Introduction
Esophageal cancer (EC) is the eighth commonest
malignancy and sixth leading cause of cancer-related mortalities
worldwide (1). In Eastern countries,
esophageal squamous cell carcinoma (ESCC) is the commonest
histological type of EC and one of the most aggressive malignancies
(2). Despite the development of
multidisciplinary treatments, the prognosis of patients with EC
remains poor owing to its high invasiveness and metastatic
potential (3). This limited
improvement in the prognosis of patients with EC has prompted the
search for novel candidates that can act as clinically useful
biomarkers and treatment targets.
Anterior gradient 2 (AGR2), the human homolog of the
Xenopus laevis cement gland gene (XAG-2), is a member of the
protein disulfide isomerase family and usually localizes to the
endoplasmic reticulum (ER) (4,5). Although it primarily functions as a
modulator of ER homeostasis, it has been reported to have a number
of functional roles, such as in cell proliferation, migration and
differentiation, in some types of human cancer (6). Among human cancers, the overexpression of
AGR2 was first reported in breast cancer (7) and similar results were subsequently
reported in several human adenocarcinomas, including prostate,
pancreas, ovary and lung carcinomas (8–11). A recent
review reported that the overexpression of AGR2 has an unfavorable
impact on overall survival in breast, prostate and gastric cancers
(12). AGR2 is also found to be highly
expressed in head and neck squamous cell carcinoma (HNSCC)
(13–15)
and its downregulation in HNSCC cells decreases cell proliferation
and induces apoptosis (15,16). In addition, in esophageal
adenocarcinoma, AGR2 has been reported to promote tumor growth and
cell migration (17).
AGR2 suppresses p53 phosphorylation and worsens
prognosis in estrogen receptor-positive TP53-wild-type
breast cancer (18). In addition,
upregulated AGR2 expression in Barrett's epithelium and
precancerous lesions suppresses the phosphorylation of p53
(19). These studies show that AGR2
suppresses p53-related pathways via the inhibition of p53
phosphorylation, which is the main step in p53 activation (20). It has been reported that p53 serves
multiple roles in a number of cellular processes, such as cell
cycle regulation, apoptosis induction, differentiation and
senescence (21,22) and a major role of p53 is the
maintenance of genomic stability and integrity (23). The phosphorylation of the p53 protein
is related to malignancy and poor prognosis in some types of cancer
(4,6,18,19). p53 serine 15 is generally
phosphorylated in response to DNA damage and this phosphorylation
activates p53 (20). Therefore, the
inactivation of p53 via the inhibition of phosphorylation
represents a critical step in preventing tumor development and
progression.
To the best of the authors' knowledge, no
comprehensive functional analyses on the role of AGR2 in ESCC have
been conducted, although several studies have focused on AGR2 in
other cancers. The present study examined the effects of AGR2
downregulation on tumor progression and the phosphorylation of p53
in ESCC cell lines. In addition, the associations between AGR2 and
p53 expression, as well as clinical outcomes, were analyzed in ESCC
tissue.
Materials and methods
Bioinformatics analysis
Gene Expression Profiling Interactive Analysis
(GEPIA, http://gepia.cancer-pku.cn/index.html) was used to
show the expression of AGR2, which was the target gene, in
esophageal cancer.
Cell lines and culture
Human ESCC cell lines TE2, TE5, TE8, TE11 and TE15
and human osteosarcoma cell lines U2OS and SAOS2 were obtained from
the RIKEN BioResource Center Cell Bank. Human ESCC cell lines
KYSE70, KYSE150 and KYSE170 were obtained from the Japanese
Collection of Research Bioresources Cell Bank. The immortalized
esophageal epithelium cell line Het-1A and mesothelial cell line
MeT-5A were obtained from the American Type Culture Collection. The
ESCC cell lines Het-1A and MeT-5A were maintained in Roswell Park
Memorial Institute medium (Nakalai Tisque) and U2OS and SaOS2 cells
were cultured in HyClone McCoy's 5A medium (Cytiva). The two media
were supplemented with 10% fetal bovine serum (System Biosciences),
100 U/ml penicillin and 100 µg/ml streptomycin. All cell lines were
cultured in a humidified 37°C incubator with 5% carbon dioxide.
RNA extraction and quantification of
mRNA expression
Total RNA was extracted from the cell lines using a
miRNeasy Mini kit (Qiagen) as per the manufacturer's instructions.
Reverse transcription reactions were performed using a High
Capacity cDNA Reverse Transcription kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.), as per the manufacturer's given
procedure, at 25°C for 10 min, followed by 37°C for 120 min and
85°C for 5 min. The level of mRNA expression was measured using
TaqMan Gene Expression Assays (Hs_00356521_m1 for AGR2 and
Hs_02758991_g1 for GAPDH; Applied Biosystems; Thermo Fisher
Scientific, Inc.), as per the manufacturer's protocol. Reverse
transcription-quantitative (RT-q) PCR was performed using a
StepOnePlus PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.) and the cycle threshold (Ct) value was calculated
using StepOne Software v2.3 (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were: 95°C for 10
min, followed by 40 cycles at 95°C for 15 sec, 55°C for 30 sec and
72°C for 30 sec. The results were examined using the
2−ΔΔCq method relative to the expression of GAPDH
(24).
Western blot analysis
The cultured cells were lysed using the M-PER
Mammalian Protein Extraction Reagent (Thermo Fisher Scientific,
Inc.) and protein was extracted. The protein concentration was
adjusted using the Protein Assay Rapid kit Wako II (FUJIFILM Wako
Pure Chemical Corporation). The prepared lysate containing 20 µg of
total protein was subjected to 12 or 15% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, followed by transfer to
a polyvinylidene difluoride membrane (Cytiva). After blocking by 5%
skimmed milk powder or 5% bovine serum albumin (Sigma-Aldrich,
Merck KGaA) at room temperature for 1 h, the membrane was
subsequently probed with each candidate antibody at 4°C overnight.
Next day, the secondary antibody was added into the membranes at
room temperature for 1 h and the level of protein expression in
each case was visualized via SuperSignal West Dura (Thermo Fisher
Scientific, Inc.) and evaluated using an Amersham Imager 680,
software version 2.0.0 (Cytiva). β-actin (ACTB) was used as a
loading control.
The antibodies were purchased and diluted as
follows: anti-AGR2 antibody (rabbit, 1:5,000; cat. no. Ab76473)
from Abcam; anti-phospho-p53 antibody (serine 15) (mouse, 1:1,000;
cat. no. 9286S) from Cell Signaling Technology, Inc.; anti-p53
antibody (DO1, Mouse, 1:1,000; cat. no. sc-126) from Santa Cruz
Biotechnology, Inc. and anti-ACTB antibody (Mouse, 1:20,000; cat.
no. A5441) from Sigma-Aldrich. The secondary antibodies were:
Anti-mouse IgG, HRP-linked anti body (horse; cat. no. 7076) and
anti-rabbit IgG, HRP-linked anti body (goat; cat. no. 7074) both
from Cell Signaling Technology, Inc. The dilution rate of the
secondary antibody varied according to the dilution rate of the
primary antibody.
