Introduction
Gastric cancer (GC) is one of the most common
gastrointestinal malignancies and remains the leading cause of
cancer-related deaths worldwide, with over half of the cases
occurring in East Asia (1,2). Adequate surgical resection is the only
curative treatment strategy for localized GC, however a high risk
of recurrence and metastasis remains after surgical resection
(3). Furthermore, most patients are
diagnosed at an advanced stage, and chemotherapy is the first-line
treatment (4). Cisplatin
[cis-diamminedichloroplatinum(II); DDP] is a widely used
platinum-based antineoplastic agent for GC treatment, especially
for patients at an advanced stage (5). However, its application is limited by
drug resistance and its systemic toxicity, such as nephrotoxicity,
ototoxicity, neurotoxicity and hepatotoxicity (6). Thus, it is essential to develop novel
chemical sensitizing agents for DDP to enhance its efficacy and
attenuate side effects in GC patients.
Resveratrol (3,4′,5-trihydroxystilbene; RES) is a
nonflavonoid polyphenolic compound notably present in grapes and
red wine (7). It plays a protective
role against a wide variety of diseases, including cardiovascular
diseases (8), neurodegenerative
diseases (9), inflammatory diseases
(10) and diabetes (11). Moreover, recent preclinical studies
have identified RES as a chemopreventive and therapeutic agent as
well as a chemical sensitizer for various malignant tumors
(12), which displays lower
cytotoxicity to normal cells than traditional therapy. Although
previous studies have demonstrated that RES exerts its anticancer
effects against GC cells by promoting apoptosis and inducing cell
cycle arrest or suppressing migration and invasion (13–15),
whether it can sensitize GC cells to DDP and the underlying
mechanisms remain to be determined.
The endoplasmic reticulum (ER) is the principle
organelle responsible for biogenesis, folding and trafficking of
over one-third of the proteins in eukaryotic cells (16). Extrinsic and intrinsic perturbations
disturb ER homeostasis, trigger ER stress and activate the unfolded
protein response (UPR), which promotes cell survival, or induce
apoptosis if ER stress is severe or chronic (17). In fact, the UPR promotes cancer cell
survival by enhancing cancer cell adaptation to hostile
environmental conditions, and regulating the UPR is a promising
anticancer strategy (18). Recent
studies have demonstrated that RES promoted ER stress-mediated
apoptosis in colon cancer cells (19), ovarian cancer cells (20), and malignant melanoma cells
(21). However, the role of RES in
ER stress-induced apoptosis in GC cells has not been studied to
date.
In the present study, it was revealed that RES
synergized with DDP in antineoplastic effects against the human GC
cell line AGS by inducing ER stress-mediated apoptosis and G2/M
cell cycle arrest. Mechanistic experiments revealed that RES/DDP
combination treatment activated the PRKR-like ER kinase
(PERK)/eukaryotic translation initiation factor 2α
(eIF2α)/activating transcription factor 4 (ATF4)-CCAAT/enhancer
binding protein homologous protein (CHOP) signaling pathway and
upregulated cleaved caspase-12. Furthermore, RES/DDP combination
treatment arrested AGS cells in G2/M phase by inactivating the
cyclin-dependent kinase 1 (CDK1)-cyclin B1 complex and upregulating
p21Waf1/Cip1 and p27Kip protein levels. The
present results provided evidence that may lead to the future
application of RES as an adjuvant for GC chemotherapy.
Materials and methods
Cell culture
The human GC cell lines AGS, KATO III, MKN-45 and
NCI-N87 were obtained from the Type Culture Collection of the
Chinese Academy of Sciences. The AGS cells were cultured in Hams
F-12K (Kaighns) medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin
and 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.)
at 37°C in a humidified incubator containing 5% CO2. The
KATO III cell line was cultured in Iscoves Modified Dulbeccos
Medium (IMDM; Gibco; Thermo Fisher Scientific, Inc.), and the
MKN-45 and NCI-N87 cell lines were cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) as described above.
Cell Counting Kit-8 (CCK-8) assay
Cell viability was detected by a CCK-8 assay
(Dojindo Molecular Technologies, Inc.). Approximately
5×103 cells/well were seeded into 96-well plates and
incubated overnight at 37°C. Next, various doses of RES (0, 5, 10,
20, 30, 40, 50, 60, 70, and 80 µM) and/or DDP (0, 0.5, 1, and 2
µg/ml) (both from Selleck Chemicals) were added. After culturing
for another 24, 48, 72 or 96 h, each well was incubated with 10 µl
CCK-8 solution for 1–3 h before measuring the absorbance at 450 nm
using a microplate reader (Bio-Rad Laboratories, Inc.).
Evaluation of the combination effect
of RES and DDP
The combination index (CI) was applied to evaluate
the combination effect of RES and DDP in AGS cells according to the
Chou-Talalay method (22). CI was
calculated by CompuSyn software (ComboSyn, Inc.), and the
combination effect was defined as follows: CI <1, synergistic
effect; CI >1, antagonistic effect; and CI=1, additive
effect.
Morphological observation
AGS cells were seeded in 6-well plates
(1×105 cells/well), incubated at 37°C overnight, and
exposed to RES (20 µM) and DDP (1 µg/ml) alone or in combination
for 48 h. Cell morphology was observed using an inverted light
microscope (magnification, ×100; Olympus Corporation).
Colony-forming assay
AGS cells were treated with RES (20 µM) and DDP (1
µg/ml) alone or in combination for 48 h. Thereafter, the cells were
trypsinized by 0.25% trypsin (Gibco; Thermo Fisher Scientific,
Inc.) and dispensed into individual wells of 6-well plates at a
density of 1×103 cells/well. Following another 14 days
of drug-free culture and changing of the medium every other day,
the cells were fixed with 4% paraformaldehyde at 25°C for 15 min
and stained with 0.1% crystal violet at 25°C for another 15 min.
Finally, colonies (≥10 cells) were counted using an inverted light
microscope (magnification, ×100; Olympus Corporation).
Cell apoptosis assay
Apoptotic cells were detected by an FITC-Annexin V
Apoptosis Detection Kit (BD Biosciences) according to the
manufacturers instructions. The cells (1×105/well) were
seeded in 6-well plates and incubated overnight at 37°C. Then, the
cells were treated with RES (0, 20, and 40 µM) for 72 h or exposed
to RES (20 µM) and DDP (1 µg/ml) alone or in combination for 48 and
72 h. Subsequently, the cells were harvested with EDTA-free trypsin
and resuspended in 500 µl of 1X binding buffer followed by
incubation with FITC-Annexin V (5 µl) and PI (5 µl) at 25°C in the
dark for 15 min. Finally, the apoptotic cells were analyzed by a BD
FACSCanto II flow cytometer (BD Biosciences) and FlowJo software
(version 10.4; BD Biosciences).
Cell cycle assay
The cell cycle assays were performed according to
the manufacturers instructions using a cell cycle staining kit
[MultiSciences (Lianke) Biotech Co., Ltd.] as previously described
(23). After administration of RES
(20 µM) and DDP (1 µg/ml) alone or in combination, the cells were
harvested and fixed with 75% ethanol at −20°C overnight.
Thereafter, the cells were hydrated with cold phosphate-buffered
saline (PBS) for 15 min and incubated with DNA staining solution at
25°C for 30 min in the dark before evaluation by a BD FACSCanto II
flow cytometer and ModFit LT software (version 5.0.9; Verity
Software House Co., Ltd.).
