Introduction
Metastasis occurs in ~90% of malignant tumors and is
the leading cause of cancer-associated mortality in patients with
cancer worldwide (1,2). A number of biological factors and
multiple signaling pathways, such as epithelial-mesenchymal
transition, resistance to apoptosis and angiogenesis have been
associated with the complex processes of metastasis (3); however, a novel approach for effectively
controlling tumor metastasis is still required.
Metastasis is defined as cancer cells leaving the
original tumor mass and disseminating to other parts of the body
via the bloodstream or lymphatic system. Therefore, the metastatic
process represents a multi-step event (3). For example, remote hematogenous
metastasis requires the cancer cells to successfully pass through
the following steps: i) Transendothelial migration into the vessel
(known as intravasation); ii) survival in the circulatory system;
iii) attachment to the vessel wall and transendothelial migration
out of the vessel (known as extravasation) and iv) eventually live
and propagate at the distal site (4,5). All of
these steps are accompanied with a change in the surrounding
microenvironment, with various biomechanical forces and oxidative
stress (6); therefore, metastasis can
be a stressful and inefficient event to the cancer cell (7).
Biomechanical forces have been demonstrated to play
critical roles in regulating cell migration and proliferation
(8,9).
With the rapid advancement of mechanobiology in recent years, it
has become a hot topic for understanding how biomechanical forces
mediate malignant tumor progression (2,8). Beyond
the mechanical stress during metastatic processes, it is also
well-known that elevated interstitial fluid hydrostatic pressure
(HP) occurs in solid tumors (10,11).
Higher interstitial fluid HP in tumor mass has been demonstrated to
be associated with a worse prognosis in patients with head and neck
cancer (12). Furthermore, the
exposure of cancer cells to 20 mmHg HP has been demonstrated to
accelerate cell motility (8).
However, it is not clear whether and how the elevation of
interstitial fluid HP in tumor mass promotes the metastasis of
cancer cells.
Notably, it has recently been reported that cyclical
mechanical force can induce the stabilization of HIF-1α and
upregulate the protein expression level of CXCL2 in monocytes
(13). HIF-1α is well-known as a
master upstream regulator of oxidative stress, metabolism and DNA
repair of cells (14–16). Therefore, we hypothesized that
elevated interstitial fluid HP in a rapid growing malignant tumor
may stabilize HIF-1α to promote the metastasis of cancer cells.
In the present study, mouse Lewis lung carcinoma
(LLC) cells were exposed to 50 mmHg HP for 24 h, then the role of
HP on the metastatic property of these cells was investigated using
both in vitro and in vivo experiments.
Materials and methods
Cells and animals
The LLC cells (LL/2) were used for the experiments.
The cells were maintained in DMEM (FUJIFILM Wako Pure Chemical
Corporation), supplemented with 10% fetal bovine serum (Cytiva) and
1% penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.),
and cultured at 37°C in a humidified incubator with 5%
CO2.
A total of 19, male C57BL/6 mice (10–12 weeks old;
weight, 23–25 g; CLEA Japan, Inc.) were used for the in vivo
study. The mice were kept in specific, pathogen-free conditions and
were allowed free access to food and water under a controlled
temperature (24±1°C) with 55% humidity in a 12-h light/dark cycle.
The animal experiments were approved by the Institutional Animal
Care and Use Committee of Nagasaki University (approval no.
1608251335-11). All the animal procedures were performed in
accordance with institutional and national guidelines. At the end
of the experiments, the mice were administered with general
anesthesia using an intraperitoneal injection of mixed anesthetics
(0.75 mg/kg medetomidine, 4 mg/kg midazolam and 5 mg/kg
butorphanol) and sacrificed by severing the abdominal aorta for
blood removal. The removal of vital organs (lung tissue) was used
as confirmation of the death of the mice following sacrifice.
HP stimulation
HP was induced in the LLC cells using a
pneumatic pressurizing system (Strex. Inc.). Briefly, the LLC cells
were seeded in 60 mm diameter Petri dishes (1×105
cells/dish) and cultured for 36 h to form an adherent monolayer.
The culture dishes were then randomly selected to move into a
sealed chamber in which 50 mmHg HP was stably applied using the
pneumatic pressurizing system and kept for 24 h (HP group). The
culture dishes without HP exposure were used as the control (CON
group).
Cell morphology observation and cell
count
Cell morphology was observed under a light
microscope (1X71S8F-3; Olympus Corporation) at ×200 magnification,
24 h following HP exposure. Then, the cells were collected as a
single cell suspension to measure the total cell number using a
TC20™ Automated Cell Counter (Bio-Rad Laboratories, Inc.).
Reverse transcription (RT)2
Profiler™ PCR array
To investigate the mRNA expression level of genes
associated with metastasis, RNA was isolated from the cells using a
Quick-RNA™ MicroPrep kit (Zymo Research Corp.). The concentration
of RNA was measured using a NanoDrop® 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). Then, 1 µg RNA
was used to generate cDNA using the RT2 First Strand kit
(Qiagen Corporation), at 25°C for 10 min, 42°C for 60 min, then
85°V for 5 min. Mouse Tumor Metastasis RT2 Profiler™ PCR
array (cat. no. 330231; Qiagen Corporation) was used with a
RT2 SYBR-Green Master mix, according to the
manufacturer's instructions and a Roche LightCycler 480 machine
(Roche Diagnostics). The array contained a total of 84 genes
associated with metastasis. The genes included in the assay were
also defined by biological function by the manufacturer. The fold
change in expression to the control was calculated using a
web-based data analysis program (https://geneglobe.qiagen.com/jp/analyze). Among the 5
available housekeeping genes (Actb, B2m, Gapdh, Gusb and
Hsp90ab1) in the array, Actb, Gusb and
Hsp90ab1 were automatically selected as the optimal set of
internal control for normalization.
Adhesion assay
To evaluate the adhesion ability, the cells from
both the HP and CON groups were harvested as single cell
suspensions. Freshly harvested cells (5×104 cells in 5
ml DMEM) were seeded onto a 25-cm2 Collagen I-coated
Flask (Thermo Fisher Scientific, Inc.). Following incubation for 60
min, the unattached cells were gently removed by washing with PBS
twice. The number of adherent cells was counted under a light
microscope at ×200 magnification. The average cell count from
>20 randomly selected fields was used for statistical
analysis.
Western blot analysis
The protein expression level of HIF-1α, SOD1 and
SOD2 was evaluated using western blot analysis, as previously
described (17). Total protein from
the cells was extracted using 1X RIPA buffer (FUJIFILM Wako Pure
Chemical Corporation) and the concentration was detected using a
BCA assay. A total of 30 µg protein from each sample was separated
using 10–12% SDS-PAGE, then transferred to 0.2-µm PVDF membranes
(Bio-Rad Laboratories, Inc.). After blocking with 5% skimmed milk
for 1 h at room temperature, the membranes were incubated with
primary antibodies against HIF-1α (1:250 dilution; cat. no. ab1;
room temperature for 2 h; Abcam), SOD1 (1:500 dilution; cat. no.
sc11407; overnight at 4°C; Santa Cruz Biotechnology, Inc.), SOD2
(1:500 dilution; cat. no. sc30080; overnight at 4°C; Santa Cruz
Biotechnology, Inc.) and β-actin (1:1,000 dilution; cat. no. 8457S;
overnight at 4°C; Cell Signaling Technology, Inc.), followed by
incubation with horseradish peroxidase-conjugated secondary
antibodies (rabbit anti-mouse, 1:1,000 dilution; cat. no. P026002;
goat anti-rabbit, 1:1,000 dilution; cat. no. P044801) (both from
Dako; Agilent Technologies, Inc.) at room temperature for 1 h. The
expression level was visualized using an enhanced chemiluminescence
detection kit (Thermo Fisher Scientific, Inc.). Semi-quantitative
analysis was done using ImageQuant LAS 4000 mini detection system
(v1.0; GE Healthcare Life Sciences).