Downregulation of AGR2 by small
interfering RNA (siRNA)
The expression of AGR2 was downregulated
using small interfering RNA (siRNA) targeting AGR2 (Stealth RNAi™
siRNA, Thermo Fisher Scientific, Inc.; cat. no. HSS116220;
UUUCUUUAAAGCUUGACUGUGUGGG). ESCC and osteosarcoma cells were
transfected either with control siRNA (Stealth RNAi™ siRNA Negative
Control, Thermo Fisher Scientific; cat. no. 12935112) or siRNA
targeting AGR2 at 10 nmol/l in a six-well culture plate using
Lipofectamine® RNAiMAX (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. After transfection
for 72 h at 37°C, AGR2 mRNA and protein expression was confirmed
using RT-qPCR and western blotting, respectively.
Proliferation assay
Cells were seeded on 96-well plates, incubated for
24 h and transfected with either control or AGR2-targeting siRNA.
After transfection, cells were incubated for 24 h at 37°C, and the
number of viable cells 0, 24, 48, 72 and 96 h after treatment was
determined using absorbance data at a wavelength of 450 nm using
Cell Count Reagent SF (Nakalai Tesque).
Colony formation assay
TE2, TE5 and TE15 cells transfected with either
control or AGR2-targeting siRNA were seeded in six-well culture
plates at a density of 1,000 cells per well. After 2 weeks, the
cells were fixed with 4% paraformaldehyde at room temperature for
10 min and briefly stained with crystal violet (Nakalai Tesque) at
room temperature until all cells were stained enough. The areas of
all stained cells were measured using the NIH ImageJ System (v1.53)
(http://rsb.info.nih.gov/ij/).
Cell cycle assay
Cell cycles were analyzed 72 h after transfection
using flow cytometry. The cells were detached from the plate using
trypsin-EDTA, treated with 0.2% Triton X-100 and stained with PI
RNase staining buffer (Becton-Dickinson and Company). The cells
were analyzed using a BD Accuri C6 (BD Biosciences) and cell cycle
distribution was recorded using the BD Accuri Software (BD
Biosciences). At least 10,000 cells were assessed for each
measurement.
Apoptosis assay
The cells were stained and evaluated using an
Annexin V-FITC kit (Beckman Coulter, Inc.) 72 h after transfection
according to the manufacturer's instructions. The proportions of
early and late apoptotic cells were measured by flow cytometry
using the BD Accuri C6. At least 10,000 cells were assessed for
each measurement.
Patients and tissue samples
A total of 81 patients with ESCC who had undergone
esophagectomy between 1999 and 2013 at the Kyoto Prefectural
University of Medicine were enrolled in the present study. Written
informed consent for the research was obtained from all patients
prior to their respective treatments. Patients who had undergone
non-curative resection or preoperative chemo- or radiation-therapy
were excluded from the present study. The median length of the
follow-up period for the censored cases was 5.98 years (range,
0.38–16.7 years). A total of 35 patients (43.2%) had recurrence
within 5 years of surgery and 26 patients (32.0%) died of primary
ESCC. The clinicopathological features were evaluated using the 8th
edition of the International Union Against Cancer
(UICC)/tumor-node-metastasis (TNM) Classification of Malignant
Tumors (25). The present study was
approved by the Faculty of Science Ethics Committee of the Kyoto
Prefectural University of Medicine (approval no. ERB-C-1414-1).
Immunohistochemistry (IHC)
IHC staining was performed using the labeled
streptavidin-biotin method using a Vectastain Universal Quick kit
(Vector Laboratories, Inc.) as per the manufacturer's protocol. The
method can be briefly described as follows: antigen retrieval was
performed using heated Dako Target Retrieval Solution (Dako;
Agilent Technologies, Inc.) at 95°C for 60 min and endogenous
peroxidase activity was disrupted by incubation in 0.3%
H2O2 at room temperature for 20 min; then,
2.5% normal horse serum (Vector Laboratories, Inc.) was used to
block non-specific binding at room temperature for 10 min. The
slides were incubated with either the anti-AGR2 antibody 1:500
(cat. no. Ab76473; Abcam) or the anti-p53 antibody 1:200 (mouse,
NCL-L-p53-DO7; Novocastra Laboratories Ltd.) at 4°C overnight. They
were then incubated with a biotinylated pan-specific universal
antibody at room temperature for 10 min and a
streptavidin/peroxidase complex (Vector Laboratories, Inc.) at room
temperature for 5 min, followed by incubation with
3,3′-diaminobenzidine tetrachloride. Counterstaining was performed
using hematoxylin. A formalin-fixed ESCC cell line overexpressing
AGR2 (TE15 cells) was used as the positive control.
The AGR2 staining was evaluated based on a score
depicting the intensity and proportion of stained cancer cells. The
intensity was scored as 3 (strong), 2 (intermediate), 1 (weak), or
0 (no staining). The proportion was evaluated as the ratio of
stained areas in the cancer tissue and was scored from 0 to 1.
Cancer tissues with an intensity score of 3 and a proportion ≥0.3
or an intensity score of 2 and a proportion ≥0.5 were considered
the high expression group and the others comprised the low
expression group.
p53 staining was evaluated in a manner similar to
that of AGR2. The intensity was scored as 3 (strong), 2
(intermediate), 1 (weak), or 0 (no staining) and the proportion was
scored from 0 to 1. Cancer tissues with an intensity score of 3 or
2 and a proportion ≥0.3 were considered the high expression group
and the others comprised the low expression group.
