Introduction
Breast cancer is the most commonly diagnosed type of
cancer and is a prevailing cause of cancer-associated mortality
among women. As previously reported, female breast cancer, with an
estimated 2.3 million new cases (11.7%) and 684,996
cancer-associated mortalities globally,surpassed lung cancer as the
most commonly diagnosed cancer type in 2020 (1). The majority of breast cancer-associated
mortalities are caused by distant metastasis (2). Decades of research on cancer have led to
a substantial progress in the treatment of primary breast tumors;
however, treatment options for metastatic cancer remain limited
(3).
Accumulating evidence has revealed that cancer stem
cells (CSCs), which constitute a small number of tumor cells in
liquid and solid tumors, possess self-renewal capability and
contribute to tumor onset, resistance, recurrence and metastasis
(4). Breast CSCs (BCSCs) can be
identified by various functional assays, including tumor sphere
formation, xenograft assay or detection of specific cell-surface
markers such as CD44, CD24, Oct-4 and aldehyde dehydrogenase (ALDH)
(5). Another important factor
contributing to metastasis is epithelial-to-mesenchymal transition
(EMT), a process during which epithelial cells lose their polarity
and cell-to-cell contact, acquiring additional migratory and
invasive properties (6), which allows
cancer cells to detach from neighboring cells, dissolve the basal
membrane and invade the extracellular matrix (ECM) (7). Several molecular mechanisms are known to
promote EMT, including the regulation of specific cell-surface
protein and ECM-degrading enzyme expression levels, and the
alteration of the expression of certain transcription factors. The
EMT process during breast carcinogenesis is considered to be
controlled by a series of signaling pathways, including PI3K/AKT
(7), Notch (8), Wnt/β-catenin (9) and Hedgehog (4,8), being
also responsible for CSC maintenance (4,10).
Ubiquilin-1 (UBQLN1) belongs to a family of
ubiquitin-like proteins that contain five major UBQLN proteins
(UBQLN1-4 and UBQLNL). These proteins are evolutionarily conserved
and structurally similar, containing an amino-terminal
ubiquitin-like (UBL) domain, a carboxy-terminal
ubiquitin-associated (UBA) domain and a series of four
chaperonin-like domains within the central part of the protein
(6,11). The UBL domain mediates interaction
with the proteasome, whereas the UBA domain preferentially binds to
ubiquitinated proteins (12). UBQLNs
appear to function as adaptors to deliver ubiquitinated proteins to
the proteasome (13). UBQLNs are
essential factors for the maintenance of proteostasis in cells,
since they are important for the regulation of different protein
degradation mechanisms and pathways, including the
ubiquitin-proteasome system (UPS), autophagy and endoplasmic
reticulum-associated protein degradation pathways (6). The UBQLN1 gene is located on
human chromosome 9q22 and is ubiquitously expressed in all human
tissues (14). The role of UBQLN1 has
mainly been studied in neurological disorders. The disruption of
UBQLN1 function has been reported in a variety of neurological
disorders caused by aberrant protein aggregation, such as
Alzheimer's disease (15),
amyotrophic lateral sclerosis (16)
and Huntington's disease (17).
Previous studies have indicated that UBQLN1
participates in the progression of certain types of cancer,
including lung adenocarcinoma (18),
gastric adenocarcinoma (12) and
ovarian cancer (19). Elevated UBQLN1
levels have been revealed to be associated with a poor prognosis of
patients with breast cancer (20),
lung and gastric cancer (12). UBQLN1
has also been reported to be associated with the stemness of cells
such as human neuronal stem cell line (21) and human embryonic stem cells (22). However, UBQLN1 expression, and
biological function and mechanisms in breast cancer remain largely
unknown. In the present study, UBQLN1 expression in breast cancer
tissues was examined, its clinical significance was explored, the
effect of UBQLN1 knockdown on cell migration, invasion, EMT and
stemness was investigated and the possible mechanisms of UBQLN1 in
the progression of breast cancer were evaluated.
Materials and methods
Cells and cell culture
The human normal mammary epithelial cell line,
MCF-10A (SCSP-575), and the human breast cancer cell lines, MCF-7
(SCSP-531) and MDA-MB-231 (SCSP-5043), were obtained from The Cell
Bank of Type Culture Collection of the Chinese Academy of Sciences.
The MCF-7 cell line was cultured in DMEM/F12 (HyClone; Cytiva) with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.). MDA-MB-231 cells
were cultured in MEM α modification medium (HyClone; Cytiva)
supplemented with 10% FBS. MCF-10A cells were maintained in
DMEM/F12 supplemented with 10% FBS, 20 ng/ml human recombinant
epidermal growth factor (R&D Systems, Inc.), 0.5 µg/ml
hydrocortisone (Sigma-Aldrich; Merck KGaA), 10 µg/ml insulin
(Sigma-Aldrich; Merck KGaA), 0.1 µg/ml cholera toxin
(Sigma-Aldrich; Merck KGaA) and L-glutamine (Invitrogen; Thermo
Fisher Scientific, Inc.). Cells were maintained at 37°C and 5%
CO2.
Lentiviral construction and virus
infection
Lentiviral vectors, including short hairpin RNA
(shRNA/sh) against UBQLN1 (shUbqln1) and negative control shRNA
(shScramble) were constructed by Shanghai GenePharma Co., Ltd.
(LV3-UQLNQ1-homo1642). The sequence of shUbqln1 was as follows:
5′-GAGTACTACTGCGCCAAAT-3′. The sequence of shScramble was as
follows: Sence, 5′-UUCUCCGAACGUGUCACGUTT-3′; antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. To silence UBQLN1, the MCF-7 and
MDA-MB-231 cells were cultured in serum-free medium containing
lentivirus (multiplicity of infection, 50) and 5 µg/ml Polybrene
(GenePharma Co., Ltd.) for 6 h and then incubated in complete
growth medium for 48 h at 37°C. The minimal lethal concentration of
puromycin (Clontech Laboratories, Inc.) (10 µg/ml) was used to
selected stably transfected cells. Knockdown efficiency was
determined by the use of western blot analysis and reverse
transcription-quantitative PCR (RT-qPCR).
Isolation of
CD44+/CD24− BCSCs
CD44+/CD24− BCSCs were
separated using the magnetic-activated cell sorting system (MACS)
(Miltenyi Biotec GmbH) with CD44 MicroBeads and CD24 Microbeads kit
(both from Miltenyi Biotec GmbH), according to the manufacturer's
protocol. Breast cancer cells were collected and resuspended in 40
µl PBS with 0.5% BSA (Beyotime Institute of Biotechnology) and 2 mM
EDTA (PBE) per 107 cells, incubated with CD24-biotin and
then incubated with anti-biotin microbeads following the
manufacturer's protocol of the CD24 Microbeads kit. Subsequently,
the cells were magnetically separated. Briefly, the cell suspension
was applied to MiniMACS columns (Miltenyi Biotec GmbH) in the
magnetic field of the MACS Separator (Miltenyi Biotec GmbH).
Unlabeled cells (CD24−) were obtained by collecting the
flow-through cells, while labeled cells (CD24+ cells)
were collected by pipetting 1 ml of PBE buffer onto the column.
CD24− cells were incubated with CD44 MicroBeads at 4°C
for 15 min. Subsequently, the cells were magnetically separated
again. CD44+/CD24− cells were obtained by
collecting labeled cells from the column.
MTT assay
For the detection of cell viability, transfected
MCF-7 and MDA-MB-231 cells were seeded in 96-well plates, at a
density of 2×103 cells/well. Cell viability was assessed
by MTT assay following 0, 24, 48 and 72 h of incubation at 37°C.
For the evaluation of cell viability, the transfected cancer cells
were seeded in 96-well plates at a density of 2×104
cells/well. Cell viability in each group was assessed by MTT assay
following the addition of various concentrations (0, 5, 10, 50 and
100 ng/ml) of paclitaxel (Beyotime Institute of Biotechnology) and
incubation at 37°C for 48 h. Subsequently, MTT (Beyotime Institute
of Biotechnology) was added to each well at a final concentration
of 5 mg/ml and the cells were incubated 4 h at 37°C. The medium was
then aspirated, followed by the addition of 100 µl DMSO to each
well. The plates were then agitated for 10 min at room temperature,
and the absorbance at 490 nm was measured using a spectrophotometer
(Thermal Fisher Scientific, Inc.). Each value represented the mean
± SEM. Responses to drug treatment were assessed by normalizing the
treatment groups to the untreated controls.
Protein extraction and western blot
analysis
Cells (1×106) were washed with cold PBS
and harvested. Total protein lysate was prepared by cell sonication
in ice-cold standard RIPA lysis buffer (Sigma-Aldrich; Merck KGaA)
with proteinase and phosphatase inhibitors (Santa Cruz
Biotechnology, Inc.) for 10 sec. The protein concentration was
measured using the Easy II Protein Quantitative kit (TransGen
Biotech Co., Ltd.). Protein lysate (50–80 µg) was separated by
8–12% SDS-PAGE and transferred onto PVDF membranes
(MilliporeSigma). After blocking in a 5% skimmed milk solution in
TBS containing 0.05% Tween-20 (TBST) buffer for 1 h at room
temperature, the membranes were incubated with the corresponding
specific antibody solution at 4°C overnight and then with
IRDye® 800 CW goat anti-rabbit second antibody (cat. no.
926-82211; LI-COR Biosciences; 1:16,000) at 37°C for 1 h. The
immunoreactive bands were detected and analyzed using an Odyssey
infrared imaging system 3.0 (LI-COR Biosciences). Protein
expression levels were normalized to GAPDH expression levels.
