Introduction
Ovarian cancer (OVCA) is one of the leading causes
of cancer-associated deaths among women (1,2).
According to the World Health Organization, OVCA accounts for
150,000 deaths annually worldwide (3). Therefore, there is an urgent need to
better understand the molecular mechanisms of tumor growth and
development of OVCA.
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is
a key regulatory enzyme, which catalyzes the oxidative
phosphorylation of glycerol-3-phosphate during glycolysis. It is
one of the most extensively investigated housekeeping genes, and is
widely used as an internal control for the analysis of gene
expression levels. However, mounting evidence now recognizes GAPDH
as a multifunctional protein involved in a range of biological
processes, including membrane fusion and transport (4,5),
apoptosis (6), DNA repair (7), DNA replication and regulation of
transcription and translation (8).
GAPDH expression is modulated by serum, epidermal growth factor and
retinoic acid (9,10). In addition, it has been reported
that miRNA-mediated regulation is partly responsible for the
variability in GAPDH expression (11). Recent studies have demonstrated that
GAPDH expression is significantly upregulated in several types of
cancer, such as renal (12,13), breast (14) and nasopharyngeal cancer (15). A dysregulation of GAPDH expression
may potentially be associated with tumorigenesis and
malignancy.
Recently, studies have suggested that the genomes of
the majority of cancers in adults contain thousands of somatic
mutations (16), and mutations in
the 3′untranslated region (3′UTR) may mediate oncogene activation
or tumor suppressor inactivation by altering microRNA
(miRNA/miR)-binding efficiency (17,18).
miRNAs are small, endogenous and non-coding RNAs that negatively
regulate gene expression at the post-transcriptional level, by
binding to the 3′UTR of their target mRNAs (19). It has also been reported that mRNAs,
long non-coding RNAs (lncRNAs), and transcribed pseudogenes can act
as competitive endogenous RNAs (ceRNAs) which co-regulate each
other's expression by competing for shared microRNAs (20,21),
and the 3′UTR of mRNA alone can act as a ceRNA (22–25).
Dysregulation of the expression of various miRNAs has been
identified in several types of cancer, and have been demonstrated
to contribute to the malignancy and tumorigenesis of cancer
(26–28).
In the present study, it was demonstrated that the
expression of GAPDH mRNA was upregulated in OVCA tissues, and a
somatic mutation in the 3′UTR of GAPDH was identified, which was
located in the seed sequence of miR-125b. The mutant GAPDH 3′UTR
promoted tumor growth and cell motility by directly binding to
miR-125b, thus upregulating the expression of the miR-125b target
gene, STAT3, in OVCA.
Materials and methods
Clinical tissue specimens and cell
lines
All experimental protocols in the present study were
approved by The Third Affiliated Hospital of Guangzhou Medical
University (Guangzhou, Guangdong, China). Between January 2015 and
2019, a total of 120 patients who were pathologically diagnosed
with OVCA were enrolled in this study. The inclusion criteria were
as follows: OVCA patients who were scheduled for cytoreductive
surgery; patients aged above 18 years. Any patient who was in
pregnancy, or who was diagnosed with a malignant tumor of another
organ and tissue was excluded. The clinicopathological
characteristics of these 120 OVCA patients are documented in
Table I. All patients included in
this study provided written informed consent prior to surgery. The
cell lines used in this study were, SKOV3, OVCAR3 and 293T cells,
and were all obtained from the American Type Culture Collection.
293T cells were cultured in DMEM (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.) in
a humidified incubator with 5% CO2 at 37°C. SKOV3 and
OVCAR3 cells were cultured in RPMI-1640 medium with 10% FBS under
the same incubation conditions.
 | Table I.Clinicopathogical characteristics of
the OVCA patients (N=120). |
Table I.
Clinicopathogical characteristics of
the OVCA patients (N=120).
| Clinicopathological
characteristics | Data |
|---|
| Age (years) |
|
| Median
age (years) | 54 |
| ≥60, n
(%) | 54 (45.00) |
| <60,
n (%) | 66 (55.00) |
| BMI,
kg/m2 |
|
| Mean
(range) | 21.34
(18.9-28.6) |
| FIGO stage, n
(%) |
|
|
I–II | 74 (61.67) |
|
III–IV | 46 (38.33) |
| Histological type,
n (%) |
|
|
Serous | 71 (59.17) |
| Clear
cell | 12 (10.00) |
|
Mucinous | 17 (14.16) |
|
Endometrioid | 20 (16.67) |
Transcriptome library preparation and
sequencing
Total RNA was extracted from cancer and normal
tissues using TRIzol® (Tiangen Biotech Co., Ltd.). RNA
degradation and contamination were assessed by running a sample on
a 1% agarose gel. RNA purity and concentration were detected using
a NanoPhotometer® spectrophotometer (Implen GmbH) and a
Qubit® RNA Assay kit with a Qubit® 2.0
Flurometer (Thermo Fisher Scientific, Inc.), respectively. RNA
integrity was assessed using an RNA Nano 6000 Assay kit with a
Bioanalyzer 2100 system (Agilent Technologies, Inc.). Subsequently,
a total of 3 µg of RNA from each sample was used to construct the
Illumina sequencing library with a NEBNext® Ultra™ RNA
Library Prep kit for Illumina® (New England BioLabs,
Inc.). The index codes were added to attribute sequences to each
sample using a TruSeq PE Cluster kit v3-cBot-HS (Illumina, Inc.),
and the prepared libraries were sequenced using an Illumina
Hiseq® 2500 platform (Illumina, Inc.).
Generation of a stable cell line
The primer sequences for the amplification of GAPDH
3′UTR-WT or the mutant (CCTAG to CCTCAG) were as follows: GAPDH
3′UTR forward, 5′-CCAGCAAGAGCACAAGAGGA-3′ and reverse,
5′-TGGTTGAGCACAGGGTACTT-3′. GAPDH 3′UTR-MUT (CCT AG to CCTCAG) and
control DNA fragments were cloned into a pLKO.1-EGFP lentivirus
vector. 293T cells were used for generation of lentiviral particles
by transfecting the expression vectors and package vectors with
PEI. A total of 72 h after transfection, supernatants containing
the lentiviral particles were harvested, and the remaining cells
were removed by concentration. For lentiviral infection, SKOV3 and
OVCAR3 cells were seeded 1 day prior to infection, and cells were
infected with a multiplicity of infection (MOI) of 20 and 30,
respectively. The lentivirus was diluted in RPMI-1640 medium
containing Polybrene (10 µg/ml) and added to the cells for 24 h at
37°C. Subsequently, the virus-containing medium was replaced with
fresh supplemented RPMI-1640 medium, and the successfully infected
cells were isolated using FACS based on eGFP fluorescence.
RNA transfection
miR-125b mimics, Ago2 small interfering (si)RNA and
their corresponding negative controls (NCs) were purchased form
Shanghai GenePharma Co., Ltd. Transfection was performed using
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.).
miR-125b mimics, Ago2 siRNA and NC at a final concentration of 50
nM were used for transfection. The sequences of the Ago2 siRNAs
used were as follows: Ago2-siRNA-1, 5′-GCCAGTAATCGAGTTTGTT-3′;
Ago2-siRNA-2, 5′-GGTACCACCTGGTGGATAA-3′; and Ago2-siRNA-3,
5′-GCAGAAACACACCTACCTT-3′.
