Introduction
Lung cancer is the leading cause of
cancer-associated mortality in males and females worldwide
(1). Non-small cell lung cancer
(NSCLC) is the most common lung cancer type, accounting for ~85% of
all lung cancers (2,3). However, NSCLC is relatively resistant
to chemotherapy and radiotherapy and the prognosis is poor, with an
estimated overall 5-year survival of <15% (4). Therefore, it is a matter of great
urgency to develop effective treatment strategies as well as
discovering novel therapeutic agents to treat lung cancer.
Traditional Chinese medicines (TCMs) obtained from
plants have been widely considered as attractive sources of
anticancer drugs owing to fewer side effects as compared with
certain other chemotherapeutics (5,6). It
has been estimated that nearly half of the modern medicines in
clinical use are small molecules derived from natural products
(7), many of which exhibit
extensive activities by downregulating the signaling cascades of
growth factors, such as insulin-like growth factor (IGF). Among
them, parthenolide (PTL), which is derived from the plant feverfew
(Tanacetum parthenium; a flowering plant in the daisy
family, Asteraceae), has been reported to exhibit a variety of
biological activities, including antioxidant, anti-inflammatory and
antitumor effects against various cancer types (8), including colorectal (9,10),
pancreatic (11) and breast cancer
(12,13). These effects may be exerted through
suppressing the proliferation, migration and invasion, and inducing
apoptosis of cancer cells. Of note, recent studies have indicated
that PTL also exhibits anticancer activity in lung cancer (14,15).
However, the exact mechanisms of action have remained largely
elusive.
Signaling through receptor tyrosine kinase pathways
is largely involved in tumor growth and progression (16). Accumulating evidence suggests that
IGF signaling, which is comprised of the IGF ligand and IGF
receptor (IGF-R), contributes to cell proliferation,
differentiation and progression of numerous tumor types (17). Numerous studies have indicated that
IGF-1R is overexpressed in numerous cancer types, including breast,
prostate and colon cancers (17),
and upregulation of IGF-1R results in chemotherapy resistance
(17,18). On the other hand, high expression
levels of IGF-1, a ligand of IGF-1R, are closely associated with
increased risk of multiple types of cancer (19). IGF-1 activates IGF-1R and then
phosphorylates the downstream substrates through tyrosine kinase
activity, resulting in activation of various downstream signaling
pathways, including mitogen-activated protein kinases (MAPKs) and
phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling
cascades (20). Therefore,
developing IGF-1R-targeted inhibitors to interfere with
IGF-1R-induced downstream signaling cascades has long been
recognized as a potentially effective treatment strategy for
various cancer types.
Over-activation of PI3K/Akt signaling cascades leads
to the activation or inactivation of numerous downstream
cytoplasmic and nuclear substrates, including forkhead box O3
(FoxO3α), which belongs to a family of transcription factors
(21). FoxO3α is a tumor suppressor
that inhibits cell growth and induces apoptosis by regulating gene
transcription and protein synthesis associated with tumorigenesis
and cancer progression (22,23).
Activation of PI3K/Akt signaling following growth factor
stimulation causes phosphorylation of FoxO3α, which leads to the
export from the nucleus to the cytoplasm, thereby inhibiting its
transcriptional activity and contributing to cell growth (22).
The present study aimed to demonstrate the
anticancer effects of PTL. By using in vitro and in
vivo analyses, the effects of PTL on the proliferation and
migration of human lung cancer cells were investigated and the
underlying molecular mechanisms were explored.
Materials and methods
Materials
Phospho-IGF-1R antibody (cat. no. 3024), IGF-1R
antibody (cat. no. 9750), phospho-Akt antibody (cat. no. 4060), Akt
antibody (cat. no. 4685), phospho-FoxO3α antibody (cat. no. 9465),
FoxO3α antibody (cat. no. 2497), MMP-2 antibody (cat. no. 87809),
MMP-9 antibody (cat. no. 13667), proliferating cell nuclear antigen
(PCNA) antibody (cat. no. 13110), β-actin antibody (cat. no. 4970)
and goat anti-rabbit horseradish peroxidase (HRP)-linked IgG
antibody (cat. no. 7074) were purchased from Cell Signaling
Technology, Inc. Ki67 antibody (cat. no. 27309-1-AP) was purchased
from Proteintech Group Inc. IGF-1, PTL and LY294002 (an inhibitor
of PI3K) were purchased from Sigma Aldrich; Merck KGaA. RPMI-1640
medium and fetal bovine serum (FBS) were purchased from Gibco
(Thermo Fisher Scientific, Inc.).
Cell culture
Human NSCLC cell lines A549 and H1299 cells were
purchased from the Cell Bank of the Chinese Academy of Sciences.
The cells were cultured in RPMI-1640 medium supplemented with 100
U/ml penicillin/streptomycin and 10% FBS and were maintained at
37°C in a humidified atmosphere containing 5% CO2.
Cell proliferation assay
A Cell Counting Kit-8 (CCK-8) assay was used to
assess the inhibitory effects of PTL. In brief, 3×103
cells/well were seeded into 96-well plates. After 24 h, the cells
were treated with PTL at various concentrations for the indicated
durations as specified in the Results. Following incubation, CCK-8
reagents were added and the cells were incubated for 1 h at room
temperature. Cell proliferation was determined by measuring the
optical density at 450 nm using an ELISA plate reader (Olympus
Corp.).
Wound healing assay
Cells were seeded in 6-well plates at a
concentration of 2×105 cells per well. Once they reached
80–90% confluence, the cells were serum-starved for 24 h and a
wound on the midline of each well was then created by scraping the
cell monolayer using a sterile 200-µl pipette tip. The cells were
then washed with PBS to rinse away any detached cells. Images of
the wound margins were captured. Subsequently, serum-free medium
containing the indicated drugs was added to each well. After
incubation for 48 h, images of the same area of the wound margins
as those prior to the application of PTL were captured. To analyze
the relative wound closure, the ratio between the unhealed wound
area at a given time point and the original wound area ×100% was
calculated using ImageJ software version 1.47t (National Institutes
of Health).
Colony formation assay
The colony formation assay was performed according
to the protocol of a previous study (24). In brief, 2×102 cells were
cultured in a 6-well plate and were treated with 10 µM PTL. The
medium containing PTL was replenished every three days. After a
2-week treatment, the cells were fixed with 4% paraformaldehyde and
then stained with 0.1% crystal violet for 10 min at room
temperature. The number of colonies containing >50 cells were
counted using ImageJ software version 1.47t (National Institutes of
Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
After treatment, the total RNA was isolated by using
TRIzol™ reagent (Thermo Fisher Scientific, Inc.). Reverse
transcription into cDNA was performed using the PrimeScript™ RT
Reagent Kit with gDNA Eraser (Perfect Real Time) according to the
manufacturer's instructions (Takara Biotechnology Co., Ltd.). qPCR
analysis was performed using TB Green™ Premix Ex Taq™ II (Tli
RNaseH Plus) (Takara Biotechnology Co., Ltd.) on the StepOnePlus™
Real-Time PCR System (Thermo Fisher Scientific, Inc.) to determine
the expression of matrix metalloproteinase (MMP)-2, MMP-9 and GAPDH
by using the following primers: MMP-2, 5′-CCGTCGCCCATCATCAAGTTC-3′
(sense) and 5′-GCAGCCATAGAAGGTGTTCAGG-3′ (antisense); MMP-9,
5′-TGGTCCTGGTGCTCCTGGTG-3′ (sense) and 5′-GCTGCCTGTCGGTGAGATTGG-3′
(antisense); GAPDH, 5′-ACAACTTTGGTATCGTGGAAGG-3′ (sense) and
5′-GCCATCACGCCACAGTTTC-3′ (antisense). The reaction volume
consisted of 20 µl, containing 2 µl template cDNA, 10 µl TB Green™
Premix Ex Taq™ II (Tli RNaseH Plus), 1 µl 10 µM each primer, and 6
µl ddH2O. The amplification program included 1 cycle of
95°C for 30 sec (initial denaturation), followed by 40 cycles of
95°C for 5 sec (denaturation) and 60°C for 30 sec (DNA synthesis).