Detection of intracellular free
calcium ions (Ca2+)
The intracellular free Ca2+ concentration
was determined by the calcium probe Fluo-4 AM (Beyotime Institute
of Biotechnology) according to the manufacturers instructions.
After exposure to RES (20 µM) and DDP (1 µg/ml) alone or in
combination for 48 h, the cells were collected, washed twice with
PBS and incubated with 2 µM Fluo-4 AM probe for 30 min at 37°C.
Subsequently, the cells were washed with PBS, and cell fluorescence
was determined using a BD FACSCanto II flow cytometer (excitation
wavelength, 488 nm; emission wavelength, 525 nm).
Western blot analysis
Following treatment with RES (10, 20, and 40 µM) and
DDP (1 µg/ml) alone or combination treatment of RES (20 µM) and DDP
(1 µg/ml) for 48 h, total protein from AGS cells was extracted
using cell lysis buffer (Cell Signaling Technology, Inc.) according
to the manufacturers instructions. Western blot analysis was
performed as previously described (24). Briefly, 20 µg of protein was loaded
onto each lane and then transferred to polyvinylidene difluoride
(PVDF) membranes. The membranes were incubated with primary
antibodies overnight at 4°C, followed by goat anti-rabbit secondary
antibody (1:5,000, cat. no. HA1001; HuaBio Co., Ltd.) or goat
anti-mouse secondary antibody (1:10,000, cat. no. BL001A, BioSharp
Co., Ltd.) for 1 h at 25°C. The primary antibodies used were rabbit
antibodies against CHOP (also named DDIT3; 1:1,000; product code
ab179823; Abcam), poly-ADP-ribose polymerase (PARP; 1:1,000; cat.
no. 9532; Cell Signaling Technology, Inc.), Bax (1:1,000, cat. no.
2774; Cell Signaling Technology, Inc.), Bcl-2 (1:1,000, cat. no.
2872, Cell Signaling Technology, Inc.), glucose-regulated protein
78 (GRP78; 1:1,000; product code ab108615; Abcam), PERK (1:1,000;
cat. no. 5683; Cell Signaling Technology, Inc.), eIF2α (1:1,000;
cat. no. 5324; Cell Signaling Technology, Inc.), p-eIF2α (1:1,000;
cat. no. 3398; Cell Signaling Technology, Inc.), caspase-12
(1:1,000; product code ab62484; Abcam), cyclin B1 (1:1,000; cat.
no. 4138; Cell Signaling Technology, Inc.), p-CDK1 (Tyr15; 1:1,000;
cat. no. 4539; Cell Signaling Technology, Inc.), Cdc25C (1:1,000;
cat. no. 4688; Cell Signaling Technology, Inc.),
p21Waf1/Cip1 (1:1,000; cat. no. 2947; Cell Signaling
Technology, Inc.), p27Kip1 (1:1,000; cat. no. 3686; Cell
Signaling Technology, Inc.) and GAPDH (1:5,000; cat. no. 2118; Cell
Signaling Technology, Inc.), and mouse monoclonal antibody against
CDK1 (1:1,000; cat. no. 9116; Cell Signaling Technology, Inc.). The
protein bands were quantified using ImageJ software (version 1.52a;
National Institutes of Health). The phosphorylated protein bands
were normalized to their total protein levels, and the other bands
were normalized to GAPDH.
Statistical analysis
The results are presented as the mean ± standard
deviation (SD). All statistical analyses were performed by Prism
7.0 (GraphPad Software, Inc.) using one-way ANOVA followed by
Tukeys multiple comparisons test. Differences were considered to be
statistically significant at P<0.05. All experiments were
performed in triplicate.
Results
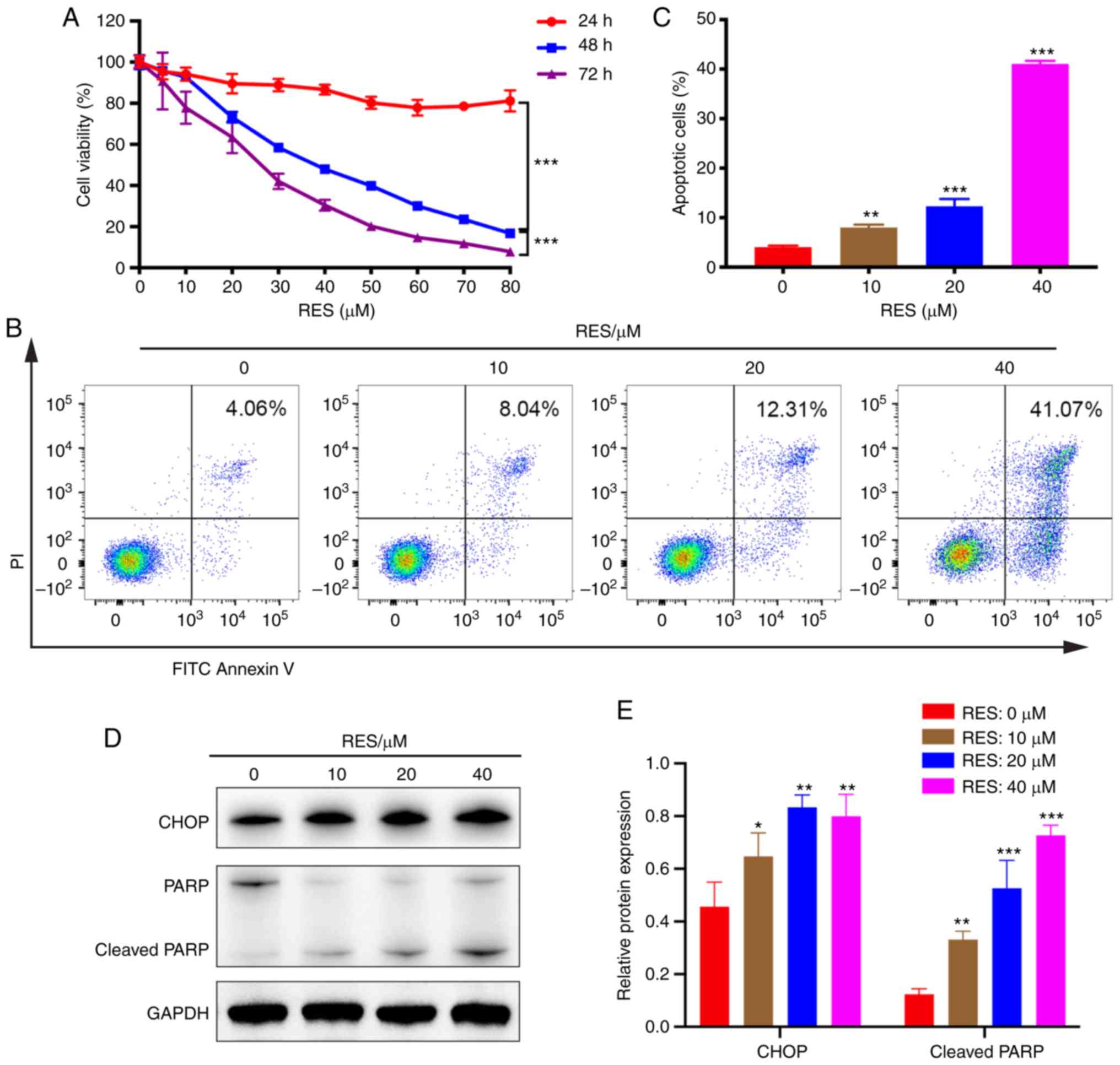
RES inhibits viability and promotes
apoptosis of AGS cells
First, cell viability assays in 4 GC cell lines
(AGS, KATO III, MKN-45 and NCI-N87) were performed and cell
apoptosis was detected in 3 GC cell lines (AGS, KATO III and
MKN-45) following RES/DDP cotreatment (Figs. S1, 2B and 3A-C). It was determined that the most
significant results were obtained in AGS cells. Therefore, the
human gastric adenocarcinoma cell line AGS was selected for the
present study. After exposure to RES, the cell viability of AGS was
explored by CCK-8 assay. The RES concentration gradient ranged from
5 to 80 µM. Our results demonstrated that RES displayed a dose-and
time-dependent inhibitory effect on AGS cellular viability
(P<0.001; Fig. 1A). Flow
cytometric analysis indicated that RES treatment (10, 20 and 40 µM)
consistently upregulated AGS cell apoptosis in a dose-dependent
manner, with the percentage of cells undergoing apoptosis
increasing from 4.06 to 41.07% with increasing RES concentration
(P<0.001; Fig. 1B and C).