Evaluation of oxidative stress
tolerance
To evaluate oxidative stress tolerance, the cells
from both groups were treated with 0, 20 or 50 µM hydrogen peroxide
(H2O2) in PBS at 37°C for 2 h. The apoptotic
cells were stained with Annexin V-FITC, while the necrotic cells
were labelled with PI using an Annexin V-FITC Apoptosis Detection
Kit (Abcam). The cells without staining were used as negative
control. Quantitative flow cytometry analysis was performed using a
FACSVerse™ flow cytometer and analyzed using BD FACSuite Software
(v1.2 Suite 1.0.2) (both from BD Biosciences).
In addition, intracellular reactive oxygen species
(ROS) was detected in the cells. Briefly, the cells from both
groups were treated with 0, 20 or 50 µM H2O2
in PBS at 37°C for 1 h, then incubated with 10 µM general oxidative
stress indicator (CM-H2DCFDA; Invitrogen; Thermo Fisher Scientific,
Inc.) for another 30 min in the dark. Cells without staining were
used as a negative control. The accumulation of intracellular ROS
was measured by fluorescence intensity using a FACSVerse™ flow
cytometer (BD Biosciences) and analyzed using BD FACSuite Software
(v1.2 Suite 1.0.2) (both from BD Biosciences).
Experimental lung cancer metastasis
model
To evaluate the metastatic potency, experimental
lung cancer metastasis was induced in mice using an intravenous
injection of LLC cells (5×105 cells in 0.5 ml saline)
from the HP (n=6) and the CON (n=7) groups. A total of 4 weeks
after the cells were injected into the mice, all the mice were
sacrificed as aforementioned. Removal of lung tissue was used for
both confirmation of mice death and experimental evaluation.
Excised lung tissue was weighed and the number of tumor lesions on
the lung surface was counted. For the mice that spontaneously died
during the 4-week follow-up period, the date of death was recorded
and the lung tissue samples were collected for evaluation.
Statistical analysis of the overall survival rate of the mice was
also determined.
Immunohistochemical staining
The cell proliferation and microvessel density in
the metastatic lesions of the lungs was detected using
immunohistochemistry staining. The lungs were fixed in 4%
paraformaldehyde for 24 h, at 4°C, and paraffin-embedded sections
(6-µm thick) were deparaffinized and rehydrated (xylene, 2×3 min
washes; xylene 1:1 with 100% ethanol, 3 min; 100% ethanol, 2×3 min
washes; 95% ethanol, 3 min; 70% ethanol, 3 min; 50% ethanol, 3 min;
running cold tap water to rinse). After blocking with 1% BSA in PBS
(Sigma-Aldrich; Merck KGaA), the sections were incubated with
rabbit anti-mouse Ki67 antibody (cat. no. ab16667; 1:100 dilution;)
and rabbit anti-mouse CD31 antibody (cat. no. ab28394; 1:150
dilution) (both from Abcam) overnight at 4°C, followed by
incubation with the Alexa fluorescent 546-conjugated goat anti
rabbit IgG(H+L) secondary antibody (cat. no. A11013; 1:350
dilution; Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 1 h. Nuclei were stained with 4,
6-diamidino-2-phenylin-dole (DAPI; cat. no. D21490; Thermo Fisher
Scientific, Inc.) at room temperature for 5 min. Positive staining
was examined under a fluorescent microscope (FV10C-W3; Olympus
Corporation). The percentage of Ki67-positive cells was calculated
from 10 randomly selected fields of view (5 fields/slide in 2
slides) and used for statistical analysis. The CD31-positive
stained structures were counted as microvessels and the average
number of microvessels counted from 10 randomly selected fields of
view (5 fields/slide in 2 slides) was used for statistical
analysis.
PKH26 red fluorescent cell
labeling
To evaluate the survival/retention of the LLC cells
in the lungs of the mice, the cells were labelled with a PKH26 Red
Fluorescent Cell Linker kit (Sigma-Aldrich; Merck KGaA). Briefly,
the cells were incubated with 2 µM PKH26 dye for 5 min at room
temperature, as the manufacturer's recommendations. Then, the mice
were intravenously injected with the PKH26-labelled cells
(1×106 cells in 0.5 ml saline) from the HP (n=3) and the
CON (n=3) groups. The mice were sacrificed 24 h following the
injection and the lung tissue samples were collected. Cryosections
(8-µm thick) of the lung tissues were used for the direct detection
of PKH26-labelled LLC cells under a fluorescent microscope
(FV10C-W3; Olympus Corporation).
Statistical analysis
The data are presented as the mean ± SD. Statistical
significance between two groups was determined using an unpaired
t-test (SPSS; v20.0; IBM Corp.). The survival of the mice was
analyzed using a Kaplan-Meier curve and statistical significance
was determined using the log-rank test (GraphPad Prism; v8.0.1;
GraphPad Software, Inc.). P<0.05 was used to indicate a
statistically significant difference.
Results
Exposure of the LLC cells to 50 mmHg
HP altered the mRNA expression level of numerous genes associated
with metastasis
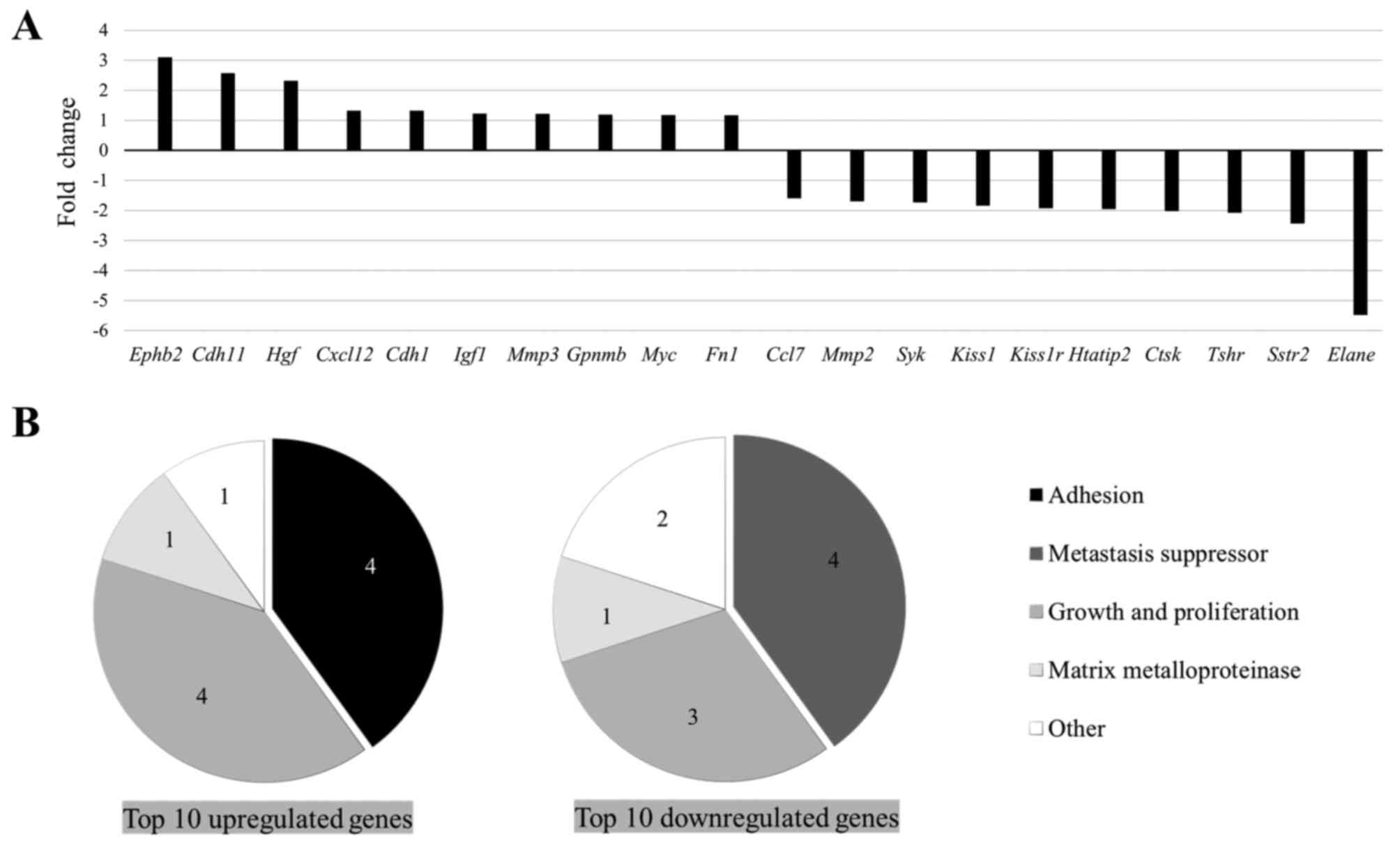
Firstly, the mRNA expression level of genes
associated with metastasis was analyzed between the HP and CON
groups. The RT2 Profiler™ PCR array revealed that
numerous genes were up- or downregulated, with a >1.3-fold
difference in the HP group compared with that in the CON group
(Table SI). The top 10 up- or
downregulated genes are shown in Fig.