The relative mRNA expression levels were analyzed using the
2−ΔΔCq method (25).
Immunofluorescence
Cells were cultured and treated with PTL in confocal
chambers. Then cells were then fixed with 4% paraformaldehyde for
15 min, followed by permeabilization with 0.25% Triton X-100 for 15
min at room temperature. Cells were then blocked with 1% bovine
serum albumin (Biosharp) and incubated with anti-Ki-67 antibody
overnight at 4°C. Alexa Fluor 568-conjugated secondary antibodies
(dilution 1:500; Thermo Fisher Scientific, Inc.) were applied for 2
h at room temperature, followed by nuclear counterstaining with
DAPI at room temperature. Images were captured using an inverted
microscope (magnification, ×200) with fluorescence filters (Olympus
Corp.).
Western blot analysis
After treatment, cells were lysed with SDS lysis
buffer (Beyotime Institute of Biotechnology) containing protease
inhibitor cocktail (Roche Diagnostics). Total protein was extracted
and the bicinchoninic acid protein assay (Beyotime Institute of
Biotechnology) was performed to determine the protein
concentration. Equal amounts of proteins (20 µg) were separated by
10% SDS-PAGE and transferred to nitrocellulose membranes (EMD
Millipore). After blocking with 5% non-fat dry milk for 1 h at room
temperature, the membranes were incubated with the following
primary antibodies overnight at 4°C: PNCA (dilution 1:2,000),
phospho-Akt (dilution 1:2,000), Akt (dilution 1:4,000),
phospho-IGF-1R (dilution 1:2,000), IGF-1R (dilution 1:2,000),
phospho-FoxO3α (dilution 1:2,000), FoxO3α (dilution 1:2,000), MMP-2
(dilution 1:2,000), MMP-9 (dilution 1:2,000) and β-actin (dilution
1:4,000). After antibody binding, the membranes were washed and
incubated with HRP-linked secondary antibodies (dilution 1:10,000)
for 2 h at room temperature. The labeled proteins were visualized
using enhanced chemiluminescence reagent (Pierce; Thermo Fisher
Scientific, Inc.) and signals were quantified by using ImageJ
software version 1.47t (National Institutes of Health).
Mouse subcutaneous xenografts and
treatments
Ten male BALB/c nude mice (age, 5-weeks; weight,
18±0.2 g) were obtained from Hunan Silaikejingda Laboratory Animal.
All animal procedures were in accordance with the protocols
approved by the Ethics Committee of the First Affiliated Hospital
of Nanchang University (Nanchang, China). A total of
5×106 in 100 µl A549 cells were subcutaneously injected
into the right flank of nude mice. After a 7-day growth (tumor size
was 250–300 mm3), the mice were randomly assigned to 2
groups (n=5 per group). The control group of mice was treated with
saline, while the experimental group was intraperitoneally injected
with PTL (10 mg/kg) daily for 24 days according to a previous study
(26). The mice were anesthetized
with 2% sodium pentobarbital (10 ml/kg) by intraperitoneal
injection, and then sacrificed by cervical dislocation to dissect
and weigh the tumors. The body weight was determined and the tumor
