Introduction
Tobacco and alcohol consumption are well-recognized
risk factors for head and neck squamous cell cancer (HNSCC).
However, over the last few decades, oncogenic human papilloma virus
(HPV) has emerged as another major etiologic factor for HNSCC and
some other cancers (1). HPV is
responsible for 4.5% of all human malignancies and is associated
with 30% of cases of HNSCC (2). HPV
is an ancient, nonenveloped, circular double-stranded DNA virus
consisting of approximately 220 distinct HPV genotypes that have
been identified to date (3,4). The HPV genome is comprised of the early
genes E1, E2, E4, E5, E6, and E7, and late genes L1 and L2. HPV E6
and E7 are considered to be the main oncoproteins that manipulate
cellular signaling pathways to promote HPV-mediated carcinogenesis
(3,4).
HPV-associated HNSCC has been shown to differ from alcohol- and
tobacco-associated HNSCC in terms of molecular pathophysiology,
presentation and prognosis. Patients with HPV-related HNSCC
generally have less exposure to alcohol and tobacco, tend to be
younger, and have a better prognosis compared to HPV-negative HNSCC
(5). However, the molecular and
cellular alterations underlying the pathobiology of HPV-related
cancers remains ambiguous. While the prevalence of patients with
HPV-related cancers is rapidly increasing, there are still many
questions that need to be addressed for proper control of
HPV-positive HNSCC. Further studies comparing the cellular pathways
between HPV-negative and HPV-positive HNSCC in more detail would
help to elucidate strategies targeting cellular pathways unique to
HPV-positive HNSCC.
A recent study which employed comprehensive
molecular and histoepigenetic analysis with epigenomic
deconvolution has identified potential novel biomarkers and
therapeutic targets for HPV-positive HNSCC (6). In this report, receptor tyrosine kinase
like orphan receptor 2 (Ror2) was proclaimed as a novel therapeutic
target for HPV-positive HNSCC. Ror2 belongs to the group of
Ror-family receptors that regulate cellular proliferation,
differentiation, polarity, and migration (7). The Ror-family receptors, consisting of
Ror1 and Ror2, act as receptors for Wnt5a to mediate the
noncanonical Wnt signaling which plays essential roles during
developmental morphogenesis (7).
Although it has been shown that Ror2 is highly expressed in various
malignant tumor cells and is involved in tumor progression by
promoting proliferation, migration, and invasion of tumor cells
(8–10), the mechanisms by which Ror2 mediates
the promotion of tumor progression remain poorly understood.
Recently, we have shown that Ror2 plays a crucial role in
regulating the G1/S phase transition of bFGF-stimulated
NIH/3T3 fibroblasts (11); however,
it remains unclear whether Ror2 regulates the G1/S
transition of tumor cells. We have also shown that Ror2 expression
in oral cancer was significantly higher than that in normal oral
mucosa, and that Ror2 is associated with cellular polarity,
motility, and tumor aggressiveness (12). However, it is currently unknown
whether Ror2 is involved in the aggressiveness of HPV-related
HNSCC. Considering the apparent difference in cellular properties
between HPV-negative and HPV-positive HNSCC, the function of Ror2
in HPV-positive HNSCC might offer a unique platform for exploring
novel diagnostic and therapeutic targets.
Hence, the aim of the present study was to
investigate the role of Ror2 in the regulation of cellular
proliferation, apoptosis, and invasive migration of HPV-positive
HNSCC.
Materials and methods
Cell culture and transfection
The human HPV16-positive HNSCC cell line UPCI:SCC152
was obtained from the American Type Culture Collection (ATCC). The
human HPV-negative HNSCC cell lines UM-SCC-22B and Ca9-22 were
obtained from RIKEN Bio Resource Center (The Institute of Physical
and Chemical Research, Tsukuba, Japan). The short-tandem repeat
profiles of UM-SCC-22B and Ca9-22 were analyzed, and we confirmed
that these cell lines are not contaminated. These cells were
cultured in Dulbecco's modified Eagle's medium (DMEM; FUJIFILM Wako
Pure Chemical Corp.) supplemented with 10% FBS at 37°C. For siRNA
transfection, UPCI:SCC152 cells were transfected with the
respective siRNAs using Lipofectamine RNAiMAX reagent (Thermo
Fisher Scientific Inc.) according to the manufacturer's
instructions. siRNAs targeting human HPV16 E6/7 (Ambion,
Thermo Fisher Scientific, Inc.), Ror2 (Sigma-Aldrich; Merck
KGaA), and their control siRNA (Silencer Select Negative Control
No. 1, cat. no. 4390843, Thermo Fisher Scientific Inc.; Mission
siRNA Universal Negative Control, cat. no. SIC_001: Sigma-Aldrich;
Merck KGaA) were used. The following target sequences were used for
the respective siRNAs: si-Ror2#1, 5′-GCAAUGUGCUAGUGUACGA-3′;
si-Ror2#2, 5′-CCUUUGAGAAGACACCAUA-3′; si-E6/7#1,
5′-CCGGACAGAGCCCAUUACA-3′; si-E6/7#2,
5′-CACCUACAUUGCAUGAAUA-3′.
Flow cytometric analysis
For cell cycle analysis, cells were synchronized
using single thymidine block. UPCI:SCC152 cells were seeded onto
6-cm (φ) dishes and were transfected with Ror2 siRNA or
control siRNA. Forty-eight hours after transfection, the cells were
arrested at the G1/S phase by treatment with 2 mM
thymidine for 24 h. Synchronized cells were released by two washes
with phosphate-buffered saline (PBS) and adding fresh medium. Cells
were fixed in 70% (v/v) ethanol and treated with 100 µg/ml RNase A
(Thermo Fisher Scientific, Inc.) and 50 µg/ml propidium iodide (PI)
(Sigma-Aldrich; Merck KGaA). DNA contents were analyzed using a
BD-LSRFortessa™ X-20 flow cytometer (BD Biosciences).
For the apoptosis assay, cells transfected with
either Ror2 siRNA or control siRNA were synchronized using a
single thymidine block as described above. Synchronized cells were
resuspended in 100 µl of binding buffer containing 5 µl
FITC-Annexin V and 5 µl PI, following the manufacturer's
instructions (BD Biosciences). Following a 15-min incubation period
at room temperature in the dark, 400 µl of binding buffer was added
to the cell suspension and the samples were analyzed using a
BD-LSRFortessa X-20 flow cytometer (BD Biosciences) within 1 h.
WST-8 assay
The WST-8 assay was carried out using a Cell
Counting Kit-8 (Dojindo, Kumamoto, Japan), according to the
manufacturer's instructions. In brief, 2 days after transfection
with the indicated siRNAs, 1,000 cells were seeded onto a 96-well
plate in triplicate. After the cells were cultured for the
indicated periods of time, WST-8 reagent was added to the culture
media. After incubation for 3 h, the absorbance at 450 nm was
measured.
