Introduction
Pancreatic cancer (PC) is a lethal disease, which
has a 5-year survival rate of <7% (1). Although surgery has improved the
overall survival (OS) of PC patients with early stage, the
prognosis of the majority of late stage PC patients is extremely
poor (2,3). The recently recommended first-line
chemotherapeutic regimen incorporating gemcitabine and paclitaxel
for PC patients who are not eligible for surgery, could prolong the
survival of PC patients. Gemcitabine is a deoxycytidine analogue
widely used for the therapy of PC patients and improved their
survival (4). Docetaxel, a
semi-synthetic analogue of paclitaxel, is a clinically well used
anti-mitotic chemotherapeutic drug for the treatment of various
carcinomas, including breast, ovarian, lung and pancreatic cancer
(1,2,5).
However, the unsatisfactory effects and unavoidable toxic side
effects are major obstacles for the present chemotherapeutic
regimens (1,2,6).
Therefore, these challenges highlight the importance of identifying
an alternative therapy with satisfactory effects and decreasing the
long-term toxic side effects.
Cell death forms include apoptosis and necrosis.
Apoptosis depends on initiator caspases (such as caspase-8 and −9)
and executioner caspases (caspase-3, −6 and −7) (7). Chemotherapeutic drugs usually induce
apoptosis of cancer cells through PARP-caspase3 pathway (8–10).
Curcumin was considered a promising agent in treating various types
of cancer by inducing apoptosis of cancer cells (11–13).
To elucidate the mechanisms of curcumin inducing apoptosis on PC
cells, we detected PARP-caspase3 pathway on three types of cell
lines.
Detached cancer cells are different from the normal
cells undergoing anoikis after dropping off their primary site and
this phenomenon is beneficial for metastasis (14,15).
Curcumin was reported to be able to enhance chemotherapeutic
effects on anoikis-resistant cancer cells (16), which would suppress metastasis of
cancer cells. Metastasis remains a serious problem affecting PC
treatment efficiency, and metastasis prevention is a promising
strategy for PC treatment and curcumin was found to be able to
inhibit cancer metastasis (17–19).
In the present study, the property of curcumin was found to
suppress PC cell metastasis and potentiate anti-metastasis
chemotherapeutic effects.
Curcumin, a hydrophobic polyphenol extracted from
dried rhizomes of turmeric, is a kind of traditional Chinese herbal
medicine and a potential drug for cancer therapy (20). A growing body of evidence has
demonstrated that curcumin has a potent anti-cancer effect for PC
treatment by inducing cell apoptosis, inhibiting cell
proliferation, and suppressing cell migration and invasion of PC
cells in pre-clinical studies (21). However, whether curcumin has
synergistic effects with either gemcitabine or docetaxel for the
treatment of PC remains elusive.
In the present study, we determined whether curcumin
has a synergistic effect with gemcitabine or docetaxel on cell
proliferation, apoptosis, migration, and invasion of PC cells.
Furthermore, the possible underlying mechanisms of the combination
treatment regimens were investigated. The results indicate that
curcumin is a promising adjuvant with the capacity of improving the
anti-cancer effects of either gemcitabine or docetaxel on PC cells
in vitro.
Materials and methods
Cell lines and reagents
Human pancreatic cell lines PANC-1, HPAF-II and
MIAPaCa-2 were purchased from the Chinese Academy of Life Science.
DMEM culture medium and fetal bovine serum (FBS) were purchased
from HyClone Laboratories Inc. and Gibco Company, respectively.
Curcumin was purchased from Xinran Company. Gemcitabine and
docetaxel were obtained from Sigma-Aldrich and dissolved in
DMSO.
Cell culture
PANC-1, HPAF-II and MIAPaCa-2 cells were cultured in
DMEM culture medium with 10% FBS and 1% penicillin and streptomycin
in an incubator with 5% CO2 at 37°C. Drugs were given at
the concentrations of 2, 5, 10, 20 and 50 µM for 48 h.
MTT assay
Drug sensitivity was detected using the MTT assay.
Briefly, cells were collected and cultured overnight, followed by
replenishment with fresh medium containing drugs and incubated at
37°C cell incubator with 5% CO2 for 48 h. A total volume
of 20 µl of MTT (Sigma-Aldrich) in PBS with the working
concentration of 5 mg/ml was added to the wells at the indicated
times. The wells were then incubated for an additional 4 h at room
temperature. The supernatant was discarded. A total volume of 100
µl of DMSO was added to the wells, followed by measurement using a
PerkinElmer 2030 VICTOR X Multilabel Plate Reader (Perkin-Elmer).
The results were collected from three independent experiments. The
percentage of live cells was represented as cell viability (%)=(OD
of treatment/OD of control) ×100. Three experimental repeats were
carried out to calculate cell viability. The average values of 50%
inhibiting concentration (IC50) of curcumin in PANC-1, MIAPaca-2
and HPAF-II were calculated from the viability values.
Analysis of cytotoxic synergy
The cell viability of PANC-1, HPAF-II and MIAPaCa-2
cells were determined by MTT assay. The CI values were then
calculated using Calcusyn 2.0 software. In detail, CI was detected
using the equation: (D)1/(Dx)1 + (D)2/(Dx)2 + α(D)1(D)2/(Dx)1(Dx)2,
where (Dx)1 and (Dx)2 are the doses for x% inhibition by drug 1 and
drug 2, respectively. (D)1 and (D)2 are representative of the
combinatory doses that inhibit cell growth by x%. A CI value of one
indicates additive effects of the two drugs and a CI value >1
indicates an antagonistic effect, while a CI value <1 suggests a
synergistic effect. Isobologram analysis in Fig. 2 shows a graphic representation of
the CI value, where CI value <1 is inside the triangle, CI value
>1 is outside the triangle, a CI value of one is on the
hypotenuse. Three experimental repeats were performed to calculate
CI values.