volume (V) was calculated every 4 days based on the following
formula: V=(the shortest diameter)2x(the longest
diameter) ×0.5.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean from at least three independent experiments. Statistical
analyses were performed using GraphPad Prism 6.0 (GraphPad
Software, Inc.) and comparisons were made by one/two-way analysis
of variance with Bonferroni's multiple-comparison tests or
Students' t-test. P<0.05 was considered to indicate statistical
significance.
Results
PTL inhibits the growth of lung cancer
cells
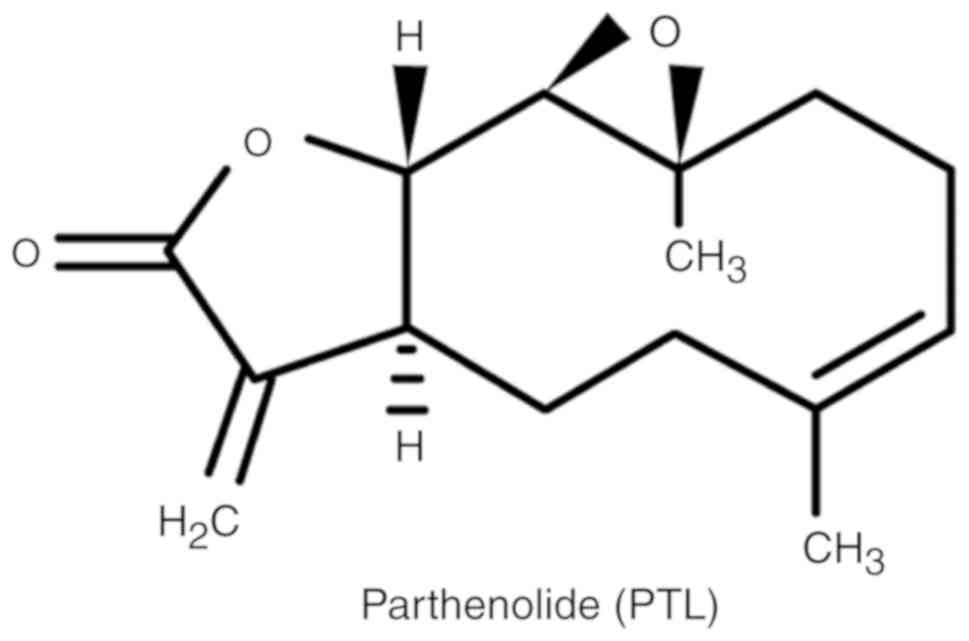
The chemical structure of PTL is presented in
Fig. 1. To determine the effect of
PLT on cancer cell proliferation, the human lung cancer cell lines
A549 and H1299 were used in the present study. The cells were
treated with increasing concentrations of PTL (0, 1, 5, 10, 20, 30
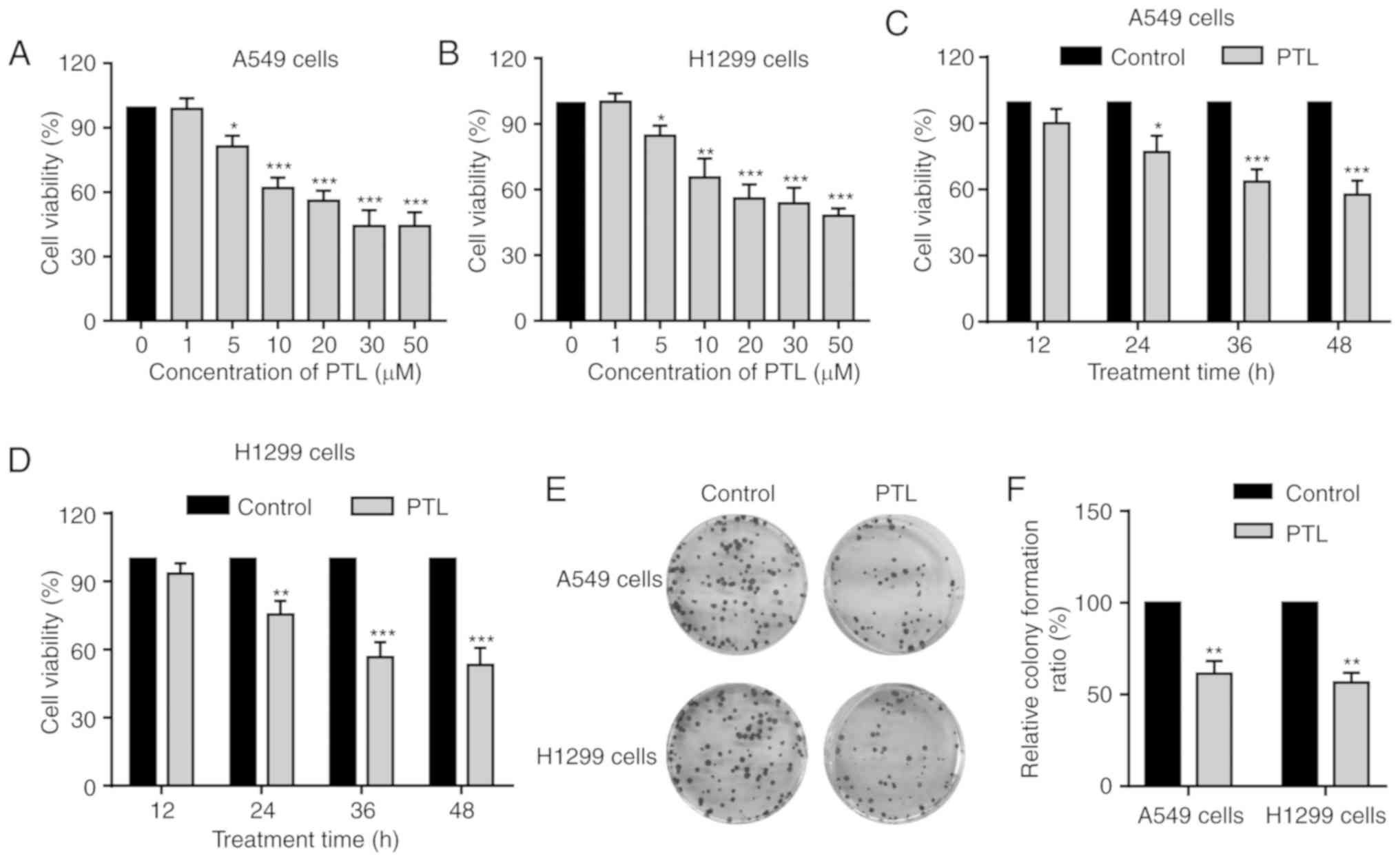
and 50 µM) for 48 h and then assessed by the CCK-8 assay. PTL
treatment decreased the cell viability of A549 and H1299 cells in a
dose-dependent manner (Fig. 2A and
B). Of note, at concentrations of <5 µM, PTL had no
noticeable effect, while significant inhibition was observed above
the concentration of 5 µM in both cell lines (Fig. 2A and B). Next, the impact of PTL
treatment for different durations on A549 and H1299 cell viability
was assessed. The cells were treated with 10 µM PTL for 12, 24, 36
or 48 h and the results indicated that PTL treatment for 24 h or
above significantly inhibited the growth of A549 and H1299 cells in
a time-dependent manner (Fig. 2C and
D).
To further confirm the growth inhibition effects of
PTL on lung cancer cells, a colony formation assay was performed.
Consistent with the results of the CCK-8 assay, PTL markedly
suppressed colony formation of A549 and H1299 cells and the number
of colonies was significantly reduced (Fig. 2E and F). These results suggest that
PTL exerts a significant growth inhibitory effect on lung cancer
cells.
PTL inhibits the expression of
proliferation markers in lung cancer cells
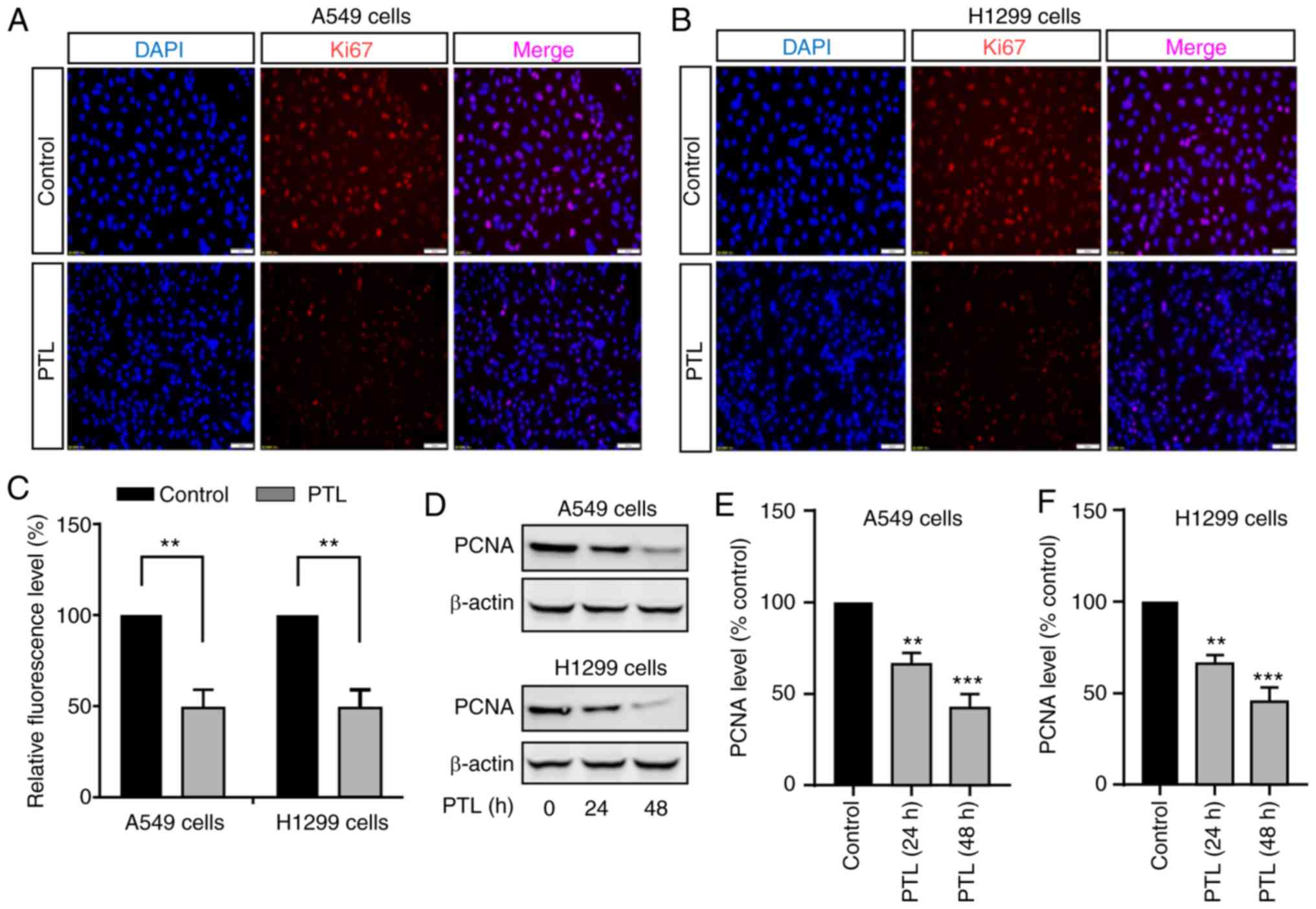
Nuclear antigen Ki67 has long been documented as one
of the most wildly used markers of proliferation in oncology
(27). Therefore, the PTL-mediated
proliferation inhibition was further investigated by detecting the
expression level of Ki67 in both lung cancer cell lines. An
immunostaining assay was performed to evaluate the effect of PTL on
cell proliferation (Fig. 3). PLT
significantly inhibited the expression of Ki67 in A549 cells
(Fig. 3A and C). Similar results
were observed in H1299 cells (Fig. 3B
and C). In addition, the expression of another cell
proliferation marker, namely proliferating cell nuclear antigen
(PCNA), was detected. PTL treatment for either 24 or 48 h
significantly reduced PCNA expression in both cell lines (Fig. 3D-F). Taken together, these results
suggest that PTL significantly inhibits the proliferation of lung
cancer cells.