Furthermore, the protein expression of CHOP, an early protein in ER
stress, was detected after administration of RES and DDP alone or
in combination for 24, 36, and 48 h. The most notable results were
obtained for the 48-h time-point (data not shown). Thus, this
time-point was selected for subsequent experiments. Western blot
analysis revealed that RES administration (10, 20 and 40 µM)
significantly increased the protein levels of CHOP and cleaved PARP
in a dose-dependent manner (P<0.05; Fig. 1D and E). Collectively, the present
results indicated that RES exerts anticancer activity against AGS
cells by inhibiting cell viability and promoting cell
apoptosis.
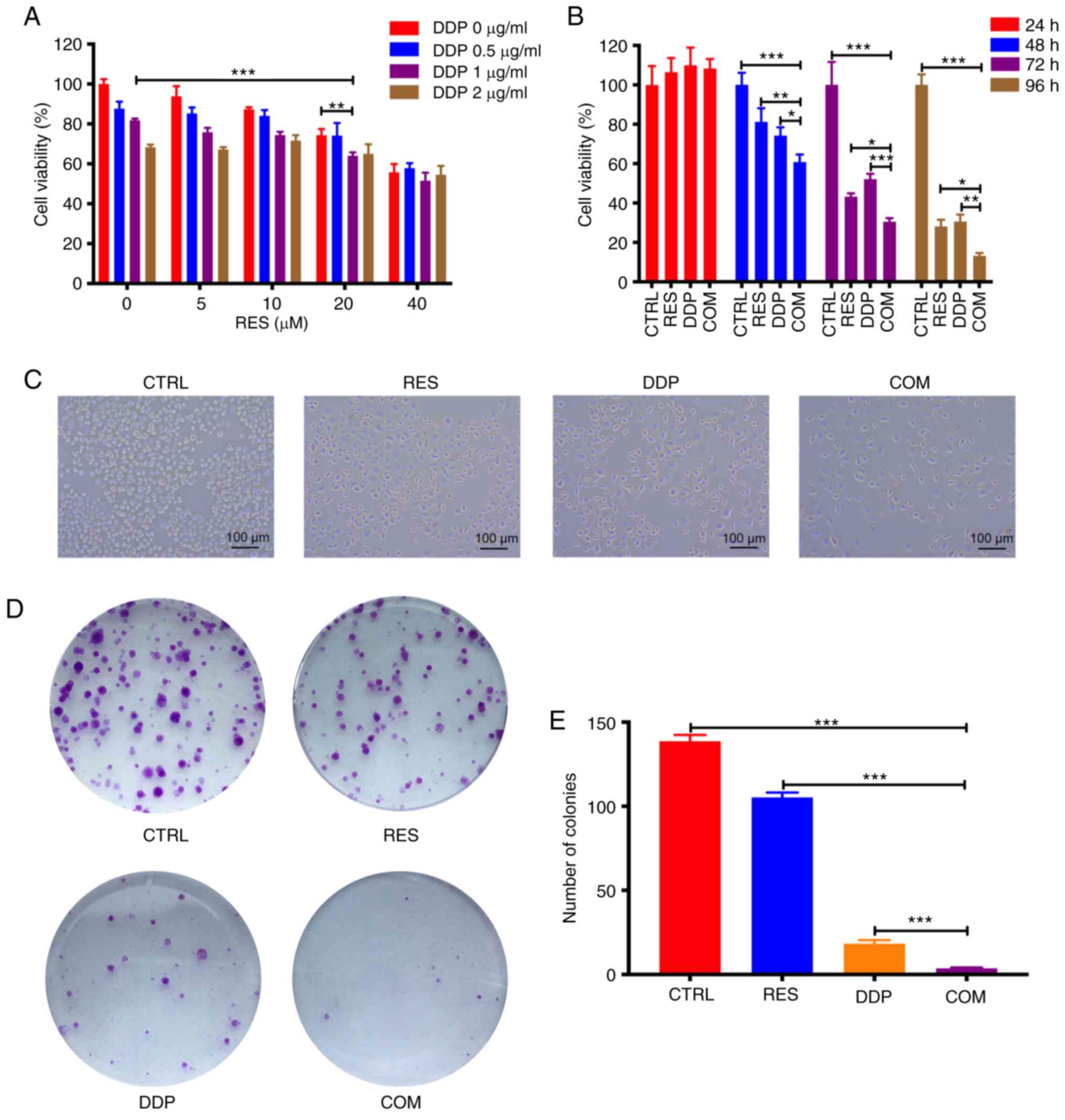 | Figure 2.RES and DDP synergistically inhibit
AGS cell viability. (A) Cell viability was determined by CCK-8
assay after cotreatment with different doses of RES and DDP for 48
h. (B) Following RES (20 µM) and DDP (1 µg/ml) combination
treatment for 24, 48, 72 and 96 h, cell viability was detected by
CCK-8 assay. (C) The morphological changes in AGS cells were
determined after RES (20 µM) and DDP (1 µg/ml) cotreatment for 48 h
(magnification, ×100). (D) Following RES (20 µM)/DDP (1 µg/ml)
combination treatment for 48 h, colony-forming assays were
performed. (E) Colonies in each well were counted. The results are
presented as the mean ± SD (n=3). *P<0.05, **P<0.01 and
***P<0.001. RES, resveratrol; DDP, cisplatin; CCK-8, Cell
Counting Kit-8; CTRL, control; COM, combination treatment. |
RES and DDP synergistically inhibit
AGS cell viability
To investigate the growth-inhibitory effect of
RES/DDP cotreatment on AGS cells, the cells were exposed to
different doses of RES (0, 5, 10, 20 and 40 µM) and DDP (0, 0.5, 1
and 2 µg/ml) alone or in combination for 48 h. The combination
treatment of RES (20 µM) and DDP (1 µg/ml) significantly suppressed
viability of AGS cells compared to the viability observed under
either RES treatment or DDP treatment alone (P<0.01; Fig. 2A). Therefore, 20 µM and 1 µg/ml were
selected as the concentrations for RES and DDP, respectively, for
the subsequent experiments. Next, cell viability was determined
following administration of RES (20 µM) and DDP (1 µg/ml) either
individually or in combination for 24, 48, 72 and 96 h. Although
exposure to RES (20 µM) or DDP (1 µg/ml) for 24 h did not
significantly inhibit cell viability, it was determined that RES
and DDP cotreatment for 48, 72 and 96 h significantly inhibited AGS
cell viability compared with that of either single-agent treatment
(P<0.05; Fig. 2B). As shown in
Fig. 2C, RES and DDP cotreatment
notably decreased the number of cells compared to the number
observed under either single-agent treatment alone. Besides, cell
shrinkage, detachment and reduced cytoplasm were evident in the
cells under RES/DDP combination treatment (Fig. 2C). In addition, the IC50
value against AGS cells decreased from 4.779 µg/ml under DDP-only
treatment to 3.154 µg/ml under combination DDP/RES (20 µM)
treatment (data not shown). Furthermore, the analysis of CI values
by CompuSyn software at ED50, ED75 and
ED90 (Table I) indicated
that RES and DDP have synergistic inhibitory effects on AGS cell
viability (22). The results of
colony-forming assays indicated that the combination treatment
significantly suppressed AGS cell viability over that attained by
RES or DDP treatment alone (P<0.001; Fig. 2D and E). Collectively, the present
results demonstrated that RES and DDP synergistically inhibit AGS
cell viability.