Invasion assay
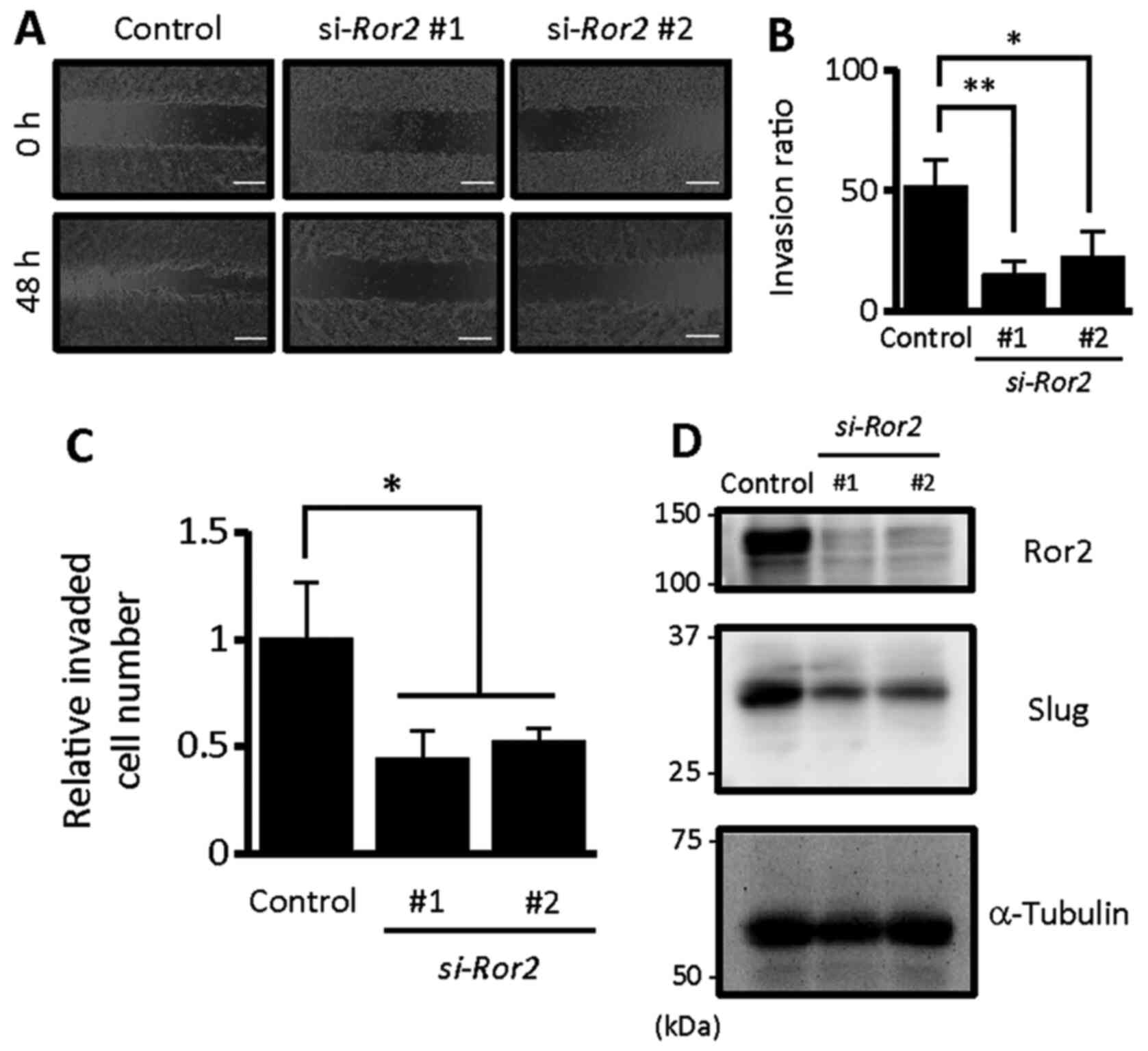
The Matrigel invasion assay was carried out as
described previously (13). Briefly,
2.5×104 cells in serum-free DMEM were loaded into the
upper wells of a Transwell chamber (8 µm φ pore size, 24 well;
Corning, Inc.) precoated with Matrigel (1:30 in DMEM). After 24 h,
the invasive cells on the lower surface of the membrane were
stained with DAPI and observed by using a fluorescence microscope
BZ-X710 (Keyence, Osaka, Japan). The number of DAPI-stained cells
was counted by using ImageJ software 1.53c (National Institutes of
Health, Bethesda, MD, USA).
Semi-3D invasion assay
Semi-3D invasion assay was carried out as described
previously (14). In brief, the
2-well culture insert with 0.5 mm gap between wells (ibidi,
Germany) was placed on a fibronectin coated 24-well plate. Cells
transfected with the respective siRNAs were seeded onto the culture
insert and grown to confluent monolayers. After removing the
insert, the monolayers were overlaid with Matrigel (BD
Biosciences), followed by incubation for 4 h before an addition of
medium. Cells were then cultured for 48 h to allow invasion toward
the space between the monolayers. The invasion ratio indicates the
percentage of filled space in a 48 h incubation.
Patient samples and
immunohistochemical analysis
Primary oropharyngeal squamous cell cancer (OPSCC)
tissue specimens were resected from 10 patients (mean age: 68.6,
age range: 53–86, sex distribution: Number of males is 8 and
females is 2) at Kobe University Hospital, fixed, and embedded in
paraffin for sectioning. All patients met the following inclusion
criteria: i) histologically diagnosed stage 1 OPSCC (15) and ii) complete resection, indicating a
homogeneous study population. Patients with a history of cancer or
any form of anticancer treatment were excluded from this study (see
the clinical profile of the patients in Table SI). Five cases were positive for p16
staining and five were negative. The resultant tissue sections were
incubated with antibodies against Ror2 (AF2064, 1:50; R&D
Systems) overnight at 4°C and then with anti-goat IgG antibodies
conjugated with HRP-labeled polymer (ImmPRESS Reagent kit
Peroxidase; Vector Laboratories) for 30 min at room temperature.
Secondary antibodies were visualized with DAB Chromogen (Dako
Cytomation) and nuclei were counterstained with hematoxylin.
Clinical tissue specimens were obtained and analyzed in accordance
with procedures approved by the Institutional Review Board of Kobe
University Hospital (no. B200096).
Extraction of DNA
Paraffin blocks were processed under strict
conditions to avoid contamination. Formalin-fixed,
paraffin-embedded (FFPE) tissues were cut into 5-µm thick sections
on glass slides, and DNA was extracted using the QIAamp DNA FFPE
tissue kit (Qiagen GmbH) according to the manufacturer's protocol.
The quantity and quality of extracted DNA was determined via
NanoDrop ND-1000 spectrophotometry (Thermo Fisher Scientific Inc.).
Isolated DNA was stored at −20°C until further use.
HPV DNA detection
As an internal control of the reaction, samples with
DNA were subjected to polymerase chain reaction (PCR) for human
β-globin gene amplification using PCO3
(5′CTTCTGACACAACTGTGTTCACTAGC3′) and PCO4
(5′TCACCACAACTTCATCCACGTTCACC3′) oligonucleotides, which flank a
sequence of approximately 110-bp (16). HPV DNA detection was carried out using
general primers GP5+/6+ (16). PCR
was performed with a final reaction volume of 50 µl, containing 5
µl of isolated DNA, 25 µl 2X buffer, 10 µl each dNTP, 0.3 µl each
of forward and reverse primers, and 1.0 unit DNA polymerase (Kod Fx
Neo; Toyobo Co.). The PCR conditions were as follows: Preheating
for 10 min at 94°C followed by 40 cycles of 1 min at 94°C, 2 min at
40°C, and 1.5 min at 72°C, and a final extension of 7 min at 72°C.