PTL suppresses the migration of lung
cancer cells
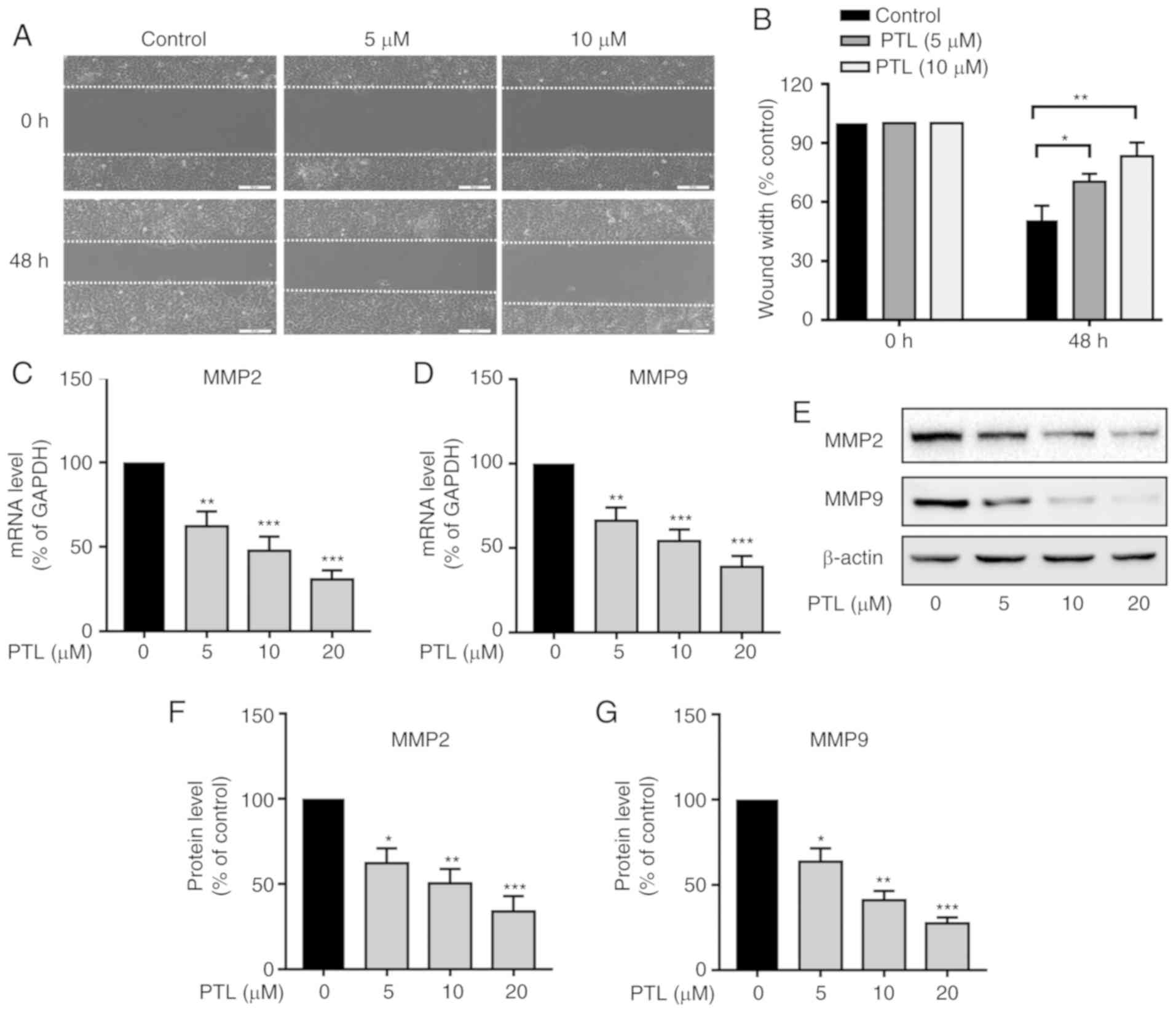
Since migration is a crucial factor that contributes
to cancer metastasis, the effect of PTL on the migration of A549
cells was then examined by a using wound-healing assay. As
presented in Fig. 4A, treatment
with PTL at 5 and 10 µM for 48 h significantly decreased the
migration ability of the A549 cells compared with that in the
control group (Fig. 4A and B).
Extracellular matrix (ECM) degradation by MMPs, such
as MMP-2 and MMP-9, plays a pivotal role in the migration and
metastasis of a variety of tumor types. It was therefore
investigated whether PTL reduces the expression of MMP-2 and MMP-9
in lung cancer cells. Compared to the control group, PTL treatment
for 48 h significantly downregulated the mRNA expression level of
MMP-2 (Fig. 4C) and MMP-9 (Fig. 4D) in a dose-dependent manner.
Furthermore, the protein levels of MMP-2 (Fig. 4E and F) and MMP-9 (Fig. 4E and G) were also significantly
inhibited by PTL treatment.
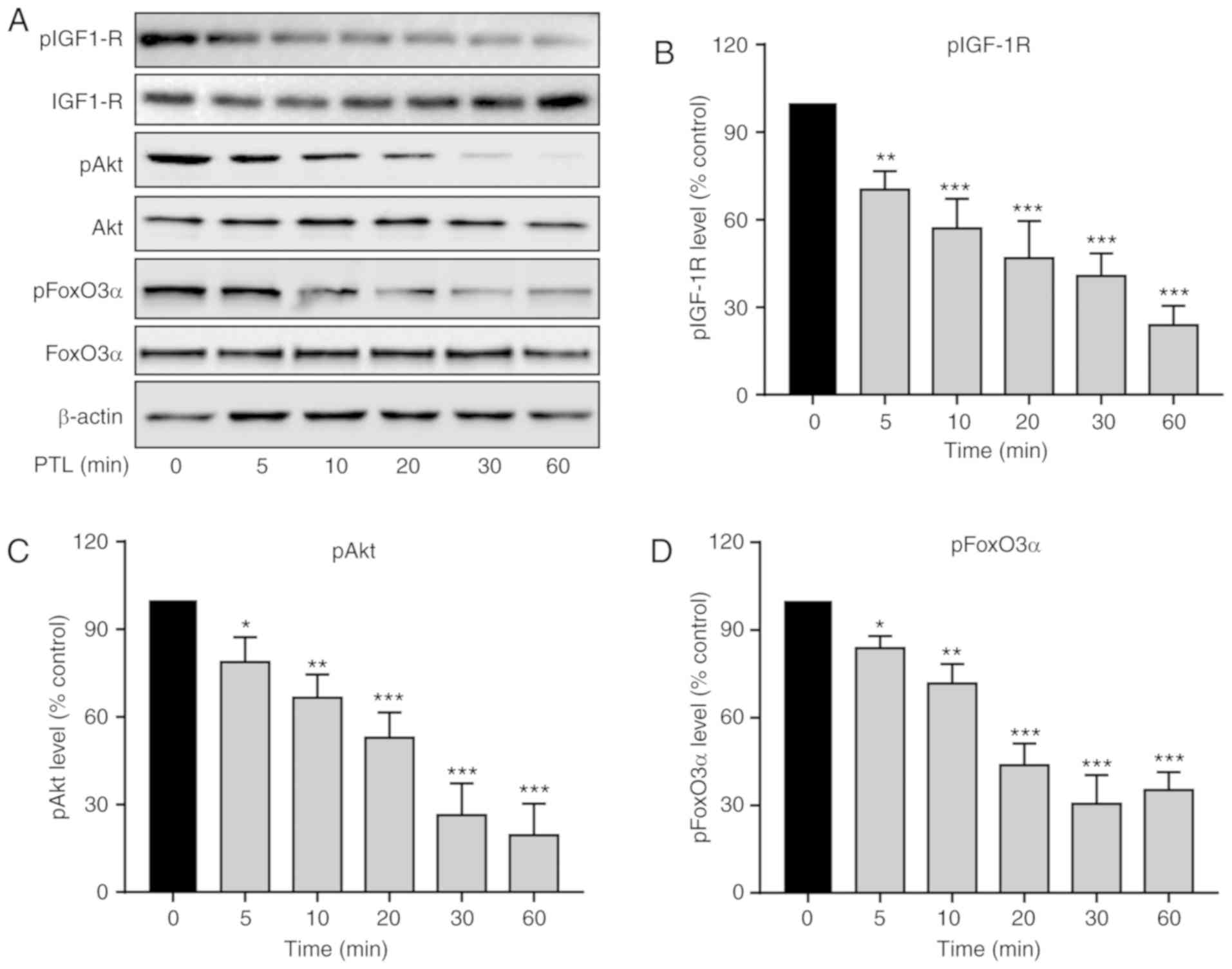
PTL inhibits the activation of the
IGF-1R/Akt/FoxO3α pathway in A549 cells
The IGF-1R-mediated PI3K/Akt signaling cascade has a
crucial role in the proliferation, survival, invasion and
angiogenesis of multiple cancers (28). First, the effect of PTL on IGF-1R
activation was explored. As presented in Fig. 5A, in comparison with the untreated
cells, PTL induced a reduction in the phosphorylation level of
IGF-1R (pIGF1-R), while it had no obvious effect on the total
expression level of IGF-1R (Fig.