The specific antibodies used for western blot
analysis were as follows: Rabbit polyclonal anti-human UBQLN1 (cat.
no. ab3341; Abcam; 1:500); rabbit monoclonal anti-human
phosphorylated (p)-AKT (Ser473) (cat. no. 4060; Cell Signaling
Technology, Inc.; 1:500); monoclonal anti-human GAPDH (cat. no.
sc-47724; Santa Cruz Biotechnology, Inc.; 1:10,000); and anti-PTEN
(cat. no. 22034-1-AP; 1:500), anti-AKT (cat. no. 10176-2-AP;
1:1,000) anti-Oct-4 (cat. no. 11263; 1:500), anti-Sox2 (cat. no.
11064; 1:500), anti-ALDH1 (cat. no. 15910; 1:500), anti-MMP2 (cat.
no. 10373; 1:1,000), anti-MMP9 (cat. no. 27306; 1:1,000),
anti-Snail (cat. no. 13099; 1:500), anti-Twist (cat. no. 25465;
1:500), anti-Bcl-2 (cat. no. 12789; 1:500), anti-Bax (cat. no.
60267-1-Ig; 1:500), anti-E-cadherin (cat. no. 20874; 1:1,000),
anti-vimentin (cat. no. 10366-1-AP; 1:1,000), anti-caspase-3 (cat.
no. 19677; 1:500), anti-caspase-9 (cat. no. 10380; 1:500) and
anti-N-cadherin (cat. no. 22018; 1:500) (all rabbit polyclonal
anti-human antibodies were purchased from ProteinTech Group,
Inc.
RNA extraction and RT-qPCR
analysis
Total RNA was extracted from 1×106 cells
using TRIzol® reagent (TransGen Biotech Co., Ltd.).
Total RNA (1 µg) was reverse transcribed into cDNA using
TransScript® All-in-One First-Strand cDNA Synthesis
SuperMiX kit (TransGen Biotech Co., Ltd.). The resulting cDNA
samples were amplified by qPCR with SYBR Premix Ex Taq Master MIx
kit (TransGen Biotech Co., Ltd.) in a 20-µl reaction mixture using
an iCycler iQ™ Real Time PCR Detection System (Agilent
Technologies, Inc.). The PCR cycles were performed as follows: One
cycle at 94°C for 10 min, 40 cycles at 95°C for 5 sec, 60°C for 15
sec, 72°C for 10 sec, and one final cycle at 72°C for 10 min,
followed by cooling to 4°C. The primers were synthesized by
Invitrogen (Thermo Fisher Scientific, Inc.), and the sequences were
as follows: UBQLN1 forward, 5′-GAACCAGGACCGAGCTTGA-3′ and reverse,
5′-TGTATTGCTCACCAAGGAAGCA-3′; ALDH1 forward,
5′-TGCAGGTTGGGCTGACAA-3′ and reverse, 5′-GCAGGCCCTATCTTCCAAATG-3′;
and GAPDH forward, 5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse,
5′-TGGTGAAGACGCCAGTGGA-3′. The relative expression levels of mRNA
were normalized to the internal control (GAPDH), and fold-changes
were calculated using the 2−ΔΔCq method (23).
Tumor cell migration and invasion
assays
Invasion assays were performed using 24-well
Transwell chambers (8.0-µm pore size; Corning, Inc.). Each
Transwell chamber was coated with Matrigel matrix (Corning, Inc.)
(100 µl at a dilution of 1:3 in DMEM; BD Biosciences), 24 h prior
to use. Cells were cultured in DMEM for 24 h and then seeded onto
cell inserts (2×104 cells/insert) in the upper chamber.
Serum-free DMEM was added to the upper chamber, while DMEM (0.5 ml)
containing 10% FBS was added to the lower chamber. After 24 h, the
upper cells were removed, and the cells on the surface of the
bottom chamber were fixed with methanol for 15 min and stained with
0.01% crystal violet (MedChemExpress) for 30 min at room
temperature. Randomly selected areas were imaged with an inverted
microscope (Olympus IX73; Olympus Corporation) connected to an
Olympus DP73 camera (Olympus Corporation), and the number of cells
was counted. The results represent the mean number of cells in five
fields per membrane for triplicate inserts. Cell migration assays
were conducted as the invasion assays, with the exception of the
Matrigel coating.
Wound-healing assay
When cultured cells reached 70–80% confluence in
6-well plates, a 100-µl pipette tip was used to produce scratches
in the cell monolayer. Subsequently, the cells were washed with PBS
and cultured in DMEM without FBS for ≤72 h. Images were observed
and captured at 0, 24, 48 and 72 h with an inverted microscope
(Olympus IX73; Olympus Corporation) connected to an Olympus DP73
camera (Olympus Corporation). All assays were performed in
triplicate.
Colony formation assay
Breast cancer cells were plated at a density of
1,000 cells/well in a 6-well plate. Following 2 weeks of incubation
at 37°C in complete medium, colonies were fixed with methanol for
15 min and stained with 0.01% crystal violet for 30 min at room
temperature. The clones were then counted and imaged using an
inverted microscope (Olympus IX73; Olympus Corporation) connected
to an Olympus DP73 camera (Olympus Corporation). The holoclones
were counted as previously reported (5).
Immunofluorescence analysis
Cells were cultured on a cover slide in 24-well
plates at 37°C, fixed in 4% paraformaldehyde for 15 min and
permeabilized with 100% ice-cold methanol at −20°C for 10 min.
After washing with PBS, the cells were blocked in 5% BSA (Beyotime
Institute of Biotechnology) containing 0.3% Triton™ X-100 for 2 h
at 4°C. The cells were then washed with TBST and incubated
overnight at 4°C with primary antibodies against UBQLN1 (cat. no.
ab3341; Abcam; 1:100), p-AKT (cat. no. 4060; Cell Signaling
Technology, Inc.; 1:100), PTEN (cat. no. 22034; 1:100), E-cadherin
(cat. no. 20874; ProteinTech Group, Inc.; 1:100) and N-cadherin
(cat. no. 22018; ProteinTech Group, Inc.; 1:100). Subsequently, the
cells were incubated with a FITC-conjugated secondary antibody
(cat. no. 408308; BioLegend, Inc.) for 1 h at 4°C. Cell nuclei were
counterstained with DAPI (Boster Biological Technology). Cells were
then observed under an Olympus BX41 fluorescence microscope
(Olympus Corporation) and photographed using an Olympus DP72 camera
(Olympus Corporation).
Mammosphere formation assay
MCF-7 and MDA-MB-231 cells were inoculated into
ultralow attachment 6-well plates (Corning, Inc.) at a density of
4×104 cells/well. The cells were grown in complete
medium supplemented with B27 (1:50; Invitrogen; Thermo Fisher
Scientific, Inc.), 5 µg/ml insulin (Sigma-Aldrich; Merck KGaA), 20
ng/ml human EGF (Sigma-Aldrich; Merck KGaA), 4 µg/ml heparin
(Sigma-Aldrich; Merck KGaA) and 20 ng/ml basic fibroblast growth
factor (Sigma-Aldrich; Merck KGaA) for 14 days. Cell colonies
>60 µm in diameter were counted under an inverted microscope
(Olympus Corporation).
Immunohistochemistry (IHC)
A tissue array slide containing 73 cases of breast
cancer tissues with complete clinicopathological data including
age, tumor diameter, lymph node metastasis, TNM stage, histology,
differentiation, molecular subtypes, estrogen receptor,
progesterone receptor, HER2 and proliferating cell nuclear antigen
expression, was used as reported previously (24). The mean age of the patients was
52.22±10.79 years with a range between 29 and 81 years. A total of
30 normal breast tissues were collected from patients with
fibrocystic breast disease. The breast cancer tissues and breast
tissues were obtained by tumor resection surgery at the Affiliated
Hospital of Dalian Medical University. Written informed consent was
obtained from all individual participants included in the study.
The present study was conducted according to the principles of the
Declaration of Helsinki and was reviewed and approved by the Ethics
Committee and Institutional Review Board (IRB) of Dalian Medical
University (IRB approval no. 2021006).
Tissue sections were routinely deparaffinized in
xylene and rehydrated in a series of 100-50% ethanol solutions.
Endogenous peroxidase activity was blocked by incubation with 3%
hydrogen peroxide in PBS for 15 min at room temperature. The tissue
sections were then blocked with 10% normal goat serum (OriGene
Technologies, Inc.) for 1 h at room temperature and then incubated
with a primary antibody against UBQLN1 (cat. no. ab3341; Abcam)
(1:100) overnight at 4°C. A biotin-streptavidin HRP and AP
detection for mouse and rabbit antibody on human tissue (cat. no.
D03-6; OriGene Technologies, Inc.) was used for immunodetection
following the manufacturer's instructions. In brief, all slides
were incubated with a secondary antibody for 1 h at 4°C and then
with a streptavidin-biotin solution for 30 min at room temperature.
The sections were then subjected to a colorimetric reaction using a
DAB detection kit (OriGene Technologies, Inc.). Finally, the
sections were briefly counterstained with hematoxylin. A negative
control staining was also performed, by replacing the primary
antibodies with normal goat serum (OriGene Technologies, Inc.). The
results of IHC were blindly evaluated by two independent
pathologists. The scoring criteria were based on staining intensity
(1, no or weak staining; or 2, medium or strong staining). The
immunostained tissues were scored by multiplying the intensity
(1–2)
and extent (0-100%) of staining (25). Total score cut-offs of ≤100 and
>100 were used to divide patients into low and high UBQLN1
expression groups, respectively.
Bioinformatics analysis
TNMplot (https://www.tnmplot.com/), Kaplan-Meier plotter
(KmPlot) (http://kmplot.com/analysis/), cBioPortal (http://www.cbioportal.org), TIMER2.0 (http://timer.cistrome.org) and UALCAN (http://ualcan.path.uab.edu) were utilized to analyze
the differential expression, prognostic value, genetic alteration,
gene promotor methylation and clinicopathological significance of
UBQLN1 in patients with breast cancer. TIMER2.0 was used to
evaluate the outcome significance of UBQLN1 mRNA expression,
optionally adjusted by clinical factors, including age, sex, stage
and race using the Cox proportional hazard model. A threshold of
P<0.05 was used to set the cut-off criterion. A protein-protein
interaction (PPI) network was obtained using STRING database
(http://string-db.org).