The amplified products were subjected to electrophoresis in 3%
agarose gels and observed using nucleic acid dye.
Western blotting
Cells were solubilized in ice-cold lysis buffer [50
mM HEPES (pH 7.5), 150 mM NaCl, 1% (v/v) Nonidet P-40 (NP-40), 1 mM
EDTA, 10 µg/ml aprotinin, 10 µg/ml leupeptin, and 1 mM p-APMSF] .
Proteins (10 µg in total per lane) was separated by SDS-PAGE (10 to
16% polyacrylamide gels were used) and transferred onto an
Immobilon-P membrane (Merck Millipore). Membranes were
immunoblotted with the following antibodies: Anti-Ror2, anti-E6
(GTX132686, 1:500, GeneTex), anti-E7 (sc-65711, 1:200, Santa Cruz
Biotechnology, Inc.), anti-α-tubulin (PM054, MBL). Immunoreactive
bands were visualized using Western Lightning Plus-ECL (Perkin
Elmer). The relative intensities of immunoblotted bands were
determined with ImageJ software.
Real-time quantitative RT-PCR
Total RNA was extracted from cultured cells using
Sepasol-RNA I SuperG (Nacalai Tesque). cDNAs were synthesized from
these RNAs as templates using PrimeScript RT Reagent (Takara Bio).
Expression levels of the respective genes of interest were measured
using the LightCycler 480 II system (Roche). The qPCR thermocycling
conditions were as follows: 95°C for 5 min followed by 60 cycles at
95°C for 10 sec, 60°C for 15 sec, 72°C for 15 sec, and final
extension step at 72°C for 5 min. The amounts of mRNAs were
normalized relative to those of 18S ribosomal RNA. Analysis
of relative gene expression was performed using the
2−∆∆Cq method (17). The
sequences of the primer pairs were as follows: Ror2
(5′-ATGTGGACTCCCTCCAGATG-3′ and 5′-GAAGACGAAGTGGCAGAAGG-3′);
E6/E7 (5′-CAATGTTTCAGGACCCACAGG-3′ and
5′-CTCACGTCGCAGTAACTGTTG-3′).
Statistical analysis
Data were analyzed using Bell Curve for Excel
(Social Survey Research Information Co., Ltd.) and are presented as
the mean ± standard deviation. Significance was determined as
*P<0.05, **P<0.01, or ***P<0.001 compared with the control
and indicated with the relevant symbols in the figures. Data were
analyzed using a Student's t-test when two groups were compared or
a one-way analysis of variance (ANOVA) with Tukey's honest
significance difference test when more than three groups were
analyzed.
Results
E6 and/or E7 oncoproteins enhance Ror2
expression in HNSCC
Ror2 is highly expressed in HPV-positive HNSCC cell
lines (6). However, the mechanism
underlying the enhanced expression of Ror2 in HPV-positive HNSCC
remains unclear. We examined the expression levels of Ror2 in
clinical specimens from patients with OPSCC using
immunohistochemical analysis. First, we confirmed the p16
expression status and examined HPV DNA expression via PCR using
G5+/6+ consensus primers for the L1 region of the viral genome.
Among the five p16-positive cases, we detected HPV DNA in four of
them, and, as expected, HPV DNA was not detected in the
p16-negative cases (Fig. 1A and B).
Among the 10 cases examined, all cases contained OPSCC cells that
were positive for Ror2 (Fig. 1C and
D). Ror2 expression was much stronger in cancer tissues than in
normal adjacent tissues (Fig. S1).
Expression of Ror2 was localized in the cytoplasm of tumor cells.
Interestingly, in one HPV-positive patient (Patient 2), metastatic
lesions in the neck lymph node showed higher expression of Ror2
than in the primary lesion, suggesting that expression levels of
Ror2 might be correlated with the invasive properties of cancer
cells (Fig. 1C). We further confirmed
that UPCI:SCC152 cells (HPV-positive HNSCC cell line) expressed
higher levels of Ror2 than UM-SCC-22B and Ca9-22 cells
(HPV-negative HNSCC cell lines) as assessed by qPCR (Fig. 1E) and western blot analyses (Fig. 1F), as previously reported (6). These results suggest that HPV enhances
the expression levels of Ror2 in HNSCCs and that Ror2 might be
involved in the progression of HNSCC.
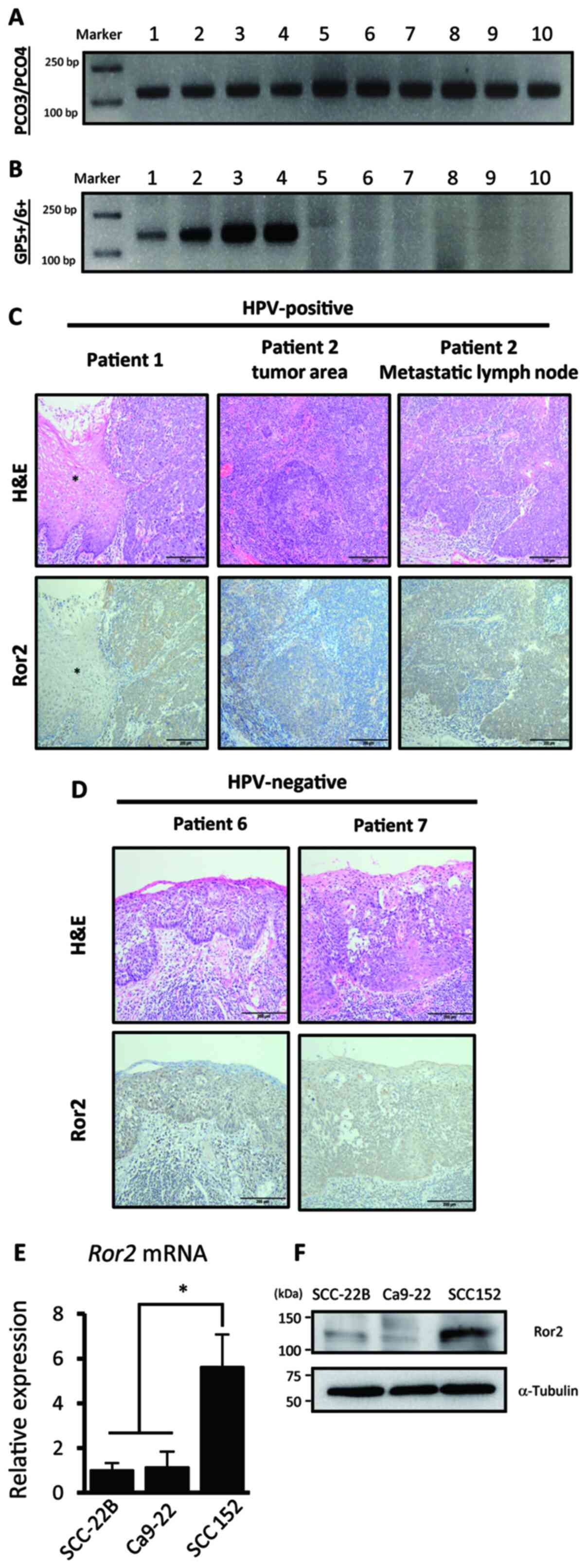 | Figure 1.Ror2 is highly expressed in HNSCC. (A
and B) Representative electrophoresis results of the amplicons from
tissues of patients with OPSCC for detecting the β-globin gene