Statistical analysis
Statistical analysis was performed using JMP Version
14 (ASA Institute). The Wilcoxon signed-rank test or Student's
t-test was used to compare differences between the paired and
unpaired samples. Differences between survival curves were examined
using the log-rank test. Multivariate analysis of survival was
carried out using the Cox regression method. Relationships between
clinicopathological factors were assessed using Fisher's exact test
or the Chi-square test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of AGR2 in tissue and cell
lines
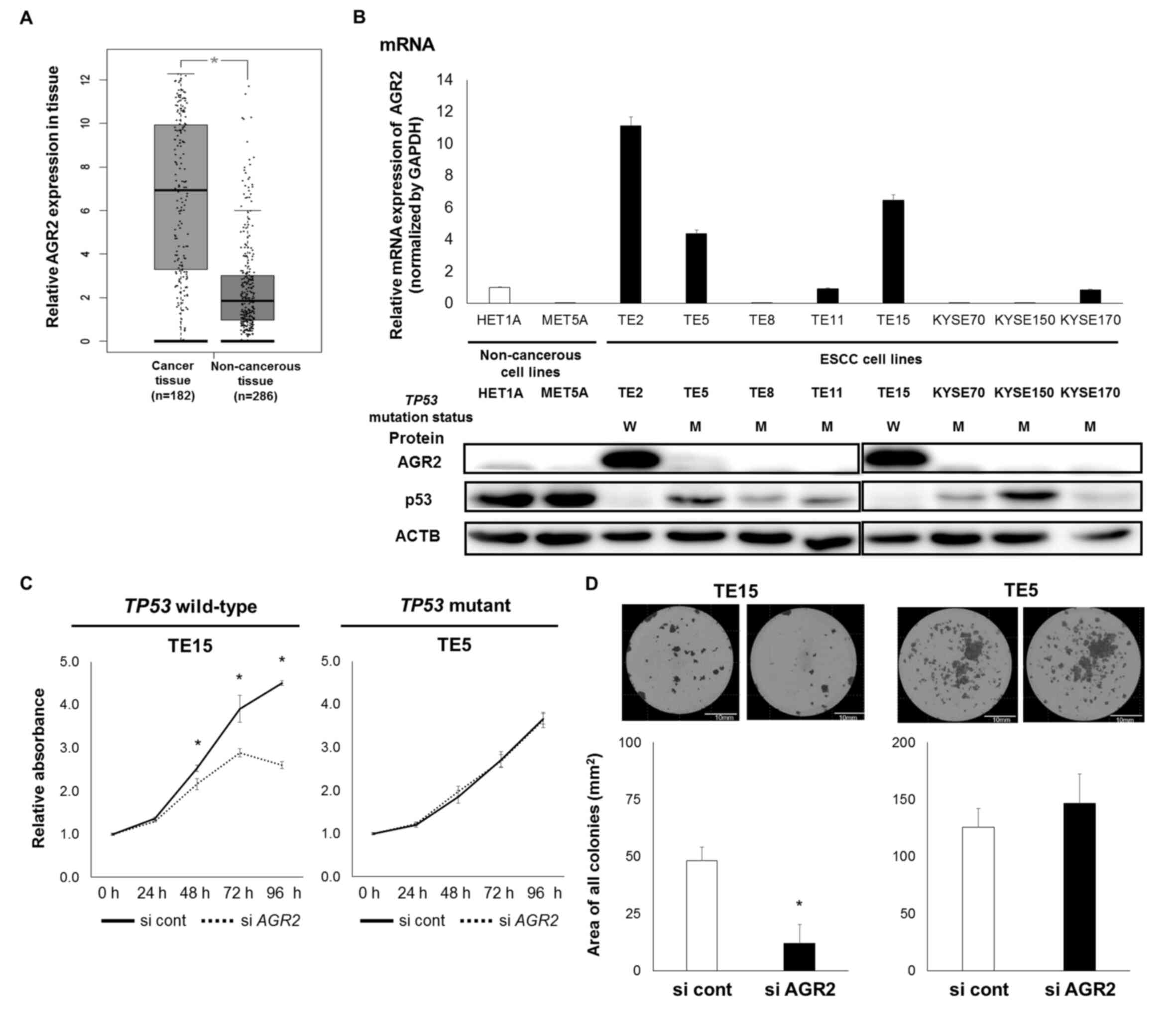
The mRNA expression of AGR2 was significantly
higher in EC tissue compared with the normal
counterpart(P<0.001; Fig. 1A),
according to GEPIA (26). Among the
nine ESCC cell lines and two non-cancerous cell lines, the mRNA and
protein expression of AGR2 was higher in the TE2, TE5 and TE15
cells compared with the other cells (Fig.
1B). p53 protein expression was also confirmed, a result that
was partly consistent with the previously reported mutation status
(Fig. 1B) (27).
Effects of AGR2 downregulation on the
functions of ESCC cells
AGR2-expressing cell lines TE15 and TE5, which are
reported to be TP53-wild-type and mutant cell lines,
respectively (27,28), were selected for the subsequent
experiments. The transient introduction of the siRNA targeting AGR2
efficiently downregulated AGR2 mRNA and protein expression levels
(Fig. S1). As determined using the
proliferation assay, the downregulation of AGR2 expression
inhibited the growth of TE15 cells but not TE5 cells (Fig. 1C). The downregulation of AGR2 also
reduced colony formation in TE15 cells but not TE5 cells (Fig. 1D).
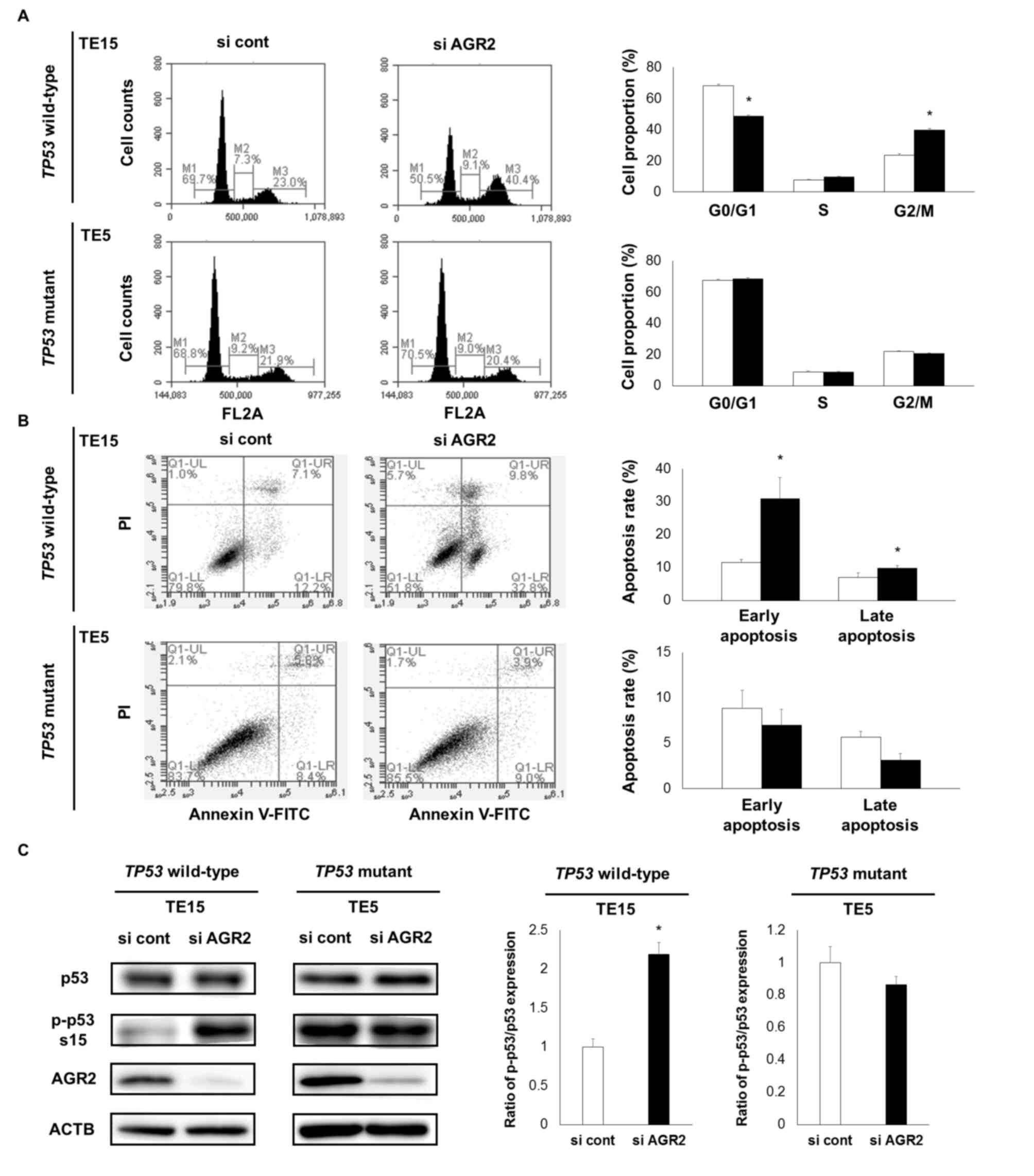
In addition, cell cycle analysis demonstrated that
AGR2 downregulation increased the proportion of TE15 cells, but not
TE5 cells, in the G2/M phase (Fig.
