underlying mechanism may provide
useful information for clinical research (17). Small breast epithelial mucin
(SBEM) is a type of secretory protein, which belongs to the
family of MUC (18). The
SBEM gene is mainly expressed in the breast and salivary
glands (19), and exhibits higher
expression in breast cancer tissue and metastatic lymph nodes
(20,21). We previously reported that
SBEM expression had the potential to serve as a useful and
specific marker for hematogenous metastasis of breast cancer
(3). Our previous study
demonstrated that the expression of SBEM was significantly
correlated with the disease-free and overall survival of patients
with TNBC, and that it appears to be a promising prognostic
biomarker for TNBC diagnosis and treatment (22). Due to its high specificity for
breast tissue, SBEM was considered to play an important role
in the metastatic process of breast cancer (23,24).
However, little is known concerning the potential role and
mechanism of action of the SBEM gene in the migration and
invasion of breast cancer cells.
In the present study, MCF-7 and MDA-MB-231 cells
with stable SBEM knockdown or overexpression were generated
by lentiviral vectors. In both cell lines, the cell repair rate in
the sh-SBEM group was markedly lower when compared with that
in the scramble group, and the cell repair rate in the SBEM
overexpression group was markedly higher when compared with that in
the vector group. These findings indicated that SBEM
knockdown obviously inhibited and SBEM overexpression
obviously enhanced the migration ability of breast cancer cells.
Similarly, in both MCF-7 and MDA-MB-231 cells, the numbers of cells
that invaded through the basement membrane over 24 h were markedly
lower in the sh-SBEM group compared with those in the
scramble group, whereas they were markedly higher in the
SBEM-overexpressing group compared with those in the vector
group. These findings indicated that SBEM knockdown
obviously inhibited and SBEM overexpression obviously
promoted the invasion ability of the breast cancer cells.
Epithelial-mesenchymal transition (EMT) is a
cellular process during which epithelial cells acquire mesenchymal
phenotypes and behavior following the downregulation of epithelial
features (25). The underlying
cytological mechanisms include changes in cell morphology, loss of
polarity, decreased adhesion, weakening of connections to the
basement membrane, and the increase of cell migration and invasion
abilities. Approximately 95% of breast cancer cells originate from
epithelial cells, whereas the surrounding cells display mesenchymal
phenotypes. Mesenchymal cells are characterized by stronger
mobility, which enables invasion of blood and lymphatic vessels and
metastasis to distant organs (26).
The loss of E-cadherin expression has been considered as the key
step during EMT in breast cancer. When the expression of E-cadherin
is decreased, the intercellular adhesions become weaker, resulting
in the loss of cell polarity. The expression of E-cadherin has been
shown to be negatively correlated with the migration and invasion
abilities of breast cancer cells (27,28).
Claudin-1 is the main cytoskeletal protein that constitutes the
tight junction chain of epithelial cells. The loss of claudin-1
expression leads to the separation of epithelial cells and an
increase of mobility, which facilitates the invasion and metastasis
of cancer cells after EMT (9).
Vimentin, a type of intermediate filament that is distributed in
mesenchymal tissues and cells, helps maintain interstitial cell
characteristics (30). When the
expression of E-cadherin is decreased, the cytoskeleton mainly
composed of keratin is transformed into vimentin-based cytoskeleton
proteins, resulting in a change of cell morphology and rendering
tumor cells more invasive (31,32).
Twist is a basic-helix-loop-helix transcription factor that
promotes cell migration and tissue recombination, which may enhance
cell invasiveness (33).
Overexpression of Twist was also found to induce angiogenesis in
breast cancer (34). In the present
study, the expression of N-cadherin, Twist and vimentin was found
to be increased, while the expression of E-cadherin and claudin-1
was decreased following SBEM overexpression in MCF-7 cells.
These findings indicated that overexpression of SBEM
downregulated the expression of the epithelial marker E-cadherin,
and upregulated the expression of the mesenchymal markers
N-cadherin and vimentin. Furthermore, SBEM promoted EMT by
upregulating the expression of the transcription factor Twist. Of
note, the increase in the expression of vimentin was not as
significant compared with that of N-cadherin and Twist following
SBEM overexpression. It was hypothesized that this may be
associated with the early observational time point, as vimentin
elevation is usually a late event during the EMT process.