(PCO3/PCO4 primer) and human papilloma virus L1 DNA (Gp5+/6+
primer). The images show the polymerase chain reaction (PCR)
products of 10 samples. (C and D) Immunohistochemical analyses of
Ror2 with surgical specimens from patients with HPV-positive (C)
and negative (D) OPSCC. Representative images of hematoxylin and
eosin (H&E) staining and anti-Ror2 immunohistochemistry of
HNSCC tissue sections are shown. The asterisks indicate the
non-tumor area. Scale bars,e 200 µm. (E) Expression levels of Ror2
in HNSCC cell lines UM-SCC-22B (HPV-negative), Ca9-22
(HPV-negative), and UPCI:SCC152 (HPV-positive); cells were measured
by qRT-PCR analysis. Relative expression levels were determined by
defining the expression level of Ror2 in UM-SCC-22B as 1. Data are
expressed as the mean ± standard deviation (n=3; *P<0.05,
Tukey's honest significance difference test). (F) Expression levels
of Ror2 and α-tubulin proteins in HNSCC cell lines
UM-SCC-22B, Ca9-22 and UPCI:SCC152; cells were evaluated by western
blot analysis. Ror2, receptor tyrosine kinase like orphan receptor
2; HNSCC, head and neck squamous cell cancer; HPV, human papilloma
virus; OPSCC, oropharyngeal squamous cell cancer; qRT-PCR,
quantitative reverse transcription-polymerase chain reaction. |
E6 and E7 oncoproteins encoded by the HPV genome
within infected HNSCC have been shown to induce the expression of
various genes in the host genome, thereby promoting tumor
progression (18). Thus, we next
examined whether E6 and E7 are involved in the enhanced expression
of Ror2 in UPCI:SCC152 cells using siRNAs against the HPV16
E6 and E7 transcripts (si-E6/E7). The
expression of Ror2 was significantly inhibited by suppressing the
expression of E6 and E7 oncoproteins with
si-E6/E7 (Fig. 2A-C). These
results indicate that E6 and/or E7 oncoproteins enhance expression
levels of Ror2 in HNSCC. However, the role of Ror2 upregulation in
HNSCC remains unclear.
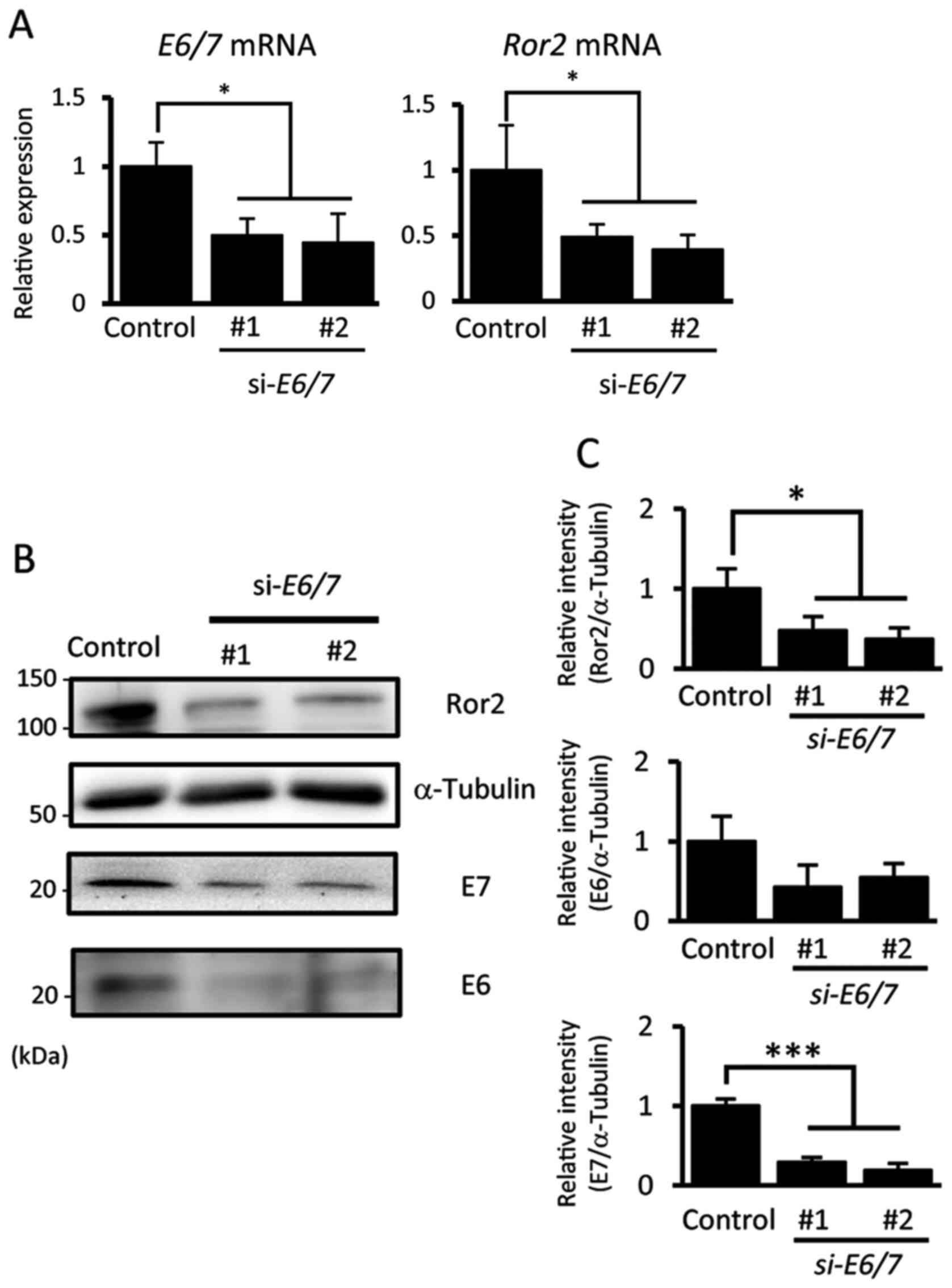 | Figure 2.Expression of Ror2 is regulated by
oncogenic E6 and/or E7 proteins in HPV16-positive HNSCCs. (A)
Expression levels of E6/7 and Ror2 mRNAs in
UPCI:SCC152 cells were measured by qRT-PCR analysis. Relative
expression values were determined by defining the expression level
in the control as 1. Data are expressed as the mean ± standard
deviation (n=3; *P<0.05, Tukey's honest significance difference
test). (B) Expression levels of Ror2, E6, E7 and α-tubulin proteins
in HPV-positive UPCI:SCC152 cells transfected with siRNA Control,
si-Ror2#1 or si-Ror2#2, were evaluated by western
blotting. (C) Relative band intensities of Ror2, E6 and E7 proteins
normalized by α-tubulin were determined. Data are expressed as mean
± standard deviation (n=3; *P<0.05, ***P<0.001 Tukey's honest
significance difference test). Ror2, receptor tyrosine kinase like
orphan receptor 2; HNSCC, head and neck squamous cell cancers; HPV,
human papilloma virus; qRT-PCR, quantitative reverse transcription
polymerase chain reaction. |
Ror2 plays an important role in
regulating proliferation of HPV-positive HNSCC cells by promoting S
phase entry
E6 and E7 oncoproteins promote the progression of
HPV-positive HNSCC cells by regulating their proliferative
activities (19). Indeed, we
confirmed that E6 and E7 knockdown resulted in
suppressed proliferative ability of UPCI:SCC152 cells (Fig. S2A). We performed an WST-8 assay to
examine whether Ror2 induced by E6 and/or E7 oncoproteins can
regulate the proliferative activities of UPCI:SCC152 cells. We
found that suppressed expression of Ror2 resulted in significantly
decreased proliferation of UPCI:SCC152 cells (Fig. 3A and B). To confirm whether the
inhibitory effect of Ror2 knockdown on the proliferation of
UPCI:SCC152 cells was partly due to induction of apoptosis, the
FITC-Annexin V apoptosis assay was performed. Both early and late
apoptotic rates of cells were unaffected by suppressed expression
of Ror2 (Fig. 3C and D). These
findings indicate that Ror2 plays an important role in regulating
the proliferation of UPCI:SCC152 cells.