5B).
The effects of PTL on Akt phosphorylation were then
assessed. Treatment of A549 cells with PTL significantly reduced
Akt phosphorylation (pAkt) that showing a similar trend with IGF-1R
(Fig. 5C). Furthermore, the effect
of PTL on the expression of FoxO3α, a transcription factor and
tumor suppressor, in A549 cells was determined. Of note, the
phosphorylation of FoxO3α (p FoxO3α) was also blocked by PTL
(Fig. 5D). These results suggest
that PTL significantly inhibited the IGF-1R/Akt/FoxO3α signaling
pathway in A549 cells.
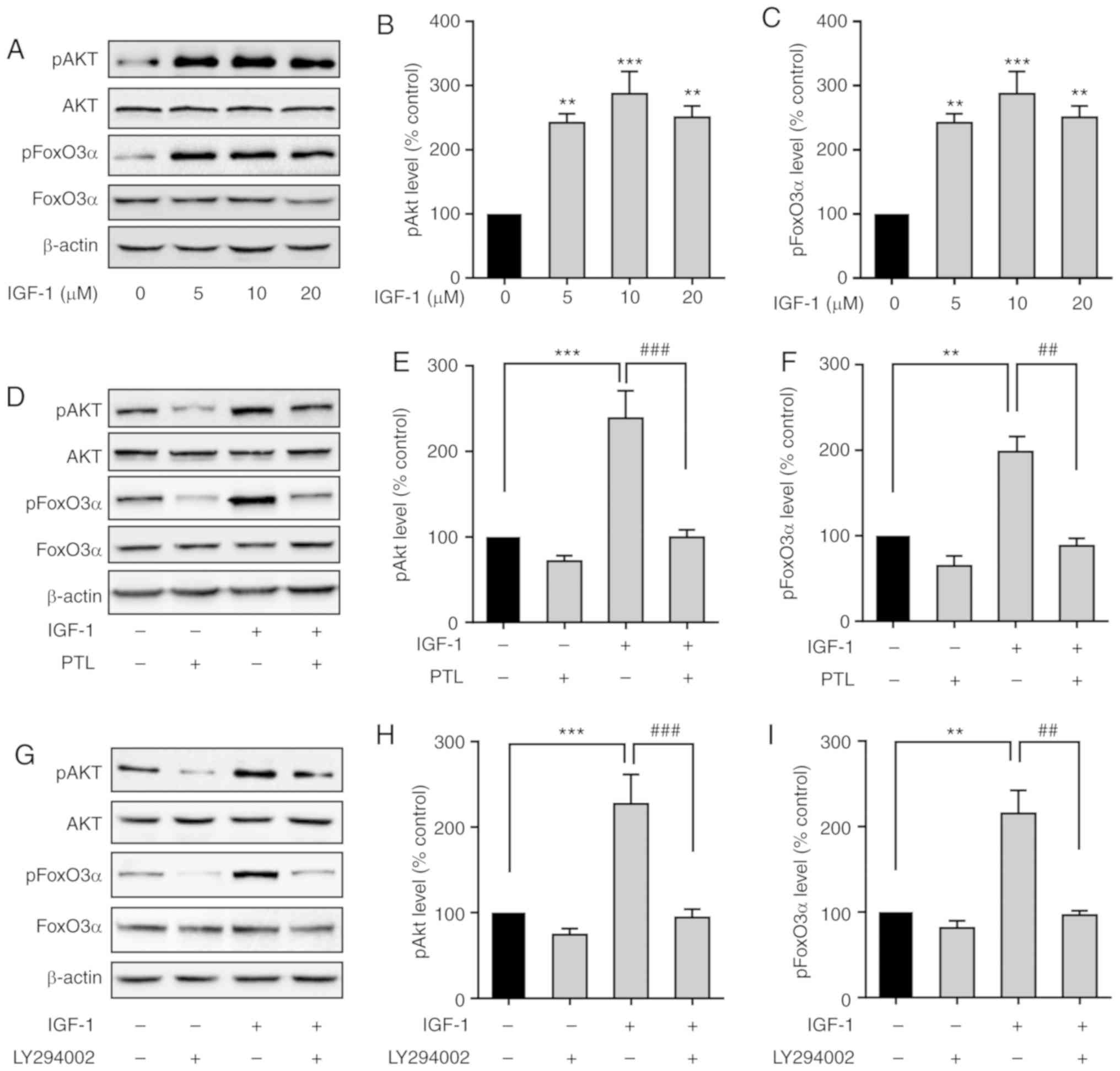
PTL inhibits IGF-1-induced activation
of the PI3K/Akt pathway in A549 cells
High circulating levels of IGF-1 are associated with
a high risk in numerous types of cancer (19). It was first confirmed that IGF-1
markedly increased Akt and FoxO3α phosphorylation (Fig. 6A-C). However, the increased
phosphorylation levels of Akt (pAkt) and FoxO3α (pFoxO3α) induced
by IGF-1 were significantly attenuated by PTL pre-treatment
(Fig. 6D-F).
To verify that Akt and FoxO3α phosphorylation was
mediated by IGF-1R in A549 cells, A549 cells were pretreated with
LY294002, an inhibitor of PI3K, for 30 min, and the levels of p-Akt
and p-FoxO3α induced by IGF-1 were then detected. The results
indicated that LY294002 markedly inhibited IGF-1-induced Akt and
FoxO3α phosphorylation (Fig. 6G-I),
further confirming that Akt and FoxO3α were downstream targets of
PI3K in A549 cells.