1A. Within the top 10 upregulated genes, the upregulation of
several adhesion molecules was found, including Cdh1, Cdh11
and Fn1 (Fig. 1B). In
addition, the upregulation of numerous metastasis-promoting genes,
such as Hgf, Cdh11 and Ephb2, was also found. Within
the top 10 downregulated genes, metastasis suppressors, including
Kiss1, Syk and Htatip2, were frequently detected
(Fig. 1B). The overall change in the
gene expression profile indicated the potential role of HP in
enhancing metastatic properties of the LLC cells.
HP exposure enhances the adhesion
property of the LLC cells
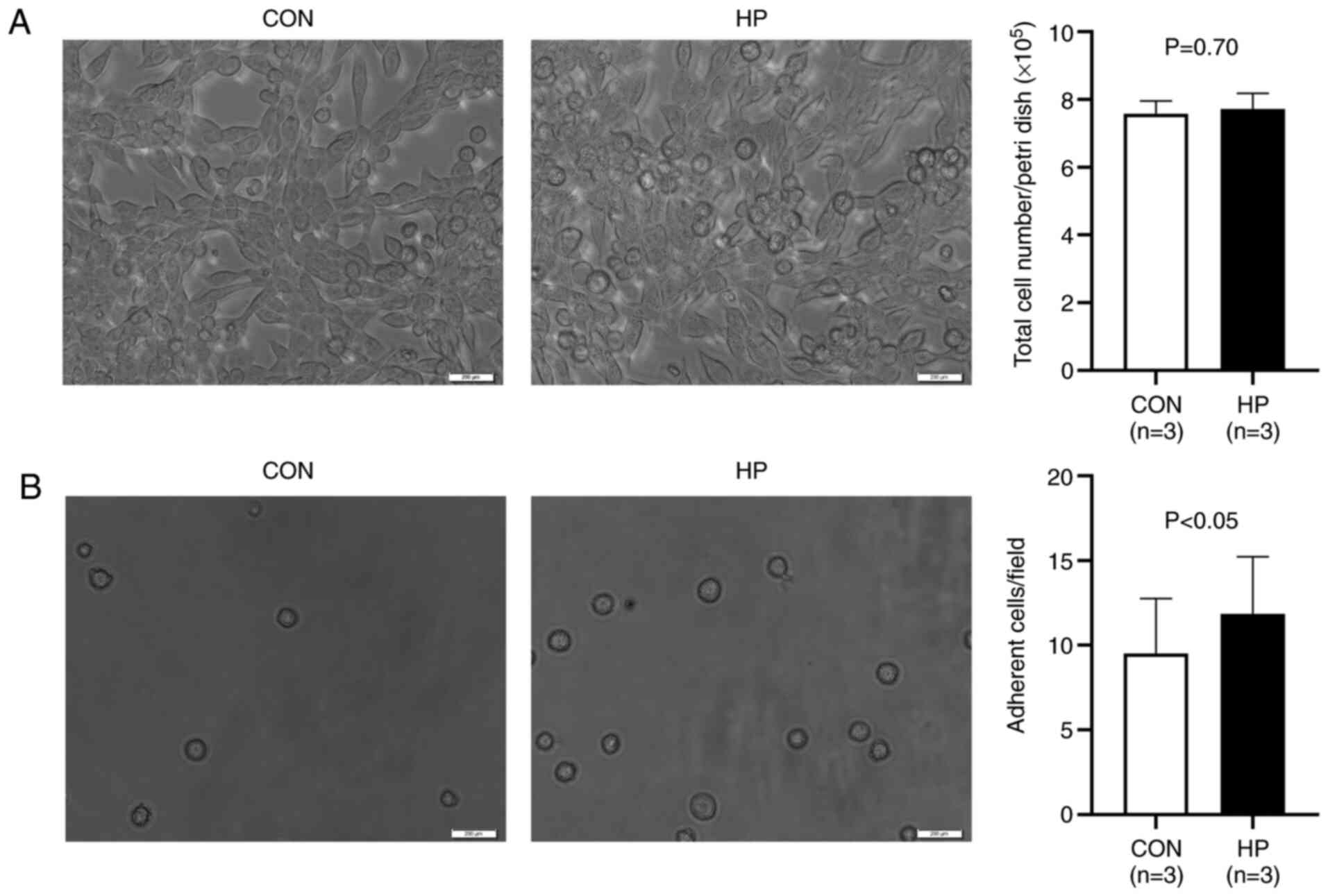
The exposure of the LLC cells to 50 mmHg HP for 24 h
did not induce notable morphological changes (Fig. 2A). The total number of harvested cells
was also comparable between the groups (P=0.70; Fig. 2A), indicating a limited effect of 50
mmHg HP exposure on cell growth.
As the PCR array data indicated the upregulation of
numerous adhesion molecules, the adhesion property of the cells was
investigated. It was found that the exposure to 50 mmHg HP
significantly increased the number of adherent cells on a collagen
I-coated flask (P<0.05; Fig.
2B).