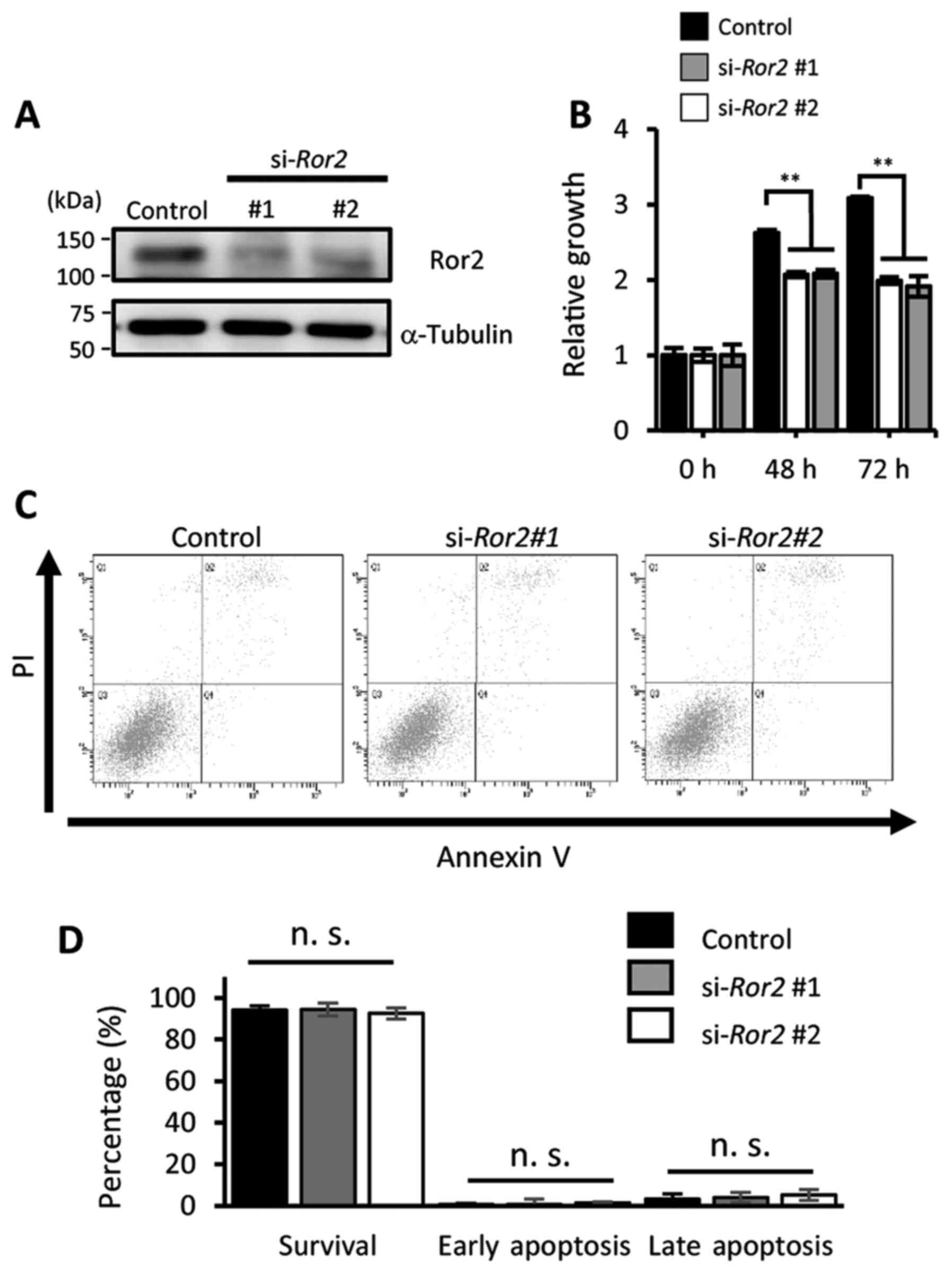 | Figure 3.Ror2 plays an important role in
regulating the proliferation of HPV-positive HNSCCs. (A) Expression
levels of Ror2 and α-tubulin proteins in UPCI:SCC152 cells treated
with the indicated siRNA control, si-Ror2#1, or
si-Ror2#2 were evaluated by western blot analysis. (B)
Viable cell numbers of HPV-positive UPCI:SCC152 cells transfected
with the indicated siRNA were measured using the WST-8 assay at the
indicated time points. Data are expressed as the mean ± standard
deviation (n=3; **P<0.01, Tukey's honest significance difference
test). (C) Proportion of apoptotic UPCI:SCC152 cells, transfected
with the indicated siRNAs, were examined by flow cytometric
analysis with PI and anti-Annexin V staining. (D) Quantification of
the proportion of apoptotic UPCI:SCC152 cells transfected with the
indicated siRNA. Data are expressed as the mean ± standard
deviation (n=3; n.s., not significant, Tukey's honest significance
difference test). Ror2, receptor tyrosine kinase like orphan
receptor 2; HNSCC, head and neck squamous cell cancer; HPV, human
papilloma virus. |
We previously showed that Ror2 is required for the
G1/S phase transition of NIH/3T3 fibroblasts (11). We thus investigated whether Ror2 is
involved in the G1/S transition of UPCI:SCC152 cells.
For this purpose, the effect of Ror2-knockdown on cell cycle
profiles of UPCI:SCC152 cells was monitored by flow cytometric
analysis with PI under non-synchronized conditions and we
re-started the cell cycle after synchronization. Since a double
thymidine block exhibited strong cell toxicity (data not shown), we
employed a single thymidine blockade to achieve overall
synchronization at the G1 phase. Any apparent effects of
Ror2-knockdown on the cell cycle profile were observed under
non-synchronized conditions (conventional cell culture conditions),
unlike the previous report (8,9) (data not
shown). UPCI:SCC152 cells transfected with either si-Ror2 or
control siRNA were subjected to one thymidine treatment to
synchronize the cell cycle at the G1 phase.