RNA extraction and reverse
transcription-quantitative PCR
TRIzol® reagent (Tiangen Biotech Co.,
Ltd.) was used to isolate total RNA. A total of 2 µg of each RNA
sample was reverse transcribed into cDNA using FastQuant RT Super
mix (Tiangen Biotech Co., Ltd.). qPCR was performed on a Bio-Rad
MiniOpticon Real-Time PCR system (Bio-Rad Laboratories, Inc.) using
a SYBR Green PCR Master mix (Tiangen Biotech Co., Ltd.). A
Bugle-Loop™ miRNA qPCR kit (Guangzhou RiboBio Co., Ltd.) was used
to quantify miR-125b and U6. Hsa-mir-125b-1_1_PR miScript Precursor
Assay (Qiagen GmbH) was used for quantification of precursor
miR-125b. The primer sequences were as follows: GAPDH forward,
5′-AGCCACAATCGCTCAGACAC-3′ and reverse, 5′-GCCCAATACGACCAAATCC-3′;
Ago2 forward, 5′-CCTCCCATGTTTACAAGTCG-3′ and reverse,
5′-TCTTTGTCCTGCCACAATG-3′; STAT3 forward,
5′-CATATGCGGCCAGCAAAGAA-3′ and reverse, 5′-ATACCTGCTCTGAAGAAACT-3′;
GUSB forward, 5′-AGCGTGGAGCAAGACAGTG-3′ and reverse,
5′-TCTGCATAGGGGTAGTGGCT-3′; and Pri-miR-125b forward,
5′-TGAACCTCGAACAGAAATTGCC-3′ and reverse,
5′-TCCACCAAATTTCCAGGATGC-3′.
Western blotting
Total cellular proteins were solubilized in RIPA
lysis buffer containing protease inhibitor cocktail (Sigma-Aldrich;
Merck KGaA) and 1 mM PMSF. Proteins were loaded on a 10% SDS gel,
resolved using SDS-PAGE and subsequently transferred
electrophoretically to a PVDF membrane (EMD Millipore). The
membranes were blocked in 5% non-fat-dried milk at room temperature
for 30–60 min, and subsequently incubated with primary antibodies
overnight at 4°C at the following dilutions: GAPDH (Santa Cruz
Biotechnology, Inc.; cat. no. sc-25778) at 1:1,000, Ago2 (Abcam;
cat. no. ab57113) at 1:2,000, STAT3 (Santa Cruz Biotechnology, Inc.
cat. no. sc-8019) at 1:300, anti-human β-actin (Cell Signaling
Technology, Inc.; cat. no. 4970) at 1:1,000. Subsequently, the
membranes were washed and incubated for 1 h at room temperature
with horseradish peroxidase-conjugated secondary antibodies (Cell
Signaling Technology, Inc.; cat. no. 7074) at a dilution of
1:2,000. Following several washes, signals were visualized using an
ECL system (Tiangen Biotech Co., Ltd.). β-actin was used as the
loading control. Western blot protein grayscale values were
detected and analyzed by ImageJ (1.48V; NIH).
Luciferase assay
For the luciferase assays, the WT and mutant GAPDH
3′UTR, and the WT and mutant (CTCAGGGAtoCTACTAGA) STAT3 3′UTR were
amplified by PCR. The primer sequences for the amplification of
STAT3 3′UTR-WT and STAT3 3′UTR-MUT were as follows: STAT3 3′UTR
forward, 5′-ACCTGGCCTTTGGTGTTGAA-3′ and reverse:
5′-GCTGAGGCAAGGTGGTTTTG-3′. The DNA fragments were cloned into a
pGL3 plasmid 3′ to the firefly luciferase gene. A total of
4×104 293T cells were co-transfected with 200 ng of
microRNA mimics, 200 ng of one of the pGL3 firefly luciferase
constructs and 20 ng of pGL3 Renilla luciferase construct,
which was used as the normalization control. After 48 h, cells were
lysed and luciferase activity was measured using a Dual-Luciferase
Reporter Assay System (Promega Corporation), according to the
manufacturer's protocol.
Cell Counting Kit-8 (CCK-8) assay
Cells were seeded in 96-well plates at a density of
2×103 cells/well in 100 µl culture medium. A total of 10
µl CCK-8 reagent (Beijing Transgen Biotech Co., Ltd.) was added at
0, 12, 24, 36, 48, 60 or 72 h. The plates were further cultured for
2 h. Cell viability was assessed by measuring the optical density
(OD) absorbance at a wavelength of 450 nm using a microplate reader
(BioTek Instruments, Inc.).
Wound healing assays
Cells were cultured to 80% confluence in 6-well
culture plates and a wound was created using a 200-µl pipette tip.
Subsequently, the cells were washed once with fresh medium to
remove the non-adherent cells. Wounds were imaged at 0 and 24 h and
the width of scratch were measured using ImageJ Software (1.48V;
NIH).
Cell migration and invasion
assays
Cells were resuspended in 200 µl serum-free
RPMI-1640 medium and were plated onto either a BD BioCoat™
Matrigel™ Invasion Chamber (BD Biosciences) for invasion assays, or
onto a BD Falcon (uncoated) Chamber for migration assays. For both
assays, supplemented medium containing 10% FBS was added to the
lower chamber. The chambers were processed after 24 h according to
the manufacturer's protocol, and migrated cells were stained with
Hoechst at room temperature for 10 min. Cells were examined using a
fluorescence microscope with 20× objective (Nikon Corporation).
microRNA stability analysis
SKOV3 cells were transfected with control WT or
mutant GAPDH 3′UTR constructs for 24 h. Subsequently, actinomycin-D
was added and cells were further cultured for 4, 8, 12, 16, 20 and
24 h. Total RNA was extracted using TRIzol reagent (Tiangen Biotech
Co., Ltd.). The expression of miR-125b was detected using
RT-qPCR.
RNA binding immunoprecipitation
Cells were lysed for at least 20 min on ice. The
cell lysates were collected following centrifugation (12,000 × g
for 10 min). Cell lysates were incubated with an anti-Ago2 antibody
(Abcam; cat. no. ab57113) at a dilution of 1:200, or isotype
control IgG (Sigma-Aldrich: Merck KGaA) at 4°C for 2 h. The
RNA-protein immunocomplexes were enriched using protein A/G Plus
agarose beads. Subsequently, the beads were washed and the
complexes were treated with DNase I and Proteinase K. RT-qPCR was
performed to assess the RNA isolated from the immunoprecipitation
material.
Statistical analysis
Data are presented as the mean ± standard error of
the mean of the indicated number of measurements. The differences
between groups were analyzed by Student's t-test. Comparisons
between multiple groups were performed using one-way ANOVA analysis
of variance followed by the Tukey's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of a frequent somatic
mutation in the 3′UTR of GAPDH
Transcriptome sequencing was performed on the OVCA
samples and their matching adjacent normal tissues from 120
patients to analyze the presence and frequency of mutations of
housekeeping genes. A summary of the top 10 housekeeping genes with
a high mutation rate in the UTR is shown in Fig. 1A. The gene with the highest
frequency of non-coding mutations in the UTR was GAPDH, which
harbored 6 mutations in 22/120 (18.3%) OVCA tissues (Table SI). Additionally, a somatic
mutation located in GAPDH 3′UTR (NM_001289745.2:c.*1325_*1326insC)
occurred most frequently (9.17%). In contrast, the matching
adjacent normal tissues did not contain this somatic mutation.