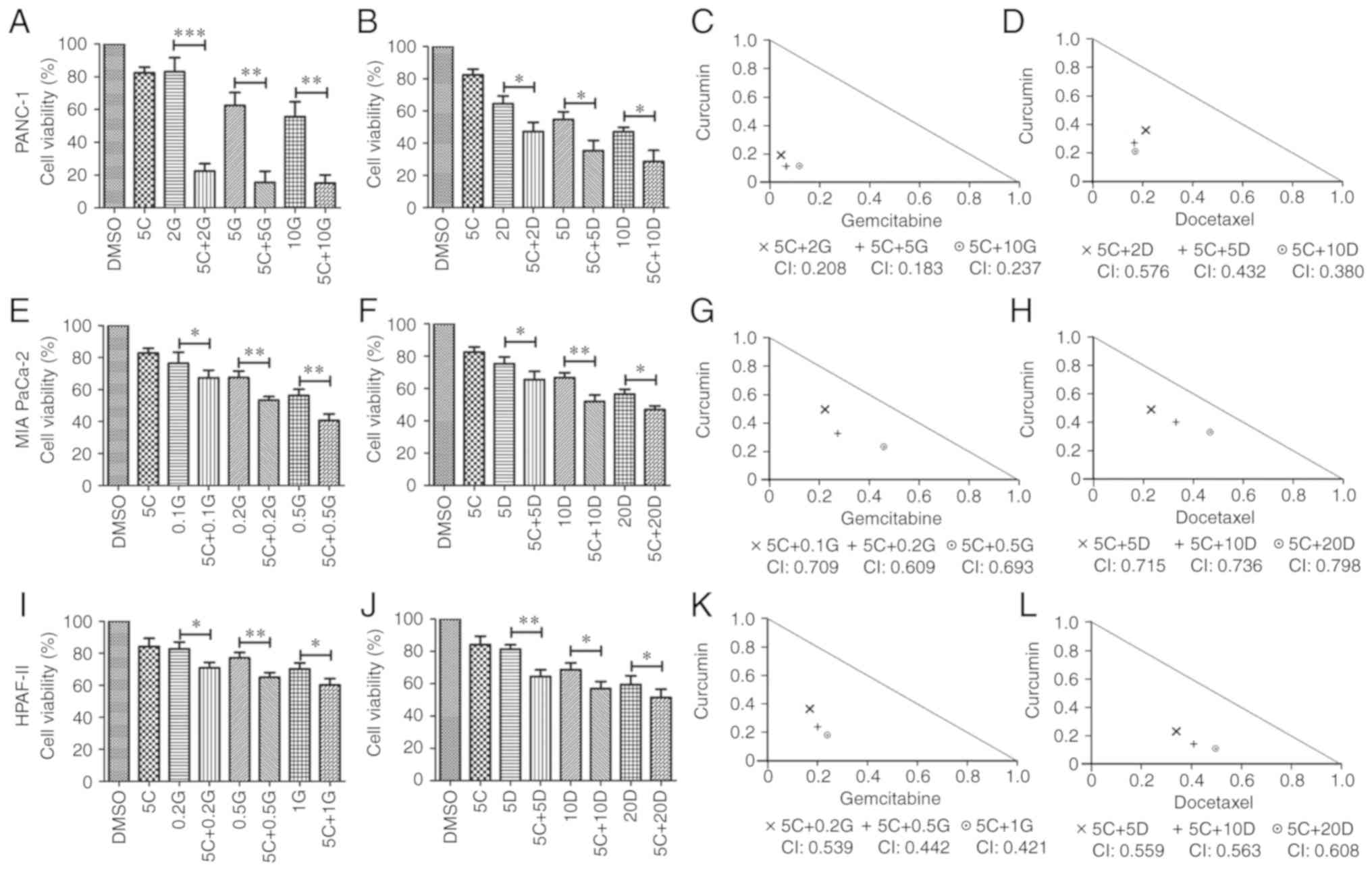 | Figure 2.Effects of curcumin with gemcitabine
or docetaxel on cell viability of PANC-1, MIA PaCa-2 and HPAF-II
cells. (A and B) Combinatory effects of curcumin with gemcitabine
or docetaxel on cell viability of PANC-1 cells were determined by
MTT assay. Data were collected from three independent experiments.
(C and D) Isobologram analysis was used to assess the synergistic
effects of curcumin with either gemcitabine or docetaxel on PANC-1
cells. The CI values indicate the synergistic effects at different
drug combinations. (E and F) Combinatory effects of curcumin with
either gemcitabine or docetaxel on cell viability of MIA PaCa-2
cells were determined by MTT assay. (G and H) Isobologram analysis
was used to assess the synergism of curcumin with either
gemcitabine or docetaxel on MIA PaCa-2 cells. The CI values depict
the synergistic effects at indicated combination drugs. (I and J)
Combinatory effects of curcumin with either gemcitabine or
docetaxel on cell viability of HPAF-II cells were determined by MTT
assay. (K and L) Isobologram analysis was conducted to evaluate the
synergism of curcumin with gemcitabine or docetaxel on HPAF-II
cells. The CI values depict the synergistic effects at different
drug combinations. All the experiments were conducted in
triplicate. CI, Combination index. 5C: 5 µM curcumin, 2G: 2 µM
gemcitabine, 5G: 5 µM gemcitabine, 10G: 10 µM gemcitabine, 5C+2G: 5
µM curcumin plus 2 µM gemcitabine, 5C+5G: 5 µM curcumin in
combination with 5 µM gemcitabine, 5C+10G: 5 µM curcumin in
combination with 10 µM gemcitabine, 2D: 2 nM docetaxel, 5D: 5 nM
docetaxel, 10D: 10 nM docetaxel, 5C+2D: 5 µM curcumin in
combination with 2 nM docetaxel, 5C+5D: 5 µM curcumin in
combination with 5 nM docetaxel, 5C+10D: 5 µM curcumin in
combination with 10 nM docetaxel. *P<0.05, **P<0.01 and
***P<0.001. |
DAPI staining assay
Live pancreatic cancer cells were plated in 6-well
plates for 24 h, followed by treatment with the indicated drugs.
After treatment for 48 h, the cells were fixed with 4%
paraformaldehyde for 20 min, followed by DAPI staining for 10 min
in the dark. Finally, cells were detected using immunofluorescence
microscopy (DSY5000X, OPPNO).
EdU (5-ethynyl-2′-deoxyuridine)
assay
Cell Light™ EdU Kit was purchased from RiboBio Co.,
Ltd. and the experiment was conducted according to the
manufacturer's instructions. Briefly, prepared 50 µM EdU DMEM
medium solutions were added to treated PC cells in 96-well plates
and incubated for 2 h, followed by washing with PBS twice. Then 4%
paraformaldehyde was used to fix the cells for 30 min and 2 mg/ml
glycine was used to neutralize the remaining paraformaldehyde.
Apollo staining reaction solution was used to incubate PC cells in
the dark for 30 min, followed by washing with 0.5% Triton X-100 PBS
three times. Finally, Hoechst-33342 was added for 30 min and images
were taken via immunofluorescence microscopy (DSY5000X, OPPNO).
Wound healing assay
PANC-1 cells were seeded in six-well plates,
followed by incubation for 24 h. Each well was initiated by
scratching with a sterile pipette tip, followed by washing with PBS
three times, and then treated with the indicated drugs in
serum-free medium for 24 h. Images at 0 and 24 h were taken using
an inverted fluorescence microscope (DSY5000X) at ×40
magnification. The blank area between two cell edges was calculated
via ImageJ software. Wound healing percentage was calculated using
the formula: [Blank area (0 h)-blank area (24 h)]/Blank area (0 h)
×100%.
Matrigel invasion assay
Cells were cultured in the culture medium in the
presence of drugs at the indicated concentrations for 48 h. DMSO
with the same volume was used as the control. Cells were then
trypsinized and resuspended into DMEM medium. The aforementioned
cells (5×104 cells per well with serum-free medium) were
plated in upper chamber coated with Matrigel (Corning). DMEM
culture medium containing 10% FBS was used as a chemoattractant in
the lower chamber. After incubation for an additional 24 h, the
invaded cells in the lower chamber were stained with 0.1% crystal
violet. Finally, cell images were obtained via light microscope
with charge-coupled device camera.
Transmission electron microscope
Cells were digested by 0.25% trypsin, followed by
centrifugation at 400 × g for 5 min at 4°C and fixation in 2.5%
glutaraldehyde overnight at 4°C. Next, the samples were fixed in 1%
osmium acid, followed by dehydration and embedding with fresh epon
resin, then incubated at 70°C vacuum oven for 2 days. Appropriate
areas of the samples were selected and ultrathin sections of 0.08
µm were stained with lead citrate and uranyl acetate for 5–10 min
at about 95°C empirically. Finally, these sections were determined
by a transmission electron microscope (TEM; JEM1230, Tokyo).
Western blotting
Cultured cells were treated with the indicated drugs
for 48 h, followed by lysing in RIPA buffer and denaturation.