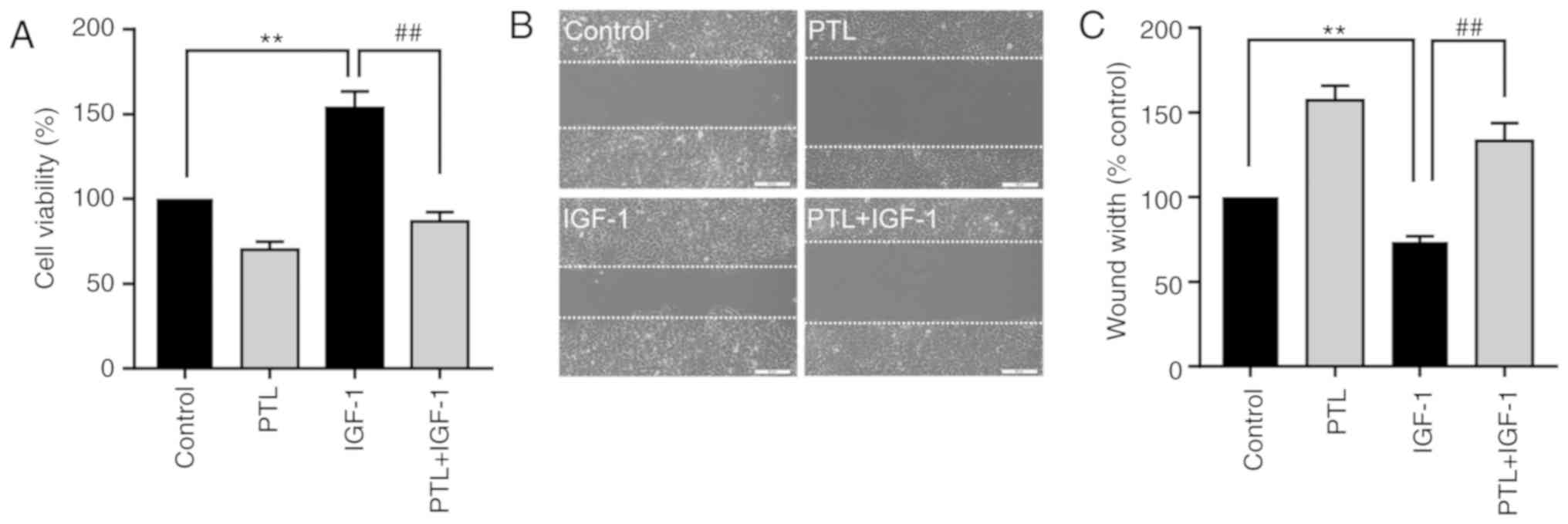
PTL inhibits IGF-1-induced A549 cell
proliferation and migration
The results revealing that PTL suppressed the cell
proliferation and migration, as well as inhibited the
phosphorylation of IGF-1R/PI3K/Akt/FoxO3α in A549 cells, strongly
suggest that the antitumor effect of PTL may be mediated by the
IGF-1R/PI3K/Akt pathway. Thus, the effect of PTL on IGF-1-induced
proliferation of A549 cells was then determined. As presented in
Fig. 7A, IGF-1 significantly
increased the proliferation of A549 cells, while pre-treatment with
PTL significantly attenuated the IGF-1-induced cell proliferation
(Fig. 7A). Similarly, PTL also
significantly inhibited IGF-1-induced cell migration ability
(Fig. 7B and C). Thus, the above
results indicate that PTL inhibits the proliferation and migration
of lung cancer cells by downregulating IGF-1R and its downstream
signaling cascades.
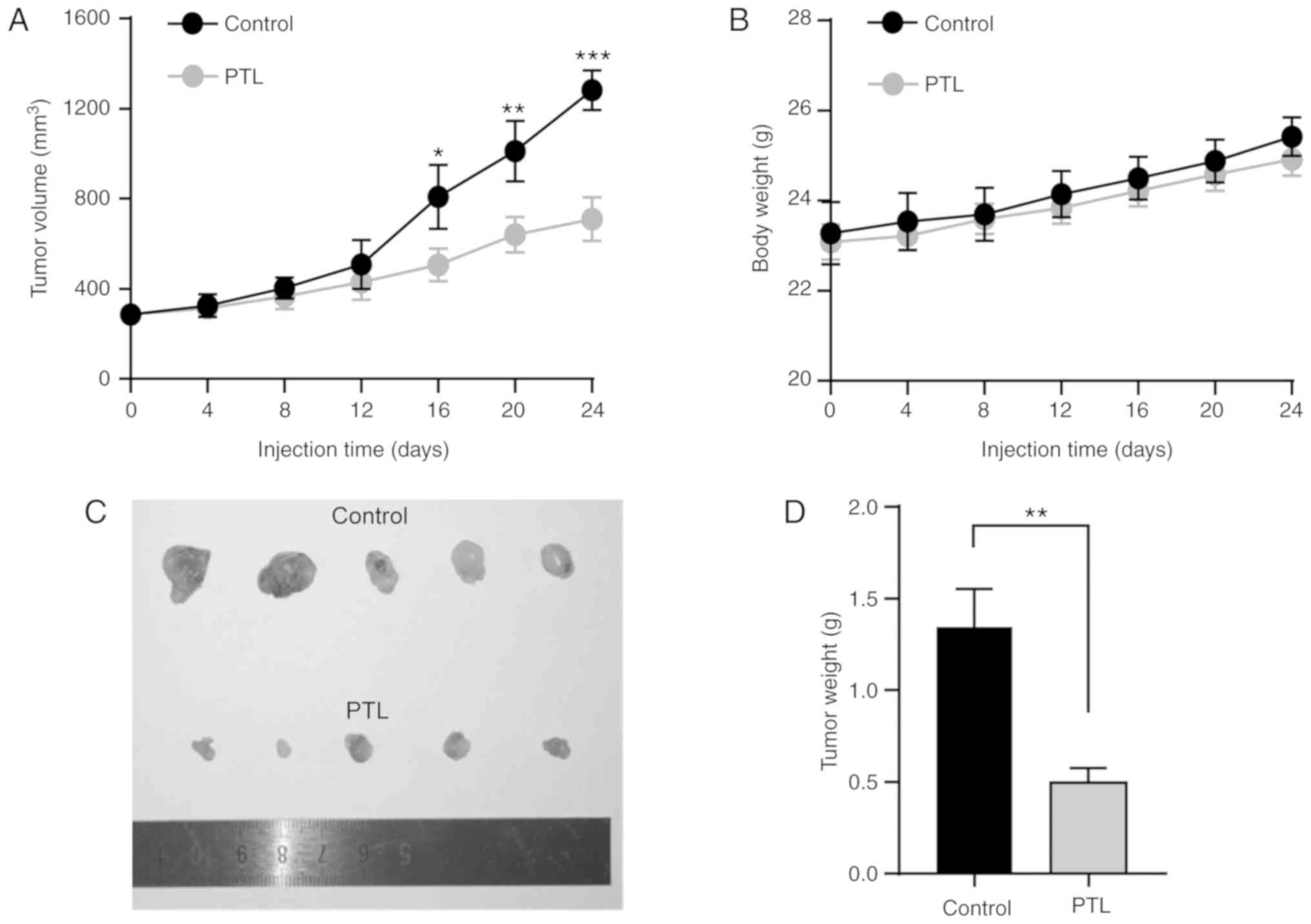
PTL inhibits the growth of lung cancer
in a mouse xenograft model
Considering the above in vitro results that
PTL suppressed the proliferation and migration of lung cancer
cells, the present study then set out to determine whether PTL is
able to inhibit the growth of lung cancer in vivo. Nude mice
were used to establish a lung cancer subcutaneous tumor xenograft
model. Compared with the control group, PTL treatment resulted in a
significant reduction in tumor growth and volume (Fig. 8A). During the entire treatment
period, no apparent alteration in body weight was observed between
the groups (Fig. 8B). Furthermore,
after treatment, the tumors were dissected from each mouse to
calculate the tumor weight in the different groups. The results
suggested that the tumor weight in PTL-treated mice was
significantly lower than that in the control group (Fig. 8C and D).
Discussion
In the present study, the anticancer effects of
parthenolide (PTL), a natural product extracted from the plant
Tanacetum parthenium (a flowering plant in the daisy family,
Asteraceae) on the proliferation and migration of lung cancer
cells, as well as the underlying molecular mechanisms, were
investigated. The results indicated that PTL reduced the expression
of matrix metalloproteinase (MMP)-2 and MMP-9, inhibited the
IGF-1R/PI3K/Akt/FoxO3α signaling pathway and consequently reduced
the migration and proliferation of lung cancer cells.