Statistical analysis
Statistical analysis was performed using SPSS
version 19.0 (IBM Corp.) and GraphPad Prism 5 (GraphPad Software,
Inc.). Associations between clinicopathological characteristics and
UBQLN1 expression were examined using the χ2 test.
Differences between two groups were evaluated using a paired or
unpaired Student's t-test. One-way ANOVA and Dunnett's multiple
comparisons test were used for comparing multiple groups. Each set
of results represents ≥3 separate experiments. All experimental
data are expressed as the mean ± SE. P<0.05 was considered to
indicate a statistically significant difference.
Results
UBQLN1 expression is significantly
increased in breast cancer, and a high UBQLN1 expression is
associated with a poor prognosis of patients with breast
cancer
UBQLN1 is abnormally expressed in certain cancer
types, including lung adenocarcinoma (11,18) and
gastric cancer (12). In the present
study, to elucidate UBQLN1 expression in breast cancer, UBQLN1
expression was detected in a tissue array slide, containing samples
from 73 breast cancer cases and 30 normal breast tissues by using
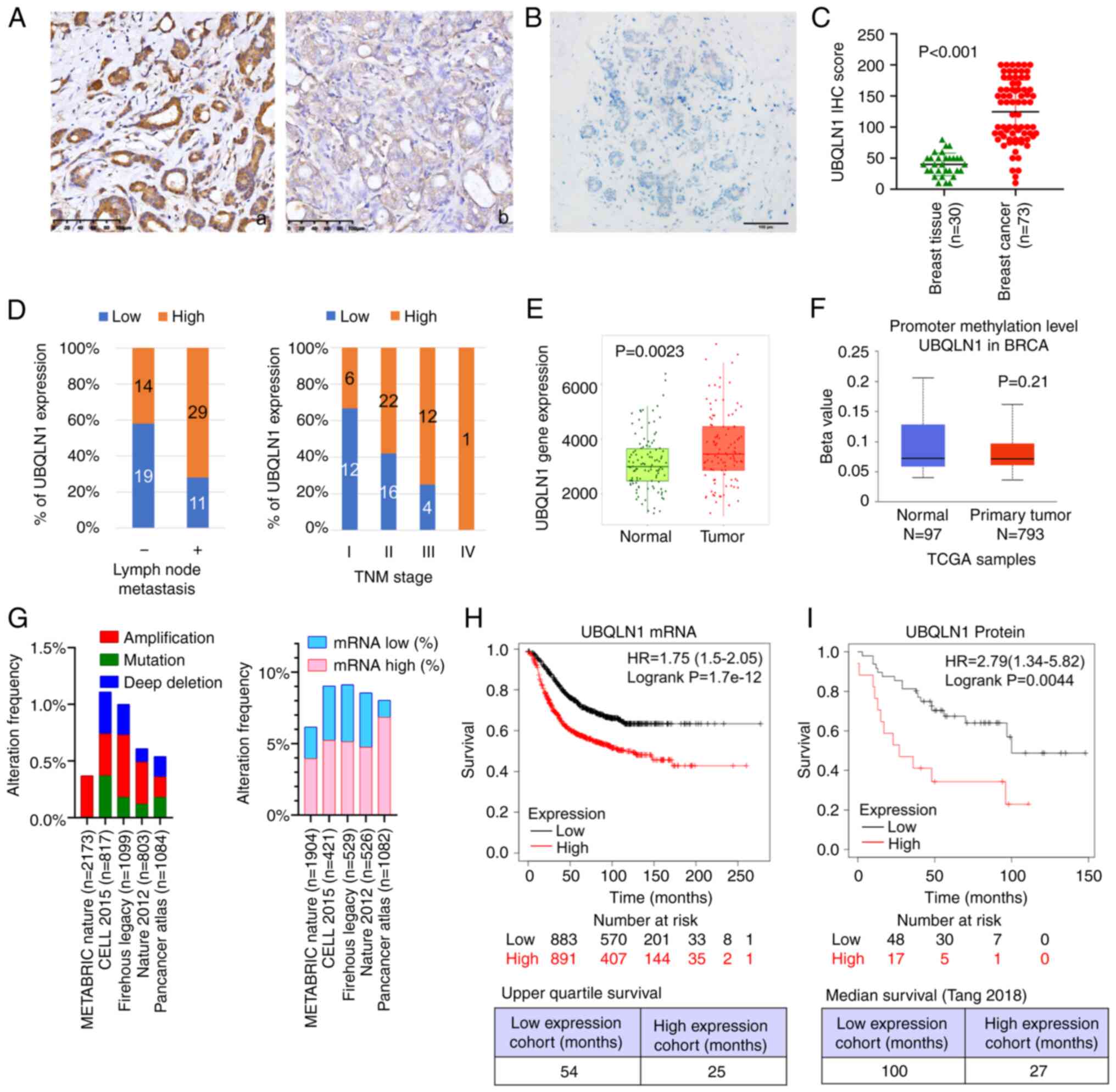
IHC. UBQLN1 immunoreactivity was mainly observed in the cytoplasm.
expression in breast cancer tissues was significantly higher than
normal breast tissue UBQLN1 expression (Fig. 1A-C).
The association between UBQLN1 expression and
clinicopathological features of patients with breast cancer was
further investigated. The results demonstrated that UBQLN1
expression was significantly associated with tumor diameter
(P=0.034), lymph node metastasis (P=0.016) and TNM staging
(P=0.047) (Fig. 1D and Table I); however, UBQLN1 expression was not
significantly associated with age, histology, differentiation,
molecular subtypes, or estrogen receptor, progesterone receptor,
HER2 or proliferating cell nuclear antigen expression (Table I). Breast cancer cases with a higher
UBQLN1 expression tended to have a more advanced stage and to
undergo lymphatic metastasis more frequently.
 | Table I.Association between UBQLN1 expression
and clinicopathological parameters of breast ductal carcinoma
cases. |
Table I.
Association between UBQLN1 expression
and clinicopathological parameters of breast ductal carcinoma
cases.
|
|
|
| UBQLN1 expression,
n (%) |
|
|---|
|
|
|
|
|
|
|---|
|
Characteristics | No. of cases | Low (n=30) | High (n=43) | P-value |
|---|
| Age | <50 | 32 | 14 (44) | 18 (56) | 0.811 |
|
| ≥50 | 41 | 16 (39) | 25 (61) |
|
| Diameter (cm) | <2 | 31 | 18 (58) | 15 (48) | 0.034a |
|
| ≥2 | 42 | 12 (29) | 38 (90) |
|
| LN | – | 33 | 19 (58) | 14 (42) | 0.016a |
|
| + | 40 | 11 (28) | 29 (73) |
|
| DM | – | 72 | 30 (42) | 42 (58) | 0.431 |
|
| + | 1 | 0 (0) | 1 (100) |
|
| TNM stage | I | 18 | 12 (67) | 6 (33) | 0.047a |
|
| II | 38 | 14 (42) | 24 (58) |
|
|
| III | 16 | 4 (25) | 12 (75) |
|
|
| IV | 1 | 0 (0) | 1 (100) |
|
| Grade | 1 | 12 | 6 (50) | 6 (50) | 0.388 |
|
| 2 | 48 | 17 (35) | 31 (65) |
|
|
| 3 | 13 | 7 (54) | 6 (46) |
|
| ER | – | 40 | 19 (48) | 21 (53) | 0.242 |
|
| + | 33 | 11 (33) | 22 (67) |
|
| PR | – | 53 | 22 (42) | 21 (40) | 0.061 |
|
| + | 20 | 8 (40) | 12 (60) |
|
| Her-2 | – | 61 | 24 (39) | 37 (61) | 0.534 |
|
| + | 12 | 6 (50) | 6 (50) |
|
| Molecular
subtype | Luminar A | 31 | 11 (35) | 20 (65) | 0.183 |
|
| Luminal B | 27 | 12 (44) | 15 (56) |
|
|
| HER2-enriched | 7 | 4 (57) | 3 (43) |
|
|
|
Triple-negative | 8 | 3 (37) | 5 (63) |
|
| PCNA | – | 28 | 12 (43) | 16 (57) | 0.812 |
|
| + | 45 | 18 (40) | 27 (60) |
|
To further explore the clinical significance of
UBQLN1 in breast cancer, a number of widely used databases were
employed, in order to clarify whether this difference was
associated with UBQLN1 promotor methylation. Using TNMPlot
(26), it was revealed that UBQLN1
mRNA expression was significantly higher in breast cancer tissues
than in paired normal breast tissues (Fig. 1E). Data from UALCAN indicated that
there was no significant difference in UBQLN1 methylation between
normal breast tissues and breast cancer tissues (Fig. 1F). The genomic alteration of
UBQLN1 was further investigated using cBioPortal. The
genetic alterations affecting UBQLN1 identified by cBioPortal in
five breast cancer studies are shown in Fig. 1G. Gene mutations and gene number
alterations were found in 39 (<0.1%) out of 5,976 patients with
breast cancer and the most frequent alteration was gene
amplification (22 cases) (Table SI).
Subsequently, mRNA expression in these breast cancer cases was
investigated in relation to mean mRNA expression in diploid samples
in breast cancer cases as follows: 223 (5.00%) out of 4,462 cases
demonstrated a higher expression, while 112 (2.51%) cases exhibited
a lower expression (Table SII).
UALCAN revealed that UBQLN1 mRNA expression was significantly
associated with tumor stage and lymph node metastasis (Table SIII).