Protein concentration was determined by bicinchoninic acid assay
system (Beyotime). Protein sample (50 µg per lane) was separated by
12% SDS-PAGE gel, followed by electrophoretical transfer to
nitrocellulose membranes. The membranes were then blocked with 5%
non-fat milk for 30 min at room temperature. Primary antibodies
including anti-caspase-3 (ABclonal, A2156), anti-cleaved-caspase-3
(ABclonal, A11021), anti-PARP (ABclonal, A19596), anti-cleaved-PARP
(ABclonal, A19612), anti-p-MLKL (Abcam, ab196436), anti-MLKL
(Abcam, ab184718), anti-N-cadherin (Abcam, ab76011),
anti-E-cadherin (Abcam, ab40772), anti-Vimentin (Abcam, ab92547),
anti-MMP2 (CST, 4022), anti-MMP9 (CST, 3852S), anti-TIMP1 (CST,
8946S) and anti-TIMP2 (CST, 5738S) antibodies were diluted in
primary antibody dilution buffer (Coolaber, SL1360) and incubated
with nitrocellulose membranes at 4°C overnight. Next, the membranes
were incubated with the corresponding secondary antibodies
conjugated with horseradish peroxidase at room temperature for 2 h,
followed by detection via an enhanced chemiluminescence detection
kit (Thermo Fisher Scientific). GAPDH (CST, 5174S) was used as the
control. Images were captured via a chemiluminescence imaging
system (ChemiScope 6000 Exp).
Statistical analysis
Data were analyzed using SPSS20.0 and presented as
mean ± standard deviation. ANOVA followed by the
Bonferroni-multiple comparison test was used for statistical
analysis to compare values among multiple groups. When the overall
difference across the multiple groups was significant,
Bonferroni-adjusted significance tests were used for pairwise
comparisons. P<0.05 indicated statistical significance.
Results
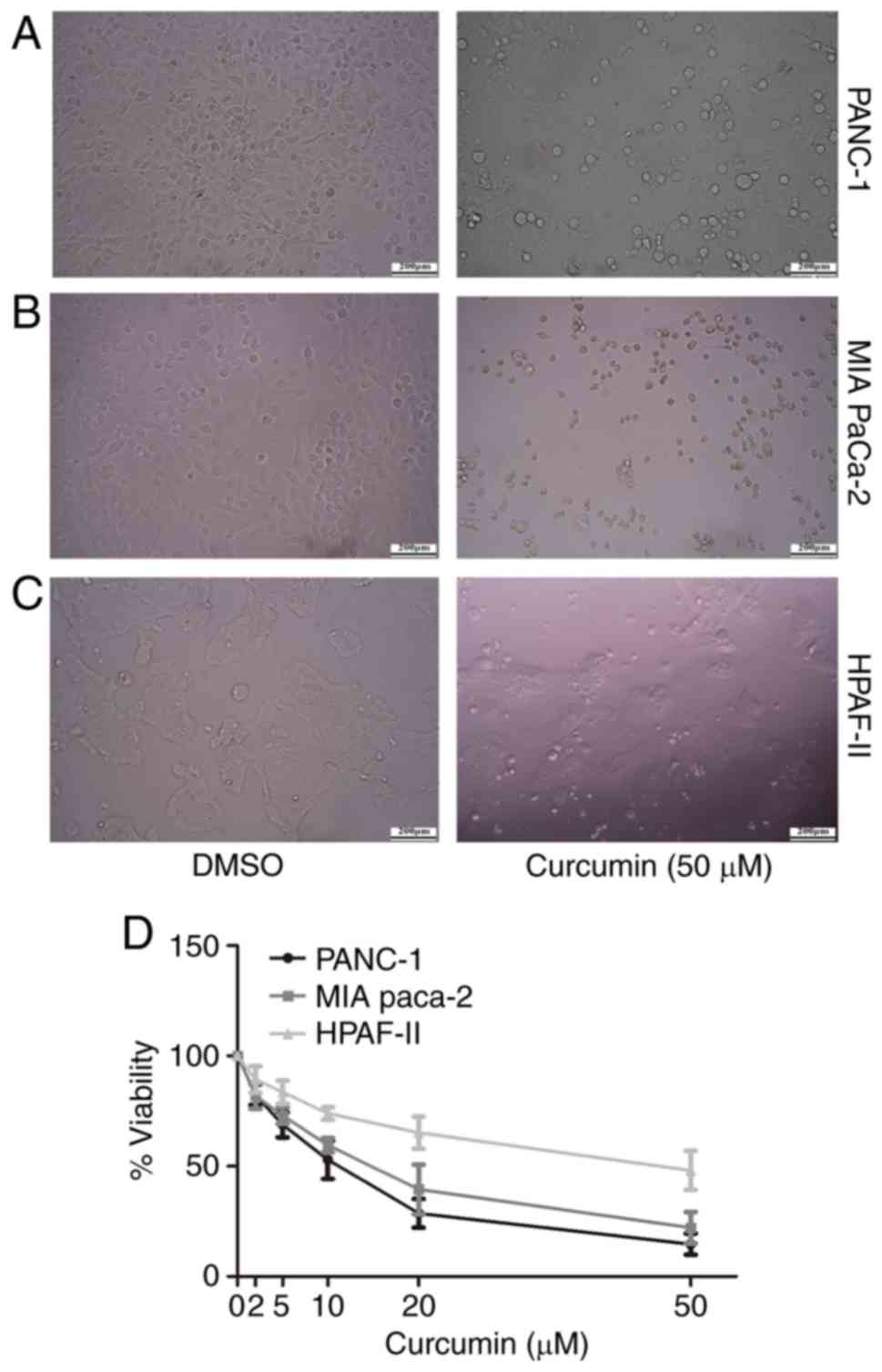
Curcumin induced morphologic changes
of PC cells and inhibited cell viability
We detected the morphologic changes of PANC-1,
MIAPaCa-2 and HPAF-II pancreatic cancer cells in the presence of
curcumin by inverted microscope. Cells treated with 50 µM curcumin
exhibited a different profile with a shrinkage of cell size, while
cells in the controlled group exhibited blurry features and
attached tightly to the well of the plate (Fig. 1A-C). The morphological changes are
more obvious with the concentrations of curcumin increasing, the
data of 0–20 µM are not shown. Next, we used MTT assay to detect
cell viability. As expected, cell viability was significantly
reduced in PC cells treated with curcumin. Notably, the suppressive
effects of curcumin on cell viability of PC cells occurred in a
dose-dependent manner (Fig. 1D).
The IC50 of curcumin in PANC-1, MIAPaca-2 and HPAF-II
were 9.87, 13.49 and 45.96 µM, respectively, in our detection.
Three experimental repeats were carried out to calculate IC
values.
Curcumin showed synergistic effects
with either gemcitabine or docetaxel on PC cells
To investigate whether curcumin has synergistic
effects with gemcitabine or docetaxel, we administered curcumin
together with gemcitabine or docetaxel of different concentrations
to PC cells. As shown in Fig. 2A,
gemcitabine at 2 µM alone mildly inhibited cell proliferation with
a cell viability of (83±5.027)%, while combination with curcumin (5
µM) induced significant inhibition on proliferation of PANC-1 cells
with viability of (22.33±2.656)%. Similarly, curcumin (5 µM) in
combination with gemcitabine (5, 10 µM) significantly inhibited
cell proliferation compared with gemcitabine (5, 10 µM) alone
[(15.52±3.928)% vs. (62.47±4.573)%, P<0.01; (15.13±2.852) vs.
(55.73±5.1)%, P<0.01]. The CI values of gemcitabine plus
curcumin were 0.208, 0.183 and 0.237 when they were given with the
ratios of 1:2.5, 1:1 and 2:1, respectively (Fig. 2C). Similarly, docetaxel had
synergistic effects with curcumin. The viability of PC cells
treated with curcumin (5 µM) plus docetaxel (2, 5, 10 nM) was
obviously decreased when compared to docetaxel alone (2, 5, 10 nM)
[(47.27±3.268)% vs. (64.57±2.735)%, P<0.05; (35.33±3.708) vs.