HP exposure increases the protein
expression level of HIF-1α and antioxidant enzymes in the LLC
cells
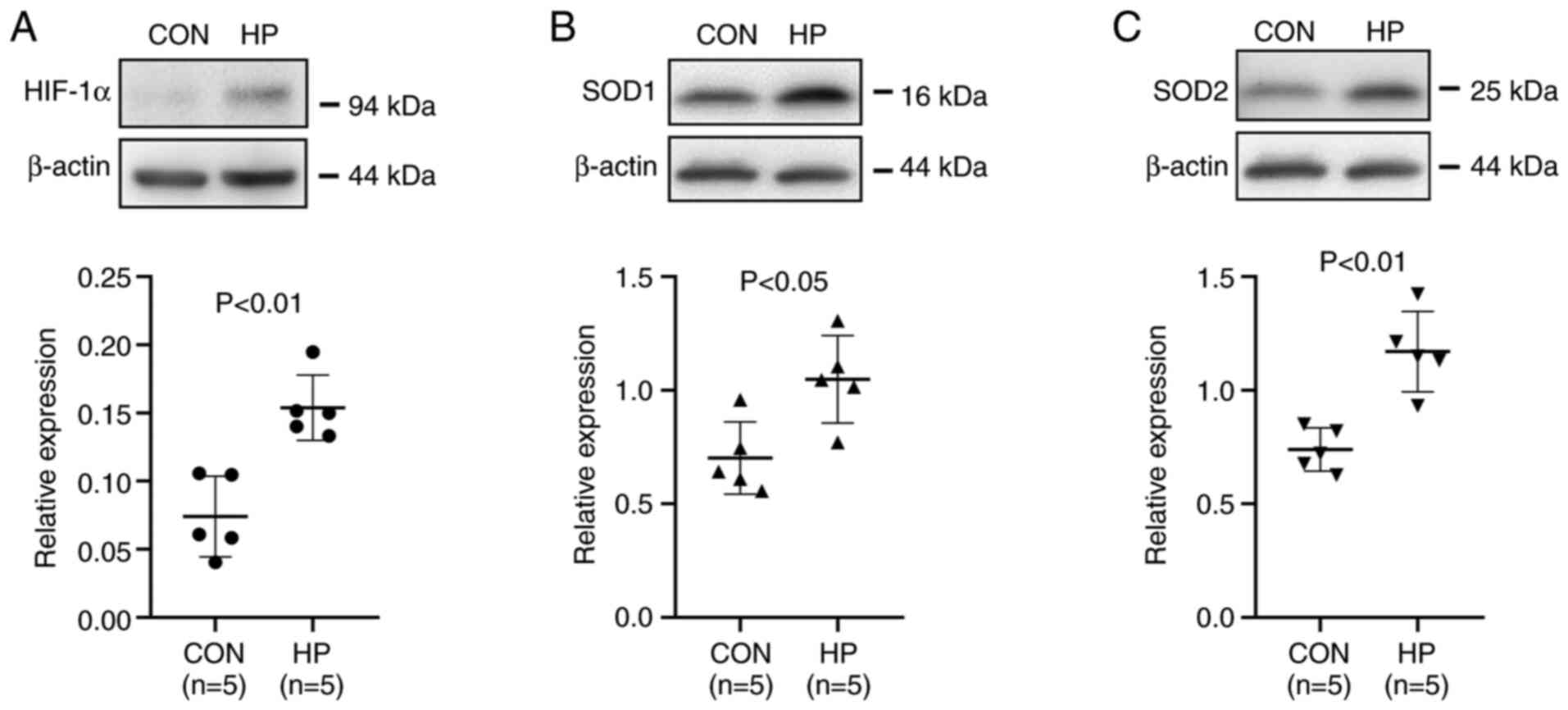
HIF-1α, a master regulator of the cellular adaptive
response to hypoxia, is known to play critical roles in metabolic
reprogramming (16) and metastasis
(14) in cancer cells. Western blot
analysis showed that the protein expression level of HIF-1α was
significantly increased in cells exposed to 50 mmHg HP for 24 h
(P<0.01; Fig. 3A).
The protein expression level of the antioxidant
enzymes, SOD1 and SOD2, which are HIF-1α downstream signals
(18,19), was also investigated. As expected, the
exposure of the cells to 50 mmHg HP for 24 h also significantly
upregulated the protein expression level of SOD1 and SOD2
(P<0.05; Fig. 3B and C).
HP exposure induces the tolerance of
the LLC cells to oxidative stress
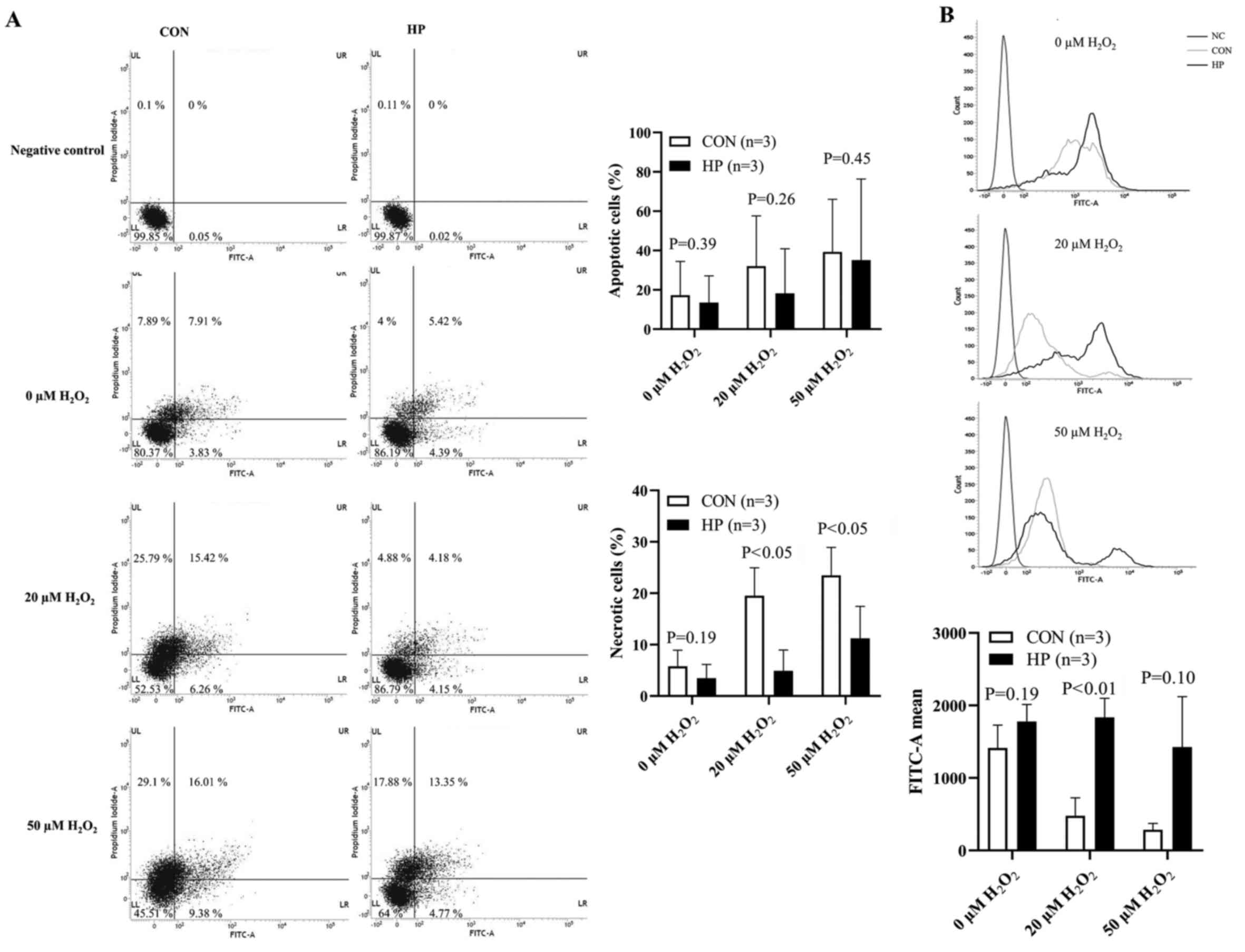
In addition, the oxidative stress tolerance of the
cells was investigated in vitro. Cell necrosis, under 20 or
50 µM H2O2 treatment was significantly
reduced in the LLC cells pretreated with 50 mmHg for 24 h
(P<0.05; Fig. 4A); however, the
percentage of apoptotic cells was not significantly different
between the 2 groups treated with 20 or 50 µM
H2O2 (P=0.26 and P=0.45, respectively;
Fig. 4A).
The intracellular ROS level at the baseline (without
H2O2 stimulation) was detected at comparable
levels between the HP and CON groups (Fig. 4B). Unexpectedly, 1-h stimulation with
20 or 50 µM H2O2 notably decreased the ROS
accumulation in the LLC cells without pretreatment with 50 mmHg HP
compared with that at baseline (Fig.