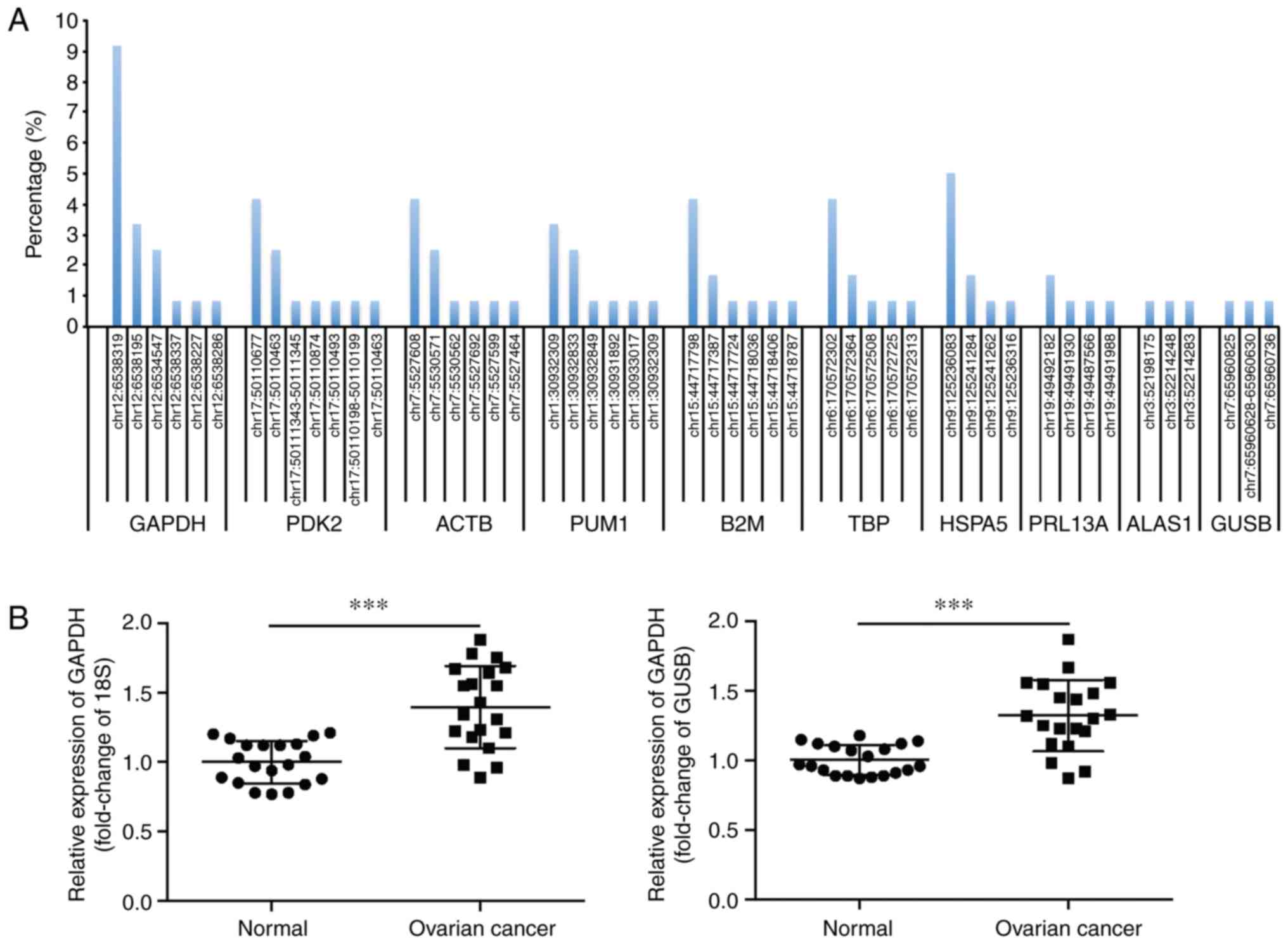 | Figure 1.Identification of a frequent somatic
mutation in the 3′UTR of GAPDH. (A) The top 10 housekeeping genes
with high mutation rates in the UTR, as determined by transcriptome
sequencing of 120 pairs of human OVCA tissues and the matching
adjacent normal tissues. (B) Relative mRNA expression levels of
GAPDH in 20 pairs of OVCA tissues and the matching adjacent normal
tissues, with 18S or GUSB as the internal control, respectively.
***P<0.001. UTR, untranslated region; OVCA, ovarian cancer;
GAPDH, glyceraldehyde 3-phosphate dehydrogenase, PDK2, pyruvate
dehydrogenase kinase 2; ACTB, β actin; PUM1, Pumilio RNA binding
family member 1; B2M, β-2-microglobulin; TBP, TATA-box binding
protein; HSPA5, heat shock protein family A (Hsp70) member 5;
RPL13A, ribosomal protein L13a; ALAS1, 5′-aminolevulinate synthase
1; GUSB, glucuronidase β. |
The expression of GAPDH in OVCA tissues was assessed
using RT-qPCR. As 18S and GUSB are reliable reference genes to
determine the profile of gene expression in OVCA (29), they were used as the internal
controls in the present study. The results showed that GAPDH mRNA
levels were significantly upregulated in OVCA tissues (Fig. 1B). This finding was supported by
data from the Oncomine database (oncomine.org/resource/main.html).
In contrast to the GAPDH mRNA expression patterns, the expression
levels of GAPDH protein did not differ significantly between the
cancerous and non-cancerous tissues (Fig. S1).
Mutant GAPDH 3′UTR promotes cell
growth and cell motility of OVCA
To determine the biological roles of the effect of
the mutant GAPDH 3′UTR (NM_001289745.2:c.*1325_*1326insC) on OVCA,
the OVCA cell lines SKOV3 and OVCAR3 stably expressing WT or mutant
GAPDH 3′UTR were constructed. A CCK-8 assay demonstrated that the
mutant but not the WT GAPDH 3′UTR-overexpressing OVCA cells
exhibited significantly increased cell growth compared with the
control cells (Fig. 2A and B).
Furthermore, the effect of the GAPDH 3′UTR on cell motility of OVCA
cells was assessed using wound healing and Transwell assays. The
results showed that overexpression of the mutant GAPDH 3′UTR in
OVCA cells significantly promoted migration as the wound closure
rate was increased (Fig. 2C and D).
Consistent with this result, Transwell assays also showed that the
mutant GAPDH 3′UTR significantly increased the migratory and
invasive capacity of OVCA cells (Fig.
2E-G). Taken together, these results suggest that mutant GAPDH
3′UTR promoted cell growth and cell motility of OVCA cells.
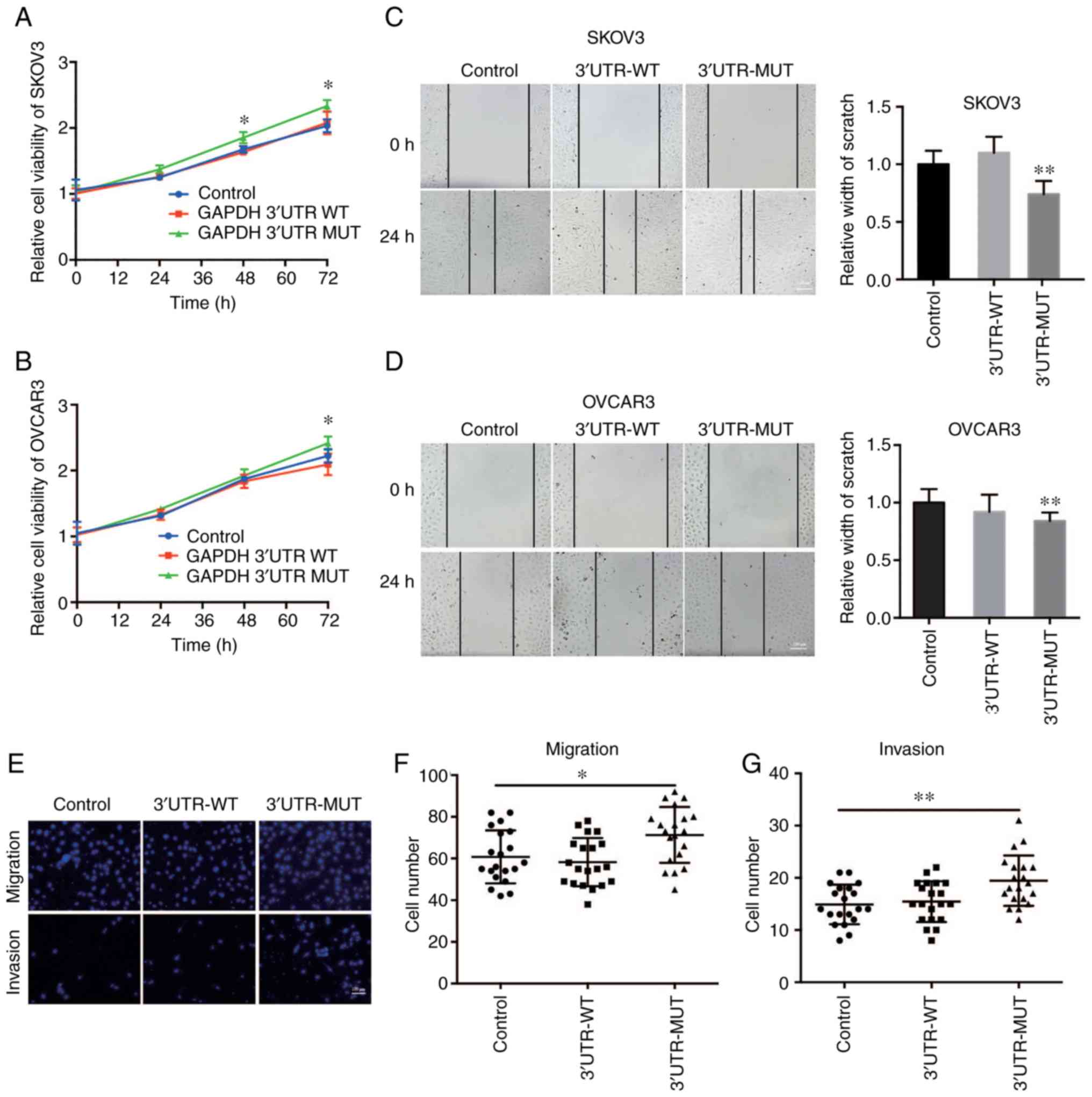 | Figure 2.Mutant GAPDH 3′UTR promotes tumor
growth and cell motility of OVCA. Cell viability of (A) SKOV3 and
(B) OVCAR3 cells stably expressing WT or mutant (MUT) GAPDH 3′UTR
was determined by CCK-8 assay. Cells infected with pLKO.1-control
lentivirus were used as the negative control. In the CCK-8 assay
experiment, 4 parallel wells were set up in each group, and the
experiment was repeated three times. Wound healing assays were
performed to assess cell migration in (C) SKOV3 and (D) OVCAR3
cells stably expressing WT or MUT GAPDH 3′UTR. Cells infected with
pLKO.1-control lentivirus were used as the negative control. n=20.