2A). The apoptosis assay demonstrated that early and late
apoptosis was increased only for the TE15 cells (Fig. 2B). The TE2 cell line, which is a
TP53-wild-type cell line with AGR2 expression, demonstrated
similar changes in cell function as TE15 cells owing to AGR2
downregulation (Fig. S2A-E).
Relationships between AGR2 expression
and phosphorylation of p53
Based on the aforementioned results, the
associations between AGR2 expression and the phosphorylation or
mutation status of TP53 were examined. The phosphorylation
of p53 Ser15 was markedly enhanced by the downregulation of AGR2 in
TP53-wild-type TE15 cells (Fig.
2C). By contrast, in TP53-mutant TE5 cells, the
phosphorylation of p53 was already high and not enhanced by the
downregulation of AGR2 (Fig. 2C).
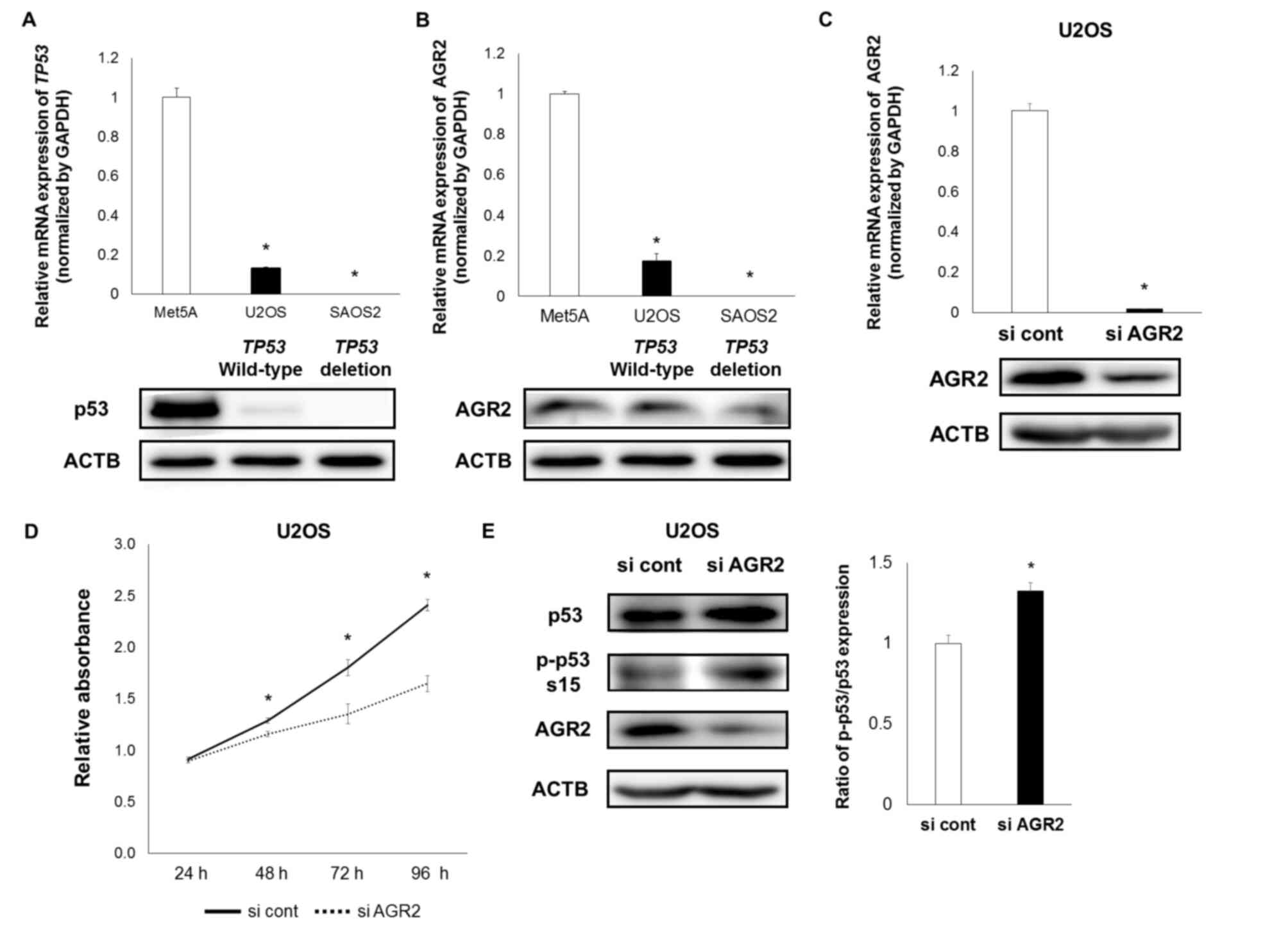
For further verification of these associations,
similar examinations were performed using the osteosarcoma cell
lines U2OS, which is a TP53-wild-type cell line and SAOS2,
which is a TP53-deficient cell line (29). AGR2 and p53 expression was confirmed in
U2OS cells, whereas in SAOS2 cells, AGR2 expression was very low
and p53 expression was not detected (Fig.