Survival analysis using KmPlot revealed that, in
breast cancer, patients with a low UBQLN1 mRNA expression had a
longer median survival time (54 months) than patients with a high
UBQLN1 expression (25 months) (Fig.
1H) (27). The analysis of data
(n=118) from Tang et al (28)
revealed that UBQLN1 protein expression was also associated with a
poor prognosis (Fig. 1I). TIMER2 was
used with a Cox proportional hazard model to evaluate the
significance of UBQLN1 mRNA expression adjusted by clinical
factors, including age, sex, ethnicity and tumor stage in 976
patients with 136 dying. The results demonstrated that UBQLN1
[hazard ratio (HR)=1.33, P=0.038], age (HR=1.03, P<0.001), stage
3 (HR=3.28, P<0.001) and stage 4 (HR=14.17, P<0.001) were
significant predictors of survival (Table SIV). These data indicated a close
association between UBQLN1 and lymph node metastasis. and suggested
that UBQLN1 may be an independent prognostic predictor for overall
survival of patients with breast cancer.
UBQLN1 knockdown attenuates breast
cancer cell migration and invasion by inhibiting EMT
The results of the analysis of UBQLN1 in breast
cancer tissues suggested that it may facilitate the metastasis of
breast cancer. In the present study, the biological function of
UBQLN1 in breast cancer cell lines was then investigated using
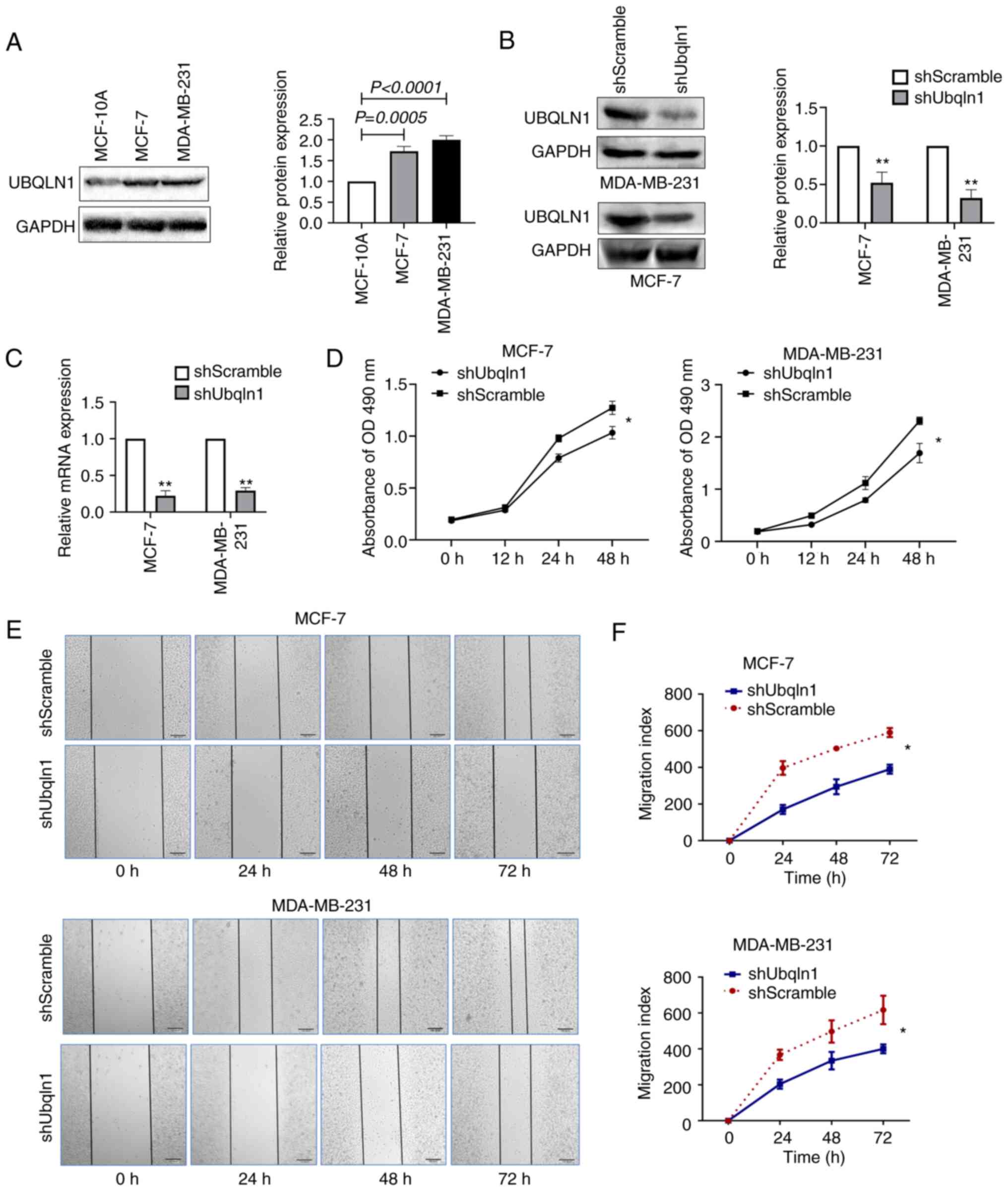
in vitro experiments. UBQLN1 expression in breast cancer
cells was first investigated using western blot analysis. Compared
with that in the human normal breast epithelial cell line, MCF-10A,
UBQLN1 expression was significantly higher in the breast cancer
cell lines, MCF-7 and MDA-MB-231 (Fig.
2A).
Subsequently, the effect of UBQLN1 silencing on the
biological behavior of breast cancer cell lines was investigated.
The expression of UBQLN1 in the MCF-7 and MDA-MB-231 cell lines was
stably knocked down using a lentivirus carrying UBQLN1 shRNA.
UBQLN1 protein and mRNA expression was significantly inhibited in
the MCF-7-shUbqln1 and MDA-MB-231-shUbqln1 cells compared with the
expression levels in the controls (MCF-7-shScramble and
MDA-MB-231-shScramble), as determined by western blot analysis
(Fig. 2B) and RT-qPCR (Fig. 2C).
MTT assay recealed that MCF-7 and MDA-MB-231 cell
viability was significantly inhibited when UBQLN1 was knocked down
(P<0.05) (Fig. 2D). The effect of
UBQLN1 knockdown on cell migration was investigated using wound
healing assay and Transwell migration assays. The results indicated
that UBQLN1 knockdown significantly suppressed cell migration
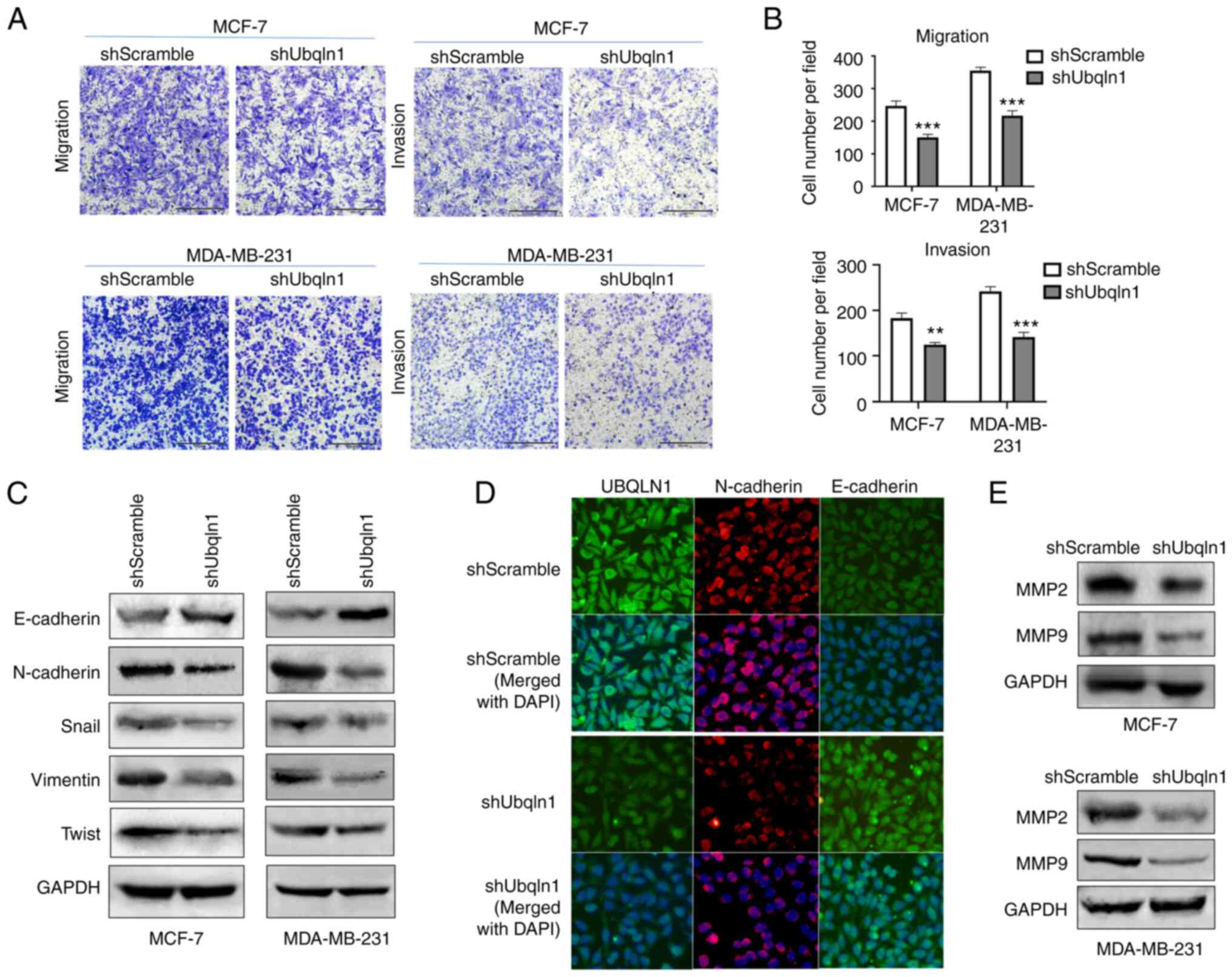
(Figs. 2E and F and 3A and B). The effect of UBQLN1 knockdown on
cell invasion was then investigated using Transwell invasion assay,
which uses Matrigel to simulate the extracellular matrix. The
results revealed that UBQLN1 knockdown significantly inhibited
breast cancer cell invasion, as indicated by the number of invaded
MCF-7-shUbqln1 and MDA-MB-231-shUbqln1 cells, which was decreased
in comparison with that in MCF-7-shScramble (P<0.01) and
MDA-MB-231-shScramble groups (P<0.001) (Fig. 3A and B).