4B). By contrast, after 1-h stimulation with 20 µM
H2O2, the ROS accumulation was slightly
increased in the LLC cells pretreated with 50 mmHg HP compared with
that at baseline. We hypothesized that the less intracellular ROS
accumulation in the LLC cells without HP exposure was due to the
severe cell damage or cell death, rather than the resistance to
oxidative stress.
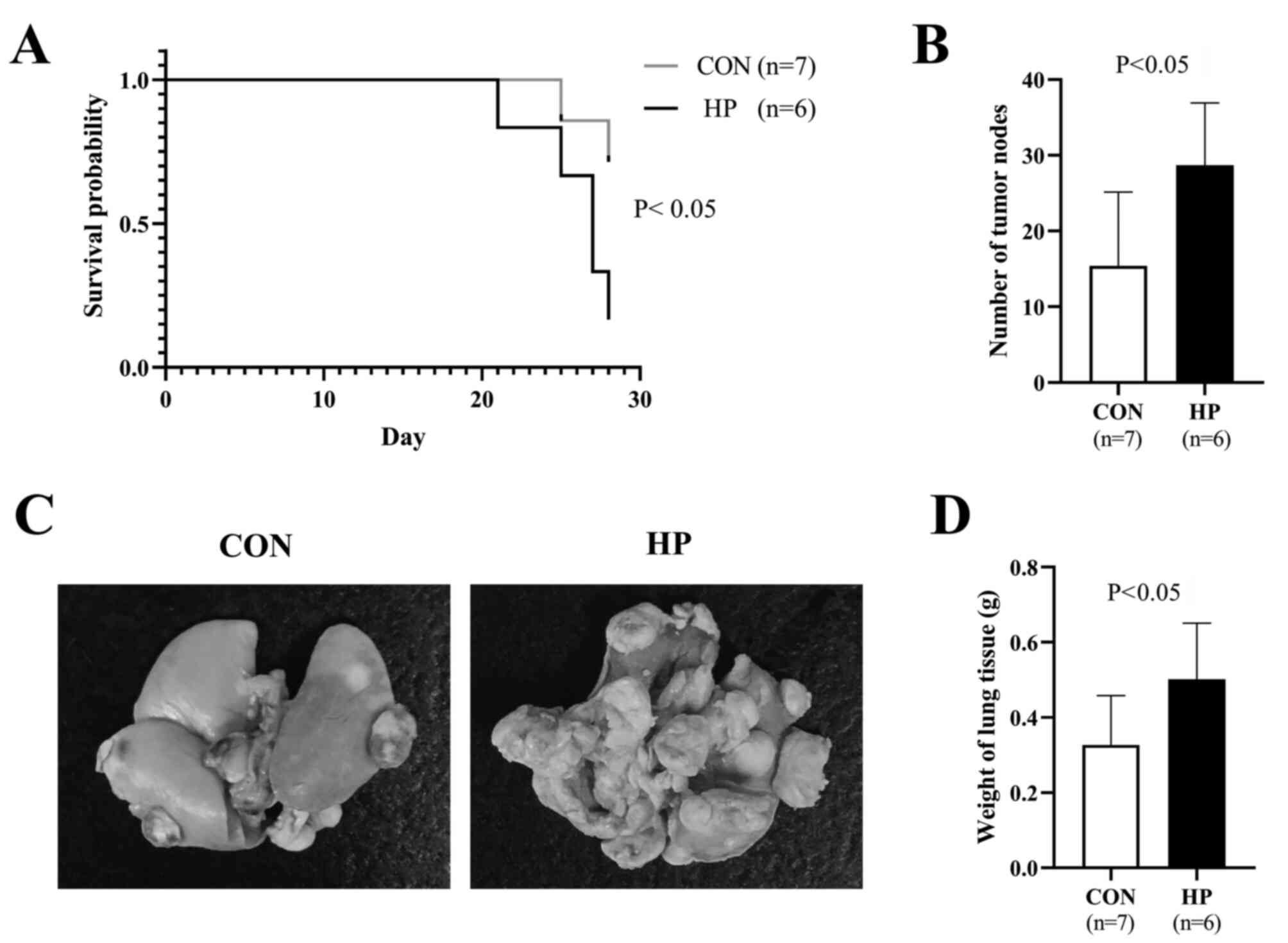
HP exposure promotes the metastasis of
the LLC cells to the lungs
To evaluate the metastatic potency in vivo,
the LLC cells were intravenously injected into healthy adult mice.
Compared with that in the mice that received LLC cells without HP
exposure, significantly worse survival was observed in the mice
that received LLC cells pretreated with 50 mmHg HP (P<0.05;
Fig. 5A). All the mice were killed 4
weeks following the injection of the cells and the maximum
percentage body weight loss observed was 9.3%. There were
significantly more metastatic tumor lesions in the lungs of the
mice in the HP group compared with that in the CON group
(P<0.05; Fig. 5B and C). The
weight of the lung tissue was also significantly higher in the HP
group compared with that in the CON group (P<0.05; Fig. 5D). These data suggested that HP
exposure promoted the metastasis of the LLC cells to the lungs.
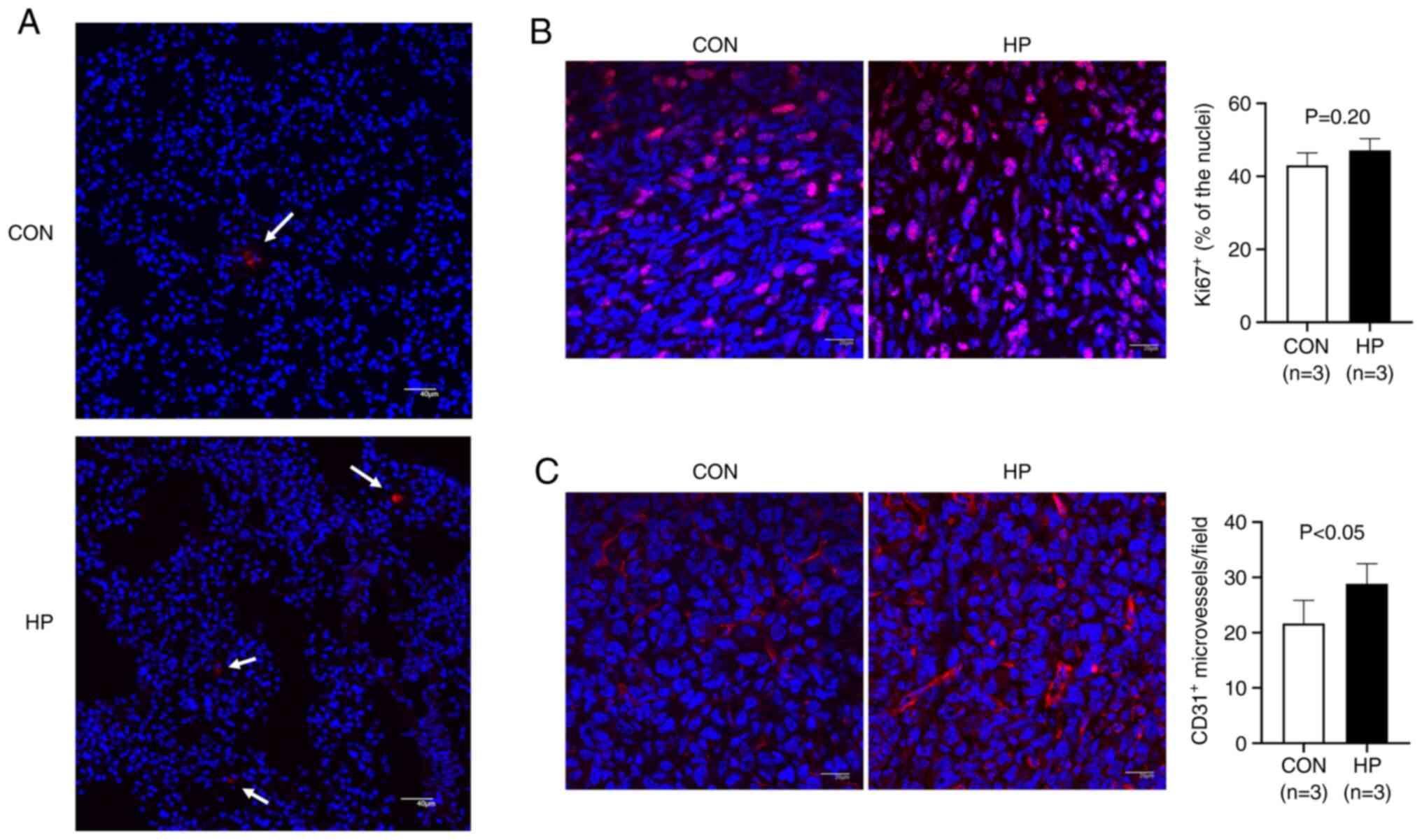
To further understand the mechanism involved, the
LLC cells were labelled with PKH26 before intravenous injection
into the mice, then the survival/retention of the cells in the
lungs was analyzed 24 h later. As expected, more LLC cells (or cell
clusters) were detected in the lungs from mice in the HP group
compared with that in the CON group (Fig.