 | Table I.Combination index values of RES and
DDP combination treatment at ED50, ED75 and
ED90. |
Table I.
Combination index values of RES and
DDP combination treatment at ED50, ED75 and
ED90.
| RES (µM)/DDP
(µg/ml) |
ED50 |
ED75 |
ED90 |
|---|
| 25:1 | 0.931 | 0.867 | 0.910 |
| 12.5:1 | 0.911 | 0.853 | 0.862 |
RES sensitizes AGS cells to DDP by
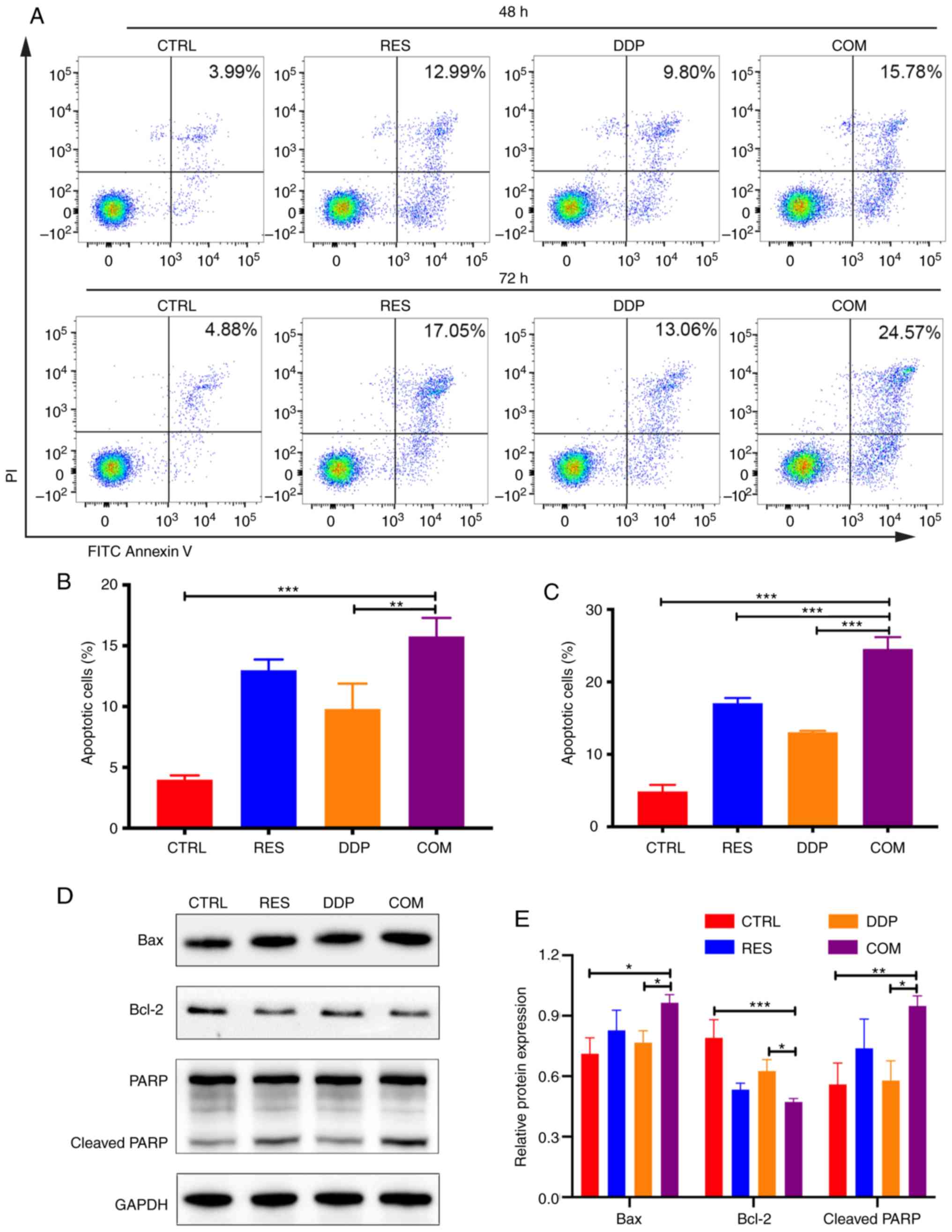
inducing cell apoptosis
FITC-Annexin V/PI double-staining assays were
conducted to assess apoptotic cells after cotreatment with RES (20
µM) and DDP (1 µg/ml) for 48 and 72 h (Fig. 3A). The present results demonstrated
that RES/DDP combination treatment for 48 h induced a higher
apoptotic rate compared with that of DDP treatment alone
(P<0.01; Fig. 3B). In
particular, when the cotreatment time was extended to 72 h, the
apoptotic rate was increased and was significantly higher than that
under either RES or DDP administration alone (P<0.01; Fig. 3C). To investigate the molecular
mechanisms underlying the proapoptotic effects of the RES/DDP
combination treatment, apoptosis-related proteins were analyzed by
western blotting. The results revealed that RES/DDP cotreatment
upregulated the levels of the proapoptotic protein Bax and the
cleaved form of PARP compared with the levels under DDP treatment
alone, whereas expression of the antiapoptotic protein Bcl-2 was
downregulated under cotreatment relative to DDP-only treatment
(P<0.05; Fig. 3D and E).
Collectively, the results indicated that RES enhanced the antitumor
effect of DDP on cell apoptosis.
RES/DDP combination treatment
activates ER stress-mediated apoptotic signaling pathways
To better understand the molecular mechanisms
underlying the proapoptotic effects of the RES/DDP combination,
western blotting was performed. The expression of the ER stress
chaperone GRP78 was significantly upregulated under RES/DDP
combination treatment compared to the expression under either
single-agent treatment alone (P<0.001; Fig. 4A and B). Moreover, cotreatment with
RES and DDP significantly increased PERK, p-eIF2α and CHOP protein
levels but had no significant effect on the expression of eIF2α,
indicating that the PERK/eIF2α/ATF4/CHOP signaling pathway was
activated by cotreatment (P<0.05; Fig. 4A-C). In addition, cleaved caspase-12
was upregulated upon RES and DDP combination treatment compared
with its expression under DDP treatment alone (P<0.05; Fig. 4A and B). The enhancement of
intracellular Ca2+ levels induces ER stress and
apoptosis (25). Herein,
Ca2+ probes and flow cytometry were applied to evaluate
intracellular Ca2+ levels. It was revealed that exposure
to RES (20 µM) alone for 48 h caused an increase in Ca2+
levels compared with levels in untreated cells (P<0.05; Fig. 4D and E). However, the RES/DDP
combination treatment significantly enhanced the cytosolic
Ca2+ levels and exhibited the greatest increase compared
with the levels under either single-agent treatment alone
(P<0.001; Fig. 4D and E). These
results demonstrated that RES/DDP cotreatment activated ER
stress-mediated apoptotic signaling pathways in AGS cells.