(54.67±2.751)%, P<0.05; (28.6±4.063) vs. (47±1.65)%, P<0.005]
(Fig. 2B). The CI values of
docetaxel (nM) plus curcumin (µM) were 0.576, 0.432 and 0.38 when
they were given with the ratios of 1:2.5, 1:1 and 2:1, respectively
(Fig. 2D). Consistently, similar
results were obtained from MIAPaCa-2 and HPAF-II cells (Fig. 2E-L). These aforementioned results
showed that curcumin has a synergistic effect with either
gemcitabine or docetaxel on suppressing cell viability of PC.
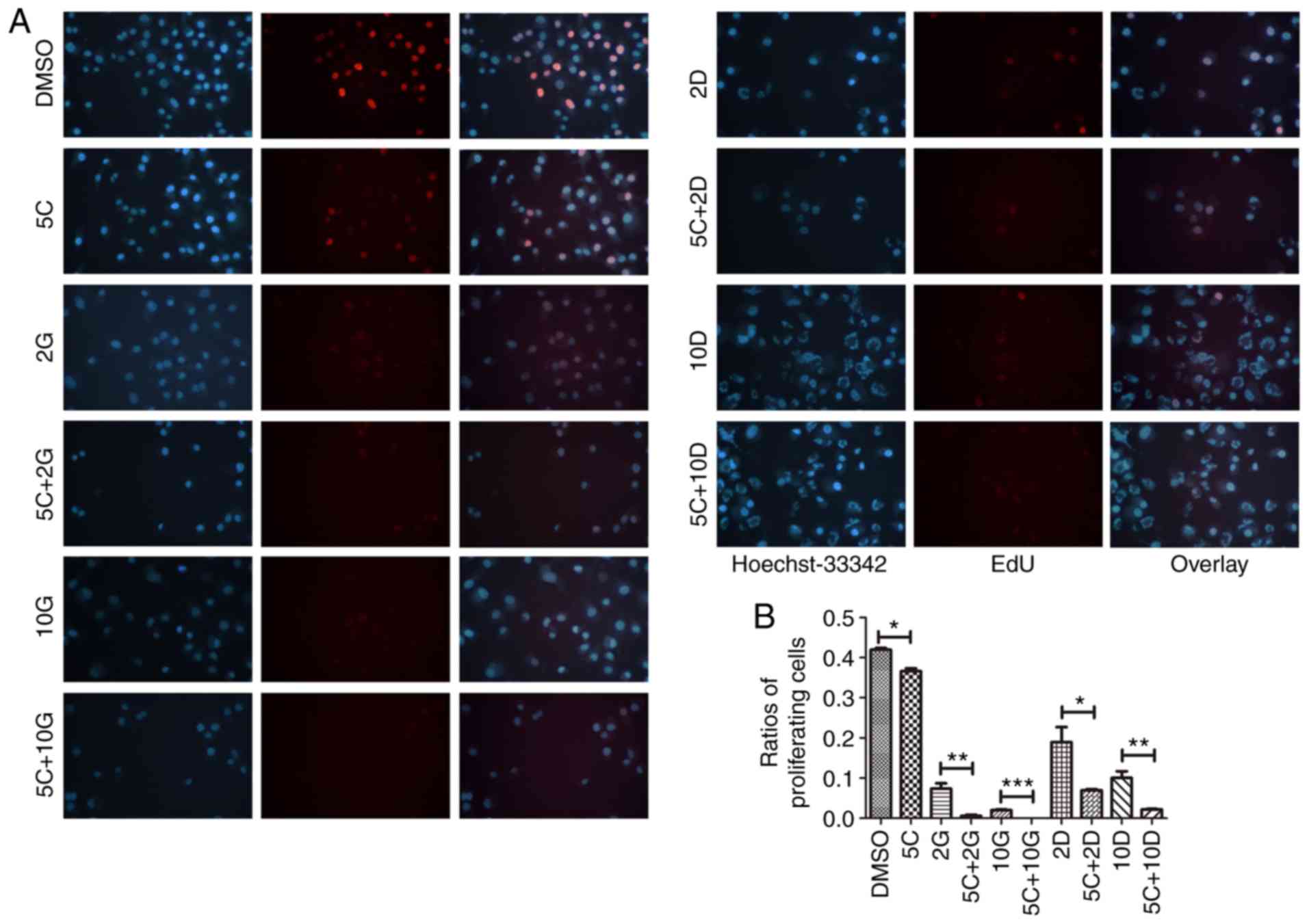
Curcumin enhanced the
anti-proliferation effects of either gemcitabine or docetaxel on PC
cells
Next, we determined whether curcumin could have
synergistic effects with gemcitabine or docetaxel on the
proliferation of PC cells. Relative proliferation ability was
assessed by the ratios of proliferating cells using EdU assay. The
proliferating cell treatment with curcumin (5 µM) was significantly
lower than that in the control group (P<0.05) (Fig. 3A and B). The proliferation of PC
cells treated with curcumin (5 µM) plus gemcitabine (2 µM) were
significantly decreased compared to gemcitabine alone (2 µM)
[(0.558±0.279) vs. (7.228±1.412)%, P<0.01]. Interestingly,
proliferation of PC cells in curcumin (5 µM) plus gemcitabine (10
µM) group was completely inhibited (Fig. 3A and B). Similarly, cell
proliferation after treatment with curcumin (5 µM) plus docetaxel
(2 or 10 nM) groups were significantly lower than docetaxel (2 or
10 nM) alone [(6.961±0.286) vs. (19.26±3.499)%, P<0.05;
(2.201±0.11) vs. (10.3±1.396)%, P<0.01] (Fig. 3A and B). Our results indicated that
curcumin is a potential adjuvant to enhance the anti-proliferation
effects of either gemcitabine or docetaxel for PC cells.