Cancer cells acquire migratory and invasive
abilities by EMT, through which, epithelial cells acquire enhanced
mobility and invasive properties by losing cell-cell adhesion
structures and polarity (29). The
present study further investigated the effect of UBQLN1 knockdown
on the expression of EMT markers. In the cells in which UBQLN1 was
silenced, the expression of the epithelial marker, E-cadherin, was
increased, while the expression of the mesenchymal markers,
vimentin and N-cadherin, was decreased. The expression of Snail,
which is an E-cadherin repressor and a major EMT inducer, was
markedly downregulated in the cells in which UBQLN1 was silenced
(Fig. 3C). Similar results were
observed using immunofluorescence analysis for N-cadherin and
E-cadherin expression (Fig. 3D).
These findings indicated that UBQLN1 knockdown promoted
mesenchymal-epithelial transition (MET) and effectively inhibited
EMT in breast cancer.
MMP2 and MMP9, two important enzymes that degrade
the extracellular matrix, are associated with EMT and the
metastatic potential of cancer cells (30). In the present study, the effect of
UBQLN1 knockdown on the expression of MMP2 and MMP9 was then
investigated. Western blot analysis revealed that the expression of
MMP2 and MMP9 was markedly decreased in the cells in which UBQLN1
was silenced, suggesting that UBQLN1 may promote invasion via the
upregulation of MMP expression (Fig.
3E).
UBQLN1 promotes the stemness and
chemoresistance of breast cancer cells
It has been reported that BCSCs possess self-renewal
capabilities and contribute to tumor onset, recurrence, metastasis
and therapy resistance (31). The
present study investigated whether UBQLN1 is associated with the
stem cell properties of breast cancer. Firstly, it was investigated
whether then downregulation of UBQLN1 affects the expression of
BCSC markers. The results revealed that ALDH1, Oct-4 and Sox2
expression levels were markedly decreased in the breast cancer
cells in which UBQLN1 was silenced (P<0.001; Fig. 4A).
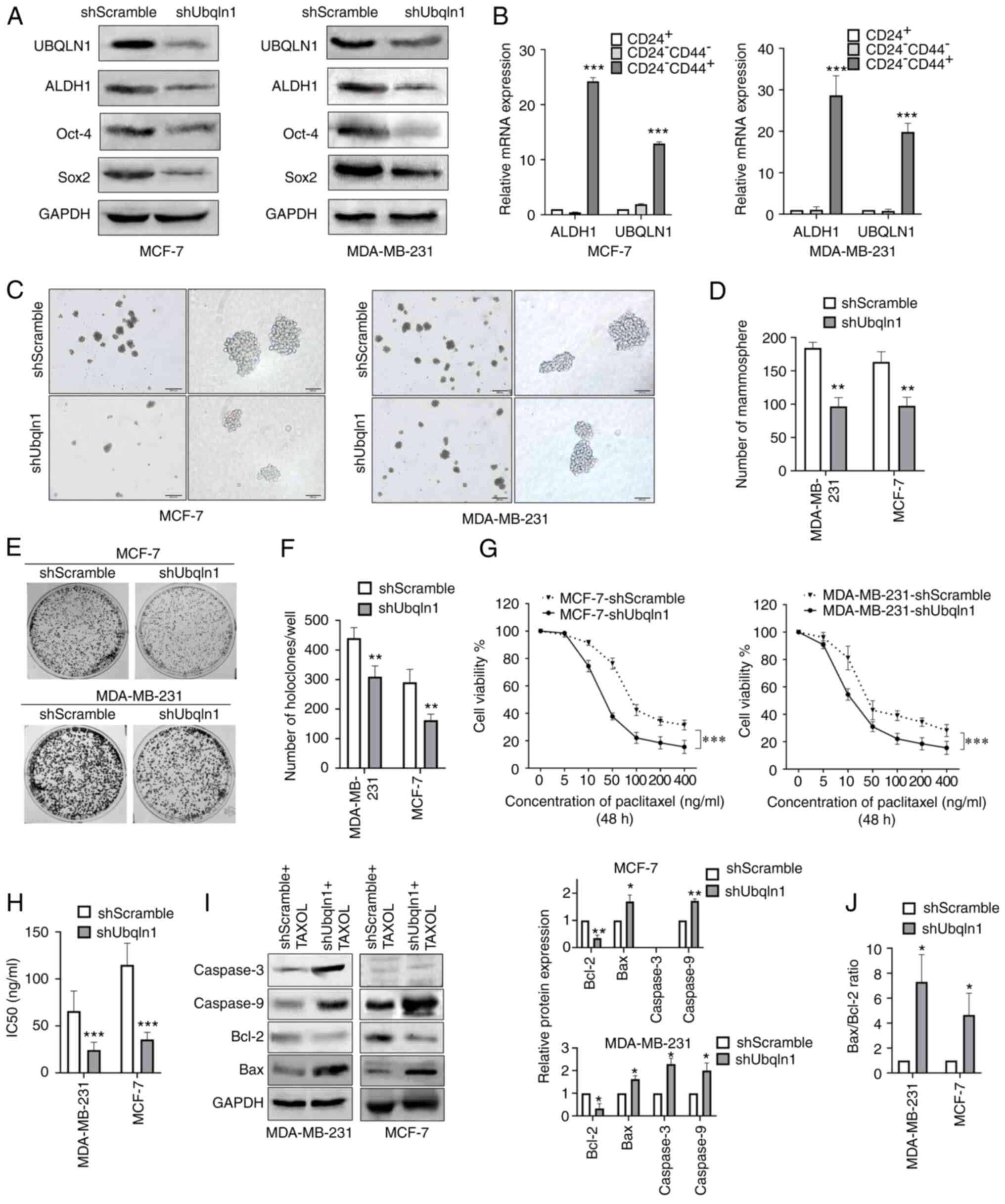 | Figure 4.UBQLN1 knockdown attenuates stemness
and chemoresistance to paclitaxel in breast cancer. MDA-MB-231 and
MCF-7 cells were stably transfected with UBQLN1 shRNA (shUbqln1) or
control shRNA (shScramble) and then subjected to different assays.
(A) Western blot analysis of UBQLN1, ALDH1, Oct-4 and Sox2. (B)
Reverse transcription-quantitative PCR analysis of UBQLN1 and the
stem cell marker, ALDH1, in breast cancer cells sorted using
magnetic-activated cell sorting system by CD24 or CD44 markers. (C)
Mammosphere formation assay and (D) quantification. (E) Holoclone
colony formation assay. (F) Histograms indicate mean holoclone
numbers formed by 1,000 starting cells. (G) MTT assay was performed
to examine cell viability after treating the cells with the
indicated dose of paclitaxel for 48 h. (H) Bar graph showing the
IC50 values. (I) Cells were exposed to 50 ng/ml
paclitaxel for 48 h, and the levels of Bcl-2, Bax, cleaved
caspase-3 and cleaved caspase-9 were then detected by western
blotting. Bar graphs illustrate relative protein expression levels.
(J) Ratio of Bax to Bcl-2. Data represent means ± SEM of three
independent experiments. *P<0.05, **P<0.01 and ***P<0.001.
UBQLN1, ubiquilin-1; shRNA, short hairpin RNA; TAXOL,
paclitaxel. |
CD24−/CD44+ cells and
ALDH1+ cells are widely considered to be BCSCs (24). Thus, in the present study, the
expression of UBQLN1 in BCSCs was determined. The
CD24−/CD44+ cell population from the
MDA-MB-231 and MCF-7 cell lines was isolated using MACS and
confirmed by their higher expression of the BCSC marker ALDH1. The
results of RT-qPCR demonstrated that UBQLN1 mRNA expression was
significantly higher in the CD24−/CD44+ cells
than in the CD24+ or CD24−/CD44−
cells (Fig. 4B).
Mammosphere formation reflects the self-renewal
potential of tumor cells (31). In
the present study, the results of mammosphere formation assays
indicated that UBQLN1 knockdown significantly decreased the number
of mammospheres (P<0.01; Fig. 4C and
D). Holoclone formation is a typical property of CSCs. The
number of holoclones in MCF-7-shUbqln1 and MDA-MB-231-shUbqln1
cells was significantly lower than that in the control groups
(P<0.01; Fig. 4E and F).
The effects of UBQLN1 knockdown on the sensitivity
of breast cancer cells to chemotherapy were then investigated in
the present study. Since paclitaxel is frequently used as the
first-line treatment drug in breast cancer (32), the cells were treated with various
concentrations of paclitaxel. MTT assay indicated that the
MCF7-shUbqln1 and MDA-MB-231-shUbqln1 cells were more sensitive to
paclitaxel than the controls (P<0.001; Fig. 4G). The drug concentrations that
inhibited cell proliferation by 50% (IC50) in the cells
in which UBQLN1 was knocked down were significantly lower than
those in the control cells (P<0.001; Fig. 4H). This result suggested that UBQLN1
knockdown promoted the cytotoxic effects of paclitaxel.