Subsequently, cell cycle progression of the resultant cells was
monitored every 3 h after the release of thymidine blockade.
Following synchronization, a vast majority of the cells were
arrested at the G1 phase. In total, 54.88% of control
cells entered the S phase within 3 h after release, while 40.98% of
si-Ror2#1 treated cells and 42.46% si-Ror2#2 treated
cells entered the S phase (Fig. 4A and
B). This significant deceleration continued for
si-Ror2#1 at 6 h after release, but lessened for
si-Ror2#2 (P=0.011 and P=0.55, respectively). These findings
indicate that Ror2 plays an important role in regulating the
proliferation of HPV-positive HNSCC cells by promoting the
G1/S phase transition.
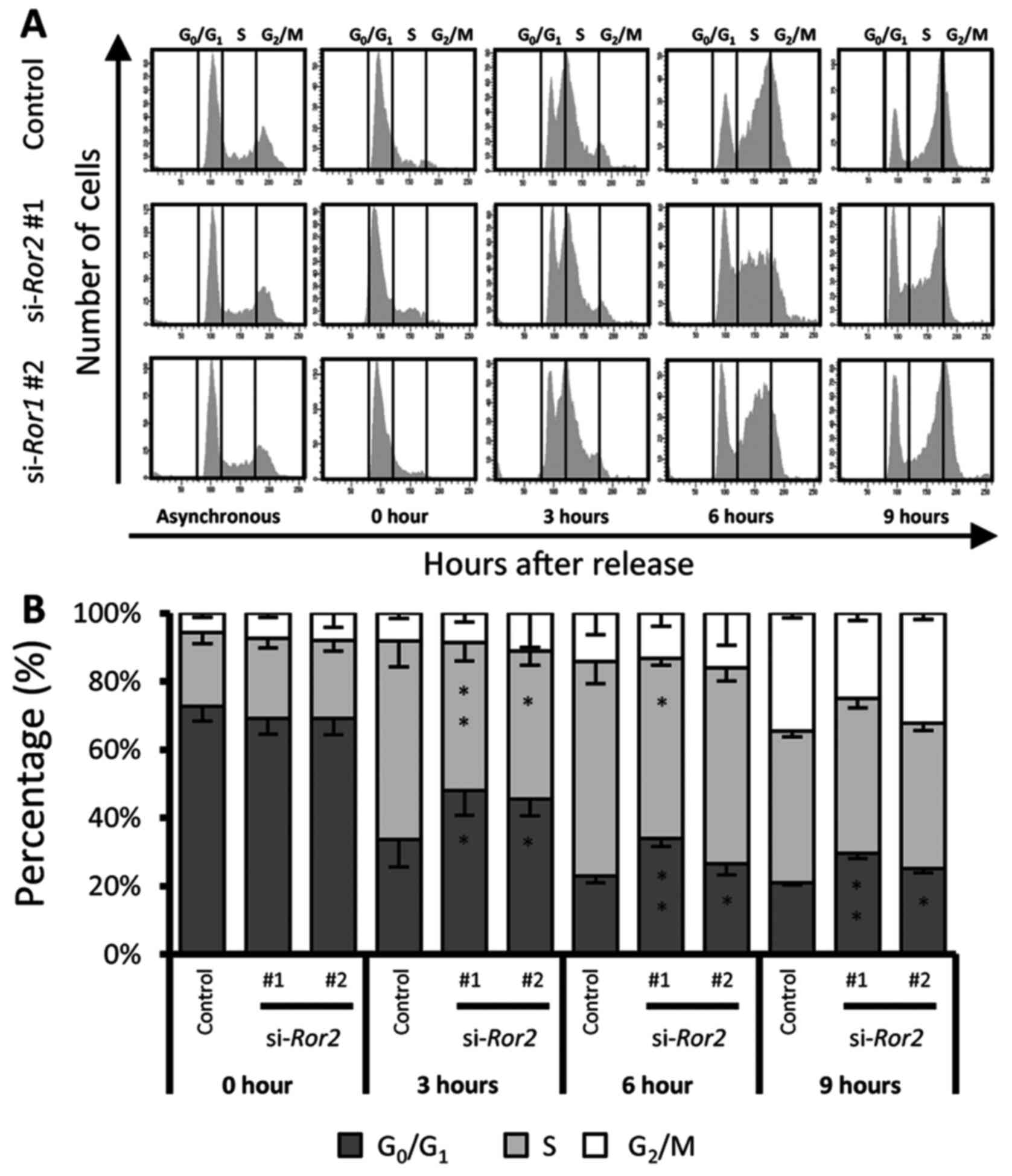 | Figure 4.The S phase entry of HPV-positive
HNSCCs is regulated by Ror2. (A) HPV-positive UPCI:SCC152 cells,
transfected with siRNA control, si-Ror2#1, or
si-Ror2#2, were cultured asynchronously or were synchronized
by treatment with thymidine. Twenty-four hours after treatment with
thymidine, the thymidine in the medium was removed and the DNA
contents of UPCI:SCC152 cells at the indicated time points were
determined by flow cytometric analysis. The DNA content histograms
reveal the proportions of cells that reside in the different cell
cycle phases at the respective time points. (B) Quantification of
the percentages of cells in each cell cycle phase at the indicated
time points. Data are expressed as the mean ± standard deviation
(n=3; *P<0.05, **P<0.01, Tukey's honest significance
difference test). Ror2, receptor tyrosine kinase like orphan
receptor 2; HNSCC, head and neck squamous cell cancer; HPV, human
papilloma virus. |
Ror2 is required to promote the
invasive ability of HPV-positive HNSCC cells
HPV has been shown to promote the invasive ability
of HNSCC cells (16). In fact, it was
confirmed that knockdown of E6 and E7 resulted in
significant inhibition of invasive ability of UPCI:SCC152 cells
(Fig. S2B). Thus, we examined
whether Ror2, induced by E6 and/or E7 oncoproteins, is also
involved in the invasive ability of HPV-positive HNSCC cells. To
this end, we employed Semi-3D and Matrigel invasion assays to
evaluate the invasive abilities of UPCI:SCC152 cells transfected
with control siRNA, si-Ror2#1, or si-Ror2#2.
Ror2-knockdown significantly inhibited the invasive ability
of UPCI:SCC152 cells (Fig. 5A-C),
indicating that Ror2 is required to promote the invasive ability of
HPV-positive HNSCC cells. Moreover, in agreement with the
undetectable expression of Ror2 in UM-SCC22B and Ca9-22 cells,
siRNAs against Ror2 failed to affect significantly their
invasiveness (Fig. S3).
Interestingly, Ror2-knockdown resulted in suppressed
expression of Slug, an epithelial-to-mesenchymal transition
(EMT)-related transcription factor, in UPCI:SCC152 cells (Fig. 5D), indicating that Ror2 might mediate
the invasive ability of HPV-positive HNSCC cells through
epithelial-mesenchymal transition (EMT).