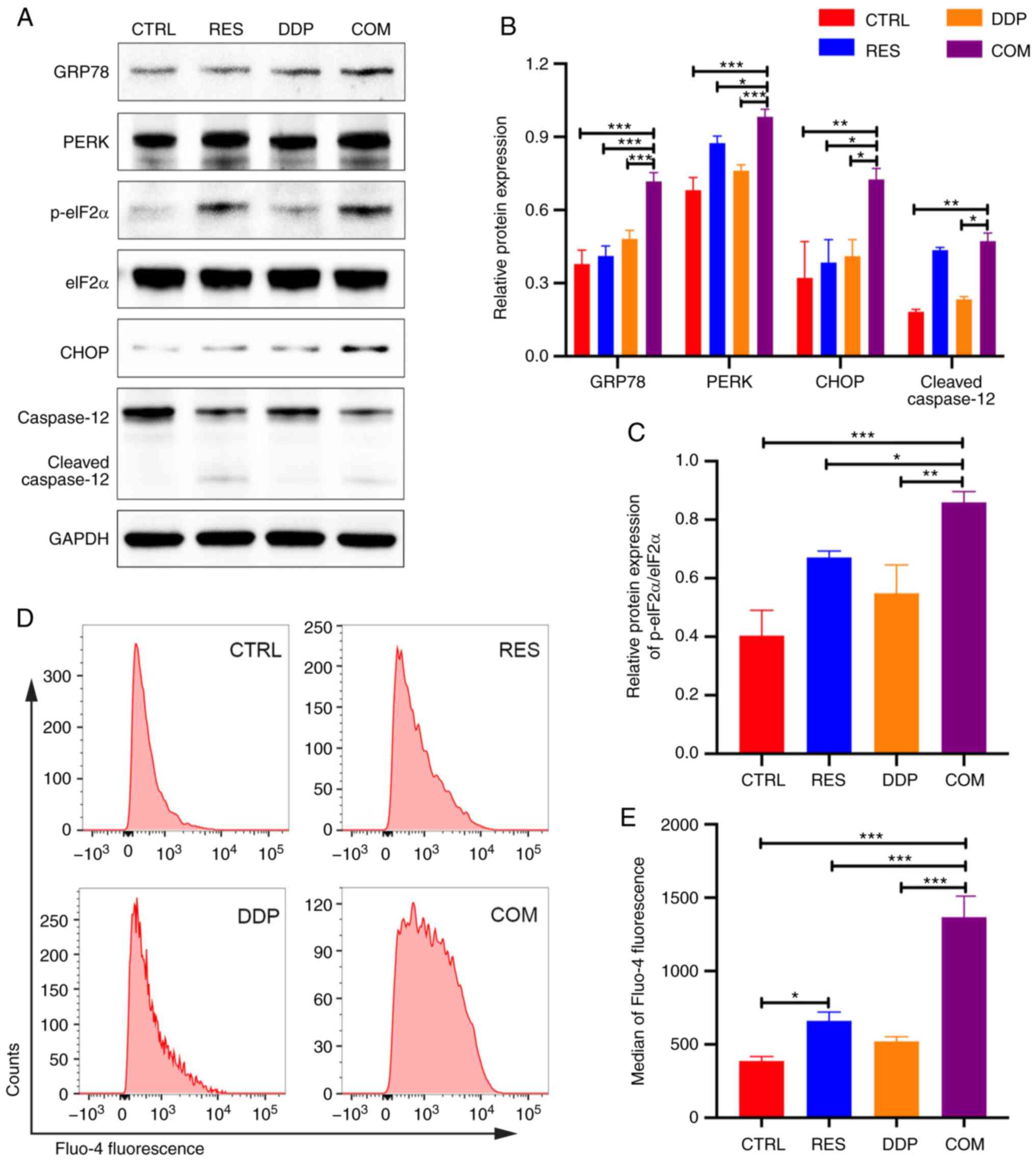 | Figure 4.RES/DDP combination treatment
activates endoplasmic reticulum stress-mediated apoptotic signaling
pathways. (A-C) The protein expression levels of GRP78, PERK,
p-eIF2α, eIF2α, CHOP and caspase-12 were determined by western
blotting. (D) Fluo-4 AM probe and flow cytometry were used to
detect intracellular Ca2+ concentrations after RES (20
µM) and DDP (1 µg/ml) cotreatment for 48 h. (E) The median Fluo-4
fluorescence intensities are presented. The data are presented as
the mean ± SD (n=3). *P<0.05, **P<0.01 and ***P<0.001.
RES, resveratrol; DDP, cisplatin; GRP78, glucose-regulated protein
78; PERK, PRKR-like ER kinase; p-eIF2α, phosphorylated eukaryotic
translation initiation factor 2α; CHOP, CCAAT/enhancer binding
protein homologous protein; CTRL, control; COM, combination
treatment. |
RES sensitizes AGS cells to DDP by
inducing G2/M phase arrest
To identify the effect of RES/DDP combination
treatment on cell cycle progression, PI staining assays and flow
cytometry were performed. As revealed in Fig. 5A-C, RES/DDP cotreatment
significantly increased the number of cells in the G2/M phase
compared to the number observed under either single-agent treatment
alone (P<0.001). Moreover, the expression levels of cell cycle
progression regulators were detected by western blotting. The
present results indicated that upon RES/DDP combination treatment,
the phosphorylation level of CDK1 (Tyr15) and the protein levels of
p21Waf1/Cip1 and p27Kip1 were significantly
increased, whereas Cdc25C protein levels were downregulated
(P<0.05; Fig. 5D-F). In
addition, it was determined that DDP administration significantly
upregulated cyclin B1 protein expression, while combination
treatment of DDP and RES significantly decreased the protein level
of cyclin B1 (P<0.001; Fig. 5D and
E). These results demonstrated that RES sensitizes AGS cells to
DDP by inducing G2/M phase arrest and by inactivating the
CDK1-cyclin B1 complex and upregulating p21Waf1/Cip1 and
p27Kip1 expression.