Natural plant extracts provide a variety of drug
candidates for cancer treatment. Accumulating evidence has
indicated that PTL exerts anticancer effects mainly through
interfering with the cancer progression process, including
suppression of the proliferation and migration, or induction of
cell apoptosis in various types of cancer cells (29,30).
The results of the present study indicated that PTL significantly
suppressed the proliferation of A549 and H1299 lung cancer cells.
Furthermore, molecular biology assays confirmed the
anti-proliferation effect of PTL on those lung cancer cells. Of
note, the expression of proliferating cell nuclear antigen (PCNA)
protein was significantly decreased after PTL treatment. Since PCNA
is found in the nucleus and has an essential role in numerous
cellular processes, including DNA replication and cell-cycle
progression (31), it has been
accepted that PCNA expression may be used as a potential indicator
for cancer prognosis (32). In
addition, the expression of Ki67, a widely used marker of tumor
aggressiveness and proliferation, was inhibited by PTL treatment.
Furthermore, in the mouse xenograft model of leukemia induced by
inoculating A549 cells into BALB/c mice, PTL significantly
inhibited the tumor growth in vivo, strongly suggesting that
PTL suppresses lung cancer growth.
To demonstrate the mechanism by which PTL regulates
lung cancer cell growth, the effects of PTL on insulin-like growth
factor 1 (IGF-1) signaling, which is fundamental for growth,
survival and migration, were evaluated. Increasing evidence
indicates that the IGF-1 receptor (IGF-1R) is abnormally expressed
in multiple cancer types (17), and
it has thus been suggested as a potential anticancer target
(33). Indeed, inhibitors targeting
IGF-1 signaling have been assessed in clinical cancer trials,
confirming IGF-1R as an effective therapeutic target (34). In addition, overexpression of IGF-1R
has been determined in lung cancer and high IGF-1R expression is
highly associated with an increased risk of developing malignant
tumors, as well as with a low survival rate of patients with NSCLC
after surgical resection (35).
Therefore, targeting the IGF-1/IGF-1R has been recognized as an
attractive strategy for the prevention and treatment of lung and
other types of cancer. The present study indicated that PTL is
effective in inhibiting IGF-1R phosphorylation. Furthermore, PTL
treatment also blocked IGF-1-induced IGF-1R activation, as well as
the proliferation of lung cancer cells. These results suggest that
PTL may inhibit the growth of lung cancer cells by inhibiting
IGF-1R signaling. However, it remains elusive how PTL acts to
suppress IGF-1R phosphorylation, which requires to be explored in
future research.
Accumulating evidence suggests that IGF-1R
activation leads to PI3K/Akt activation, which is mostly involved
in regulating the growth of cancer cells (29). In addition, clinical studies have
revealed that PI3K/Akt activation is associated with tumor
progression. Aberrant activation of the PI3K/Akt pathway confers
drug resistance in a variety of cancer types (36). Of note, a previous study has
demonstrated that PTL suppresses cell growth and induces apoptosis
through attenuating the PI3K/Akt signaling cascade in cervical
cancer cells (29). In the present
study, it was confirmed that in lung cancer cells, PTL suppressed
Akt phosphorylation and blocked IGF-1-induced Akt activation. In
line with this, IGF-1-induced proliferation of A549 cells was also
inhibited by PTL treatment, suggesting that PTL may be an
attractive candidate for lung cancer therapy. In addition, PTL
significantly inhibited the phosphorylation of transcription factor
FoxO3α. However, the exact mechanisms of how PTL-induced
de-phosphorylation of FoxO3α functions to suppress lung cancer cell
growth remain to be fully elucidated. Phosphorylation of FoxO3α
promotes its interaction with 14-3-3 protein, which ultimately
leads to nuclear export to the cytoplasm to inhibit the
transcriptional functions of FoxO3α thus facilitating cell growth
(37). By contrast, inhibition of
the phosphorylation of PI3K/Akt induces de-phosphorylation of
FoxO3α, leading to FoxO3α nuclear translocation to arrest cell
cycle progression and promote apoptosis. In future studies, it is
required to demonstrate whether PTL-induced de-phosphorylation of
FoxO3α is responsible for enhancing apoptosis of lung cancer
cells.
MMPs are calcium-dependent zinc-containing
endopeptidases that have essential roles in degrading ECM and
basement membranes. MMPs are widely known to be linked to the
metastatic ability of cancer cells (38). Overexpression of MMP-2 and MMP-9 is
frequently observed in tumor tissues and is highly associated with
tumor metastasis and aggressiveness of various cancer types
(39). Of note, the expression of
MMP-2 and MMP-9 is upregulated, which contributes to the migration
of A549 cancer cells during lung cancer progression (40). Consistent with this, the present
study demonstrated that PTL markedly inhibited MMP-2 and MMP-9
expression, as well as attenuated IGF-1-induced migration of A549
cells. Of note, upregulation of MMP-2 and MMP-9 has been reported
to be influenced by PI3K/Akt activation in numerous types of cancer
cell, including human melanoma (41) and A549 cells (42), and this was consistent with the
present result that PTL suppressed the PI3K/Akt pathway in A549
cells.
In summary, the present study demonstrated that PTL,
a natural product isolated from the medicinal plant Tanacetum
parthenium, not only inhibits the proliferation and migration
of lung cancer cells in vitro, but also suppresses lung
cancer growth in vivo. The anticancer effect of PTL may be
exerted by blocking IGF-1R-mediated PI3K/Akt signaling cascades,
including inhibition of FoxO3α activity. Furthermore, PTL
suppressed the migration of lung cancer cells, which involved
changes in the expression of migration-associated genes, including
MMP-2 and MMP-9. The present results suggest that PTL may be a
potential candidate for lung cancer treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81960523,
81460444 and 81660391).
Availability of data and materials
The datasets used during the current study are
available from the corresponding author upon reasonable
request.
Authors' contributions
LS and WY conducted most of the experiments. GW, BY
and FX analyzed the data. XG, JT and QZ performed the animal
experiments. LZ and CC performed the immunofluorescence
experiments. WZ conceived and designed the study. LS, WY and WZ
wrote the manuscript. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
All animal experimentation was approved by the
Ethics Committee of the First Affiliated Hospital of Nanchang
University (Nanchang, Jiangxi, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PTL
|
parthenolide
|
|
NSCLC
|
non-small cell lung cancer
|
|
IGF-1
|
insulin-like growth factor 1
|
|
IGF-1R
|
insulin-like growth factor 1
receptor
|
|
PI3K
|
phosphoinositide 3-kinase
|
References
|
1
|
Bittoni MA, Focht BC, Clinton SK,
Buckworth J and Harris RE: Prospective evaluation of C-reactive
protein, smoking and lung cancer death in the Third National Health
and Nutrition Examination Survey. Int J Oncol. 47:1537–1544. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nawaz K and Webster RM: The non-small-cell
lung cancer drug market. Nat Rev Drug Discov. 15:229–230. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Di X, Jin X, Li R, Zhao M and Wang K:
CircRNAs and lung cancer: Biomarkers and master regulators. Life
Sci. 220:177–185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Youlden DR, Cramb SM and Baade PD: The
international epidemiology of lung cancer: Geographical
distribution and secular trends. J Thorac Oncol. 3:819–831. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li LC and Kan LD: Traditional Chinese
medicine for pulmonary fibrosis therapy: Progress and future
prospects. J Ethnopharmacol. 198:45–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Efferth T, Li PC, Konkimalla VS and Kaina
B: From traditional Chinese medicine to rational cancer therapy.