6A), suggesting an improved survival/retention of the LLC cells
by pretreatment with 50 mmHg HP.
Cell proliferation and neovascularization in the
metastatic lesions was also analyzed using immunostaining. The
percentage of Ki67-positive cells was not significantly different
between the HP and CON groups (P=0.20; Fig. 6B). However, the density of the
CD31-positive microvessels in the metastatic lesions was
significantly higher in the HP group compared with that in the CON
group (P<0.05; Fig. 6C).
Discussion
Various mechanical forces within the
microenvironment of the tumor mass have been reported to play
critical roles in the progression of malignant tumors (2). Owing to the hyper-permeability of
immature capillaries, the elevation of the interstitial fluid HP
could be commonly induced by the presence of excess fluid
accumulation within malignant tumors (20,21). A
previous study has reported that HP may drive breast cancer cells
toward a more invasive phenotype (9);
however, the precise role and relevant mechanism of the mechanical
forces in mediating metastasis is still not well understood.
To investigate the role of HP on the metastatic
property of cancer cells, the LLC cells were exposed to 50 mmHg HP
to mimic the in vivo tumor microenvironment, then the mRNA
expression level of genes associated with tumor metastasis was
analyzed. A PCR array indicated noticeable changes, including the
upregulation of metastasis promotors (Hgf, Cdh11 and
Ephb2) and the downregulation of metastasis suppressors
(Kiss1, Syk and Htatip2) in the LLC cells following exposure
to 50 mmHg HP for 24 h. The most upregulated gene, Ephb2,
has been demonstrated to modulate the metastatic phenotype
(22) and induce angiogenesis
(23). The most downregulated gene,
Elane, has been demonstrated to modulate neutrophil
expression, inflammation and repair (24). Using an experimental lung metastasis
model in mice, it was further confirmed that the LLC cells
pretreated with 50 mmHg HP developed a significantly higher number
of tumor metastasis lesions in the lungs. These data suggested that
an elevated interstitial fluid HP in a rapidly growing malignant
tumor may enhance the metastatic property of cancer cells.
Additional experiments were performed to further
understand how HP enhanced the metastatic property of the LLC cells
from different aspects, according to the multi-step processes of
hematogenous metastasis. It is well-known that cancer cells enter
the circulation system and are exposed to hyperoxic arterial blood
for hematogenous metastasis (6).
Accumulating evidence suggests that oxidative stress kills most of
the circulating cancer cells, resulting in a very poor efficiency
of metastasis (6,7). Therefore, oxidative stress tolerance is
essential for the successful metastasis of cancer cells. HIF-1α is
well-known as an important mediator of metabolism reprogramming of
cancer cells by regulating antioxidant enzymes and antioxidant
properties (25). Notably, it has
been recently demonstrated that cyclic mechanical force stabilizes
HIF-1α by reducing protein degradation (13). Consistently, the results from the
present study showed the upregulation of HIF-1α protein expression
level in the LLC cells following exposure to 50 mmHg HP for 24 h.
In addition, the protein expression level of the antioxidant
enzymes, SOD1 and SOD2, the direct downstream targets of HIF-1α
(18,19), were also significantly increased in
cells exposed to 50 mmHg HP. This could contribute to enhancing
antioxidant capacity of cancer cells for remote hematogenous
metastasis. Consistent with the upregulation of various adhesion
molecules, the exposure to 50 mmHg HP also enhanced the adhesion
property of the LLC cells, as shown by the results of the in
vitro adhesion and the in vivo cell tracking assays.
HIFs are heterodimeric proteins composed of HIF-α
and HIF-1β subunits. HIF-1α is an O2-regulated subunit,
while HIF-1β is a constitutively expressed subunit (26). The protein expression level of HIF-1α
has been reported to be overexpressed in numerous malignant tumors,
including lung, prostate, breast and colon carcinomas (27,28). It
has been demonstrated that the enhanced protein expression level of
HIF-1α was associated with poor prognosis in patients with breast,
oropharyngeal and prostate cancer (29–31). As a
master regulator of cellular response to hypoxia, HIF-1 can induce
the transcription of several genes involved in angiogenesis, cell
proliferation and cell metabolism (15,32). One
of the most popularly recognized downstream genes of HIF-1 is
vascular endothelial growth factor (VEGF), which is known to induce
angiogenesis for the rapid growth of malignant tumors (33). VEGF, originally named as vascular
permeability factor, was first identified as a tumor-secreted
factor, which increases vascular permeability and promotes the
accumulation of ascite fluid (21).
Considering the hyper-permeability of microvessels in the tumor
(34), it is reasonable to
hypothesize that an excess accumulation of exudate in the
interstitial space contributes, at least in part, to the increase
of the interstitial fluid HP in the tumor mass. As a result,
elevated HP may stabilize HIF-1α, which thereby induces VEGF and
antioxidant enzymes to accelerate the growth and metastasis of
malignant tumors.
There is a caution in Cellosaurus that the LLC cells
(LL/2) could be identical to 3LL cells. It is reported that LL/2
cell line could be identical to 3LL cell line because both of them
are from mouse Lewis lung carcinoma and show the same biological
characteristics. Therefore, this will not affect the conclusion of
the present study.
A total of 4 weeks after the cells were injected
into the mice was used as the humane endpoint, based on clinical
signs (reduced intake and activity) and pathophysiological changes
(weight loss >20%). During the follow-up for 4 weeks, the
progression of tumor metastasis in the lungs of the mice was not
directly monitored; however, it was indirectly monitored by
observing the clinical signs (intake and activity) and
pathophysiological changes (weight loss) of mice.
For the 7 mice that died spontaneously, prior to the
end of the 4-week follow-up time in the lung metastasis mice model,
it was confirmed that the 7 mice did not die from lung
metastasis-induced respiratory failure or systemic cachexia. We
hypothesized that the cause of spontaneous death may be due to
another cause, based on the following: i) There were no signs of
severe clinical symptoms in the 7 mice following daily monitoring;
ii) based on the assessment of the exercised lungs, there were
fewer metastatic lesions in the lungs of the 7 mice; iii) according
to the examination of the 7 mice after sacrifice, there was no
serious bleeding, inflammation or purulent secretions in the body,
no notable signs of metastasis or organ necrosis was found in the
chest cavity or in any of the other organs and no obvious
occurrence of cachexia was found.