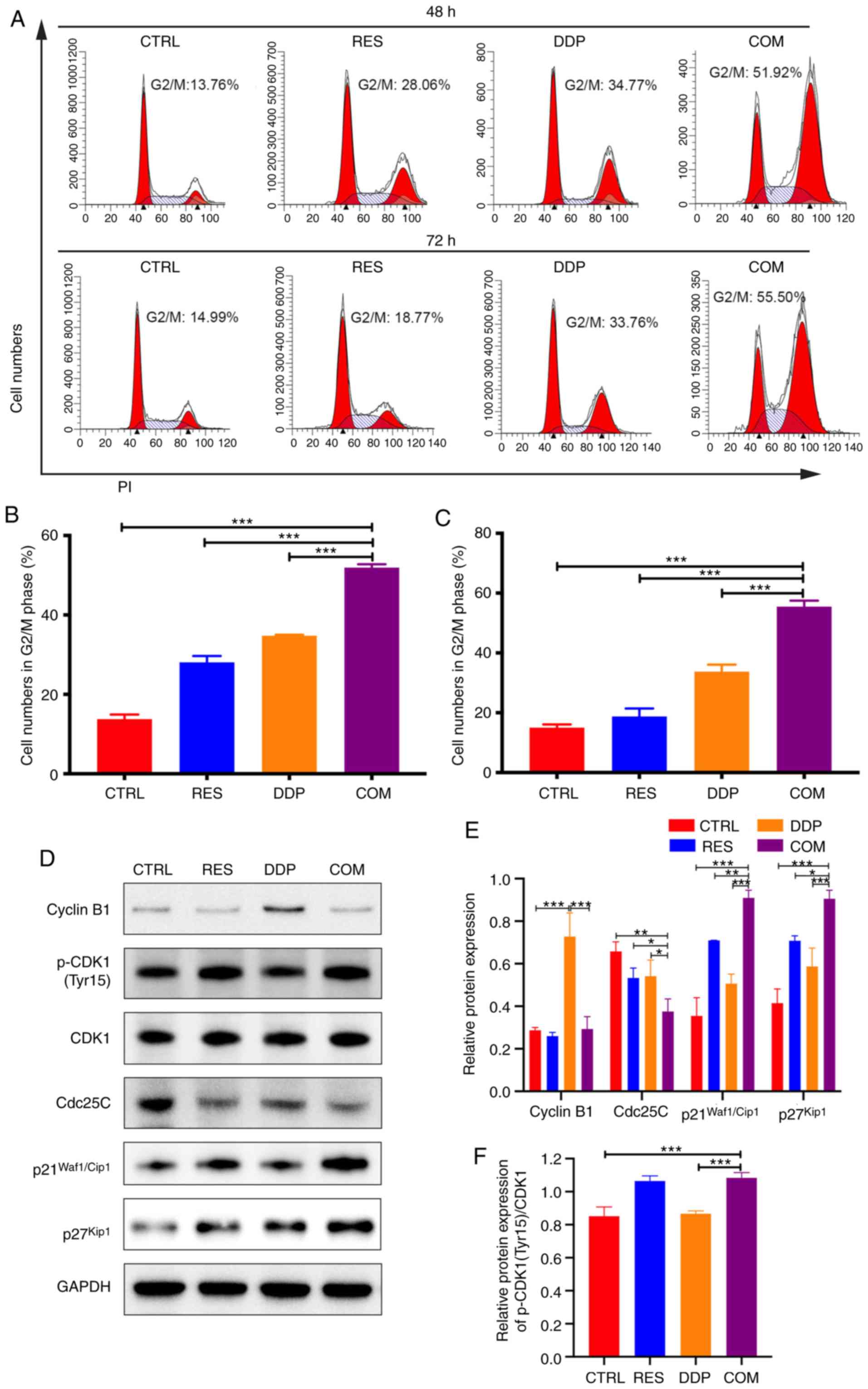 | Figure 5.RES sensitizes AGS cells to DDP by
inducing G2/M cell cycle arrest. (A) PI staining and flow cytometry
were performed to determine cell cycle progression after RES (20
µM) and DDP (1 µg/ml) cotreatment for 48 and 72 h. (B and C) The
percentage of cells in the G2/M phase was examined. (D-F) After RES
(20 µM) and DDP (1 µg/ml) combination treatment for 48 h, the
protein levels of cyclin B1, p-CDK1 (Tyr15), CDK1, Cdc25C,
p21Waf1/Cip1 and p27Kip1 were detected by
western blotting. All data are presented as the mean ± SD (n=3).
*P<0.05, **P<0.01 and ***P<0.001. RES, resveratrol; DDP,
cisplatin; p-CDK1, phosphorylated cyclin-dependent kinase 1; CTRL,
control; COM, combination treatment. |
Discussion
Severe side effects and resistance to chemotherapy
have been pressing issues hampering the success of GC treatment
(26). Hence, new effective
chemotherapeutic strategies are urgently required. RES has been
revealed to act as an antineoplastic drug in numerous types of
cancers, including GC (12–15,27).
However, whether RES and DDP have synergistic effects against GC,
and if so, what mechanisms underlie these effects are unknown. In
the present study, evidence was provided that RES and DDP
synergistically inhibited cell viability, prompted cell apoptosis
and induced G2/M cell cycle arrest in gastric adenocarcinoma AGS
cells. Furthermore, ER stress-induced apoptosis signaling pathways
were significantly activated and the CDK1-cyclin B1 complex was
inactivated under RES/DDP combined treatment.
The combination of naturally occurring agents with
conventional chemotherapeutic drugs has been applied in the
treatment of various types of cancers and can improve therapeutic
efficiency and reduce dosages to alleviate toxicity (28). Despite recent advances in molecular
targeted therapy and immunotherapy, DDP remains an important
component of chemotherapeutic regimens for GC (5,26).
However, its systemic toxicity and drug resistance impede its use.
RES, a natural polyphenol compound found in plants, is considered
to have potent anticancer activity against various types of cancers
(12–15,27).
Moreover, RES has been reported to act as a chemical sensitizer and
to enhance the anticancer effect of DDP in prostate cancer
(29), lung cancer (30,31)
and hepatoma (32). Herein, it was
revealed that RES exerted antineoplastic effects against the GC
cell line AGS by suppressing cell viability and promoting cell
apoptosis in a dose-dependent manner. Furthermore, the present
results demonstrated that RES and DDP synergistically inhibited
cell viability and promoted apoptosis of AGS cells, revealing RES
as a promising effective chemical sensitizer for DDP treatment in
GC patients.
Adverse conditions in the tumor microenvironment,
such as nutrient deprivation, oxidative stress, energy
perturbations and hypoxia, dysregulate proteostasis of the ER,
resulting in a cellular state termed ‘ER stress’ (33). ER stress triggers the UPR, which
subsequently induces prosurvival programs to maintain homeostasis
or activates apoptosis when damage is irreversible (34). The UPR is vital for cancer cell
survival in adverse environments and induces drug resistance to
conventional chemotherapy. Furthermore, blocking the adaptive
pathway or promoting the apoptotic pathway of the UPR is a
potential antineoplastic strategy since ER stress acts as a
‘double-edged sword’ during carcinogenesis (18). Previous studies have reported that
RES promotes ER stress-mediated apoptosis in colon cancer (19), ovarian cancer (20), and malignant melanoma (21,35).
However, the role of RES in regulating the UPR in GC cells is
unknown. In the present study, it was revealed that RES/DDP
combination treatment significantly upregulated the expression of
the ER stress hallmark GRP78 and significantly increased
cytoplasmic Ca2+ levels in AGS cells, indicating the
occurrence of ER stress under cotreatment.
The PERK/eIF2α/ATF4/CHOP signaling pathway is an
important modulator of ER stress-mediated apoptosis (16). As a UPR signal transducer on the ER
membrane, upon activation, PERK phosphorylates eIF2α at Ser51 to
decrease global protein translation but increase ATF4 mRNA
expression, thereby upregulating CHOP expression (36). CHOP is an ER stress-associated
apoptosis marker that further upregulates the expression of
proapoptotic proteins such as death receptor 5 (DR5) and Bim and
inhibits the expression of the antiapoptotic protein Bcl-2
(16). Caspase-12 is an ER
membrane-resident caspase that is upregulated only upon ER stress,
and its activation leads to cleavage of caspase-9 and caspase-3,
resulting in apoptosis (37).