3A and B). The present study therefore only examined the
effects of AGR2 downregulation in U2OS cells. The downregulation of
AGR2 also suppressed the proliferation of and enhanced p53 Ser15
phosphorylation in U2OS cells (Fig.
3C-E).
Immunohistochemistry of AGR2 and p53
in ESCC tissues
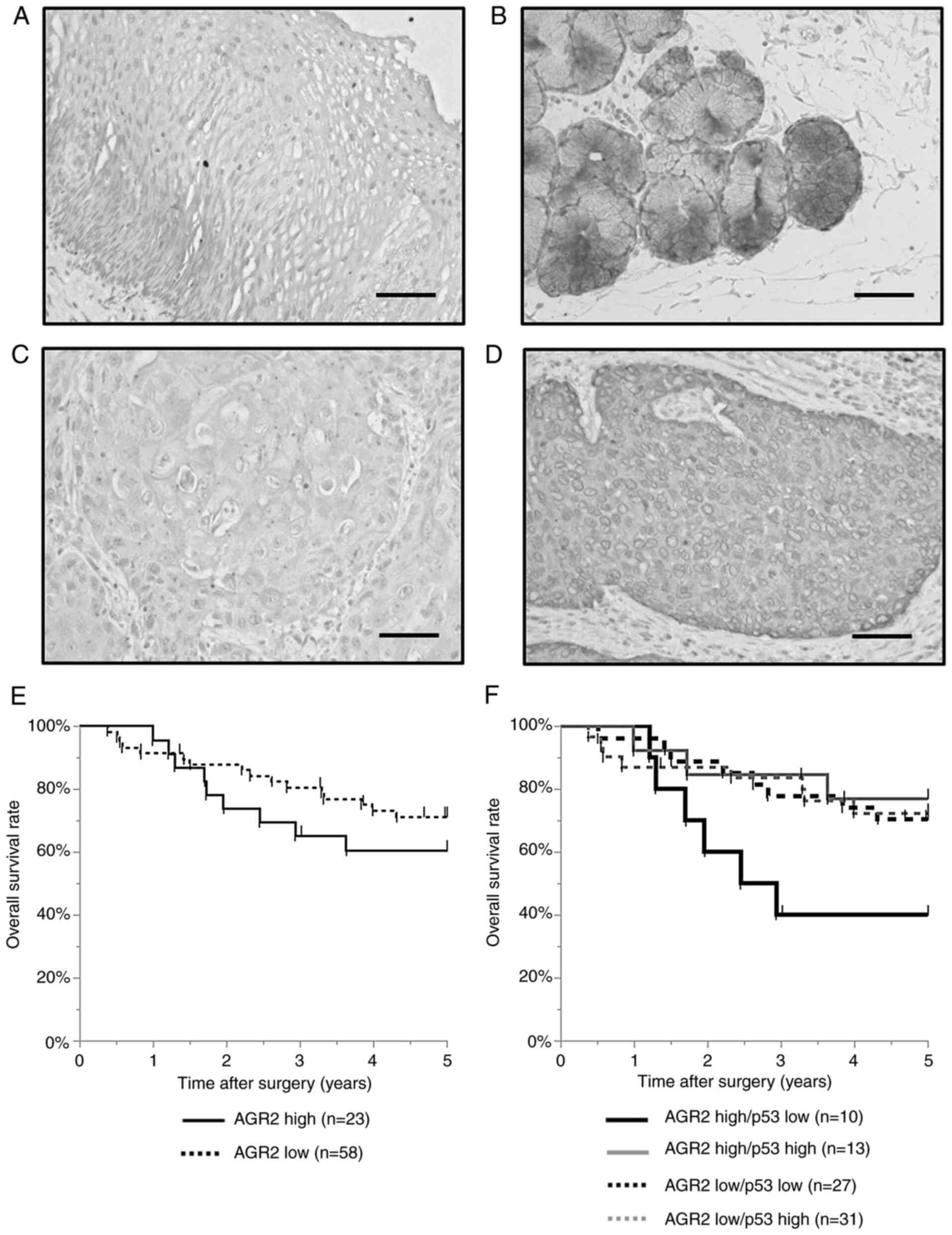
AGR2 was expressed in the esophageal gland but not
the non-cancerous esophageal epithelium (Fig. 4A and B). In ESCC tissue, AGR2 was
detected in the cell membrane and cytoplasm and the level of AGR2
expression was categorized as either low or high (Fig. 4C and D). No significant differences
were observed in the 5-year overall survival rate was between the
high and low expression groups (60.5 vs. 71.2%; P=0.32; Fig. 4E). In addition, no significant
differences were observed in the clinicopathological features
between these groups (Table I).
 | Table I.Relationship between AGR2 expression
and clinicopathological features. |
Table I.
Relationship between AGR2 expression
and clinicopathological features.
| Factors | AGR2 (−) n=58 | AGR2 (+) n=23 | P-value |
|---|
| Age (years) |
|
| 0.63 |
|
≥65 | 29 | 10 |
|
|
<65 | 29 | 13 |
|
| Sex |
|
| 1.00 |
|
Female | 9 | 4 |
|
|
Male | 49 | 19 |
|
| T
factora |
|
| 0.11 |
| T3 | 22 | 4 |
|
|
T1/2 | 36 | 19 |
|
| N
factora |
|
| 0.78 |
|
N2/N3 | 13 | 6 |
|
|
N0/N1 | 45 | 17 |
|
| Stagea |
|
| 0.80 |
| III,
Iva | 21 | 9 |
|
| I,
II | 37 | 14 |
|
| Histopathological
typeb |
|
| 0.78 |
| Well,
moderate | 42 | 15 |
|
|
Poorly | 15 | 7 |
|
| Tumor size
(mm) |
|
| 0.81 |
|
≥40 | 33 | 12 |
|
|
<40 | 25 | 11 |
|
| Lymphatic
invasion |
|
| 0.09 |
|
Present | 28 | 16 |
|
|
Absent | 30 | 7 |
|
| Venous
invasion |
|
| 1.00 |
|
Present | 26 | 10 |
|
|
Absent | 32 | 13 |
|
| Adjuvant
chemotherapy |
|
| 0.34 |
|
Present | 28 | 14 |
|
|
Absent | 30 | 9 |
|
| IHC status of
p53 |
|
| 1.00 |
|
High | 31 | 13 |
|
|
Low | 27 | 10 |
|
The result of IHC for p53 was also examined and
categorized into low and high expression groups (Fig. S3A and B). No significant differences
were observed in the 5-year overall survival rate between the high
and low p53 expression groups (73.7 vs. 61.9%, P=0.28; Fig. S3C). However, with respect to AGR2 and
p53 expression in combination, patients with high-AGR2/low-p53
expression demonstrated significantly worse prognosis than other
patients. The 5-year overall survival rate was 40.0% in the
high-AGR2/low-p53 expression group, whereas it was 70.3% in the
low-AGR2/low-p53 expression group (P=0.046; Fig. 4F). The survival analysis of patients
with low p53 expression demonstrated that a large tumor of >40
mm in size (HR=3.82, P=0.03) and high AGR2 expression (HR=3.66;
P=0.03) were independent prognostic factors (Table II).
| Table II.Survival analysis of patients with
low p53 expression. |
Table II.