The results of western blot analysis demonstrated
that UBQLN1 knockdown promoted paclitaxel-induced apoptosis. This
was evidenced by the finding that cleaved caspase-3 and caspase-9
expression was significantly upregulated in the MDA-MB-231 cells in
which UBQLN1 was knocked down, and caspase-9 expression was
upregulated in the MCF-7 cells in which UBQLN was knocked down and
treated with paclitaxel for 48 h (P<0.01; Fig. 4I). Caspase-3 expression was not
detected in MCF-7 cells as these cells do not express caspase-3
(33).
The expression of Bcl-2, an anti-apoptotic member of
the Bcl-2 protein family, was significantly decreased, while that
of Bax, a pro-apoptotic effector protein, was increased in the
cells in which UBQLN1 was knocked down (Fig. 4I). As a result, the Bax/Bcl-2 ratio
significantly increased (P<0.05; Fig.
4J), thus indicating that UBQLN1 effectively inhibited
paclitaxel-induced apoptosis through Bcl-2 family members.
UBQLN1 sustains stemness and EMT by
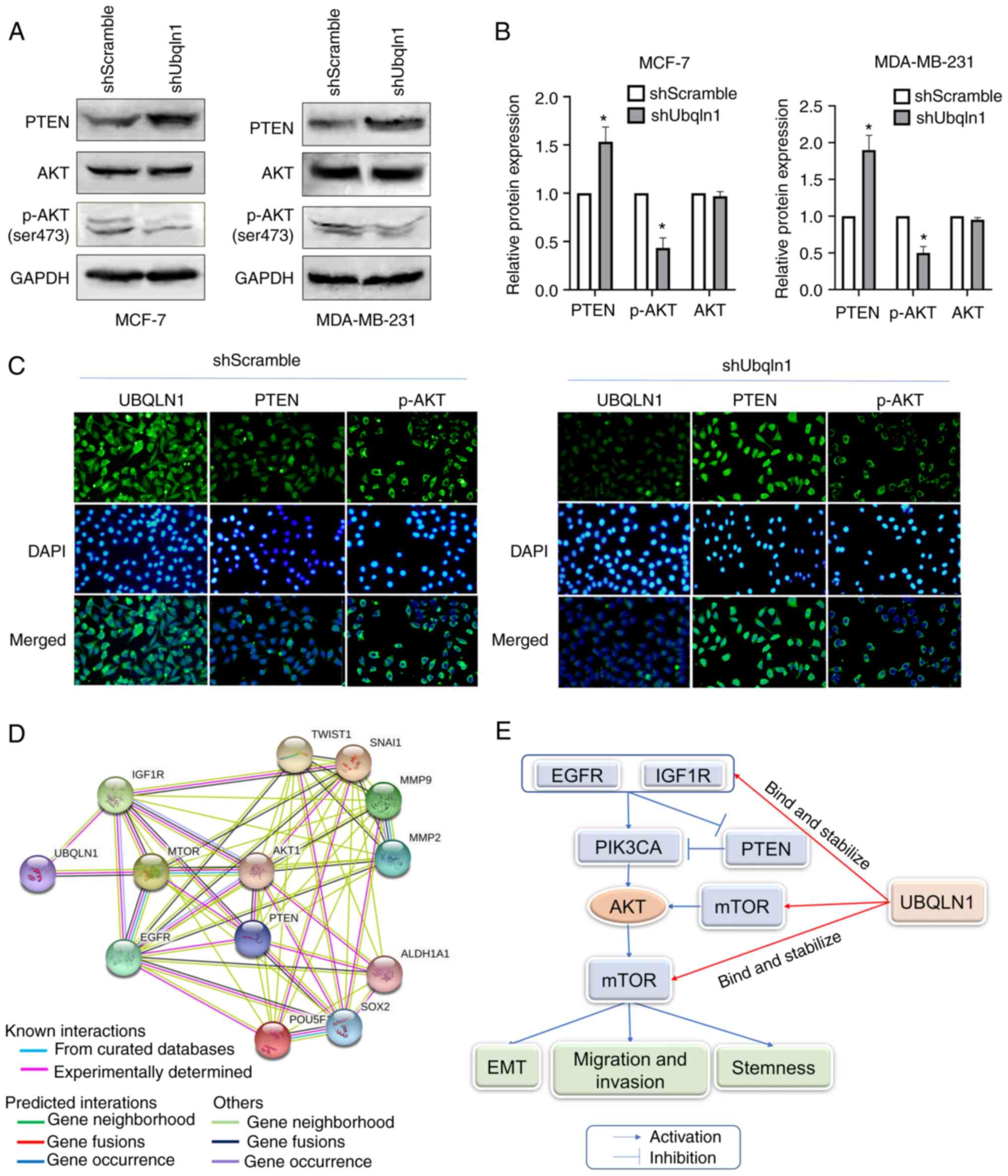
regulating PI3K/AKT signaling
UBQLN1 is important for the regulation of protein
degradation. Beverly et al (34) determined that UBQLN1 was associated
with specific biological functions or canonical biological pathways
by Ingenuity Pathway Analysis (IPA), and found that AKT signaling
is the most markedly canonical pathway represented by this gene.
The present study investigated the effect of UBQLN1 on the
expression of molecules involved in AKT signaling. AKT expression
was not altered, while p-AKT expression was significantly decreased
in the cells in which UBQLN1 was knocked down. PTEN, an inhibitor
of AKT signaling, was significantly increased in UBQLN1-silenced
cells (P<0.05; Fig. 5A and B).
Similar results were also observed by immunofluorescence analysis.
Following the loss of UBQLN1 in breast cancer, PTEN expression was
increased, while p-AKT expression was decreased in the MDA-MB-231
cells (Fig. 5C).
Furthermore, PPI network analysis was conducted
using STRING to analyze UBQLN1-related molecules, including EGFR,
insulin like growth factor 1 receptor (IGF1R), mTOR, AKT, PTEN;
stem cell markers, such as ALDH1, Oct-4 and Sox2; and EMT-related
molecules, such as Twist, Snail, MMP2 and MMP9, and to explore the
potential interactions among them. It was observed that these
molecules were all closely associated with AKT signaling (Fig. 5D). Therefore, UBQLN1 may inhibit
migration, invasion, EMT and the stemness of breast cancer by
affecting the activation of PI3K/AKT signaling (Fig. 5E).
Discussion
In the present study, UBQLN1 expression and function
in breast cancer tissues and cell lines was investigated. The
findings for breast cancer tissues using IHC, and the data from
UALCAN, KmPlot and TIMER2.0 demonstrated that UBQLN1 was highly
expressed in breast cancer tissues and breast cancer cell lines
both at the mRNA and protein level. UBQLN1 expression in breast
cancer was associated with TNM stage, lymph node metastasis and a
poor prognosis. These results suggested that UBQLN1 is associated
with metastasis and may be an independent prognostic predictor for
overall survival of patients with breast cancer. While the total
number of cases included in the IHC analysis in present study was
relatively small, including only 1 case with stage IV, further more
well-characterized largescale studies are required to validate the
association of UBQLN protein expression with the distant metastasis
of breast cancer. UBQLN1 expression has been estimated in several
cancer types, including non-small cell lung cancer (NSCLC)
(6,11,34–36), and
breast (20), gastric (12) and ovarian cancer (19). The majority of the findings in the
literature cited above are consistent with the findings of the
present study, suggesting that UBQLN1 promotes cancer progression
and may play an oncogenic role in cancer. In gastric and breast
cancer, UBQLN1 has been found to be highly expressed in cancer
tissues, and to be positively associated with TNM stage, tumor
invasion and lymph node metastasis. Higher UBQLN1 expression levels
have been shown to be associated with a shorter survival of
patients with gastric cancer and breast cancer (12,20). In
ovarian cancer, UBQLN1 has been found to be significantly
upregulated when exposed to cisplatin (19), indicating that UBQLN1 may mediate
cisplatin resistance in ovarian cancer. In lung cancer, both UBQLN1
mRNA (34,36) and protein (36) have been found to be highly expressed
in primary lung adenocarcinoma, and higher UBQLN1 levels have been
shown to be associated with the shorter survival of patients with
lung cancer (34). However, in NSCLC,
certain contradictory results have been reported. UBQLN1 has been
observed to be absent and underexpressed in ~50% of tumor tissues
(6). The silencing of UBQLN1 in a
NSCLC cell line has been shown to increase cell proliferation,
migration and invasion, actin cytoskeleton reorganization, and the
induction of EMT, leading to a more invasive cell phenotype
(6,11,35). These
data raise doubts concerning the biological role of UBQLN1 in
cancer, and further research is therefore warranted to clarify the
role of UBQLN1 in cancer.
To further elucidate the biological role of UBQLN1
in breast cancer, the influence of UBQLN1 knockdown on biological
behavior in vitro was investigated, in particular regarding
metastasis. In contrast to the tumor suppressor role reported in
lung cancer cells (6,11,35), the
present study found that the knockdown of UBQLN1 inhibited cell
migration and invasion, EMT, and MMP2 and MMP9 expression in breast
cancer cell lines. EMT has been implicated in carcinogenesis, and
confers metastatic properties to cancer cells by enhancing mobility
and invasion (37). MMP2 and MMP9
belong to a group of zinc-containing enzymes that are responsible
for the degradation of extracellular matrix components and play
pivotal roles in tumor growth and metastasis (30). The secretion and activation of MMPs is
a critical step of EMT (38). These
data suggested that UBQLN1 is associated with a more migratory and
invasive phenotype of breast cancer, and also that UBQLN1 knockdown
can effectively inhibit EMT.