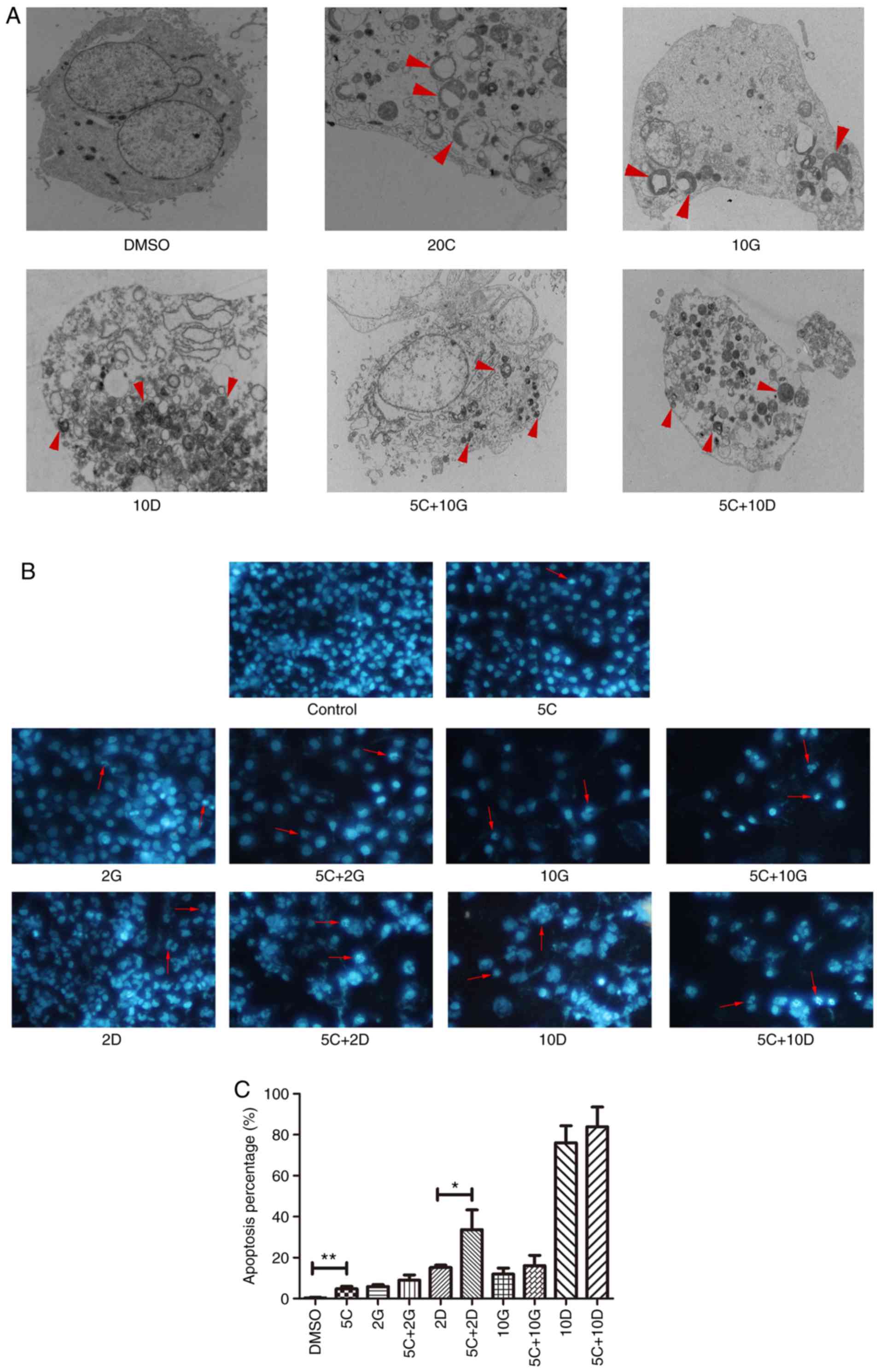
Curcumin induced apoptosis of PC cells
and enhanced the pro-apoptotic effects of either gemcitabine or
docetaxel on PC cells
To assess whether curcumin mediated suppression of
PC cell growth was correlated with cell apoptosis, we used TEM,
DAPI staining assay as well as western blot analysis after
treatment with 5, 10, 20 µM curcumin, 2, 10 µM gemcitabine, 2, 10
µM docetaxel, 5 µM curcumin plus 2, 10 µM gemcitabine or docetaxel
for 48 h. As expected, treatment with different drugs at the
indicated concentrations triggered increased morphological changes
of apoptosis and necrosis in PC cells by TEM (Fig. 4A). Curcumin at 5 µM induced
significantly increased cell apoptosis in PC cells compared with
the control [(4.633±1.11) vs. (0.327±0.119)%, P<0.001; Fig. 4B and C]. These data demonstrated
that curcumin could induce cell apoptosis and necrosis of PC cells
in vitro.
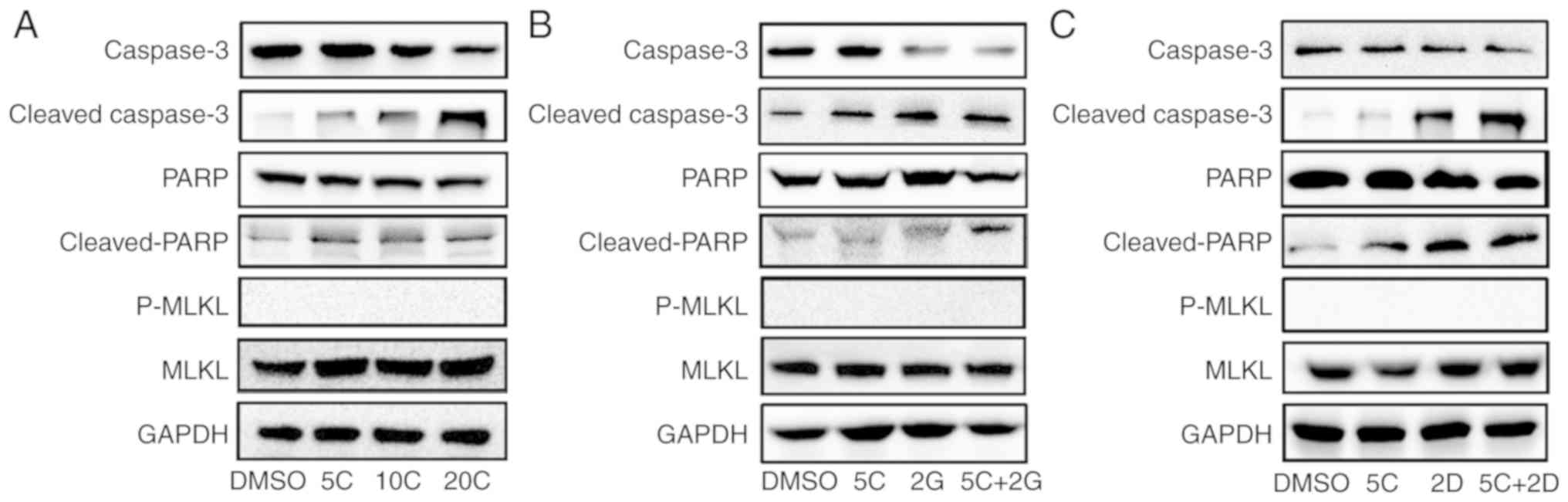
To further investigate the molecular mechanism of
apoptosis and necrosis induced by curcumin, western blotting was
used to determine the protein levels of classical apoptosis-related
molecules including caspase-3, cleaved-caspase-3, PARP and
cleaved-PARP, which were considered as classical apoptosis-related
molecules. The results showed that the expression level of
caspase-3 was downregulated, accompanied by concomitantly increased
expression level of cleaved-caspase-3 after treatment with curcumin
alone. Western blotting showed protein level of cleaved-PARP was
significantly increased after curcumin treatment, while there was
no significant change in PARP (Fig.
5A). These results indicated that the apoptosis-related
caspase-3/PARP signaling pathway may play a critical role in
curcumin-induced apoptosis of PC cells. p-MLKL, which was regarded
as a hallmark of necroptosis, was also detected by western blot
analysis in the present study. However, no significant change in
p-MLKL was observed (Fig. 5A).
Then, we investigated whether curcumin enhanced the
pro-apoptotic effects of gemcitabine or docetaxel on PC cells by
DAPI staining. Notably, treatment with curcumin (5 µM) together
with docetaxel (2 nM) induced significantly increased apoptosis of
PC cells in comparison with docetaxel (2 nM) alone [(33.73±4.787)
vs. (15.63±1.589)%; P<0.05]. Similar trends were observed in
other groups but no statistical significance (Fig. 4B and C). Additionally, we observed
significant apoptotic features of cells in each group treated with
drugs by TEM (Fig. 5A).
Furthermore, caspase-3/PARP signaling pathway was obviously
activated in the presence of gemcitabine plus curcumin or docetaxel
plus curcumin (Fig. 5B and C).
These results suggested that curcumin has a potent ability to
enhance the pro-apoptotic effects of chemotherapy drugs on PC cells
in vitro and exerts different influences on pro-apoptosis
effect of different drugs.