Scale bar in D represents 100 µm. (E) Cell migration and invasive
capacity was examined in SKOV3 cells stably expressing WT or MUT
GAPDH 3′UTR using a Transwell assay. SKOV3 cells infected with
pLKO.1-control lentivirus were used as the negative control. Scale
bar in E represents 100 µm. Quantification of (F) Transwell
migration and (G) Transwell invasion assays, n=20. In the Transwell
assay experiment, three parallel wells were set up in each group,
and the experiment was repeated three times. *P<0.05,
**P<0.01, GAPDH, glyceraldehyde 3-phosphate dehydrogenase; UTR,
untranslated region; OVCA, ovarian cancer. |
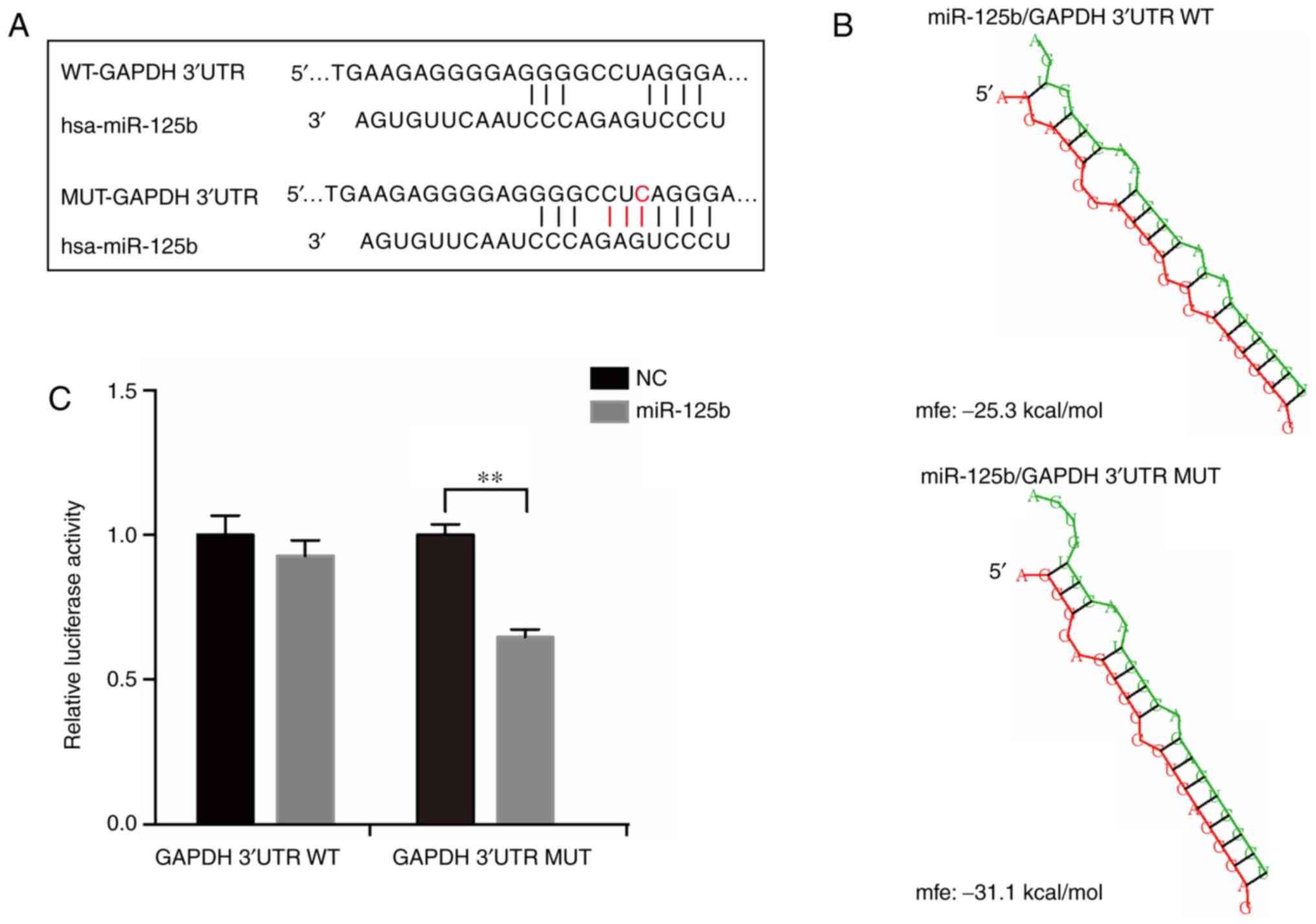
Mutation in GAPDH 3′UTR results in the
creation of a miR-125b binding site
To further analyze the molecular mechanisms of the
frequent mutation in GAPDH 3′UTR, bioinformatics algorithms were
used to predict miRNAs which may interact with the mutation site,
including miRDB (30) and miRcode
(31). The results of the alignment
analysis suggested that the mutated sequence matches a sequence
located in the ‘seed region’ of the binding sites of miR-125b
(Fig. 3A). The structure and free
energy of potential binding site interactions between miR-125b with
GAPDH mRNA 3′UTR (WT and mutant) were analyzed using RNAhybrid
(32). The results showed that the
mutant GAPDH 3′UTR had a stronger affinity for miR-125b compared
with the WT (Fig. 3B). To further
confirm the binding activation of GAPDH 3′UTR with miR-125b,
luciferase reporter plasmids with the WT or mutant GAPDH 3′UTR were
constructed and co-transfected with miR-125b mimics in 293T cells.
The results showed that only when transfected with the mutant
construct was the activity of luciferase significantly reduced by
miR-125b (Fig. 3C). Based on these
results, it was concluded that the frequent mutation in GAPDH 3′UTR
created a new binding site for miR-125b.