EMT marks the initiation of the malignant phenotype
transformation process, and it is the first step in the
invasion-metastasis cascade of breast cancer cells through the
basement membrane (35). There are
a number of EMT-related signaling pathways in breast cancer,
including TGF-β, NF-κB, Notch, Wnt/β-catenin and PI3K/AKT, among
others (36–40). Determining which signaling pathways
are involved in SBEM-induced EMT constitutes an important
research direction. There remain a number of relevant mechanisms
and pathways to be investigated.
In summary, the effects of SBEM on the
invasion-metastasis cascade suggest that it may be a potential
effective target for anti-metastasis treatment in breast cancer.
However, there are yet no data from experiments in vivo to
validate our in vitro findings, and there are no exact data
on EMT-related signaling pathways of SBEM. The findings of
the present study may prove helpful as the basis for further
research on the mechanisms of action and relevant pathways of
SBEM. More focus will be placed on these two aspects of
research to acquire relevant data in the future.
Acknowledgements
We thank Professor Guo-Xin Li from The Second
Affiliated Hospital of Liaoning University of Traditional Chinese
Medicine for his secretarial and organizational support in our
experiments.
Funding
The present study was financially supported by
Foundation projects: 2019 Subsidization Project of Liaoning
Province Natural Foundation (2019-MS-351 and 2019-MS-210); 2018
Guidance Project of Liaoning Province Natural Foundation
(20180550359); 2017 Liaoning Province TCM Clinics (Specialized)
Branch Capacity Building Project; 2018 Liaoning Province Doctoral
Start-up Foundation (20180540043).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
QHL, ZZL and BL contributed to the study conception
and design. Material preparation, data collection and analysis were
performed by QHL, ZZL, YNG, XL, XDX, ZDZ and YHM. The first draft
of the manuscript was written by ZZL. All authors read, revised and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Li T, Mello-Thoms C and Brennan PC:
Descriptive epidemiology of breast cancer in China: Incidence,
mortality, survival and prevalence. Breast Cancer Res Treat.
159:395–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schindlbeck C, Andergassen U, Jueckstock
J, Rack B, Janni W and Jeschke U: Disseminated and circulating
tumor cells in bone marrow and blood of breast cancer patients:
Properties, enrichment, and potential targets. J Cancer Res Clin.
142:1883–1895. 2016. View Article : Google Scholar
|
|
3
|
Skliris GP, Hubé F, Gheorghiu I, Mutawe
MM, Penner C, Watson PH, Murphy LC, Leygue E and Myal Y: Expression
of small breast epithelial mucin (SBEM) protein in tissue
microarrays (TMAs) of primary invasive breast cancers.
Histopathology. 52:355–369. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu ZZ, Xie XD, Qu SX, Zheng ZD and Wang
YK: Small breast epithelial mucin (SBEM) has the potential to be a
marker for predicting hematogenous micrometastasis and response to
neoadjuvant chemotherapy in breast cancer. Clin Exp Metastasis.
27:251–259. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ayerbes MV, Diaz-Prado S, Ayude D, Campelo
RG, Iglesias P, Haz M, Medina V, Gallegos I, Quindos M and Aparicio
LA: In silico and in vitro analysis of small breast epithelial
mucin as a marker for bone marrow micrometastasis in breast cancer.
Adv Exp Med Biol. 617:331–339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Voulgari A and Pintzas A:
Epithelial-mesenchymal transition in cancer metastasis: Mechanisms,
markers and strategies to overcome drug resistance in the clinic.
Biochim Biophys Acta. 1796:75–90. 2009.PubMed/NCBI
|
|
7
|
Corso G, Pravettoni G, Galimberti V and
Veronesi P: Clinical implication of E-cadherin deficiency in
lobular breast cancer. Breast Cancer Res Treat. 173:751–752. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luo CW, Wu CC, Chang SJ, Chang TM, Chen
TY, Chai CY, Chang CL, Hou MF and Pan MR: CHD4-mediated loss of
E-cadherin determines metastatic ability in triple-negative breast
cancer cells. Exp Cell Res. 363:65–72. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou B, Blanchard A, Wang N, Ma X, Han J,
Schroedter I, Leygue E and Myal Y: Claudin 1 promotes migration and
increases sensitivity to tamoxifen and anticancer drugs in
luminal-like human breast cancer cells MCF7. Cancer Invest.