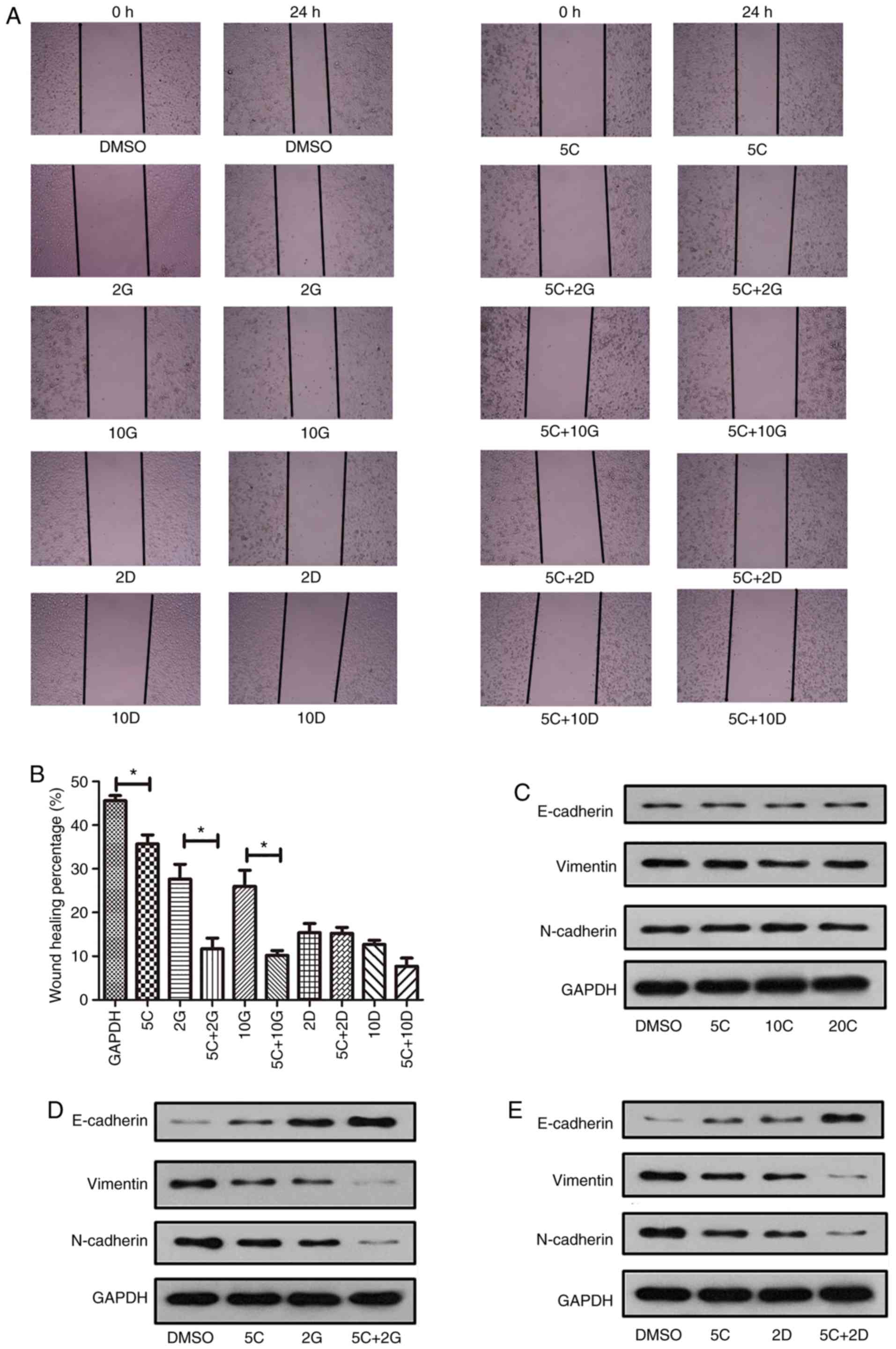
Curcumin inhibited PC cell migration
and benefitted the suppressive ability of gemcitabine or docetaxel
on cell migration
To further detect the effects of curcumin on cell
migration, we used wound healing assay to determine the migration
ability of PC cells under different drug treatment. As expected,
curcumin at 5 µM resulted in obvious reduction in recovery ratio of
PC cells in comparison with control [(35.7±2.155) vs.
(46.43±1.105)%; P<0.05] (Fig. 6A and
B).
Next, we determined whether curcumin reinforced
suppressive ability of gemcitabine or docetaxel on cell migration
by using scratching assay. Treatment of curcumin (5 µM) together
with gemcitabine (2 µM) exhibited stronger ability to inhibit cell
migration in comparison with gemicitabine alone (2 µM)
[(11.67±3.159) vs. (28.17±2.906)%; P<0.05] (Fig. 6A and B). Similarly, a combination of
curcumin (5 µM) and gemcitabine (10 µM) showed significantly
suppressive effect on PC cells in comparison with gemcitabine alone
(10 µM) [(10.83±2.677) vs. (25.97±3.302)%; P<0.05] (Fig. 6A-C). Moreover, expression of
N-cadherin and Vimentin were obviously reduced in the gemcitabine
(2 µM) plus curcumin (5 µM) group (Fig.
6D), while the expression of E-cadherin was obviously increased
in the gemcitabine (2 µM) plus curcumin (5 µM) group (Fig. 6D). Although curcumin showed no
significant enhancement to inhibitory effect of docetaxel,
expression of N-cadherin and Vimentin were slightly reduced in the
docetaxel (2 nM) plus curcumin (5 µM) group, while E-cadherin
showed slight increase (Fig. 6E).
These results suggest that curcumin reinforced the ability of
gemcitabine to suppress migration of PC cells.
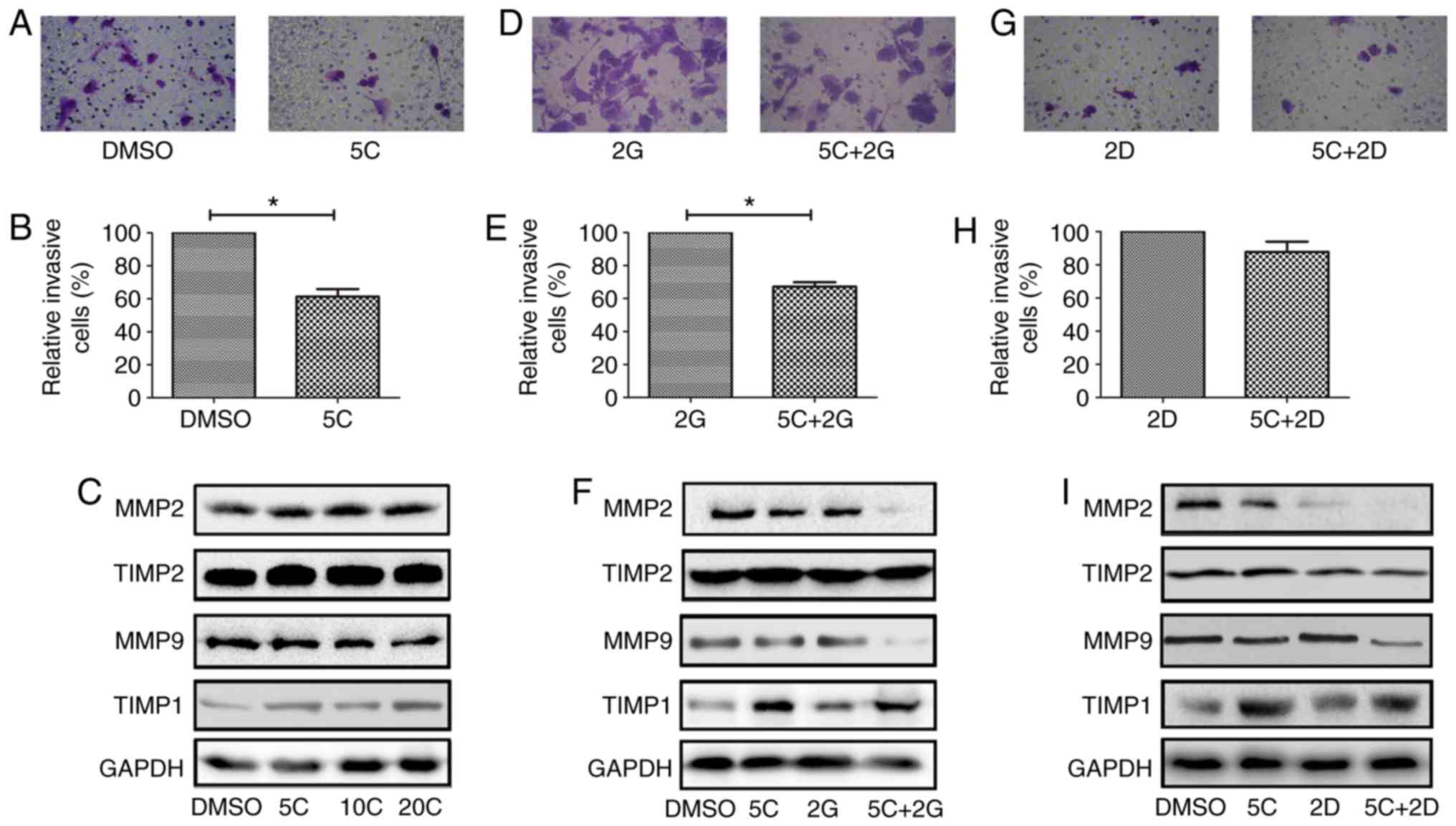
Combination application of curcumin
with gemcitabine or docetaxel inhibited invasion of PC cells
To gain a better understanding of the anti-invasion
ability of curcumin in combination with gemcitabine or docetaxel,
we conducted Matrigel invasion assay to assess the invasion ability
of PC cells under different drug combination. As expected, curcumin
at 5 µM induced an average of 38.67% reduction of invaded PC cells
compared with control (Fig. 7A and
B). Interestingly, TIMP1, a member of natural inhibitor for the
matrix metalloproteinases (MMPs), was upregulated in a
dose-dependent manner when treated with curcumin alone (Fig. 7C). Similarly, an average of 32.77%
reduction of invaded PC cells was found in curcumin (5 µM) plus
gemcitabine (2 µM) group compared with gemcitabine (2 µM) alone
(Fig. 6D and E). In addition, MMP2
and MMP9 expression was significantly reduced after treatment with
gemcitabine (2 µM) plus curcumin (5 µM) group while the expression
of TIMP1 was upregulated in curcumin (5 µM) group and gemcitabine
(2 µM) plus curcumin (5 µM) group (Fig.