Survival analysis of patients with
low p53 expression.
|
|
|
|
| Multivariate |
|---|
|
|
|
|
|
|
|---|
| Factors | n | 5-year OS (%) | Univariate
P-value | HR | 95% CI | P-value |
|---|
| Age (years) |
|
| 0.82 |
|
|
|
|
≥65 | 16 | 62.5 |
|
|
|
|
|
<65 | 21 | 61.9 |
|
|
|
|
| Sex |
|
| 1.00 |
|
|
|
|
Female | 5 | 60.0 |
|
|
|
|
|
Male | 17 | 62.2 |
|
|
|
|
| T
factora |
|
| 0.69 |
|
|
|
| T3 | 13 | 69.2 |
|
|
|
|
|
T1/2 | 24 | 57.7 |
|
|
|
|
| N
Factora |
|
| 0.04 | 1.98 | 0.63–5.92 | 0.23 |
|
N2/N3 | 10 | 40.0 |
|
|
|
|
|
N0/N1 | 27 | 70.1 |
|
|
|
|
| Stagea |
|
| 0.19 |
|
|
|
| III,
IVa | 12 | 50.0 |
|
|
|
|
| I,
II | 25 | 67.7 |
|
|
|
|
| Histopathological
typeb |
|
| 0.86 |
|
|
|
| Well,
moderate | 26 | 61.3 |
|
|
|
|
|
Poorly | 11 | 63.6 |
|
|
|
|
| Tumor size
(mm) |
|
| 0.03 | 3.82 | 1.17–14.83 | 0.03 |
|
≥40 | 18 | 50.0 |
|
|
|
|
|
<40 | 19 | 78.6 |
|
|
|
|
| Lymphatic
invasion |
|
| 0.75 |
|
|
|
|
Present | 22 | 63.0 |
|
|
|
|
|
Absent | 15 | 60.0 |
|
|
|
|
| Venous
invasion |
|
| 0.36 |
|
|
|
|
Present | 18 | 55.0 |
|
|
|
|
|
Absent | 19 | 68.4 |
|
|
|
|
| Adjuvant
chemotherapy |
|
| 0.64 |
|
|
|
|
Present | 21 | 66.6 |
|
|
|
|
|
Absent | 16 | 55.5 |
|
|
|
|
| IHC status of
AGR2 |
|
| 0.047 | 3.66 | 1.12–11.56 | 0.03 |
|
High | 10 | 40.0 |
|
|
|
|
|
Low | 27 | 70.3 |
|
|
|
|
Discussion
Several studies have shown that the downregulation
of AGR2 decreases cell proliferation (15–17,30) and induces cell cycle arrest (31) or apoptosis (32,33) in some
types of cancer. The present study demonstrated that AGR2
downregulation in TP53-wild-type ESCC cells suppressed cell
proliferation and increased the proportion of G2/M-phase and
apoptotic cells. In addition, it enhanced the phosphorylation of
p53 Ser15. However, these changes were not observed in
TP53-mutant ESCC cells. Similar findings were also verified
in TP53-wild-type osteosarcoma cells. In addition, the IHC
results demonstrated that high AGR2 and low p53 expression in ESCC
tissue was associated with worse prognosis. These results suggest
that AGR2 expression enhances cell proliferation via the inhibition
of p53 phosphorylation in TP53-wild-type ESCC and that this
mechanism does not regulate cell functions in TP53-mutant
ESCC because the function of p53 is already impaired.
The correlation between AGR2 overexpression in solid
tumors and poor prognosis has been reported (12) and it is particularly clear in
adenocarcinoma tissue (8,17,34). There
are some reports on SCC of the lung or head and neck, but the
potential of AGR2 as a prognostic biomarker would be weaker in
those cells compared with adenocarcinoma (6,8,13–15,34). Indeed, AGR2 is reported to be a useful
positive marker of esophageal adenocarcinoma (35). However, there are no reports of AGR2
functions in ESCC; to the best of the authors' knowledge, the
present study is the first report on the molecular mechanism and
clinical significance of AGR2 expression in SCC of the
esophagus.
Phosphorylation regulation as a protein disulfide
isomerase is an important mechanism in the function of AGR2 as an
oncogenic factor. Pohler et al (19) report that AGR2 attenuates p53 activity,
suppressing its phosphorylation even in preneoplastic Barrett's
esophageal epithelium. Hrstka et al (18) mention that AGR2 upregulates DUSP10
expression, which results in the inhibition of p38
mitogen-activated protein kinase and p53 activation. In addition,
Sicari et al (36) report that
ER stress in cancer cells causes AGR2 migration to the cytoplasm
and the inhibition of p53 signaling via the binding of AGR2 to p53
protein. TP53, one of the most well-known tumor-suppressive
genes, controls cell proliferation via the regulation of the cell
cycle and apoptosis (21,22). A mutation in TP53 is reported to
reduce the anti-proliferative and apoptotic functions of p53 and
occasionally even exert an oncogenic function (37). In the present study, AGR2
downregulation increased the phosphorylation of p53 Ser15 and
decreased malignant potential in the TP53-wild-type cell
lines TE15, TE2 and U2OS, but not in the TP53-mutant cell
line TE5 because the phosphorylation of p53 is frequently induced
without AGR2 expression in p53-mutant cells.
In the present study, prognosis, as determined by
AGR2 expression in ESCC tissues, demonstrated no significant
differences between groups. However, higher AGR2 expression was
correlated with worse prognosis in patients with lower p53
expression. The presence of a TP53 gene mutation generally
induces the accumulation of the p53 protein because the half-life
of a mutant p53 protein is extended (38–40). Thus,
the level of mutant p53 protein expression can be evaluated by IHC,
but wild-type p53 cannot be detected. Therefore, as in the present
study, lower expression of p53 based on IHC closely reflected
wild-type p53 protein expression. These IHC results are consistent
with altered AGR2 regulation in p53-wild-type cell lines. The
present study also performed IHC of phosphorylated p53; however,
almost all tissues were negative (data not shown). This is due to
the fact that the expression level of phosphorylated p53 in
p53-wild-type cells was much lower compared with p53-mutant cells,
as shown in the present study, indicating that the expression of
phosphorylated p53 is unstable and difficult to detect by IHC.