33:429–439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Upmanyu N, Bulldan A, Papadopoulos D,
Dietze R, Malviya VN and Scheiner-Bobis G: Impairment of the
Gnα11-controlled expression of claudin-1 and MMP-9 and collective
migration of human breast cancer MCF-7 cells by DHEAS. J Steroid
Biochem Mol Biol. 182:50–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sheehan GM, Kallakury BV, Sheehan CE,
Fisher HA, Kaufman RP Jr and Ross JS: Loss of claudins-1 and −7 and
expression of claudins-3 and −4 correlate with prognostic variables
in prostatic adenocarcinomas. Hum Pathol. 38:564–569. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blanchard AA, Ma X, Dueck KJ, Penner C,
Cooper SC, Mulhall D, Murphy LC, Leygue E and Myal Y: Claudin 1
expression in basal-like breast cancer is related to patient age.
BMC Cancer. 13:2682013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu X, Huang S, Zeng L, Ma J, Sun S, Zeng
F, Kong F and Cheng X: HMOX-1 inhibits TGF-β-induced
epithelial-mesenchymal transition in the MCF-7 breast cancer cell
line. Int J Mol Med. 40:411–417. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta DeltaC(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Torre LA, Islami F, Siegel RL, Ward EM and
Jemal A: Global cancer in women: Burden and trends. Cancer
Epidemiol Biomarkers Prev. 26:444–457. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
de Boer M, van Deurzen CH, van Dijck JA,
Borm GF, van Diest PJ, Adang EM, Nortier JW, Rutgers EJ, Seynaeve
C, Menke-Pluymers MB, et al: Micrometastases or isolated tumor
cells and the outcome of breast cancer. N Engl J Med. 361:653–663.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hubé F, Mutawe M, Leygue E and Myal Y:
Human small breast epithelial mucin: The promise of a new breast
tumor biomarker. DNA Cell Biol. 23:842–849. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park HK and Seov UH: MUC1 from the mucin
family as potential tools in breast cancer immunotherapy. J Breast
Cancer. 12:1252009. View Article : Google Scholar
|
|
19
|
Kontani K, Taguchi O, Narita T, Izawa M,
Hiraiwa N, Zenita K, Takeuchi T, Murai H, Miura S and Kannagi R:
Modulation of MUC1 mucin as an escape mechanism of breast cancer
cells from autologous cytotoxic T-lymphocytes. Br J Cancer.
84:1258–1264. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heublein S, Mayr D, Egger M, Karsten U,
Goletz S, Angele M, Gallwas J, Jeschke U and Ditsch N:
Immunoreactivity of the fully humanized therapeutic antibody
PankoMab-GEX™ is an independent prognostic marker for breast cancer
patients. J Exp Clin Cancer Res. 34:502015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gerber PA, Hevezi P, Buhren BA, Martinez
C, Schrumpf H, Gasis M, Grether-Beck S, Krutmann J, Homey B and
Zlotnik A: Systematic identification and characterization of novel
human skin-associated genes encoding membrane and secreted
proteins. PLoS One. 8:e639492013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu L, Liu Z, Qu S, Zheng Z, Liu Y, Xie X
and Song F: Small breast epithelial mucin tumor tissue expression
is associated with increased risk of recurrence and death in
triple-negative breast cancer patients. Diagn Pathol. 8:712013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Miksicek RJ, Myal Y, Watson PH, Walker C,
Murphy LC and Leygue E: Identification of a novel breast- and
salivary gland-specific, mucin-like gene strongly expressed in
normal and tumor human mammary epithelium. Cancer Res.
62:2736–2740. 2002.PubMed/NCBI
|
|
24
|
Liu Z, Guo F and Xie X: Individual
detection significances of small breast epithelial mucin (SBEM) and
human mammaglobin (hMAM) expressions in peripheral blood of breast
cancer patients. Chin-Ger J Clin Oncol. 11:716–720. 2012.