7F). Treatment with curcumin (5 µM) plus docetaxel (2 nM)
showed little inhibition for invasion of PANC-1 cells compared with
docetaxel alone (2 nM), without statistical significance (Fig. 7G and H). Expression of MMP2 and MMP9
was obviously reduced in docetaxel (2 nM) plus curcumin (5 µM)
group, accompanied by the upregulation of TIMP1 (Fig. 7I). These results indicated that
curcumin would be a promising adjuvant to inhibit invasion of PC
cells.
Discussion
Increasing evidence has indicated that curcumin
exerts anti-tumor effects on the proliferation, apoptosis,
migration as well as invasion of PC cells (22–25).
Multiple signaling pathways may be involved in this circumstance,
including modulation of inhibitor of apoptosis proteins (IAP) and
downregulation of YAP/TAZ expression (26,27).
Other studies have shown that downregulation of NEDD4 with
concomitant upregulation of PTEN and p53 may be involved in this
procedure (28). Youns and
Fathy have demonstrated that activation of TNFR, CASP8,
CASP3, BID, BAX, and downregulation of NF-κB, NDRG1 and
BCL2L10 genes may be involved in curcumin-mediated effects
on PC cells (29). Our results were
in accordance with these findings. Curcumin was reported to produce
fluorescence emission (excitation spectra from 300 to 540 nm)
(26,30,31).
Consequently, we did not use flow cytometry to detect apoptosis but
selected TEM to detect apoptosis. In the present study, we have
found that caspase-3/PARP signaling pathway is involved in
curcumin-induced apoptosis of PC cells.
To the best of our knowledge, the current study has
several novelties. Firstly, the present study is the first to
determine the combinatory effects of curcumin plus docetaxel or
gemcitabine by using the CalcuSyn software. Secondly, we employed
three pancreatic cells lines with different genetic backgrounds to
test the combinatory effects of the drugs, providing convincing
data for the combinatory drug use. PANC-1 and MIA paca-2 have
different chromosomal aberrations, while HPAF-II is a kind of cell
line derived from pancreatic adenocarcinoma patients with
metastases. Notably, the combinatory treatment of gemcitabine or
docetaxel with curcumin could significantly strengthen this
process. Thirdly, necroptosis has been demonstrated to be involved
in some drug-induced tumor cell death and the p-MLKL is a marker of
necroptosis (32–35). Whether necroptosis participates in
the cell death caused by curcumin, gemcitabine or docetaxel is
unknown; however, we detected the marker p-MLKL to check the
necroptosis pathway, which is novel, compared to other similar
studies. However, we did not find significant change in the
expression of MLKL/p-MLKL, suggesting that necroptosis is not
involved in curcumin-induced death of PC cells.
Targeting metastasis is crucial to the treatment of
PC. Wang et al have reported that curcumin has the capacity
to inhibit mesenchymal transition (EMT) by targeting
cancer-associated fibroblasts (CAFs), which plays a pivotal role in
the metastasis of PC (36). In the
cancer process, it is considered that epithelial-derived cancer
cells are reversible, trans-differentiated and with low affinity in
cell-cell adhesion, and then disseminated through blood or
lymphatics to other sites via invasion (37,38).
E-cadherin, N-cadherin, Vimentin, MMP2, MMP9, TIMP1, TIMP2 are
common molecules involved in metastasis (38,39).
In addition to cell migration during metastasis, EMT also
influences resistance to anoikis and apoptosis, blocks senescence,
enhances survival, facilitates genomic instability, causes cancer
stem cell (CSC) activity, alters metabolism, and induces drug
resistance and immune suppression. Notably, both cadherin and
Vimentin seem to be involved in this procedure. Our results showed
that combinatory treatment significantly downregulated
MMP2/MMP9/N-cadherin/Vimentin and upregulated
TIMP1/TIMP2/E-cadherin. Therefore, curcumin is a potential
candidate to strengthen the current chemotherapeutic regimens for
metastatic PC. Yoshida et al have reported that curcumin can
sensitize chemo-resistant cancer cells via downregulating the
expression of EZH2 and lncRNA PVT1, suggesting that curcumin may
have the synergistic effects with gemcitabine on PC cells (40). In our experiments, curcumin
exhibited strong synergistic effects with either gemcitabine or
docetaxel on three PC cell lines by using Calcusyn software. The CI
values of gemcitabine plus curcumin with the ratios of 1:2.5, 1:1
and 2:1 were all <0.5, indicating the strong synergistic effects
of the two drugs. Similarly, docetaxel also exhibited a synergistic
effect with curcumin on PC cells. These data suggest that addition
of low dosage of curcumin into the chemotherapeutic regimens
containing gemcitabine or docetaxel for the treatment of PC
patients may be a promising strategy.
In conclusion, results of the present study have
demonstrated that curcumin has synergistic effects with either
gemcitabine or docetaxel on PC cells. Combination of
chemotherapeutic drugs with curcumin may be an alternative choice
for the treatment of clinical PC patients.
Acknowledgements
We would like to thank Professor Qui-ping Zhang of
Wuhan University School of Basic Medical Sciences for experiment
technical guidance. We would like to thank Yu-fang Zhu and Ming Xu
for assistance with cell culture.
Funding
This study was supported by the Fundamental Research
Funds for the Central Universities in Wuhan University (China)
(grant no. 2042019kf0131).