Trends Mol Med. 13:353–361. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu D, Chen L, Zhao H, Vaziri ND, Ma SC
and Zhao YY: Small molecules from natural products targeting the
Wnt/β-catenin pathway as a therapeutic strategy. Biomed
Pharmacother. 117:1089902019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ghantous A, Sinjab A, Herceg Z and
Darwiche N: Parthenolide: From plant shoots to cancer roots. Drug
Discov Today. 18:894–905. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang S, Ong CN and Shen HM: Critical
roles of intracellular thiols and calcium in parthenolide-induced
apoptosis in human colorectal cancer cells. Cancer Lett.
208:143–153. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu YC, Kim SL, Park YR, Lee ST and Kim
SW: Parthenolide promotes apoptotic cell death and inhibits the
migration and invasion of SW620 cells. Intest Res. 15:174–181.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu JW, Cai MX, Xin Y, Wu QS, Ma J, Yang
P, Xie HY and Huang DS: Parthenolide induces proliferation
inhibition and apoptosis of pancreatic cancer cells in vitro. J Exp
Clin Cancer Res. 29:1082010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wyrębska A, Szymański J, Gach K, Piekielna
J, Koszuk J, Janecki T and Janecka A: Apoptosis-mediated cytotoxic
effects of parthenolide and the new synthetic analog MZ-6 on two
breast cancer cell lines. Mol Biol Rep. 40:1655–1663. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jafari N, Nazeri S and Enferadi ST:
Parthenolide reduces metastasis by inhibition of vimentin
expression and induces apoptosis by suppression elongation factor
α-1 expression. Phytomedicine. 41:67–73. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Talib WH and Al Kury LT: Parthenolide
inhibits tumor- promoting effects of nicotine in lung cancer by
inducing P53-dependent apoptosis and inhibiting VEGF expression.
Biomed Pharmacother. 107:1488–1495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin M, Bi H, Yan Y, Huang W, Zhang G,
Zhang G, Tang S, Liu Y, Zhang L, Ma J and Zhang J: Parthenolide
suppresses non-small cell lung cancer GLC-82 cells growth via
B-Raf/MAPK/Erk pathway. Oncotarget. 8:23436–23447. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Regad T: Targeting RTK signaling pathways
in cancer. Cancers (Basel). 7:1758–1784. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Denduluri SK, Idowu O, Wang Z, Liao Z, Yan
Z, Mohammed MK, Ye J, Wei Q, Wang J, Zhao L and Luu HH:
Insulin-like growth factor (IGF) signaling in tumorigenesis and the
development of cancer drug resistance. Genes Dis. 2:13–25. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Casa AJ, Dearth RK, Litzenburger BC, Lee
AV and Cui X: The type I insulin-like growth factor receptor
pathway: A key player in cancer therapeutic resistance. Front
Biosci. 13:3273–3287. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weroha SJ and Haluska P: The insulin-like
growth factor system in cancer. Endocrinol Metab Clin North Am.
41335–350. (vi)2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mayer IA and Arteaga CL: The PI3K/AKT
pathway as a target for cancer Treatment. Annu Rev Med. 67:11–28.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Farhan M, Wang H, Gaur U, Little PJ, Xu J
and Zheng W: FOXO signaling pathways as therapeutic targets in
cancer. Int J Biol Sci. 13:815–827. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coomans de Brachène A and Demoulin JB:
FOXO transcription factors in cancer development and therapy. Cell
Mol Life Sci. 73:1159–1172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hornsveld M, Dansen TB, Derksen PW and
Burgering BMT: Re-evaluating the role of FOXOs in cancer. Semin
Cancer Biol. 50:90–100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang J, Wen G, Sun L, Yuan W, Wang R,
Zeng Q, Zhang G and Yu B: Cryptotanshinone inhibits cellular
proliferation of human lung cancer cells through downregulation of
IGF-1R/PI3K/Akt signaling pathway. Oncol Rep. 40:2926–2934.
2018.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nakabayashi H and Shimizu K: Involvement
of Akt/NF-κB pathway in antitumor effects of parthenolide on
glioblastoma cells in vitro and in vivo. BMC Cancer. 12:4532012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jakobsen JN and Sørensen JB: Clinical
impact of ki-67 labeling index in non-small cell lung cancer. Lung
Cancer. 79:1–7. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Altorki NK, Markowitz GJ, Gao D, Port JL,
Saxena A, Stiles B, McGraw T and Mittal V: The lung
microenvironment: An important regulator of tumour growth and
metastasis. Nat Rev Cancer. 19:9–31. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jeyamohan S, Moorthy RK, Kannan MK and
Arockiam AJV: Parthenolide induces apoptosis and autophagy through
the suppression of PI3K/Akt signaling pathway in cervical cancer.
Biotechnol Lett. 38:1251–1260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu HJ, Jung JY, Jeong JH, Cho SD and Lee
JS: Induction of apoptosis by parthenolide in human oral cancer
cell lines and tumor xenografts. Oral Oncol. 51:602–609. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stoimenov I and Helleday T: PCNA on the
crossroad of cancer. Biochem Soc Trans. 37:7(Pt 3). 605–613. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Juríková M, Danihel L, Polák Š and Varga
I: Ki67, PCNA, and MCM proteins: Markers of proliferation in the
diagnosis of breast cancer. Acta Histochem. 118:544–552. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peled N, Wynes MW, Ikeda N, Ohira T,
Yoshida K, Qian J, Ilouze M, Brenner R, Kato Y, Mascaux C and
Hirsch FR: Insulin-like growth factor-1 receptor (IGF-1R) as a
biomarker for resistance to the tyrosine kinase inhibitor gefitinib
in non-small cell lung cancer. Cell Oncol (Dordr). 36:277–288.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Singh P, Alex JM and Bast F: Insulin
receptor (IR) and insulin-like growth factor receptor 1 (IGF-1R)
signaling systems: Novel treatment strategies for cancer. Med
Oncol. 31:8052014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gong Y, Yao E, Shen R, Goel A, Arcila M,
Teruya-Feldstein J, Zakowski MF, Frankel S, Peifer M, Thomas RK, et
al: High expression levels of total IGF-1R and sensitivity of NSCLC
cells in vitro to an anti-IGF-1R antibody (R1507). PLoS One.
4:e72732009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang J, Nie J, Ma X, Wei Y, Peng Y and Wei
X: Targeting PI3K in cancer: Mechanisms and advances in clinical
trials. Mol Cancer. 18:262019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tzivion G, Dobson M and Ramakrishnan G:
FoxO transcription factors; Regulation by AKT and 14-3-3 proteins.
Biochim Biophys Acta. 1813:1938–1945. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Piperigkou Z, Manou D, Karamanou K and
Theocharis AD: Strategies to target matrix metalloproteinases as
therapeutic approach in cancer. Methods Mol Biol. 1731:325–348.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cathcart J, Pulkoski-Gross A and Cao J:
Targeting matrix metalloproteinases in cancer: Bringing new life to
old ideas. Genes Dis. 2:26–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hu P, He J, Liu S, Wang M, Pan B and Zhang
W: β2-adrenergic receptor activation promotes the proliferation of
A549 lung cancer cells via the ERK1/2/CREB pathway. Oncol Rep.
36:1757–1763. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yao X, Jiang W, Yu D and Yan Z: Luteolin
inhibits proliferation and induces apoptosis of human melanoma
cells in vivo and in vitro by suppressing MMP-2 and MMP-9 through
the PI3K/AKT pathway. Food Funct. 10:703–712. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gao Y, Guan Z, Chen J, Xie H, Yang Z, Fan
J, Wang X and Li L: CXCL5/CXCR2 axis promotes bladder cancer cell
migration and invasion by activating PI3K/AKT-induced upregulation
of MMP2/MMP9. Int J Oncol. 47:690–700. 2015. View Article : Google Scholar : PubMed/NCBI
|