Regarding AGR2 overexpression, Hrstka et al
(41) report on the AKT-dependent
pathway in tamoxifen-resistant cancer. Ondrouskova et al
(42) mention that HER2 expression
upregulates AGR2 expression in a hormone-independent manner in
breast cancer. In addition, Pohler et al (19) speculate that the activity of the AGR2
pathway in suppressing p53 could be related to the TP53
mutation status. In the present study, AGR2 overexpression was
detected in 33% (3/9) of the cell lines and 28% (23/81) of the
tissue samples. These rates were consistent with the results of a
previous report showing that AGR2 overexpression could be detected
in 36% (15/41) of ESCC samples (35).
However, the level of AGR2 expression was not significantly related
to that of p53 expression in the ESCC tissue samples (Table I). There might be several mechanisms
that upregulate AGR2 expression in p53-wild-type cancer cells to
attenuate the active tumor-suppressive functions of p53. In this
case, AGR2 inhibition, such as blocking with an antibody targeting
this protein (43), might become a
potential therapeutic option for TP53-wild-type ESCC.
There are some limitations to the present study. The
number of patients was small and it was difficult to arrive at a
definite conclusion. Therefore, examinations based on more samples,
particularly for patients with SCC, are needed. No definitive
TP53 mutation status in cell lines and tissues was confirmed
in the present study. However, the results obtained from the cell
lines are similar to previous findings (27). The frequency of the TP53
mutation in the ESCC tissues was reported to be in the range of 40
to 80% (27,44). These results are consistent with the
data from the present study showing that the high p53 expression
group comprised 54% (44/81) of cases, although some functionally
suppressed p53 mutants might be mixed in with the low-p53
expression group and it remains unclear whether p53 activity in
p53-wild-type cells is a clinically effective marker for cancer
progression.
In conclusion, AGR2 serves an important role in the
progression of TP53-wild-type ESCC by inhibiting the
phosphorylation of p53. AGR2 might be a useful prognostic marker
and potential therapeutic target in patients with
TP53-wild-type ESCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
All authors conceived and planned the experiments.
KT carried out the experiments. KT and HK confirm the authenticity
of all the raw data. KT, HK, TA and KS performed data analysis. KT,
HK, TA, SK, JS, HF, WT, KS, HS, YY, SK and AS contributed to the
interpretation of the results. OK and OE assisted in research
planning based on the results obtained. KT and HK wrote the first
draft of the manuscript. All authors provided critical feedback and
helped shape the research, analysis and manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participant
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki Declaration and its later amendments or comparable ethical
standards. The present study was approved by the Faculty of Science
Ethics Committee of Kyoto Prefectural University of Medicine.
Informed consent was obtained from all individual participants
included in the study (approval no. ERB-C-1414-1).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ACTB
|
β-actin
|
|
AGR2
|
anterior gradient 2
|
|
EC
|
esophageal cancer
|
|
ER
|
endoplasmic reticulum
|
|
ESCC
|
esophageal cell carcinoma
|
|
HNSCC
|
head and neck squamous cell
carcinoma
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
siRNA
|
small interfering RNA
|
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rustgi AK and El-Serag HB: Esophageal
carcinoma. N Engl J Med. 371:2499–2509. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cohen DJ and Leichman L: Controversies in
the treatment of local and locally advanced gastric and esophageal
cancers. J Clin Oncol. 33:1754–1759. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gray TA, Alsamman K, Murray E, Sims AH and
Hupp TR: Engineering a synthetic cell panel to identify signalling
components reprogrammed by the cell growth regulator anterior
gradient-2. Mol Biosyst. 10:1409–1425. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park SW, Zhen G, Verhaeghe C, Nakagami Y,
Nguyenvu LT, Barczak AJ, Killeen N and Erle DJ: The protein
disulfide isomerase AGR2 is essential for production of intestinal
mucus. Proc Natl Acad Sci USA. 106:6950–6955. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brychtova V, Vojtesek B and Hrstka R:
Anterior gradient 2: A novel player in tumor cell biology. Cancer
Lett. 304:1–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thompson DA and Weigel RJ: hAG-2, the
Human Homologue of the Xenopus laevis Cement Gland Gene XAG-2, is
coexpressed with estrogen receptor in breast cancer cell lines.
Biochem Biophys Res Commun. 251:111–116. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alavi M, Mah V, Maresh EL, Bagryanova L,
Horvath S, Chia D, Goodglick L and Liu AY: High expression of AGR2
in lung cancer is predictive of poor survival. BMC Cancer.
15:6552015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Armes JE, Davies CM, Wallace S, Taheri T,
Perrin LC and Autelitano DJ: AGR2 expression in ovarian tumours: A
potential biomarker for endometrioid and mucinous differentiation.
Pathology. 45:49–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lowe AW, Olsen M, Hao Y, Lee SP, Taek Lee
K, Chen X, Van de Rijn M and Brown PO: Gene expression patterns in
pancreatic tumors, cells and tissues. PLoS One. 2:e3232007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Forootan SS, Liu D, Barraclough
R, Foster CS, Rudland PS and Ke Y: Increased expression of anterior
gradient-2 is significantly associated with poor survival of
prostate cancer patients. Prostate Cancer Prostatic Dis.
10:293–300. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tian SB, Tao KX, Hu J, Liu ZB, Ding XL,
Chu YN, Cui JY, Shuai XM, Gao JB, Cai KL, et al: The prognostic
value of AGR2 expression in solid tumours: A systematic review and
meta-analysis. Sci Rep. 7:155002017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma SR, Wang WM, Huang CF, Zhang WF and Sun
ZJ: Anterior gradient protein 2 expression in high grade head and
neck squamous cell carcinoma correlated with cancer stem cell and
epithelial mesenchymal transition. Oncotarget. 6:8807–8821. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun B, Cheng Z and Sun J: Associations of
MACC1, AGR2, and KAI1 expression with the metastasis and prognosis
in head and neck squamous cell carcinoma. Int J Clin Exp Patho.