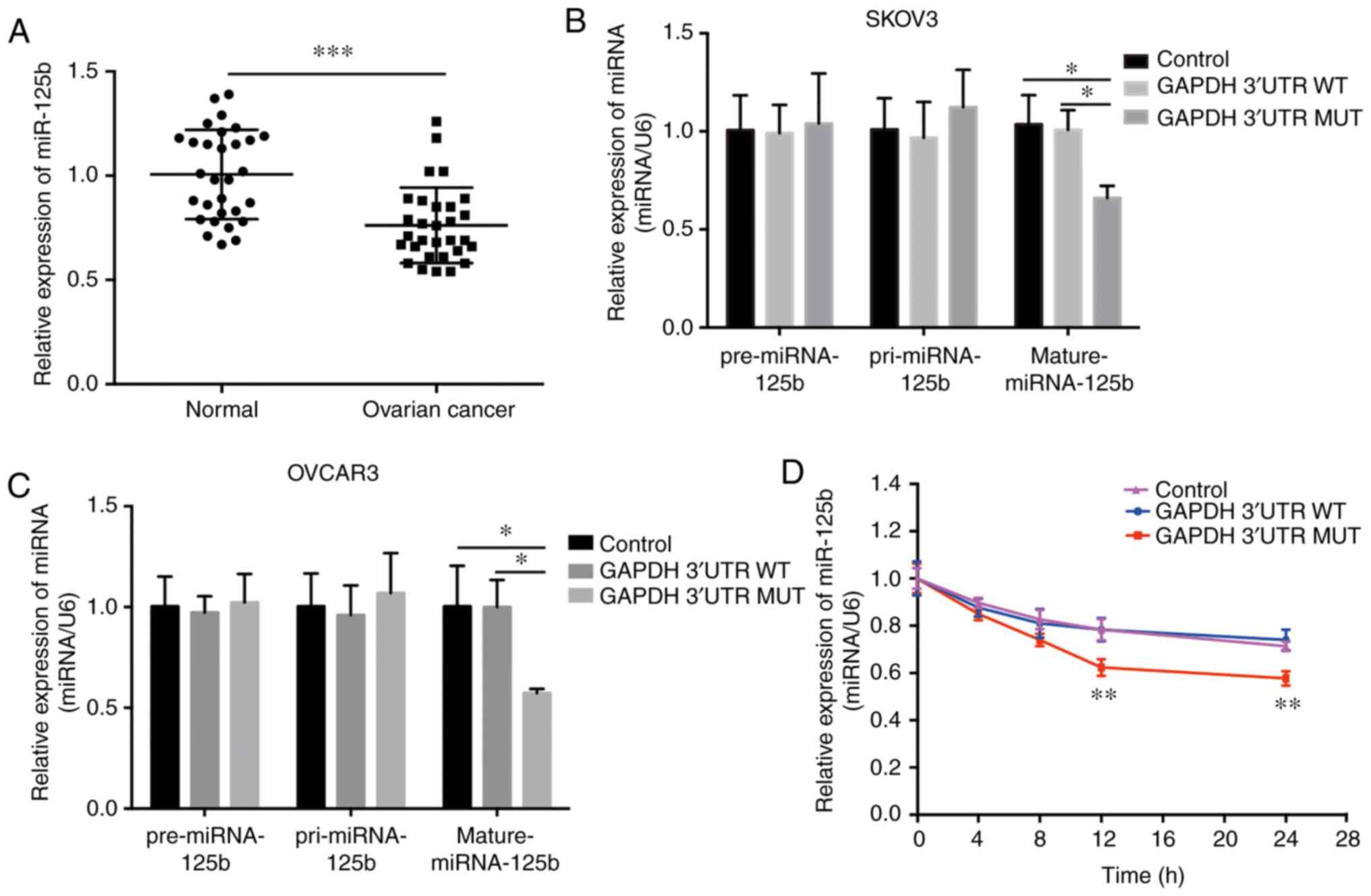
Mutation in GAPDH 3′UTR results in the
downregulation of mature miR-125b levels in OVCA cells
Previous studies have shown that miR-125b is
dysregulated in different types of cancer (33–36).
The expression of miR-125b was quantified in 30 paired normal and
OVCA tissues using RT-qPCR, and it was found that miR-125b was
significantly downregulated in the OVCA tissues (Fig. 4A). This finding is supported by
several recent studies (33,37).
Additionally, miR-125b levels may have been regulated by the mutant
GAPDH 3′UTR in OVCA cells. Expression of the mutant GAPDH 3′UTR
significantly decreased the levels of mature miR-125b, whereas
neither pre-miRNA nor pri-miRNA levels were altered (Fig. 4B and C); and the stability of mature
miR-125b was significantly decreased in cells stably expressing the
mutant GAPDH 3′UTR following inhibition of intracellular RNA
transcription with actinomycin D treatment (Fig. 4D). These phenomena were abrogated in
cells with the WT 3′UTR of GAPDH (Fig.
4B-D). Additionally, the mature miR-125b level in the OVCA
mutant tissues (NM_001289745.2:c.*1325_*1326insC) and OVCA
wild-type tissues was analyzed. The results showed there was no
difference between these two group, and the main reason was the low
abundance of GAPDH mutation (Fig.
S2). Together, these results suggest that the mutant GAPDH
3′UTR reduced the relative levels of mature miR-125b, and this
effect was not associated with transcriptional regulation or miRNA
precursor processing.
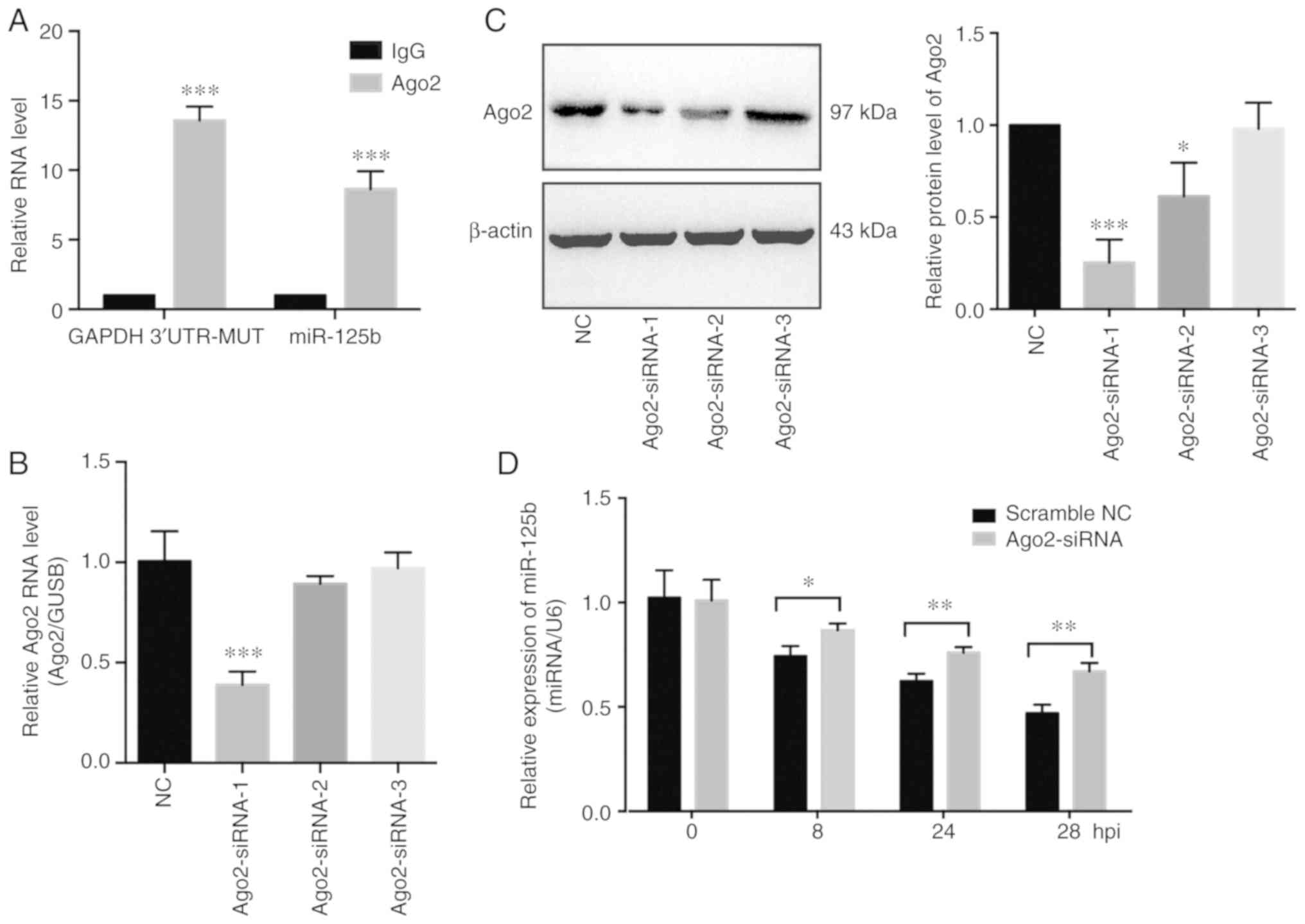
Mutant GAPDH 3′UTR regulates miR-125b
levels in an Ago2-dependent manner
It was previously reported that miRNAs guide
argonaute RISC catalytic component 2 (Ago2) protein to distinct
target sites on mRNAs (38).