Availability of data and materials
The datasets used and/or analyzed in the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LS and XYL designed the study. XYL performed the
statistical analysis. PL, QY, HL, SQY and LPB acquired the data and
performed the experiments. PL analyzed the data and wrote the
manuscript. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Humphris JL, Johns AL, Simpson SH, Cowley
MJ, Pajic M, Chang DK, Nagrial AM, Chin VT, Chantrill LA, Pinese M,
et al: Clinical and pathologic features of familial pancreatic
cancer. Cancer. 120:3669–3675. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stan SD, Singh SV and Brand RE:
Chemoprevention strategies for pancreatic cancer. Nat Rev
Gastroenterol Hepatol. 7:347–356. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu P, Feng J, Sun M, Yuan W, Xiao R,
Xiong J, Huang X, Xiong M, Chen W, Yu X, et al: Synergistic effects
of baicalein with gemcitabine or docetaxel on the proliferation,
migration and apoptosis of pancreatic cancer cells. Int J Oncol.
51:1878–1886. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Von Hoff DD, Ervin T, Arena FP, Chiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang H, Ge W, Jiang W, Li D and Ju X:
SRPK1siRNA suppresses K562 cell growth and induces apoptosis via
the PARPcaspase3 pathway. Mol Med Rep. 17:2070–2076.
2018.PubMed/NCBI
|
|
9
|
Liu L, Han C, Yu H, Zhu W, Cui H, Zheng L,
Zhang C and Yue L: Chloroquine inhibits cell growth in human A549
lung cancer cells by blocking autophagy and inducing
mitochondrialmediated apoptosis. Oncol Rep. 39:2807–2816.
2018.PubMed/NCBI
|
|
10
|
Zhu W, Liang Q, Yang X, Yu Y, Shen X and
Sun G: Combination of sorafenib and Valproic acid synergistically
induces cell apoptosis and inhibits hepatocellular carcinoma growth
via down-regulating notch3 and pAkt. Am J Cancer Res. 7:2503–2514.
2017.PubMed/NCBI
|
|
11
|
Yang C, Ma X, Wang Z, Zeng X, Hu Z, Ye Z
and Shen G: Curcumin induces apoptosis and protective autophagy in
castration-resistant prostate cancer cells through iron chelation.
Drug Des Dev Ther. 11:431–439. 2017. View Article : Google Scholar
|
|
12
|
Giordano A and Tommonaro G: Curcumin and
cancer. Nutrients. 5:23762019. View Article : Google Scholar
|
|
13
|
Liczbinski P, Michalowicz J and Bukowska
B: Molecular mechanism of curcumin action in signaling pathways:
Review of the latest research. Phytother Res. 6:10022020.
|
|
14
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Simpson CD, Anyiwe K and Schimmer AD:
Anoikis resistance and tumor metastasis. Cancer Lett. 272:177–185.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
San TT, Khaenam P, Prachayasittikul V,
Sripa B, Kunkeaw N and Chan-On W: Curcumin enhances
chemotherapeutic effects and suppresses ANGPTL4 in
anoikis-resistant cholangiocarcinoma cells. Heliyon. 6:e032552020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia W, Deng F, Fu W, Hu J, Chen G, Gao X,
Tan X, Li G, Liu G and Zhu S: Curcumin suppresses wilms' tumor
metastasis by inhibiting RECK methylation. Biomed Pharmacother.
111:1204–1212. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bachmeier BE, Killian PH and Melchart D:
The role of curcumin in prevention and management of metastatic
disease. Int J Mol Sci. 9:17162018. View Article : Google Scholar
|
|
19
|
Coker-Gurkan A, Celik M, Ugur M, Arisan
ED, Obakan- Yerlikaya P, Durdu ZB and Palavan-Unsal N: Curcumin
inhibits autocrine growth hormone-mediated invasion and metastasis
by targeting NF-κB signaling and polyamine metabolism in breast
cancer cells. Amino Acids. 50:1045–1069. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yassine F, Salibi E and Gali-Muhtasib H:
Overview of the formulations and analogs in the taxanes' story.
Curr Med Chem. 23:4540–4558. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mantripragada KC and Safran H: Optimizing
initial chemotherapy for metastatic pancreatic cancer. Future
Oncol. 12:1125–1133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Azimi H, Khakshur AA, Abdollahi M and
Rahimi R: Potential new pharmacological agents derived from
medicinal plants for the treatment of pancreatic cancer. Pancreas.
44:11–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bar-Sela G, Epelbaum R and Schaffer M:
Curcumin as an anti-cancer agent: Review of the gap between basic
and clinical applications. Curr Med Chem. 17:190–197. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Devassy JG, Nwachukwu ID and Jones PJ:
Curcumin and cancer: Barriers to obtaining a health claim. Nutr
Rev. 73:155–165. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hosseini M, Hassanian SM, Mohammadzadeh E,
ShahidSales S, Maftouh M, Fayazbakhsh H, Khazaei M and Avan A:
Therapeutic potential of curcumin in treatment of pancreatic
cancer: Current status and future perspectives. J Cell Biochem.
118:1634–1638. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Diaz Osterman CJ, Gonda A, Stiff T,
Sigaran U, Valenzuela MM, Bennit HR, Moyron RB, Khan S and Wall NR:
Curcumin induces pancreatic adenocarcinoma cell death via reduction
of the inhibitors of apoptosis. Pancreas. 45:101–109. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou X, Su J, Feng S, Wang L, Yin X, Yan J
and Wang Z: Antitumor activity of curcumin is involved in
down-regulation of YAP/TAZ expression in pancreatic cancer cells.
Oncotarget. 7:79076–79088. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su J, Zhou X, Yin X, Wang L, Zhao Z, Hou
Y, Zheng N, Xia J and Wang Z: The effects of curcumin on
proliferation, apoptosis, invasion, and NEDD4 expression in
pancreatic cancer. Biochem Pharmacol. 140:28–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Youns M and Fathy GM: Upregulation of
extrinsic apoptotic pathway in curcumin-mediated antiproliferative
effect on human pancreatic carcinogenesis. J Cell Biochem.
114:2654–2665. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fujioka S, Sclabas GM, Schmidt C,
Frederick WA, Dong QG, Abbruzzese JL, Evans DB, Baker C and Chiao
PJ: Function of nuclear factor kappaB in pancreatic cancer
metastasis. Clin Cancer Res. 9:346–354. 2003.PubMed/NCBI
|
|
31
|
Aggarwal BB: Nuclear factor-kappaB: The
enemy within. Cancer Cell. 6:203–208. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang X, Yousefi S and Simon HU:
Necroptosis and neutrophil-associated disorders. Cell Death Dis.
9:1112018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Chen X, Gueydan C and Han J:
Plasma membrane changes during programmed cell deaths. Cell Res.
28:9–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yoon S, Kovalenko A, Bogdanov K and
Wallach D: MLKL, the protein that mediates necroptosis, also
regulates endosomal trafficking and extracellular vesicle
generation. Immunity. 47:51–65. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Weinlich R, Oberst A, Beere HM and Green
DR: Necroptosis in development, inflammation and disease. Nat Rev
Mol Cell Biol. 18:127–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Q, Qu C, Xie F, Chen L, Liu L, Liang
X, Wu X, Wang P and Meng Z: Curcumin suppresses
epithelial-to-mesenchymal transition and metastasis of pancreatic
cancer cells by inhibiting cancer-associated fibroblasts. Am J
Cancer Res. 7:125–133. 2017.PubMed/NCBI
|
|
37
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: Acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bhatia S, Wang P, Toh A and Thompson EW:
New insights into the role of phenotypic plasticity and EMT in
driving cancer progression. Front Mol Biosci. 7:712020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yoshida K, Toden S, Ravindranathan P, Han
H and Goel A: Curcumin sensitizes pancreatic cancer cells to
gemcitabine by attenuating PRC2 subunit EZH2, and the lncRNA PVT1
expression. Carcinogenesis. 38:1036–1046. 2017. View Article : Google Scholar : PubMed/NCBI
|