Recently, it was reported that RES promotes ER stress-mediated
apoptosis in numerous types of cancer cells, such as colon cancer
(19), ovarian cancer (20), lung cancer (38), malignant melanoma (21) and nasopharyngeal carcinoma (39). The mechanisms included upregulation
of CHOP and activation of PERK and caspase-12. Herein, it was
revealed that RES administration upregulated CHOP expression in a
dose-dependent manner. Moreover, RES combined with DDP treatment
significantly activated the PERK/eIF2α/ATF4/CHOP signaling pathway
and induced caspase-12 cleavage in AGS gastric adenocarcinoma
cells. Therefore, it is suggested that RES/DDP combination
treatment promotes ER stress-mediated apoptosis in the GC cell line
AGS by activating the PERK/eIF2α/ATF4/CHOP signaling pathway and
caspase-12.
Dysregulation of cell cycle progression is a
hallmark of tumor development, and targeting the cell cycle is an
effective antineoplastic strategy (40). Unlike previous studies that revealed
that RES blocked GC cells at the G0/G1 phase (41), the present results revealed that
RES/DDP combination treatment significantly induced G2/M cell cycle
arrest. Moreover, Suttie et al (42) demonstrated that a low dose (0.156
µg/ml, LD10) of DDP arrested AGS cells in G0/G1 phase,
whereas a high dose (5 µg/ml, LD50) of DDP induced G2/M
phase arrest. In the present study, it was similarly revealed that
exposure to low dose of DDP (0.5 µg/ml) for 72 h induced G0/G1
phase arrest in AGS cells (data not shown). However, DDP treatment
(1 µg/ml) for 48 or 72 h led to G2/M cell cycle arrest. Based on
the previous literature and our experimental results, we speculate
that differences in cell cycle phase arrest in AGS cells are
associated with differences in the dose of DDP. In addition, the
proteins regulating cell cycle progression at the G2/M checkpoint
were detected by western blotting. The CDK1-cyclin B1 complex plays
a critical role in the G2/M transition (43). The downregulation or inactivation of
either CDK1 (also known as Cdc2) or cyclin B1 blocks cell cycle
progression to the G2 phase. Furthermore, the activity of CDK1 is
regulated by Cdc25C, a phosphatase, which dephosphorylates CDK1 at
Thr14 and Tyr15 residues, leading to the activation of the
CDK1-cyclin B1 complex (44).
p21Waf1/Cip1 and p27Kip1 are cyclin-dependent
kinase inhibitors (CKIs) that reduce the activity of CDK1 (45), and both are thought to be tumor
suppressive. In the present study, it was determined that RES/DDP
combination treatment upregulated the protein levels of p-CDK1
(Tyr15), p21Waf1/Cip1 and p27Kip,
downregulated Cdc25C expression, and reduced cyclin B1 protein
expression (which was increased by DDP administration alone),
leading to G2/M phase arrest.
There are certain limitations to the present study
worth mentioning. The primary limitation is that the target protein
and molecular mechanisms are currently not fully defined and
further research is required. In this study, we only presented the
possible molecular mechanisms underlying the antineoplastic effects
for RES/DDP cotreatment by activating ER stress-mediated apoptosis
and suppressing the activity of CDK1-cyclin B1 complex. Further
exploration of the specific mechanisms would be addressed in a
future study. In addition, we may also perform rescue experiments
to validate the specificity of the signaling pathways. Since
increase of Ca2+ during ER stress has been revealed to
be mainly associated with inositol trisphosphate receptors (IP3R)
(46), future experiments such as
blocking or downregulating IP3R are required to clarify the
relationship between RES/DDP combination treatment and
Ca2+ levels.
In conclusion, it was revealed that RES and DDP
synergistically inhibited cell growth of the GC cell line AGS by
inducing ER stress-mediated apoptosis and G2/M phase arrest.
Further experiments revealed that the PERK/eIF2α/ATF4/CHOP
signaling pathway and caspase-12 were activated by RES/DDP
cotreatment. Additionally, RES/DDP combination treatment suppressed
the activity of the CDK1-cyclin B1 complex and upregulated
p21Waf1/Cip1 and p27Kip1 expression to arrest
AGS cells in G2/M phase. These findings identify RES as a promising
adjuvant for GC chemotherapy.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank the State Key
Laboratory for Diagnosis and Treatment of Infectious Diseases of
the First Affiliated Hospital of Zhejiang University for excellent
technical assistance.
Funding
The present study was supported by Zhejiang
Traditional Chinese Medicine Science and Technology Project (grant
no. 2017ZZ010), the Key Research and Development Program of
Zhejiang Province (grant no. 2019C03031), and the Zhejiang Medical
Science and Technology Program (grant no. 2018266817).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
MR and XZ contributed equally to this study. MR, XZ
and MG designed and performed the experiments. WJ and JY performed
part of the experiments. MY, YW and SL contributed to the data
analysis. MR wrote the manuscript. XZ and MG reviewed and edited
the manuscript. FJ conceived the study and was responsible for the
revision of the manuscript and final decision to submit the article
for publication. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. Ca Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. Ca Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van Cutsem E, Sagaert X, Topal B,
Haustermans K and Prenen H: Gastric cancer. Lancet. 388:2654–2664.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang FH, Shen L, Li J, Zhou ZW, Liang H,
Zhang XT, Tang L, Xin Y, Jin J, Zhang YJ, et al: The Chinese
society of clinical oncology (CSCO): Clinical guidelines for the
diagnosis and treatment of gastric cancer. Cancer Commun (Lond).
39:102019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamada Y, Higuchi K, Nishikawa K, Gotoh M,
Fuse N, Sugimoto N, Nishina T, Amagai K, Chin K, Niwa Y, et al:
Phase III study comparing oxaliplatin plus S-1 with cisplatin plus
S-1 in chemotherapy-naive patients with advanced gastric cancer.
Ann Oncol. 26:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Quintanilha JCF, Saavedra KF, Visacri MB,
Moriel P and Salazar LA: Role of epigenetic mechanisms in
cisplatin-induced toxicity. Crit Rev Oncol Hematol. 137:131–142.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Berman AY, Motechin RA, Wiesenfeld MY and
Holz MK: The therapeutic potential of resveratrol: A review of
clinical trials. NPJ Precis Oncol. 1:352017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bonnefont-Rousselot D: Resveratrol and
cardiovascular diseases. Nutrients. 8:2502016. View Article : Google Scholar
|
|
9
|
Ahmed T, Javed S, Javed S, Tariq A, Šamec
D, Tejada S, Nabavi SF, Braidy N and Nabavi SM: Resveratrol and
Alzheimers disease: Mechanistic insights. Mol Neurobiol.
54:2622–2635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nunes S, Danesi F, Del Rio D and Silva P:
Resveratrol and inflammatory bowel disease: The evidence so far.
Nutr Res Rev. 31:85–97. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wong RHX and Howe PRC: Resveratrol
counteracts insulin resistance-potential role of the circulation.
Nutrients. 10:11602018. View Article : Google Scholar
|
|
12
|
Jiang Z, Chen K, Cheng L, Yan B, Qian W,
Cao J, Li J, Wu E, Ma Q and Yang W: Resveratrol and cancer
treatment: Updates. Ann N Y Acad Sci. 1403:59–69. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zulueta A, Caretti A, Signorelli P and
Ghidoni R: Resveratrol: A potential challenger against gastric
cancer. World J Gastroenterol. 21:10636–10643. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou HB, Chen JJ, Wang WX, Cai JT and Du
Q: Anticancer activity of resveratrol on implanted human primary
gastric carcinoma cells in nude mice. World J Gastroenterol.