It has been reported that BCSCs play a vital role in
metastasis and therapeutic resistance (37). The number of studies on the
association between UBQLN1 and stem cells is limited. In a study on
proteomic analysis of a proliferating and differentiating human
neuronal stem cell line, the results revealed a significantly
decreased UBLQN1 protein expression during stem cell
differentiation (21). RNA-sequencing
in human embryonic stem cells has demonstrated that UBQLN1 is
highly expressed in a cluster of cells with long telomeres and a
higher expression of known pluripotency markers (22). These studies indicated an increased
level of UBQLN1 expression in stem cells. In the present study, a
higher expression of UBQLN1 mRNA in BCSCs was detected, in
comparison with that of non-BCSCs. Additionally, UBQLN1 silencing
in breast cancer significantly downregulated the expression of the
stem cell markers, ALDH1, Oct-4 and Sox2, decreased the mammosphere
formation ability and increased the sensitivity of breast cancer
cells to paclitaxel therapy. These data thus suggest that UBQLN1 is
highly expressed in BCSCs and maintains the stemness properties of
breast cancer, including self-renewal and chemoresistance.
The molecular mechanisms underlying the functions of
UBQLN1 in the metastasis and stemness of breast cancer are unknown.
UBQLN1 was first identified as one of the DAN-binding proteins
(DA41) that was expressed at low levels in quiescent cells, and was
significantly increased between the G1 and S phases of
the cell cycle (39). It was also
identified as a protein linking IAP with the cytoskeleton (PLIC),
which mediates the interaction between integrin-associated protein
(IAP) and vimentin-containing intermediate filaments (40). The overexpression of PLICs has been
shown to increase IAP-dependent cell spreading and increase the
vimentin association with IAP at the plasma membrane (40). UBQLNs have been found to be
functionally linked to the UPS and act as ubiquitin receptors
(41). UBQLN1 is a component of the
protein quality control system, and is important for protein
degradation and stabilization (42).
UBQLN1 facilitates the proteasome-mediated degradation of certain
proteins, including ataxin 3 (14,43),
epidermal growth factor receptor pathway substrate 15 (14,43,44),
Homo sapiens J domain protein 1a (43) and the viral polymerase nonstructural
protein 5B (45). For certain
ubiquitin-dependent proteasome substrates such as IκBα and P53,
overexpression of UBQLN1 interferes with degradation (46). UBQLN1 binds and stabilizes presenilin
1/2 (47), γ-aminobutyric acid type A
(48), BCLb (34), EGFR (11), IGF1R (18), mTOR (35,49) and
extended synaptotagmin 2 (14). The
mechanism determining the fate of UBQLN1 interacting partners
remains unknown.
Among these proteins, EGFR, IGF1R and mTOR are all
closely related to AKT signaling, which has been widely reported to
participate in tumorigenesis (Fig.
5D) (9), particularly in
metastasis and stemness (7,10). EGFR and IGF1R function as oncogenes
and promote the development and progression of numerous cancer
types (50,51). PI3K/AKT signaling is one of the most
critical cancer-promoting pathways through the upregulation of
tyrosine kinase receptors (52).
Kurlawala et al (11,18) reported that the loss of UBQLN1 led to
a marked decrease in total EGFR and IGF1R, particularly when
stimulated with their ligands, and the interactions with UBQLN1
stabilized these receptors without affecting the activation of
these molecules. However, the effect of this interaction on the
activation of downstream signaling was not detected. UBQLN1 can
also interact with another molecule involved in PI3K/AKT signaling,
namely mTOR (35,49). It has been reported that the loss of
UBQLN1 markedly inhibits mTOR phosphorylation and activates
autophagy activity (35). Therefore,
UBQLN1 may be crucial for maintaining the stability of EGFR, IGF1R
and mTOR, thus influencing the activation of PI3K-AKT signaling
(Fig. 5E). The UBQLN1-related
signaling pathway was estimated by IPA, and the results showed that
the canonical pathways of UBQLN1 were involved in the process of
neoplastic transformation, including AKT signaling, MYC signaling
and cell cycle regulation (34).
Although previous evidence has suggested that UBQLN1 may affect the
activation of AKT signaling (34,35,49),
whether and how UBQLN1 modulates AKT signaling in cancer remains
unclear. The present study revealed that UBQLN1 knockdown
significantly decreased the expression of p-AKT without affecting
total AKT protein levels. In addition, PTEN, a plasma-membrane
lipid phosphatase antagonizing the PI3K/AKT pathway (53), demonstrated a significantly increased
expression in UBQLN1-depleted breast cancer cells. PPI network
analysis by STRING indicated that UBQLN1 and molecules associated
with EMT and stemness are all closely correlated with AKT
signaling. The present results and those of previous studies
suggest that UBQLN1 promotes the migration, invasion, EMT and the
stemness of breast cancer by maintaining the activation of AKT
signaling, possibly by stabilizing EGFR, IGF1R and mTOR (Fig. 5D and E).
In conclusion, the present study revealed that
UBQLN1 expression was upregulated in breast cancer tissues and was
associated with a poor prognosis. UBQLN1 knockdown inhibited the
migration, invasion, EMT and the stemness of breast cancer cells,
and attenuated AKT signaling activation. It was also suggested that
UBQLN1 facilitated tumor progression by maintaining the activation
of AKT signaling. However, further studies are required to validate
the current findings on UBQLN1 function in breast cancer
progression by gain-of-function studies and to investigate the
specific molecular mechanism of this protein degradation-related
protein in the regulation of AKT signaling activation.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Dalian
Science and Technology Innovation Fund (grant no. 2019J12SN53).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
LL, BS and XY conceived and designed the
experiments. XF, TQ, AC, SF and BW performed the experiments. XF,
QZ and XY performed the data collection and analyses. XY and XF
wrote the original draft. XF, XY and LL confirm the authenticity of
the all raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study and experimental procedures were
approved by the Ethics Committee of Dalian Medical University
[Dalian, China; (IRB approval no. 2021006)]. Written informed
consent was obtained from all patients or patients' families. The
study was conducted according to the principles outlined in the
Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
UBQLN1
|
ubiquilin-1
|
|
CSCs
|
cancer stem cells
|
|
BCSCs
|
breast cancer stem cells
|
|
EMT
|
epithelial-to-mesenchymal
transition
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
MACS
|
magnetic-activated cell sorting
|
|
IGF1R
|
insulin like growth factor 1
receptor
|
|
PLIC
|
protein linking IAP with
cytoskeleton
|
|
IAP
|
integrin-associated protein
|
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality Worldwide for 36
cancers in 185 Countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thorat MA and Balasubramanian R: Breast
cancer prevention in high-risk women. Best Pract Res Clin Obstet
Gynaecol. 65:18–31. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nandy SB and Lakshmanaswamy R: Cancer stem
cells and metastasis. Prog Mol Biol Transl Sci. 151:137–176. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Najafi M, Farhood B and Mortezaee K:
Cancer stem cells (CSCs) in cancer progression and therapy. J Cell
Physiol. 234:8381–8395. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu X, Zhang F, Mao J, Lu Y, Li J, Ma W,
Fan S, Zhang C, Li Q, Wang B, et al: Protein tyrosine phosphatase
receptor-type δ acts as a negative regulator suppressing breast
cancer. Oncotarget. 8:98798–98811. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shah PP, Lockwood WW, Saurabh K, Kurlawala
Z, Shannon SP, Waigel S, Zacharias W and Beverly LJ: Ubiquilin1
represses migration and epithelial-to-mesenchymal transition of
human non-small cell lung cancer cells. Oncogene. 34:1709–1717.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Karimi Roshan M, Soltani A, Soleimani A,
Rezaie Kahkhaie K, Afshari AR and Soukhtanloo M: Role of AKT and
mTOR signaling pathways in the induction of epithelial-mesenchymal
transition (EMT) process. Biochimie. 165:229–234. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tao C, Luo J, Tang J, Zhou D, Feng S, Qiu
Z, Putti TC, Xiang T, Tao Q, Li L and Ren G: The tumor suppressor
Zinc finger protein 471 suppresses breast cancer growth and
metastasis through inhibiting AKT and Wnt/β-catenin signaling. Clin
Epigenetics. 12:1732020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xia P and Xu XY: PI3K/Akt/mTOR signaling
pathway in cancer stem cells: From basic research to clinical
application. Am J Cancer Res. 5:1602–1609. 2015.PubMed/NCBI
|
|
11
|
Kurlawala Z, Saurabh K, Dunaway R, Shah
PP, Siskind LJ and Beverly LJ: Ubiquilin proteins regulate EGFR
levels and activity in lung adenocarcinoma cells. J Cell Biochem.
122:43–52. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bao J, Jiang X, Zhu X, Dai G, Dou R, Liu
X, Sheng H, Liang Z and Yu H: Clinical significance of ubiquilin 1
in gastric cancer. Medicine(Baltimore). 97:e97012018.PubMed/NCBI
|
|
13
|
Jantrapirom S, Piccolo LL, Pruksakorn D,
Potikanond S and Nimlamool W: Ubiquilin networking in cancers.
Cancers (Basel). 12:15862020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kurlawala Z, Shah PP, Shah C and Beverly
LJ: The STI and UBA Domains of UBQLN1 are critical determinants of
substrate interaction and proteostasis. J Cell Biochem.
118:2261–2270. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X, Zhou J, Chen H, Wang F, Mei Q and
Sun H: The association between the UBQLN1 polymorphism and
Alzheimer's disease risk: A systematic review. Cell Mol Biol
(Noisy-le-grand). 63:94–96. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim SH, Shi Y, Hanson KA, Williams LM,
Sakasai R, Bowler MJ and Tibbetts RS: Potentiation of amyotrophic
lateral sclerosis (ALS)-associated TDP-43 aggregation by the
proteasome-targeting factor, ubiquilin 1. J Biol Chem.