View Article : Google Scholar
|
|
25
|
Yang J, Antin P, Berx G, Blanpain C,
Brabletz T, Bronner M, Campbell K, Cano A, Casanova J, Christofori
G, et al: Guidelines and definitions for research on
epithelial-mesenchymal transition. Nat Rev Mol Cell Bio. Apr
16–2020.(Epub ahead of print). View Article : Google Scholar
|
|
26
|
Creighton CJ, Gibbons DL and Kurie JM: The
role of epithelial-mesenchymal transition programming in invasion
and metastasis: A clinical perspective. Cancer Mang Res. 5:187–195.
2013.
|
|
27
|
Vergara D, Simeone P, Latorre D, Cascione
F, Leporatti S, Trerotola M, Giudetti AM, Capobianco L, Lunetti P,
Rizzello A, et al: Proteomics analysis of E-cadherin knockdown in
epithelial breast cancer cells. J Biotechnol. 202:3–11. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Carrizo MN: Loss of E-cadherin expression
and epithelial mesenchymal transition (EMT) as key steps in tumor
progression. J Cancer Prev Curr Res. 7:002352017. View Article : Google Scholar
|
|
29
|
Bar I, Merhi A, Larbanoix L, Constant M,
Haussy S, Laurent S, Canon JL and Delrée P: Silencing of casein
kinase 1 delta reduces migration and metastasis of triple negative
breast cancer cells. Oncotarget. 9:30821–30836. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu F, Gu LN, Shan BE, Geng CZ and Sang
MX: Biomarkers for EMT and MET in breast cancer: An update. Oncol
Lett. 12:4869–4876. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Libring S and Solorio L: Cancer
mechanobiology: Interaction of biomaterials with cancer cells.
Biomaterials for Cancer Therapeutics. 445–470. 2020. View Article : Google Scholar
|
|
32
|
Trogden KP, Battaglia RA, Kabiraj P,
Madden VJ, Herrmann H and Snider NT: An image-based small-molecule
screen identifies vimentin as a pharmacologically relevant target
of simvastatin in cancer cells. FASEB J. 32:2841–2854. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Y, Qin L, Sun T, Wu H, He T, Yang Z, Mo
Q, Liao L and Xu J: Twist1 promotes breast cancer invasion and
metastasis by silencing Foxa1 expression. Oncogene. 36:1157–1166.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mironchik Y, Winnard PT Jr, Vesuna F, Kato
Y, Wildes F, Pathak AP, Kominsky S, Artemov D, Bhujwalla Z, Van
Diest P, et al: Twist overexpression induces in vivo angiogenesis
and correlates with chromosomal instability in breast cancer.
Cancer Res. 65:10801–10809. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mali AV, Joshi AA, Hegde MV and Kadam SS:
Enterolactone modulates the ERK/NF-κB/Snail signaling pathway in
triple-negative breast cancer cell line MDA-MB-231 to revert the
TGF-β-induced epithelial-mesenchymal transition. Cancer Biol Med.
15:137–156. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pires BR, Mencalha AL, Ferreira GM, de
Souza WF, Morgado-Díaz JA, Maia AM, Corrêa S and Abdelhay ES:
NF-kappaB is involved in the regulation of EMT genes in breast
cancer cells. PLoS One. 12:e01696222017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li L, Tang P, Li S, Qin X, Yang H, Wu C
and Liu Y: Notch signaling pathway networks in cancer metastasis: A
new target for cancer therapy. Med Oncol. 34:1802017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang Z, Ji L, Jiang G, Liu R, Liu Z, Yang
Y, Ma Q and Zhao H: FL118, a novel camptothecin analogue,
suppressed migration and invasion of human breast cancer cells by
inhibiting epithelial-mesenchymal transition via the Wnt/β-catenin
signaling pathway. Biosci Trends. 12:40–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen L, Fu H, Luo Y, Chen L, Cheng R,
Zhang N and Guo H: cPLA2α mediates TGF-β-induced
epithelial-mesenchymal transition in breast cancer through PI3k/Akt
signaling. Cell Death Dis. 8:e27282017. View Article : Google Scholar : PubMed/NCBI
|