Discussion
Recently, the expression of several possible target
cancer cell surface proteins for human papilloma virus
(HPV)-negative and HPV-positive head and neck squamous cell cancer
(HNSCC) has been established. Ror2 has been found to be more highly
expressed in HPV-positive HNSCC cell lines than in HPV-negative
cell lines (6). Thus, Ror2 (receptor
tyrosine kinase like orphan receptor 2) signaling pathways have
become a focus in the cancer community because of their critical
roles in cancer progression. In the present study, we used
molecular biology and cell biology methods to determine the role of
Ror2 in the regulation of HPV-positive HNSCC.
It has been suggested that Ror2 is important for the
regulation of cancer cell proliferation. However, the precise role
of Ror2 in the regulation of cancer cell proliferation, as well as
its underlying mechanism, remains unclear. Huang et al
demonstrated that knockdown of Ror2 led to cell cycle arrest
in osteosarcoma cells (8). Similarly,
Jiang et al found that miR-208b inhibited the proliferation
of osteosarcoma cell lines by targeting Ror2 (20). Conversely, it has been shown that the
expression levels of Ror2 were reduced in gastric carcinoma tissues
compared with that noted in matched normal adjacent tissues. In
addition, overexpression of Ror2 inhibited gastric cancer cell
proliferation, while inducing cell apoptosis and cell cycle arrest
at the G1 phase (9). In
the present study, Ror2 knockdown decreased the
proliferation of UPCI:SCC152 cells without inducing apoptosis.
Furthermore, we also confirmed that Ror2 plays an important role in
G1/S phase transition. The findings of this study are
consistent with our previous study, which showed that Ror2 promotes
G1/S phase transition in bFGF-stimulated NIH/3T3
fibroblasts by regulating E2F-target genes (11).
Interestingly, HPV-related HNSCC has a higher risk
for lympho-vascular invasion and lymph node metastasis (21), in spite of a better prognosis compared
with HPV-negative HNSCC (21). This
indicates an essential role of HPV in carcinogenic processes,
although its role in tumor metastasis to the lymph nodes remains
unclear. Considering that lymph node metastasis is a major risk
factor related to poor outcomes in patients with HNSCC,
identification of the underlying mechanism associated with HPV
infection may reveal potential strategies for managing lymph node
metastasis in these patients. Here, we found that the invasive
ability of UPCI:SCC152 cells was downregulated by Ror2
knockdown. Although some reports showed that Ror2 promotes invasion
by acting as a receptor for Wnt5a (22), others have indicated that Ror2
regulates invasion of cancer cells independently of Wnt5a (23,24). It is
presently unclear whether Ror2 can promote invasion of UPCI:SCC152
cells by acting as a receptor for Wnt5a. Therefore, it is important
to understand the molecular mechanism of Ror2-mediated invasion of
HPV-positive HNSCC in future studies.
Our results uncovered a novel relationship between
E6/7 oncoproteins and Ror2, which may be relevant for cancer
progression. We found that HPV16-E6/7 oncogenes can upregulate the
expression of Ror2 in UPCI:SCC152 cells, suggesting a possible
interaction between E6/7 and Ror2 during carcinogenesis. It is well
known that the HPV oncogenic potential is associated with the E6
and E7 oncoproteins which mediate the degradation of p53 and Rb
tumor suppressor proteins, respectively. We previously showed that
E2F1 induces Ror2 expression in bFGF-treated NIH/3T3 fibroblasts by
binding to the promoter region of the Ror2 gene. Therefore,
the E7 oncoprotein might be sufficient to induce Ror2 expression
through E2F1 by degradation of the Rb protein. Moreover, it has
recently been shown that E6 and E7 promote cancer progression
through p53- and Rb-independent pathways (25–27).
Therefore, the expression of Ror2 might not depend on the
degradation of p53 and Rb. Future studies investigating how E6/7
regulate the expression level of Ror2 are required.
We found that Ror2 is expressed in both HPV-negative
and HPV-positive oropharyngeal squamous cell cancer (OPSCC)
patients. In contrast, we and others showed that expression levels
of Ror2 are significantly higher in HPV-positive cancer cell lines
(UPCI:SCC152) than in HPV-negative cancer cell lines (UM-SCC22B,
Ca9-22). It can be assumed that this discrepancy is at least partly
due to the difference between in vitro and in vivo
microenvironmental factors. HPV-negative cancer cell lines might
lose some carcinogenic features such as Ror2 expression with the
loss of tumor stroma, while HPV-positive cell lines do not. This
might be explained by the cell-autonomous interaction between E6/7
and Ror2 expression in the HPV-positive cell line. Although the
present study indicates that E6/7 induces the expression of Ror2 in
UPCI:SCC152 cells, the molecular mechanism of Ror2 upregulation in
HPV-negative HNSCC remains unclear. The tumor microenvironment
plays a critical role in regulating cancer progression. We
previously showed that CXCL16 derived from mesenchymal stem cells,
one of the components of the tumor microenvironment, induces the
expression of Ror1 in gastric cancer cells (28). Therefore, this might indicate the
possibility that the tumor microenvironment regulates Ror2
expression in HPV-negative HNSCC through an E6/7 independent
pathway. Future studies investigating the relationship between the
tumor microenvironment and Ror2 upregulation in HPV-negative HNSCC
are required.
Recently, treatment for HPV-positive cases has been
changed to biopsy followed by chemoradiotherapy. Thus, the
incidence of excisional HPV-positive cancer tissues is declining.
The main limitation of the present study was that the number of
OPSCC tissues was not sufficient to compare the association between
the expression levels of Ror2 in HPV-negative and HPV-positive
samples. Furthermore, we were not able to examine the clinical
characteristics of patients, such as the tumor grade, TNM stage,
and metastasis. Consequently, future studies with more HPV-negative
and HPV-positive cancer cell lines and larger sample sizes are
required to compare Ror2 expression in HPV-negative and
HPV-positive cases.
Although it has been four decades since the
association between HPV and human cancer was first proposed
(29), the epidemic of HPV-positive
disease still requires the identification of novel biomarkers. Our
findings indicate that: i) E6/7 induce the expression of Ror2 to
promote proliferation and invasion, suggesting a novel mechanism of
Ror2 function in HPV-related carcinogenesis and ii) E6/7 have the
potential to be novel targets in HPV-related cancers. Since
HPV-induced cancers depend on the expression of E6/7, targeting
E6/7 may alter Ror2 signaling pathways. Advances in understanding
the role and characteristics of Ror2 in HPV-related cancers and
understanding how to manipulate Ror2 signaling by HPV oncogenes may
serve as novel approaches to treat HNSCC and other HPV-related
cancers.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Tokyo Biochemical
Research Foundation.
Availability of data and materials
Original data and material will be made available
upon reasonable request.
Author's contributions
MOA, KK and YM designed the study. MOA, KK and NJ
performed the experiments and analyzed the data. MOA, KK and YM
interpreted the data. NJ, HS, KIN and MN were involved in the
interpretation and evaluation of the data and findings. MOA, KK and
YM prepared the manuscript. NJ, HS, KIN and MN reviewed and edited
manuscript. All authors approved the final manuscript.