Therefore, the involvement of Ago2 on the interaction of miR-125b
and GAPDH 3′UTR-MUT was assessed. Subsequently, to further explore
the potential mechanisms involved, an anti-Ago2 RIP experiment was
performed on extracts of SKOV3 cells (stably expressing mutant
GAPDH 3′UTR). The mutant GAPDH 3′UTR fragment and miR-125b were
enriched in Ago2-containing complexes (Fig. 5A). In addition, to investigate the
role of Ago2 on the biological function of the mutant GAPDH 3′UTR,
Ago2 expression was knocked down using siRNA (Fig. 5B and C). Knockdown of Ago2 partially
inhibited the degradation of miR-125b induced by the mutant GAPDH
3′UTR (Fig. 5D). Previous studies
showed that miRNAs act as miRNA ribonucleoprotein complexes
containing Ago2, an important component of RNA induced silencing
complex (RISC). The above results showed that miR-125b and the
mutant GAPDH 3′UTR were involved in a RISC, suggesting that the
mutant GAPDH 3′UTR may function as a sponge of miR-125b in OVCA
cells.
Mutant GAPDH 3′UTR promotes cancer
development by sponging miR-125b, thereby regulating STAT3
expression in OVCA cells
To investigate whether the effects of the mutant
GAPDH 3′UTR on OVCA development were mediated by miR-125b, OVCA
cells stably expressing the mutant GAPDH 3′UTR were transfected
with miR-125b mimics. Transfection of miR-125b mimics abrogated the
tumor promoting effects induced by mutant GAPDH 3′UTR, which
suggested that the mutant GAPDH 3′UTR promoted cancer development
by regulating miR-125b (Fig. 6A-C).
To examine the effects of miR-125b on the mechanisms and functions
of the mutant GAPDH 3′UTR, direct target genes of miR-125b were
selected using TargetScan. The results showed that STAT3 may be an
effective target gene. Luciferase reporter assays were performed to
assess whether STAT3 directly interacted with miR-125b. The results
demonstrated that miR-125b reduced luciferase activity in the
pGL3-STAT3-3′UTR-WT transfected cells (Fig. 6D). To determine the underlying
relationship between GAPDH 3′UTR and STAT3, the expression of STAT3
was assessed in SKOV3 cells stably expressing GAPDH 3′UTR. The
results showed that overexpression of the mutant GAPDH 3′UTR
upregulated STAT3 expression (Fig. 6E
and F). As it has been reported that elevated STAT3 expression
in OVCA contributes to tumorigenesis and progression (39,40),
it was concluded that the mutant GAPDH 3′UTR facilitates OVCA cell
growth and cell motility by sponging miR-125b and thus upregulating
STAT3 levels.
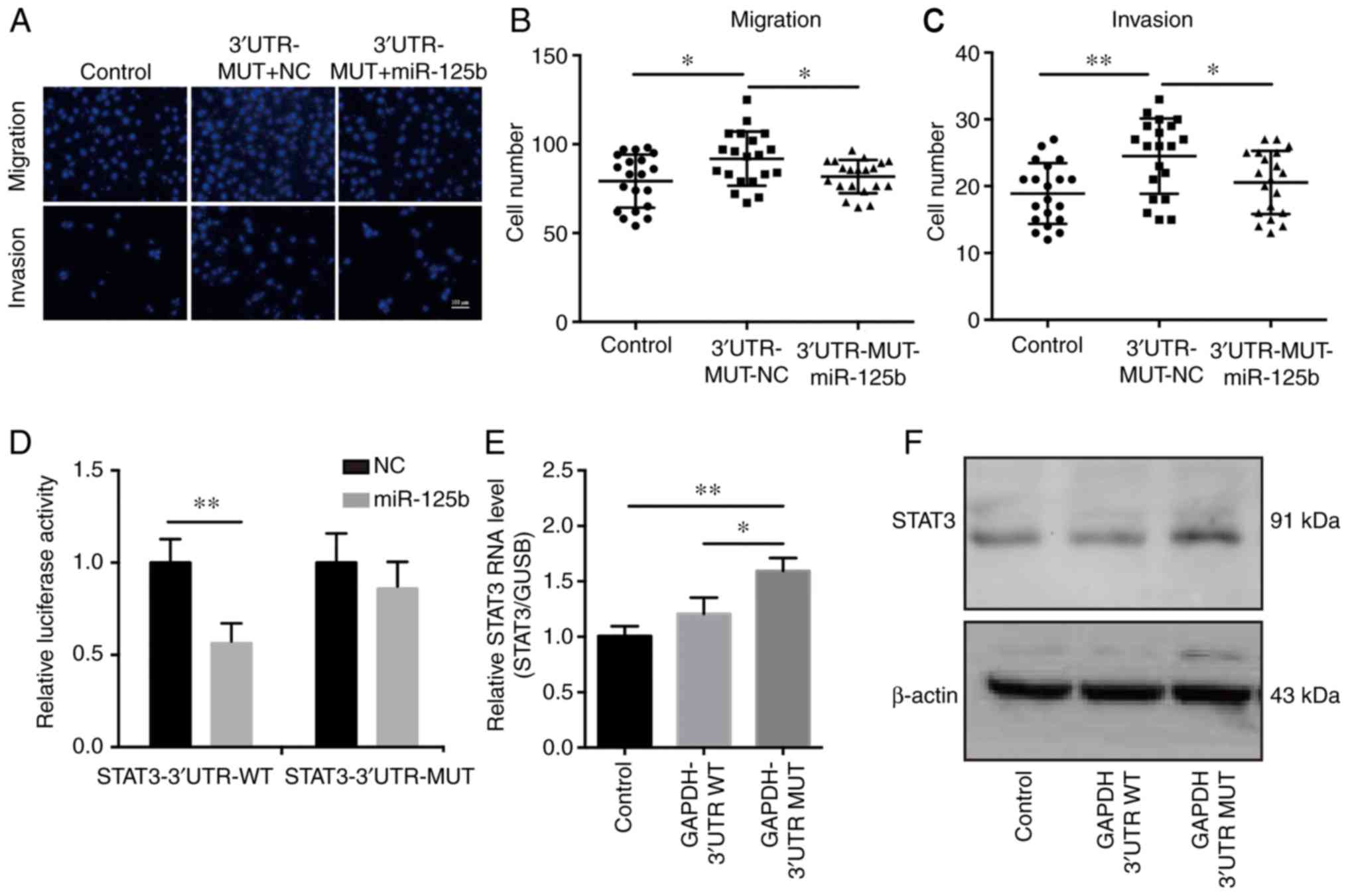 | Figure 6.Mutant GAPDH 3′UTR promotes cancer
development by sponging miR-125b, thereby regulating STAT3
expression in OVCA cells. (A) miR-125b mimics or scramble NC were
transfected into SKOV3 cells stably expressing the mutant (MUT)
GAPDH 3′UTR. Migration and invasion assays were performed to assess
the effects of miR-125b on cell motility. SKOV3 cells infected with
pLKO.1-control lentivirus were used as the negative control. Scale
bar in A represents 100 µm. Quantification of (B) migration and (C)
invasion, n=20. (D) Luciferase reporter assays showed that
overexpression of miR-125b reduced luciferase activity in 293T
cells transfected with the STAT3 3′UTR-WT vector. (E) RT-qPCR and
(F) western blot analysis of STAT3 expression in SKOV3 cells stably
expressing WT of mutant GAPDH 3′UTR. SKOV3 cells infected with
pLKO.1-control lentivirus were used as the negative control.