The present study has several limitations. First, a
single cell line was used and the cells were only exposed to 50
mmHg HP for all the experiments. This is due to the following
reasons: i) The present study was designed to examine whether an
elevated HP could promote the metastasis of cancer cells; ii) the
C57BL/6 mice were used for in vivo experiments and the LLC
cell line is the most reproducible syngeneic model for evaluating
lung metastasis to date (35); iii)
interstitial fluid pressure in solid malignant tumors could be
elevated to ~30 mmHg HP (12,36) and iv) the exposure of the LLC cells to
50 mmHg HP altered the mRNA expression level of genes associated
with metastasis; however, higher pressure (100 mmHg) induced cell
death and cell debris production (data not shown). Therefore,
further experiments are required to exposure different cancer cell
lines with different HPs. Second, the PCR array was not repeated
due to a limited budget. In addition, the fold-change result may
also have greater variations if P>0.05; therefore, it is
important to have a sufficient number of biological replicates to
validate the array data. However, a mixture of RNA samples was used
from three independent experiments to generate the cDNA for a
single PCR array in each group. Therefore, the PCR array data was
expressed as the average level in 3 samples from each group. Third,
further interventional experiments, such as the interference of the
HIF-1α signaling pathway was not performed, as silencing HIF-1α
alone would change cell biological properties. Furthermore,
multiple factors, including the increase in mRNA expression level
of HIF-1α and adhesion molecules could be involved in the
HP-induced cancer cell metastasis; therefore, a genetic
intervention approach to directly confirm the role of HIF-1α was
not performed in the present study. Forth, Annexin V-positive
apoptotic cells were only analyzed using flow cytometry and the
expression level of other apoptotic proteins, such as the caspase
family, can also be used to indicate apoptosis. In addition, a
colony-forming assay was not included, as the potential role of HP
in cancer cell metastasis, rather than tumorigenesis and tumor
growth was the aim of the present study.
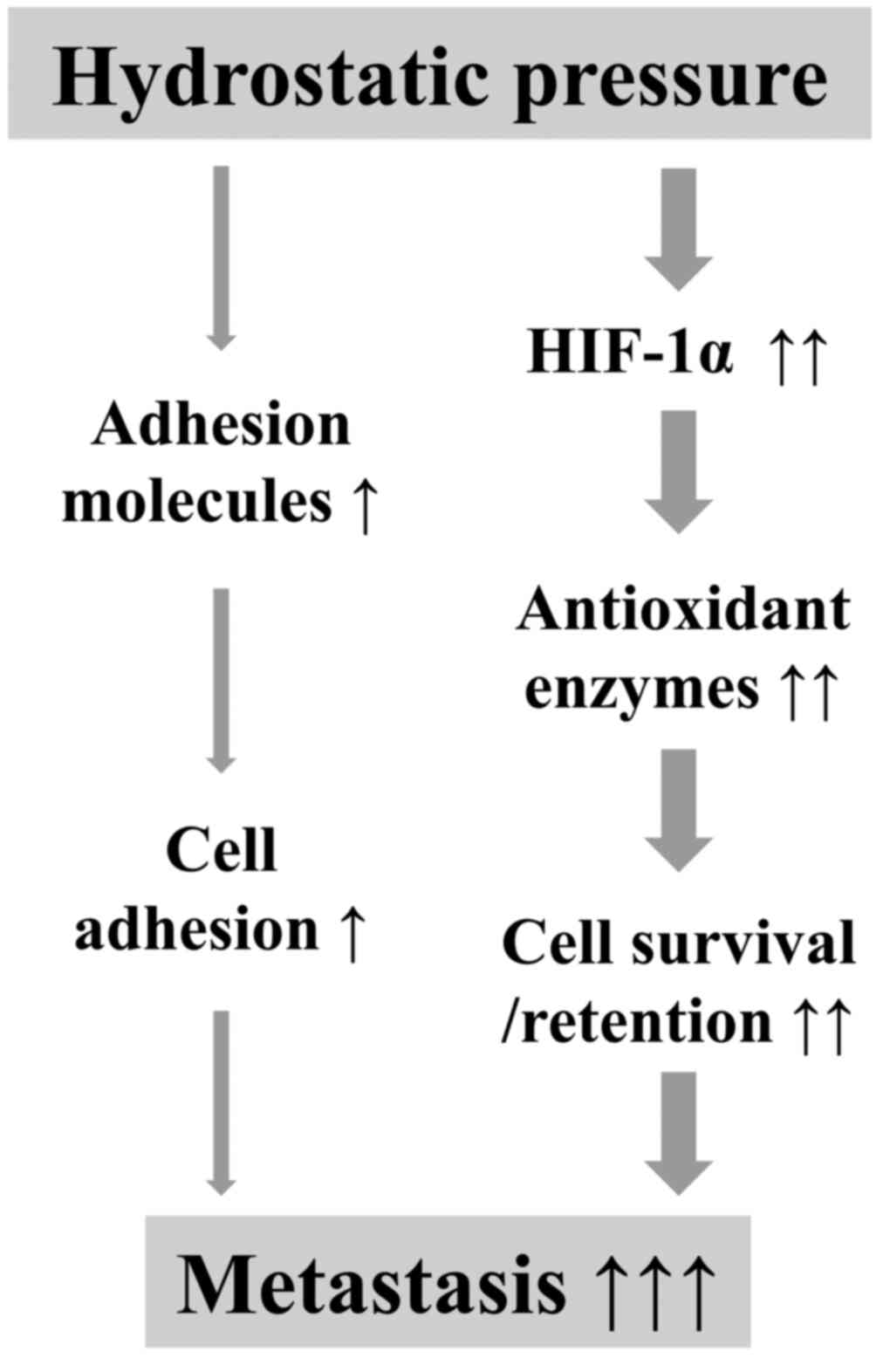
From the results in the present study, an elevated
HP in rapidly growing malignant tumors may enhance the metastatic
potency of cancer cells via complex mechanisms, including the
increase in the mRNA expression level of adhesion molecules to
improve cell adhesion and the stabilization of HIF-1α to induce the
expression of antioxidant enzymes to defend against oxidative
damage during metastasis (Fig. 7). It
is critical to elucidate the comprehensive molecular mechanisms
underlying the stabilization of HIF-1α by HP in further
investigations.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported in part by a Grant-in-Aid
from the Ministry of Education, Science, Sports, Culture and
Technology, Japan (grant no. 17H04265), the Collaborative Research
Program of the Atomic-bomb Disease Institute of Nagasaki University
and the Japan China Sasakawa Medical Fellowship.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
All the authors contributed to the conception and
design of the study. DZ, YX and LA performed the experiments and
acquired the data. TSL, DZ, YX, LA and XZ analyzed and interpreted
the data. DZ and SZ drafted the manuscript. TSL, JL and CG
critically revised the manuscript for important intellectual
content. TSL and DZ confirmed the authenticity of all the raw data.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the
Institutional Animal Care and Use Committee of Nagasaki University
(approval no. 1608251335-11) and all animal procedures were
performed in accordance with institutional and national
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brabletz T, Lyden D, Steeg PS and Werb Z:
Roadblocks to translational advances on metastasis research. Nat
Med. 19:1104–1109. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bregenzer ME, Horst EN, Mehta P, Novak CM,
Repetto T and Mehta G: The role of cancer stem cells and mechanical
forces in ovarian cancer metastasis. Cancers. 11:10082019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eccles SA and Welch DR: Metastasis: Recent
discoveries and novel treatment strategies. Lancet. 369:1742–1757.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fidler IJ: The pathogenesis of cancer
metastasis: The ‘seed and soil’ hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zijl FV, Krupitza G and Mikulits W:
Initial steps of metastasis: Cell invasion and endothelial
transmigration. Mutat Res. 728:23–34. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Piskounova E, Agathocleous M, Murphy MM,
Hu Z, Huddlestun SE, Zhao Z, Leitch AM, Johnson TM, DeBerardinis RJ
and Morrison SJ: Oxidative stress inhibits distant metastasis by
human melanoma cells. Nature. 527:186–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vanharanta S and Massague J: Origins of
metastatic traits. Cancer Cell. 24:410–421. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kao YC, Jheng JR, Pan HJ, Liao WY, Lee CH
and Kuo PL: Elevated hydrostatic pressure enhances the motility and
enlarges the size of the lung cancer cells through aquaporin
upregulation mediated by caveolin-1 and ERK1/2 signaling. Oncogene.