284:8083–8092. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Safren N, El Ayadi A, Chang L, Terrillion
CE, Gould TD, Boehning DF and Monteiro MJ: Ubiquilin-1
overexpression increases the lifespan and delays accumulation of
Huntingtin aggregates in the R6/2 mouse model of Huntington's
disease. PLoS One. 9:e875132014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kurlawala Z, Dunaway R, Shah PP, Gosney
JA, Siskind LJ, Ceresa BP and Beverly LJ: Regulation of
insulin-like growth factor receptors by Ubiquilin1. Biochem J.
474:4105–4118. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guidi F, Puglia M, Gabbiani C, Landini I,
Gamberi T, Fregona D, Cinellu MA, Nobili S, Mini E, Bini L, et al:
2D-DIGE analysis of ovarian cancer cell responses to cytotoxic gold
compounds. Mol Biosyst. 8:985–993. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Lu J, Zhao X, Feng Y, Lv S, Mu Y,
Wang D, Fu H, Chen Y and Li Y: Prognostic significance of
Ubiquilin1 expression in invasive breast cancer. Cancer Biomark.
15:635–643. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hoffrogge R, Mikkat S, Scharf C, Beyer S,
Christoph H, Pahnke J, Mix E, Berth M, Uhrmacher A, Zubrzycki IZ,
et al: 2-DE proteome analysis of a proliferating and
differentiating human neuronal stem cell line (ReNcell VM).
Proteomics. 6:1833–1847. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang H, Zhang K, Liu Y, Fu Y, Gao S, Gong
P, Wang H, Zhou Z, Zeng M, Wu Z, et al: Telomere heterogeneity
linked to metabolism and pluripotency state revealed by
simultaneous analysis of telomere length and RNA-seq in the same
human embryonic stem cell. BMC Biol. 15:1142017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang F, Wang B, Qin T, Wang L, Zhang Q,
Lu Y, Song B, Yu X and Li L: IL-6 induces tumor suppressor protein
tyrosine phosphatase receptor type D by inhibiting miR-34a to
prevent IL-6 signaling overactivation. Mol Cell Biochem. 473:1–13.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qin T, Li B, Feng X, Fan S, Liu L, Liu D,
Mao J, Lu Y, Yang J, Yu X, et al: Abnormally elevated USP37
expression in breast cancer stem cells regulates stemness,
epithelial-mesenchymal transition and cisplatin sensitivity. J Exp
Clin Cancer Res. 37:2872018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bartha Á and Győrffy B: TNMplot.com: A web
tool for the comparison of gene expression in normal, tumor and
metastatic tissues. Int J Mol Sci. 22:26222021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Györffy B, Lanczky A, Eklund AC, Denkert
C, Budczies J, Li Q and Szallasi Z: An online survival analysis
tool to rapidly assess the effect of 22,277 genes on breast cancer
prognosis using microarray data of 1,809 patients. Breast Cancer
Res Treat. 123:725–731. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tang W, Zhou M, Dorsey TH, Prieto DA, Wang
XW, Ruppin E, Veenstra TD and Ambs S: Integrated
proteotranscriptomics of breast cancer reveals globally increased
protein-mRNA concordance associated with subtypes and survival.
Genome Med. 10:942018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jia D, Park JH, Kaur H, Jung KH, Yang S,
Tripathi S, Galbraith M, Deng Y, Jolly MK, Kaipparettu BA, et al:
Towards decoding the coupled decision-making of metabolism and
epithelial-to-mesenchymal transition in cancer. Br J Cancer.
124:1902–1911. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fu Y, Shao ZM, He QZ, Jiang BQ, Wu Y and
Zhuang ZG: Hsa-miR-206 represses the proliferation and invasion of
breast cancer cells by targeting Cx43. Eur Rev Med Pharmacol Sci.
19:2091–2104. 2015.PubMed/NCBI
|
|
31
|
Yu JM, Sun W, Wang ZH, Liang X, Hua F, Li
K, Lv XX, Zhang XW, Liu YY, Yu JJ, et al: TRIB3 supports breast
cancer stemness by suppressing FOXO1 degradation and enhancing SOX2
transcription. Nat Commun. 10:57202019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Abu Samaan TM, Samec M, Liskova A, Kubatka
P and Büsselberg D: Paclitaxel's mechanistic and clinical effects
on breast cancer. Biomolecules. 9:7892019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jänicke RU: MCF-7 breast carcinoma cells
do not express caspase-3. Breast Cancer Res Treat. 117:219–221.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Beverly LJ, Lockwood WW, Shah PP,
Erdjument-Bromage H and Varmus H: Ubiquitination, localization, and
stability of an anti-apoptotic BCL2-like protein, BCL2L10/BCLb, are
regulated by Ubiquilin1. Proc Natl Acad Sci USA. 109:E119–E126.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang X, Su Y, Lin H and Yao X: The
impacts of ubiquilin 1 (UBQLN1) knockdown on cells viability,
proliferation, and apoptosis are mediated by p53 in A549 lung
cancer cells. J Thorac Dis. 12:5887–5895. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen G, Wang X, Yu J, Varambally S, Yu J,
Thomas DG, Lin MY, Vishnu P, Wang Z, Wang R, et al: Autoantibody
profiles reveal ubiquilin 1 as a humoral immune response target in
lung adenocarcinoma. Cancer Res. 67:3461–3467. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mittal V: Epithelial mesenchymal
transition in tumor metastasis. Annu Rev Pathol. 13:395–412. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Agraval H and Yadav UCS: MMP-2 and MMP-9
mediate cigarette smoke extract-induced epithelial-mesenchymal
transition in airway epithelial cells via EGFR/Akt/GSK3β/β-catenin
pathway: Amelioration by fisetin. Chem Biol Interact.
314:1088462019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hanaoka E, Ozaki T, Ohira M, Nakamura Y,
Suzuki M, Takahashi E, Moriya H, Nakagawara A and Sakiyama S:
Molecular cloning and expression analysis of the human DA41 gene
and its mapping to chromosome 9q21.2-q21.3. J Hum Genet.
45:188–191. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu AL, Wang J, Zheleznyak A and Brown EJ:
Ubiquitin-related proteins regulate interaction of vimentin
intermediate filaments with the plasma membrane. Mol Cell.
4:619–625. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gadhave K, Kumar P, Kapuganti SK, Uversky
VN and Giri R: Unstructured biology of proteins from
ubiquitin-proteasome system: Roles in cancer and neurodegenerative
diseases. Biomolecules. 10:7962020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang C and Saunders AJ: An emerging role
for Ubiquilin 1 in regulating protein quality control system and in
disease pathogenesis. Discov Med. 8:18–22. 2009.PubMed/NCBI
|
|
43
|
Heir R, Ablasou C, Dumontier E, Elliott M,
Fagotto-Kaufmann C and Bedford FK: The UBL domain of PLIC-1
regulates aggresome formation. EMBO Rep. 7:1252–1258. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Regan-Klapisz E, Sorokina I, Voortman J,
de Keizer P, Roovers RC, Verheesen P, Urbé S, Fallon L, Fon EA,
Verkleij A, et al: Ubiquilin recruits Eps15 into ubiquitin-rich
cytoplasmic aggregates via a UIM-UBL interaction. J Cell Sci.
118((Pt 19)): 4437–4450. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gao L, Tu H, Shi ST, Lee KJ, Asanaka M,
Hwang SB and Lai MM: Interaction with a ubiquitin-like protein
enhances the ubiquitination and degradation of hepatitis C virus
RNA-dependent RNA polymerase. J Virol. 77:4149–4159. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kleijnen MF, Shih AH, Zhou P, Kumar S,
Soccio RE, Kedersha NL, Gill G and Howley PM: The hPLIC proteins
may provide a link between the ubiquitination machinery and the
proteasome. Mol Cell. 6:409–419. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mah AL, Perry G, Smith MA and Monteiro MJ:
Identification of ubiquilin, a novel presenilin interactor that
increases presenilin protein accumulation. J Cell Biol.
151:847–862. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bedford FK, Kittler JT, Muller E, Thomas
P, Uren JM, Merlo D, Wisden W, Triller A, Smart TG and Moss SJ:
GABA(A) receptor cell surface number and subunit stability are
regulated by the ubiquitin-like protein Plic-1. Nat Neurosci.
4:908–916. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu S, Mikhailov A, Kallo-Hosein H, Hara K,
Yonezawa K and Avruch J: Characterization of ubiquilin 1, an
mTOR-interacting protein. Biochim Biophys Acta. 1542:41–56. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cao J and Yee D: Disrupting insulin and
IGF receptor function in cancer. Int J Mol Sci. 22:5552021.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Santos ED, Nogueira KA, Fernandes LC,
Martins JR, Reis AV, Neto JB, Júnior IJ, Pessoa C, Petrilli R and
Eloy JO: EGFR targeting for cancer therapy: Pharmacology and
immunoconjugates with drugs and nanoparticles. Int J Pharm.
592:1200822021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gallardo A, Lerma E, Escuin D, Tibau A,
Muñoz J, Ojeda B, Barnadas A, Adrover E, Sánchez-Tejada L, Giner D,
et al: Increased signalling of EGFR and IGF1R, and deregulation of
PTEN/PI3K/Akt pathway are related with trastuzumab resistance in
HER2 breast carcinomas. Br J Cancer. 106:1367–1373. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Trotman LC, Wang X, Alimonti A, Chen Z,
Teruya-Feldstein J, Yang H, Pavletich NP, Carver BS, Cordon-Cardo
C, Erdjument-Bromage H, et al: Ubiquitination regulates PTEN
nuclear import and tumor suppression. Cell. 128:141–156. 2007.
View Article : Google Scholar : PubMed/NCBI
|