*P<0.05, **P<0.01. GAPDH, glyceraldehyde 3-phosphate
dehydrogenase; STAT3, signal transducer and activator of
transcription 3; UTR, untranslated region; OVCA, ovarian cancer;
RT-qPCR, reverse transcription-quantitative PCR; miR, microRNA; NC,
negative control. |
Discussion
Recently, the genomic sequencing of cancer tissues
has provided evidence that the majority of cancer genomes contain
thousands of somatic mutations, and some of these somatic mutations
may exhibit a functional effect on tumorigenesis and metastasis
(16,41,42).
In the present study, GAPDH mRNA expression levels were upregulated
in ovarian cancer (OVCA) tissues, and a frequent somatic mutation
in the 3′UTR of GAPDH (NM_001289745.2:c.*1325_*1326insC) was
identified. The mutant GAPDH 3′UTR significantly promoted OVCA cell
growth and cell motility, whereas the WT GAPDH 3′UTR had no effect
on these biological processes.
Numerous studies have shown that somatic mutations
in the 3′UTR may promote tumor progression by altering miRNA
targeting efficiency and consequently affecting miRNA-mRNA
interactions (18,43,44).
To assess the functional mechanisms of a frequent somatic mutation
located in the 3′UTR of GAPDH, its effect on miRNA binding was
analyzed. The results showed that the mutation created a new
binding site for miR-125b. RT-qPCR was performed on OVCA cell lines
stably expressing WT or mutant GAPDH 3′UTR, and the results showed
that the mutant 3′UTR resulted in a ~40% decrease in mature
miR-125b levels, and this regulation was dependent on the
expression of Ago2. These results suggest that the GAPDH 3′UTR
mutation, which is able to bind to miR-125b target sites, may
impair miRNA-mRNA interaction and sponge miR-125b.
It was reported that miR-125b was involved in the
initiation and progression of several types of cancer, such as
urothelial tumors (45), liver
(46,47), bladder cancer (48) and ovarian cancer (33). miR-125b may act as a tumor
suppressor, inhibiting invasion, migration and angiogenesis
(48,49). In the present study, miR-125b was
downregulated in OVCA tissues compared with the matching normal
tissues. Transfection of miR-125b mimics suppressed OVCA cell
growth, and decreased cell motility. In addition, transfection of
miR-125b mimics counteracted the tumor promoting effects induced by
the mutant GAPDH 3′UTR. Taken together, these results indicated
that the mutant GAPDH 3′UTR promoted the development of OVCA by
sponging miR-125b.
Several recent studies have reported that mRNAs,
transcribed pseudogenes and other RNAs may bind to miRNAs and act
as potent miRNA sponges (50–52).
These molecules have been termed competitive endogenous RNAs
(20). ceRNAs can sequester miRNAs,
thereby protecting the target RNAs of the respective miRNA from
repression (23). In the present
study, the mutant GAPDH 3′UTR regulated miR-125b levels in an
Ago2-dependent manner, which suggested that the mutant GAPDH may
act as a ceRNA. Bioinformatics algorithms and a luciferase reporter
assay identified STAT3 as the target of miR-125b. Moreover, this
finding was supported by several recent studies (53,54).
To further determine the effect of the mutant GAPDH 3′UTR on
miR-125b-mediated regulation of STAT3, the expression levels of
STAT3 in OVCA cells stably expressing GAPDH 3′UTR were assessed.
Overexpression of the mutant GAPDH 3′UTR resulted in a significant
increase in STAT3 levels. Several studies have shown that STAT3
levels are elevated in several different types of cancer, and it is
a critical regulator of numerous cellular processes, including
proliferation, invasion and metastasis (55–57).
Therefore, the mutant GAPDH 3′UTR may promote tumor growth and cell
motility through increasing the expression of STAT3.
In conclusion, the primary finding of the present
study was that mutations in GAPDH 3′UTR promoted development of
OVCA by creating a new miR-125b binding site, thereby promoting
oncogenic STAT3 expression in ovarian cancer.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
We would like to thank the Research Fund for
Academician Lin He New Medicine for the financial support of this
study, and Genex Health Co., Ltd. (Beijing, China) for their
assistance during the preparation of this manuscript.
Funding
The study was supported by grants from Research Fund
for Academician Lin He New Medicine.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
PL, YZ and HH contributed to the design of this
study. PL, YZ, TC, XS and HH performed the experiments, and
analyzed and interpreted the data. PL, YZ and HH wrote the
manuscript. XS revised the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The Third Affiliated Hospital of Guangzhou Medical University
(Guangzhou, Guangdong, China), and written informed consent was
obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Glaser PE and Gross RW: Rapid
plasmenylethanolamine-selective fusion of membrane bilayers
catalyzed by an isoform of glyceraldehyde-3-phosphate
dehydrogenase: Discrimination between glycolytic and fusogenic
roles of individual isoforms. Biochemistry. 34:12193–12203. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tisdale EJ, Kelly C and Artalejo CR:
Glyceraldehyde-3-phosphate dehydrogenase interacts with Rab2 and
plays an essential role in endoplasmic reticulum to Golgi transport
exclusive of its glycolytic activity. J Biol Chem. 279:54046–54052.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hara MR and Snyder SH: Nitric
oxide-GAPDH-Siah: A novel cell death cascade. Cell Mol Neurobiol.
26:527–538. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Baxi MD and Vishwanatha JK: Uracil
DNA-glycosylase/glyceraldehyde-3-phosphate dehydrogenase is an Ap4A
binding protein. Biochemistry. 34:9700–9707. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng L, Roeder RG and Luo Y: S phase
activation of the histone H2B promoter by OCA-S, a coactivator
complex that contains GAPDH as a key component. Cell. 114:255–266.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Barroso I, Benito B, Garcí-Jiménez C,
Hernández A, Obregón MJ and Santisteban P: Norepinephrine,
tri-iodothyronine and insulin upregulate glyceraldehyde-3-phosphate
dehydrogenase mRNA during Brown adipocyte differentiation. Eur J
Endocrinol. 141:169–179. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Valenti MT, Bertoldo F, Dalle Carbonare L,
Azzarello G, Zenari S, Zanatta M, Balducci E, Vinante O and Lo
Cascio V: The effect of bisphosphonates on gene expression: GAPDH
as a housekeeping or a new target gene? BMC Cancer. 6:492006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sikand K, Singh J, Ebron JS and Shukla GC:
Housekeeping gene selection advisory: Glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) and β-actin are targets of miR-644a. PLoS
One. 7:e475102012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bjerregaard H, Pedersen S, Kristensen SR
and Marcussen N: Reference genes for gene expression analysis by
real-time reverse transcription polymerase chain reaction of renal
cell carcinoma. Diagn Mol Pathol. 20:212–217. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma Y, Dai H, Kong X and Wang L: Impact of
thawing on reference gene expression stability in renal cell
carcinoma samples. Diagn Mol Pathol. 21:157–163. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Majidzadeh-A K, Esmaeili R and Abdoli N:
TFRC and ACTB as the best reference genes to quantify Urokinase
plasminogen activator in breast cancer. BMC Res Notes. 4:2152011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo Y, Chen JX, Yang S, Fu XP, Zhang Z,
Chen KH, Huang Y, Li Y, Xie Y and Mao YM: Selection of reliable
reference genes for gene expression study in nasopharyngeal
carcinoma. Acta Pharmacol Sin. 31:1487–1494. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stratton MR: Exploring the genomes of
cancer cells: Progress and promise. Science. 331:1553–1558. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nicoloso MS, Sun H, Spizzo R, Kim H,
Wickramasinghe P, Shimizu M, Wojcik SE, Ferdin J, Kunej T, Xiao L,
et al: Single-nucleotide polymorphisms inside microRNA target sites
influence tumor susceptibility. Cancer Res. 70:2789–2798. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Y, Du X, Zhou Z, Jiang J, Zhang Z, Ye
L and Hong H: A gain-of-function ACTC1 3′UTR mutation that
introduces a miR-139-5p target site may be associated with a
dominant familial atrial septal defect. Sci Rep. 6:254042016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Flynt AS and Lai EC: Biological principles
of microRNA-mediated regulation: Shared themes amid diversity. Nat
Rev Genet. 9:831–842. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fang L, Du WW, Yang X, Chen K, Ghanekar A,
Levy G, Yang W, Yee AJ, Lu WY, Xuan JW, et al: Versican
3′-untranslated region (3′-UTR) functions as a ceRNA in inducing
the development of hepatocellular carcinoma by regulating miRNA
activity. FASEB J. 27:907–919. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Karreth FA, Tay Y, Perna D, Ala U, Tan SM,
Rust AG, DeNicola G, Webster KA, Weiss D, Perez-Mancera PA, et al:
In vivo identification of tumor-suppressive PTEN ceRNAs in an
oncogenic BRAF-induced mouse model of melanoma. Cell. 147:382–395.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee DY, Shatseva T, Jeyapalan Z, Du WW,
Deng Z and Yang BB: A 3′-untranslated region (3′UTR) induces organ
adhesion by regulating miR-199a* functions. PLoS One. 4:e45272009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sumazin P, Yang X, Chiu HS, Chung WJ, Iyer
A, Llobet-Navas D, Rajbhandari P, Bansal M, Guarnieri P, Silva J
and Califano A: An extensive microRNA-mediated network of RNA-RNA
interactions regulates established oncogenic pathways in
glioblastoma. Cell. 147:370–381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stahlhut C and Slack FJ: MicroRNAs and the
cancer phenotype: Profiling, signatures and clinical implications.