Ethics approval and consent to
participate
This study is approved by the institutional review
board of Kobe University Hospital (no. B200096). The Opt-out method
has been used.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HPV
|
human papilloma virus
|
|
HNSCC
|
head and neck squamous cell cancer
|
|
OPSCC
|
oropharyngeal squamous cell cancer
|
References
|
1
|
Leemans CR, Snijders PJF and Brakenhoff
RH: The molecular landscape of head and neck cancer. Nat Rev
Cancer. 18:269–282. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Martel C, Plummer M, Vignat J and
Franceschi S: Worldwide burden of cancer attributable to HPV by
site, country and HPV type. Int J Cancer. 141:664–670. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van Doorslaer K, Chen Z, Bernard HU, Chan
PKS, DeSalle R, Dillner J, Forslund O, Haga T, McBride AA, Villa
LL, et al: ICTV virus taxonomy profile: Papillomaviridae. J Gen
Virol. 99:989–990. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kumar A, Rathi E, Hariharapura RC and Kini
SG: Is viral E6 oncoprotein a viable target? A critical analysis in
the context of cervical cancer. Med Res Rev. 40:2019–2048. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marur S, D'Souza G, Westra WH and
Forastiere AA: HPV-associated head and neck cancer: A virus-related
cancer epidemic. Lancet Oncol. 11:781–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carrero I, Liu HC, Sikora AG and
Milosavljevic A: Histoepigenetic analysis of HPV- and
tobacco-associated head and neck cancer identifies both
subtype-specific and common therapeutic targets despite divergent
microenvironments. Oncogene. 38:3551–3568. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kamizaki K, Endo M, Minami Y and Kobayashi
Y: Role of noncanonical Wnt ligands and Ror-family receptor
tyrosine kinases in the development, regeneration, and diseases of
the musculoskeletal system. Dev Dyn. 250:27–38. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang J, Shi Y, Li H, Tan D, Yang M and Wu
X: Knockdown of receptor tyrosine kinase-like orphan receptor 2
inhibits cell proliferation and colony formation in osteosarcoma
cells by inducing arrest in cell cycle progression. Oncol Lett.
10:3705–3711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yan L, Du Q, Yao J and Liu R: ROR2
inhibits the proliferation of gastric carcinoma cells via
activation of non-canonical Wnt signaling. Exp Ther Med.
12:4128–4134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saji T, Nishita M, Ogawa H, Doi T, Sakai
Y, Maniwa Y and Minami Y: Critical role of the Ror-family of
receptor tyrosine kinases in invasion and proliferation of
malignant pleural mesothelioma cells. Genes Cells. 23:606–613.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Endo M, Tanaka Y, Otsuka M and Minami Y:
E2F1-Ror2 signaling mediates coordinated transcriptional regulation
to promote G1/S phase transition in bFGF-stimulated NIH/3T3
fibroblasts. FASEB J. 34:3413–3428. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi M, Shibuya Y, Takeuchi J, Murata
M, Suzuki H, Yokoo S, Umeda M, Minami Y and Komori T: Ror2
expression in squamous cell carcinoma and epithelial dysplasia of
the oral cavity. Oral Surg Oral Med Oral Pathol Oral Radiol Endod.
107:398–406. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Enomoto M, Hayakawa S, Itsukushima S, Ren
DY, Matsuo M, Tamada K, Oneyama C, Okada M, Takumi T, Nishita M and
Minami Y: Autonomous regulation of osteosarcoma cell invasiveness
by Wnt5a/Ror2 signaling. Oncogene. 28:3197–3208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Aoki T, Nishita M, Sonoda J, Ikeda T,
Kakeji Y and Minami Y: Intraflagellar transport 20 promotes
collective cancer cell invasion by regulating polarized
organization of Golgi-associated microtubules. Cancer Sci.
110:1306–1316. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Amin MB, Greene FL, Edge SB, Compton CC,
Gershenwald JE, Brookland RK, Meyer L, Gress DM, Byrd DR and
Winchester DP: The Eighth edition AJCC Cancer Staging Manual:
Continuing to build a bridge from a population-based to a more
‘personalized’ approach to cancer staging. CA Cancer J Clin.
67:93–99. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tao X, Zheng B, Yin F, Zeng Z, Li Z,
Griffith CC, Luo B, Ding X, Zhou X and Zhao C: Polymerase Chain
reaction human papillomavirus (HPV) detection and HPV genotyping in
invasive cervical cancers with prior negative HC2 test results. Am
J Clin Pathol. 147:477–483. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hoppe-Seyler K, Bossler F, Braun JA,
Herrmann AL and Hoppe-Seyler F: The HPV E6/E7 oncogenes: Key
factors for viral carcinogenesis and therapeutic targets. Trends
Microbiol. 26:158–168. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee SH, Lee CR, Rigas NK, Kim RH, Kang MK,
Park NH and Shin KH: Human papillomavirus 16 (HPV16) enhances tumor
growth and cancer stemness of HPV-negative oral/oropharyngeal
squamous cell carcinoma cells via miR-181 regulation.
Papillomavirus Res. 1:116–125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang Z, Jiang C, Yu C and Fang J:
MicroRNA-208b inhibits human osteosarcoma progression by targeting
ROR2. Tumour Biol. 39:10104283177057512017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Benson E, Li R, Eisele D and Fakhry C: The
clinical impact of HPV tumor status upon head and neck squamous
cell carcinomas. Oral Oncol. 50:565–574. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
O'Connell MP, Fiori JL, Xu M, Carter AD,
Frank BP, Camilli TC, French AD, Dissanayake SK, Indig FE, Bernier
M, et al: The orphan tyrosine kinase receptor, ROR2, mediates Wnt5A
signaling in metastatic melanoma. Oncogene. 29:34–44. 2010.
View Article : Google Scholar
|
|
23
|
Xu J, Shi J, Tang W, Jiang P, Guo M, Zhang
B and Ma G: ROR2 promotes the epithelial-mesenchymal transition by
regulating MAPK/p38 signaling pathway in breast cancer. J Cell
Biochem. 121:4142–4153. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rasmussen NR, Debebe Z, Wright TM, Brooks
SA, Sendor AB, Brannon AR, Hakimi AA, Hsieh JJ, Choueiri TK,
Tamboli P, et al: Expression of Ror2 mediates invasive phenotypes
in renal cell carcinoma. PLoS One. 9:e1161012014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yeo-Teh NSL, Ito Y and Jha S: High-risk
human papillomaviral oncogenes E6 and E7 target key cellular
pathways to achieve oncogenesis. Int J Mol Sci. 19:17062018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Charette ST and McCance DJ: The E7 protein
from human papillomavirus type 16 enhances keratinocyte migration
in an Akt-dependent manner. Oncogene. 26:7386–7390. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carrillo D, Muñoz JP, Huerta H, Leal G,
Corvalán A, León O, Calaf GM, Urzúa U, Boccardo E, Tapia JC and
Aguayo F: Upregulation of PIR gene expression induced by human
papillomavirus E6 and E7 in epithelial oral and cervical cells.
Open Biol. 7:1701112017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ikeda T, Nishita M, Hoshi K, Honda T,
Kakeji Y and Minami Y: Mesenchymal stem cell-derived CXCL16
promotes progression of gastric cancer cells by STAT3-mediated
expression of Ror1. Cancer Sci. 111:1254–1265. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
zur Hausen H: Papillomaviruses and cancer:
From basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002. View
Article : Google Scholar : PubMed/NCBI
|