11:280–284. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang Z, Xie Q, Chen Z, Ni H, Xia L, Zhao
Q, Chen Z and Chen P: Resveratrol suppresses the invasion and
migration of human gastric cancer cells via inhibition of
MALAT1-mediated epithelial-to-mesenchymal transition. Exp Ther Med.
17:1569–1578. 2019.PubMed/NCBI
|
|
16
|
Wang M and Kaufman RJ: Protein misfolding
in the endoplasmic reticulum as a conduit to human disease. Nature.
529:326–335. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hetz C and Papa FR: The unfolded protein
response and cell fate control. Mol Cell. 69:169–181. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim C and Kim B: Anti-cancer natural
products and their bioactive compounds inducing ER stress-mediated
apoptosis: A review. Nutrients. 10:10212018. View Article : Google Scholar
|
|
19
|
Park JW, Woo KJ, Lee JT, Lim JH, Lee TJ,
Kim SH, Choi YH and Kwon TK: Resveratrol induces pro-apoptotic
endoplasmic reticulum stress in human colon cancer cells. Oncol
Rep. 18:1269–1273. 2007.PubMed/NCBI
|
|
20
|
Gwak H, Kim S, Dhanasekaran DN and Song
YS: Resveratrol triggers ER stress-mediated apoptosis by disrupting
N-linked glycosylation of proteins in ovarian cancer cells. Cancer
Lett. 371:347–353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Heo JR, Kim SM, Hwang KA, Kang JH and Choi
KC: Resveratrol induced reactive oxygen species and endoplasmic
reticulum stressmediated apoptosis, and cell cycle arrest in the
A375SM malignant melanoma cell line. Int J Mol Med. 42:1427–1435.
2018.PubMed/NCBI
|
|
22
|
Chou TC: Drug combination studies and
their synergy quantification using the Chou-Talalay method. Cancer
Res. 70:440–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gu ML, Wang YM, Zhou XX, Yao HP, Zheng S,
Xiang Z and Ji F: An inhibitor of the acetyltransferases CBP/p300
exerts antineoplastic effects on gastrointestinal stromal tumor
cells. Oncol Rep. 36:2763–2770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang YM, Gu ML, Meng FS, Jiao WR, Zhou XX,
Yao HP and Ji F: Histone acetyltransferase p300/CBP inhibitor C646
blocks the survival and invasion pathways of gastric cancer cell
lines. Int J Oncol. 51:1860–1868. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bahar E, Kim H and Yoon H: ER
stress-mediated signaling: Action potential and ca(2+) as key
players. Int J Mol Sci. 17:15582016. View Article : Google Scholar
|
|
26
|
Wagner AD, Syn NL, Moehler M, Grothe W,
Yong WP, Tai BC, Ho J and Unverzagt S: Chemotherapy for advanced
gastric cancer. Cochrane Database Syst Rev.
8:CD0040642017.PubMed/NCBI
|
|
27
|
Athar M, Back JH, Kopelovich L, Bickers DR
and Kim AL: Multiple molecular targets of resveratrol:
Anti-carcinogenic mechanisms. Arch Biochem Biophys. 486:95–102.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin SR, Chang CH, Hsu CF, Tsai MJ, Cheng
H, Leong MK, Sung PJ, Chen JC and Weng CF: Natural compounds as
potential adjuvants to cancer therapy: Preclinical evidence. Br J
Pharmacol. 177:1409–1423. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martínez-Martínez D, Soto A, Gil-Araujo B,
Gallego B, Chiloeches A and Lasa M: Resveratrol promotes apoptosis
through the induction of dual specificity phosphatase 1 and
sensitizes prostate cancer cells to cisplatin. Food Chem Toxicol.
124:273–279. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li W, Shi Y, Wang R, Pan L, Ma L and Jin
F: Resveratrol promotes the sensitivity of small-cell lung cancer
H446 cells to cisplatin by regulating intrinsic apoptosis. Int J
Oncol. 53:2123–2130. 2018.PubMed/NCBI
|
|
31
|
Ma L, Li W, Wang R, Nan Y, Wang Q, Liu W
and Jin F: Resveratrol enhanced anticancer effects of cisplatin on
non-small cell lung cancer cell lines by inducing mitochondrial
dysfunction and cell apoptosis. Int J Oncol. 47:1460–1468. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Z, Peng Q, Li Y and Gao Y: Resveratrol
enhances cisplatin-induced apoptosis in human hepatoma cells via
glutamine metabolism inhibition. BMB Rep. 51:474–479. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cubillos-Ruiz JR, Bettigole SE and
Glimcher LH: Tumorigenic and immunosuppressive effects of
endoplasmic reticulum stress in cancer. Cell. 168:692–706. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Corazzari M, Gagliardi M, Fimia GM and
Piacentini M: Endoplasmic reticulum stress, unfolded protein
response, and cancer cell fate. Front Oncol. 7:782017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Limonta P, Moretti RM, Marzagalli M,
Fontana F, Raimondi M and Montagnani Marelli M: Role of endoplasmic
reticulum stress in the anticancer activity of natural compounds.
Int J Mol Sci. 20:9612019. View Article : Google Scholar
|
|
36
|
Urra H, Dufey E, Avril T, Chevet E and
Hetz C: Endoplasmic reticulum stress and the hallmarks of cancer.
Trends Cancer. 2:252–262. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rao RV, Ellerby HM and Bredesen DE:
Coupling endoplasmic reticulum stress to the cell death program.
Cell Death Differ. 11:372–380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gu S, Chen C, Jiang X and Zhang Z:
ROS-mediated endoplasmic reticulum stress and mitochondrial
dysfunction underlie apoptosis induced by resveratrol and arsenic
trioxide in A549 cells. Chem Biol Interact. 245:100–109. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chow SE, Kao CH, Liu YT, Cheng ML, Yang
YW, Huang YK, Hsu CC and Wang JS: Resveratrol induced ER expansion
and ER caspase-mediated apoptosis in human nasopharyngeal carcinoma
cells. Apoptosis. 19:527–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Otto T and Sicinski P: Cell cycle proteins
as promising targets in cancer therapy. Nat Rev Cancer. 17:93–115.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shin KO, Park NY, Seo CH, Hong SP, Oh KW,
Hong JT, Han SK and Lee YM: Inhibition of sphingolipid metabolism
enhances resveratrol chemotherapy in human gastric cancer cells.
Biomol Ther (Seoul). 20:470–476. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Suttie SA, Park KG and Smith TA:
[18F]2-fluoro-2-deoxy-D-glucose incorporation by AGS gastric
adenocarcinoma cells in vitro during response to epirubicin,
cisplatin and 5-fluorouracil. Br J Cancer. 97:902–909. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Satyanarayana A and Kaldis P: Mammalian
cell-cycle regulation: Several Cdks, numerous cyclins and diverse
compensatory mechanisms. Oncogene. 28:2925–2939. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Perdiguero E and Nebreda AR: Regulation of
Cdc25C activity during the meiotic G2/M transition. Cell Cycle.
3:733–737. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu X and Moscinski LC: Cdc2: A monopotent
or pluripotent CDK? Cell Prolif. 44:205–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Berridge MJ: Inositol trisphosphate and
calcium signalling. Nature. 361:315–325. 1993. View Article : Google Scholar : PubMed/NCBI
|