Genome Med. 5:1112013. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li YL, Ye F, Hu Y, Lu WG and Xie X:
Identification of suitable reference genes for gene expression
studies of human serous ovarian cancer by real-time polymerase
chain reaction. Anal Biochem. 394:110–116. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang J, Yu H, Ye L, Jin L, Yu M and Lv Y:
Integrated regulatory mechanisms of miRNAs and targeted genes
involved in colorectal cancer. Int J Clin Exp Pathol. 8:517–529.
2015.PubMed/NCBI
|
|
31
|
Jeggari A, Marks DS and Larsson E:
miRcode: A map of putative microRNA target sites in the long
non-coding transcriptome. Bioinformatics. 28:2062–2063. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rehmsmeier M, Steffen P, Hochsmann M and
Giegerich R: Fast and effective prediction of microRNA/target
duplexes. RNA. 10:1507–1517. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ying X, Wei K, Lin Z, Cui Y, Ding J, Chen
Y and Xu B: MicroRNA-125b suppresses ovarian cancer progression via
suppression of the epithelial-mesenchymal transition pathway by
targeting the SET protein. Cell Physiol Biochem. 39:501–510. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vriens MR, Weng J, Suh I, Huynh N,
Guerrero MA, Shen WT, Duh QY, Clark OH and Kebebew E: MicroRNA
expression profiling is a potential diagnostic tool for thyroid
cancer. Cancer. 118:3426–3432. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jia HY, Wang YX, Yan WT, Li HY, Tian YZ,
Wang SM and Zhao HL: MicroRNA-125b functions as a tumor suppressor
in hepatocellular carcinoma cells. Int J Mol Sci. 13:8762–8774.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Henson BJ, Bhattacharjee S, O'Dee DM,
Feingold E and Gollin SM: Decreased expression of miR-125b and
miR-100 in oral cancer cells contributes to malignancy. Genes
Chromosomes Cancer. 48:569–582. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Iorio MV, Visone R, Di Leva G, Donati V,
Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, et
al: MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Meister G, Landthaler M, Patkaniowska A,
Dorsett Y, Teng G and Tuschl T: Human Argonaute2 mediates RNA
cleavage targeted by miRNAs and siRNAs. Mol Cell. 15:185–197. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Saini U, Naidu S, ElNaggar AC, Bid HK,
Wallbillich JJ, Bixel K, Bolyard C, Suarez AA, Kaur B, Kuppusamy P,
et al: Elevated STAT3 expression in ovarian cancer ascites promotes
invasion and metastasis: A potential therapeutic target. Oncogene.
36:168–181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wen W, Liang W, Wu J, Kowolik CM, Buettner
R, Scuto A, Hsieh MY, Hong H, Brown CE, Forman SJ, et al: Targeting
JAK1/STAT3 signaling suppresses tumor progression and metastasis in
a peritoneal model of human ovarian cancer. Mol Cancer Ther.
13:3037–3048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Boehm JS and Hahn WC: Towards systematic
functional characterization of cancer genomes. Nat Rev Genet.
12:487–498. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chin L, Andersen JN and Futreal PA: Cancer
genomics: From discovery science to personalized medicine. Nat Med.
17:297–303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu W, Wu L, Zhu M, Wang Z, Wu M, Li P, Nie
Y, Lin X, Hu J, Eskilsson E, et al: miRNA mediated noise making of
3′UTR mutations in cancer. Genes (Basel). 9(pii): E5452018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lopes-Ramos CM, Barros BP, Koyama FC,
Carpinetti PA, Pezuk J, Doimo NTS, Habr-Gama A, Perez RO and
Parmigiani RB: E2F1 somatic mutation within miRNA target site
impairs gene regulation in colorectal cancer. PLoS One.
12:e01811532017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Veerla S, Lindgren D, Kvist A, Frigyesi A,
Staaf J, Persson H, Liedberg F, Chebil G, Gudjonsson S, Borg A, et
al: MiRNA expression in urothelial carcinomas: Important roles of
miR-10a, miR-222, miR-125b, miR-7 and miR-452 for tumor stage and
metastasis, and frequent homozygous losses of miR-31. Int J Cancer.
124:2236–2242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Alpini G, Glaser SS, Zhang JP, Francis H,
Han Y, Gong J, Stokes A, Francis T, Hughart N, Hubble L, et al:
Regulation of placenta growth factor by microRNA-125b in
hepatocellular cancer. J Hepatol. 55:1339–1345. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou JN, Zeng Q, Wang HY, Zhang B, Li ST,
Nan X, Cao N, Fu CJ, Yan XL, Jia YL, et al: MicroRNA-125b
attenuates epithelial-mesenchymal transitions and targets stem-like
liver cancer cells through small mothers against decapentaplegic 2
and 4. Hepatology. 62:801–815. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Han Y, Liu Y, Zhang H, Wang T, Diao R,
Jiang Z, Gui Y and Cai Z: Hsa-miR-125b suppresses bladder cancer
development by down-regulating oncogene SIRT7 and oncogenic long
non-coding RNA MALAT1. FEBS Lett. 587:3875–3882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu N, Zhang L, Meisgen F, Harada M,
Heilborn J, Homey B, Grandér D, Ståhle M, Sonkoly E and Pivarcsi A:
MicroRNA-125b down-regulates matrix metallopeptidase 13 and
inhibits cutaneous squamous cell carcinoma cell proliferation,
migration, and invasion. J Biol Chem. 287:29899–29908. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tay Y, Kats L, Salmena L, Weiss D, Tan SM,
Ala U, Karreth F, Poliseno L, Provero P, Di Cunto F, et al:
Coding-independent regulation of the tumor suppressor PTEN by
competing endogenous mRNAs. Cell. 147:344–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jin XJ, Chen XJ, Zhang ZF, Hu WS, Ou RY,
Li S, Xue JS, Chen LL, Hu Y and Zhu H: Long noncoding RNA SNHG12
promotes the progression of cervical cancer via modulating
miR-125b/STAT3 axis. J Cell Physiol. 234:6624–6632. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu LH, Li H, Li JP, Zhong H, Zhang HC,
Chen J and Xiao T: miR-125b suppresses the proliferation and
migration of osteosarcoma cells through down-regulation of STAT3.
Biochem Biophys Res Commun. 416:31–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bromberg J and Darnell JE Jr: The role of
STATs in transcriptional control and their impact on cellular
function. Oncogene. 19:2468–2473. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Darnell JE: Validating Stat3 in cancer
therapy. Nat Med. 11:595–596. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Levy DE and Darnell JE Jr: Stats:
Transcriptional control and biological impact. Nat Rev Mol Cell
Biol. 3:651–662. 2002. View
Article : Google Scholar : PubMed/NCBI
|