36:863–874. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tse JM, Cheng G, Tyrrell JA,
Wilcox-Adelman SA, Boucher Y, Jain RK and Munn LL: Mechanical
compression drives cancer cells toward invasive phenotype. Proc
Natl Acad Sci USA. 109:911–916. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Less JR, Posner MC, Boucher Y, Borochovitz
D, Wolmark N and Jain RK: Interstitial hypertension in human breast
and colorectal tumors. Cancer Res. 52:6371–6374. 1992.PubMed/NCBI
|
|
11
|
Nathan SS, DiResta GR, Casas-Ganem JE,
Hoang BH, Sowers R, Yang R, Huvos AG, Gorlick R and Healey JH:
Elevated physiologic tumor pressure promotes proliferation and
chemosensitivity in human osteosarcoma. Clin Cancer Res.
11:2389–2397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gutmann R, Leunig M, Feyh J, Goetz AE,
Messmer K, Kastenbauer E and Jain RK: Interstitial hypertension in
head and neck tumors in patients: Correlation with tumor size.
Cancer Res. 52:1993–1995. 1992.PubMed/NCBI
|
|
13
|
Solis AG, Bielecki P, Steach HR, Sharma L,
Harman CC, Yun S, de Zoete MR, Warnock JN, To SDF, York AG, et al:
Mechanosensation of cyclical force by PIEZO1 is essential for
innate immunity. Nature. 573:69–74. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rankin EB, Nam JM and Giaccia AJ: Hypoxia:
Signaling the metastatic cascade. Trends Cancer. 2:295–304. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Semenza GL: Hypoxia-inducible factors in
physiology and medicine. Cell. 148:399–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Semenza GL: Regulation of cancer cell
metabolism by hypoxia-inducible factor 1. Semin Cancer Biol.
19:12–16. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Urata Y, Goto S, Luo L, Doi H, Kitajima Y,
Masuda S, Ono Y and Li TS: Enhanced Nox1 expression and oxidative
stress resistance in c-kit-positive hematopoietic stem/progenitor
cells. Biochem Biophys Res Commun. 454:376–380. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu XQ, Song R and Zhang L: Effect of
oxidative stress on the estrogen-NOS-NO-KCa channel pathway in
uteroplacental dysfunction: Its implication in pregnancy
complications. Oxid Med Cell Longev. 2019:91942692019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Novak S, Drenjancevic I, Vukovic R,
Kellermayer Z, Cosic A, Tolusic Levak M, Balogh P, Culo F and
Mihalj M: Anti-inflammatory effects of hyperbaric oxygenation
during DSS-induced colitis in BALB/c mice include changes in gene
expression of HIF-1α, proinflammatory cytokines, and antioxidative
enzymes. Mediators Inflamm. 2016:71414302016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jain RK, Martin JD and Stylianopoulos T:
The role of mechanical forces in tumor growth and therapy. Annu Rev
Biomed Eng. 16:321–346. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Senger DR, Galli SJ, Dvorak AM, Perruzzi
CA, Harvey VS and Dvorak HF: Tumor cells secrete a vascular
permeability factor that promotes accumulation of ascites fluid.
Science. 219:983–985. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu YL, Horning AM, Lieberman B, Kim M,
Lin CK, Hung CN, Chou CW, Wang CM, Lin CL, Kirma NB, et al: Spatial
EGFR dynamics and metastatic phenotypes modulated by upregulated
EphB2 and Src pathways in advanced prostate cancer. Cancers
(Basel). 11:19102019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sato S, Vasaikar S, Eskaros A, Kim Y,
Lewis JS, Zhang B, Zijlstra A and Weaver AM: EPHB2 carried on small
extracellular vesicles induces tumor angiogenesis via activation of
ephrin reverse signaling. JCI Insight. 4:e1324472019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Makaryan V, Zeidler C, Bolyard AA, Skokowa
J, Rodger E, Kelley ML, Boxer LA, Bonilla MA, Newburger PE,
Shimamura A, et al: The diversity of mutations and clinical
outcomes for ELANE-associated neutropenia. Curr Opin Hematol.
22:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakashima R, Goto Y, Koyasu S, Kobayashi
M, Morinibu A, Yoshimura M, Hiraoka M, Hammond EM and Harada H:
UCHL1-HIF-1 axis-mediated antioxidant property of cancer cells as a
therapeutic target for radiosensitization. Sci Rep. 7:68792017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Semenza GL: Pharmacologic targeting of
hypoxia-inducible factors. Annu Rev Pharmacol Toxicol. 59:379–403.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhong H, De Marzo AM, Laughner E, Lim M,
Hilton DA, Zagzag D, Buechler P, Isaacs WB, Semenza GL and Simons
JW: Overexpression of hypoxia-inducible factor 1alpha in common
human cancers and their metastases. Cancer Res. 59:5830–5835.
1999.PubMed/NCBI
|
|
28
|
Talks KL, Turley H, Gatter KC, Maxwell PH,
Pugh CW, Ratcliffe PJ and Harris AL: The expression and
distribution of the hypoxia-inducible factors HIF-1alpha and
HIF-2alpha in normal human tissues, cancers, and tumor-associated
macrophages. Am J Pathol. 157:411–421. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Generali D, Berruti A, Brizzi MP, Campo L,
Bonardi S, Wigfield S, Bersiga A, Allevi G, Milani M, Aguggini S,
et al: Hypoxia-inducible factor-1alpha expression predicts a poor
response to primary chemoendocrine therapy and disease-free
survival in primary human breast cancer. Clin Cancer Res.
12:4562–4568. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aebersold DM, Burri P, Beer KT, Laissue J,
Djonov V, Greiner RH and Semenza GL: Expression of
hypoxia-inducible factor-1alpha: A novel predictive and prognostic
parameter in the radiotherapy of oropharyngeal cancer. Cancer Res.
61:2911–2916. 2001.PubMed/NCBI
|
|
31
|
Nanni S, Benvenuti V, Grasselli A, Priolo
C, Aiello A, Mattiussi S, Colussi C, Lirangi V, Illi B, D'Eletto M,
et al: Endothelial NOS, estrogen receptor beta, and HIFs cooperate
in the activation of a prognostic transcriptional pattern in
aggressive human prostate cancer. J Clin Invest. 119:1093–1108.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Semenza GL: Defining the role of
hypoxia-inducible factor 1 in cancer biology and therapeutics.
Oncogene. 29:625–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Apte RS, Chen DS and Ferrara N: VEGF in
signaling and disease: Beyond discovery and development. Cell.
176:1248–1264. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dvorak HF: Vascular permeability
factor/vascular endothelial growth factor: A critical cytokine in
tumor angiogenesis and a potential target for diagnosis and
therapy. J Clin Oncol. 20:4368–4380. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kellar A, Egan C and Morris D: Preclinical
murine models for lung cancer: Clinical trial applications. Biomed
Res Int. 2015:6213242015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mori T, Koga T, Shibata H, Ikeda K,
Shiraishi K, Suzuki M and Iyama K: Interstitial fluid pressure
correlates clinicopathological factors of lung cancer. Ann Thorac
Cardiovasc Surg. 21:201–208. 2015. View Article : Google Scholar : PubMed/NCBI
|