11:822–830. 2018.PubMed/NCBI
|
|
15
|
Sweeny L, Liu Z, Bush BD, Hartman Y, Zhou
T and Rosenthal EL: CD147 and AGR2 expression promote cellular
proliferation and metastasis of head and neck squamous cell
carcinoma. Exp Cell Res. 318:1788–1798. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma SR, Mao L, Deng WW, Li YC, Bu LL, Yu
GT, Zhang WF and Sun ZJ: AGR2 promotes the proliferation, migration
and regulates epithelial-mesenchymal transition in salivary adenoid
cystic carcinoma. Am J Transl Res. 9:507–519. 2017.PubMed/NCBI
|
|
17
|
Wang Z, Hao Y and Lowe AW: The
Adenocarcinoma-associated antigen, AGR2, promotes tumor growth,
cell migration, and cellular transformation. Cancer Res.
68:492–497. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hrstka R, Bouchalova P, Michalova E,
Matoulkova E, Muller P, Coates PJ and Vojtesek B: AGR2 oncoprotein
inhibits p38 MAPK and p53 activation through a DUSP10-mediated
regulatory pathway. Mol Oncol. 10:652–662. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pohler E, Craig AL, Cotton J, Lawrie L,
Dillon JF, Ross P, Kernohan N and Hupp TR: The Barrett's antigen
anterior gradient-2 silences the p53 transcriptional response to
DNA damage. Mol Cell Proteomics. 3:534–547. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Siliciano JD, Canman CE, Taya Y, Sakaguchi
K, Appella E and Kastan MB: DNA damage induces phosphorylation of
the amino terminus of p53. Genes Dev. 11:3471–3481. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Joruiz SM and Bourdon JC: p53 Isoforms:
Key regulators of the cell fate decision. Cold Spring Harb Perspect
Med. 6:a0260392016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lane DP: Cancer. p53, guardian of the
genome. Nature. 358:15–16. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Albrechtsen N, Dornreiter I, Grosse F, Kim
E, Wiesmüller L and Deppert W: Maintenance of genomic integrity by
p53: Complementary roles for activated and non-activated p53.
Oncogene. 18:7706–7717. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Brierley JD, Gospodarowicz MK and
Wittekind C: TNM classification of malignant tumours. John Wiley
& Sons; 2016
|
|
26
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wagata T, Shibagaki I, Imamura M, Shimada
Y, Toguchida J, Yandell DW, Ikenaga M, Tobe T and Ishizaki K: Loss
of 17p, mutation of the p53 gene, and overexpression of p53 protein
in esophageal squamous cell carcinomas. Cancer Res. 53:846–850.
1993.PubMed/NCBI
|
|
28
|
Saeki H, Kitao H, Yoshinaga K, Nakanoko T,
Kubo N, Kakeji Y, Morita M and Maehara Y: Copy-neutral loss of
heterozygosity at the p53 locus in carcinogenesis of esophageal
squamous cell carcinomas associated with p53 mutations. Clin Cancer
Res. 17:1731–1740. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Janssens V, Van Hoof C, De Baere I,
Merlevede W and Goris J: The phosphotyrosyl phosphatase activator
gene is a novel p53 target gene. J Biol Chem. 275:20488–20495.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tsuji T, Satoyoshi R, Aiba N, Kubo T,
Yanagihara K, Maeda D, Goto A, Ishikawa K, Yashiro M and Tanaka M:
Agr2 mediates paracrine effects on stromal fibroblasts that promote
invasion by gastric signet-ring carcinoma cells. Cancer Res.
75:356–366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu Z, Gu Y, Han B, Zhang J, Li Z, Tian K,
Young CY and Yuan H: Knockdown of AGR2 induces cellular senescence
in prostate cancer cells. Carcinogenesis. 33:1178–1186. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu QG, Li YJ and Yao L: Knockdown of AGR2
induces cell apoptosis and reduces chemotherapy resistance of
pancreatic cancer cells with the involvement of ERK/AKT axis.
Pancreatology. 18:678–688. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu R, Qian M, Zhou T and Cui P: TP53
mediated miR-3647-5p prevents progression of cervical carcinoma by
targeting AGR2. Cancer Med. 8:6095–6105. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pizzi M, Fassan M, Balistreri M,
Galligioni A, Rea F and Rugge M: Anterior gradient 2 overexpression
in lung adenocarcinoma. Appl Immunohistochem Mol Morphol. 20:31–36.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
DiMaio MA, Kwok S, Montgomery KD, Lowe AW
and Pai RK: Immunohistochemical panel for distinguishing esophageal
adenocarcinoma from squamous cell carcinoma: A combination of p63,
cytokeratin 5/6, MUC5AC, and anterior gradient homolog 2 allows
optimal subtyping. Hum Pathol. 43:1799–1807. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sicari D, Centonze FG, Pineau R, Le Reste
PJ, Negroni L, Chat S, Mohtar MA, Thomas D, Gillet R, Hupp T, et
al: Reflux of endoplasmic reticulum proteins to the cytosol
inactivates tumor suppressors. EMBO Rep. 22:e514122021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Soussi T and Lozano G: p53 mutation
heterogeneity in cancer. Biochem Biophys Res Commun. 331:834–842.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Barnas C, Martel-Planche G, Furukawa Y,
Hollstein M, Montesano R and Hainaut P: Inactivation of the p53
protein in cell lines derived from human esophageal cancers. Int J
Cancer. 71:79–87. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bennett WP, Hollstein MC, He A, Zhu SM,
Resau JH, Trump BF, Metcalf RA, Welsh JA, Midgley C, Lane DP, et
al: Archival analysis of p53 genetic and protein alterations in
Chinese esophageal cancer. Oncogene. 6:1779–1784. 1991.PubMed/NCBI
|
|
40
|
Finlay CA, Hinds PW, Tan TH, Eliyahu D,
Oren M and Levine AJ: Activating mutations for transformation by
p53 produce a gene product that forms an hsc70-p53 complex with an
altered half-life. Mol Cell Biol. 8:531–539. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hrstka R, Murray E, Brychtova V, Fabian P,
Hupp TR and Vojtesek B: Identification of an AKT-dependent
signalling pathway that mediates tamoxifen-dependent induction of
the pro-metastatic protein anterior gradient-2. Cancer Lett.
333:187–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ondrouskova E, Sommerova L, Nenutil R,
Coufal O, Bouchal P, Vojtesek B and Hrstka R: AGR2 associates with
HER2 expression predicting poor outcome in subset of estrogen
receptor negative breast cancer patients. Exp Mol Pathol.
102:280–283. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Arumugam T, Deng D, Bover L, Wang H,
Logsdon CD and Ramachandran V: New blocking antibodies against
novel AGR2-C4.4A pathway reduce growth and metastasis of pancreatic
tumors and increase survival in mice. Mol Cancer Ther. 14:941–951.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li LY, Tang JT, Jia LQ and Li PW:
Mutations of p53 gene exons 4–8 in human esophageal cancer. World J
Gastroenterol. 11:2998–3001. 2005. View Article : Google Scholar : PubMed/NCBI
|