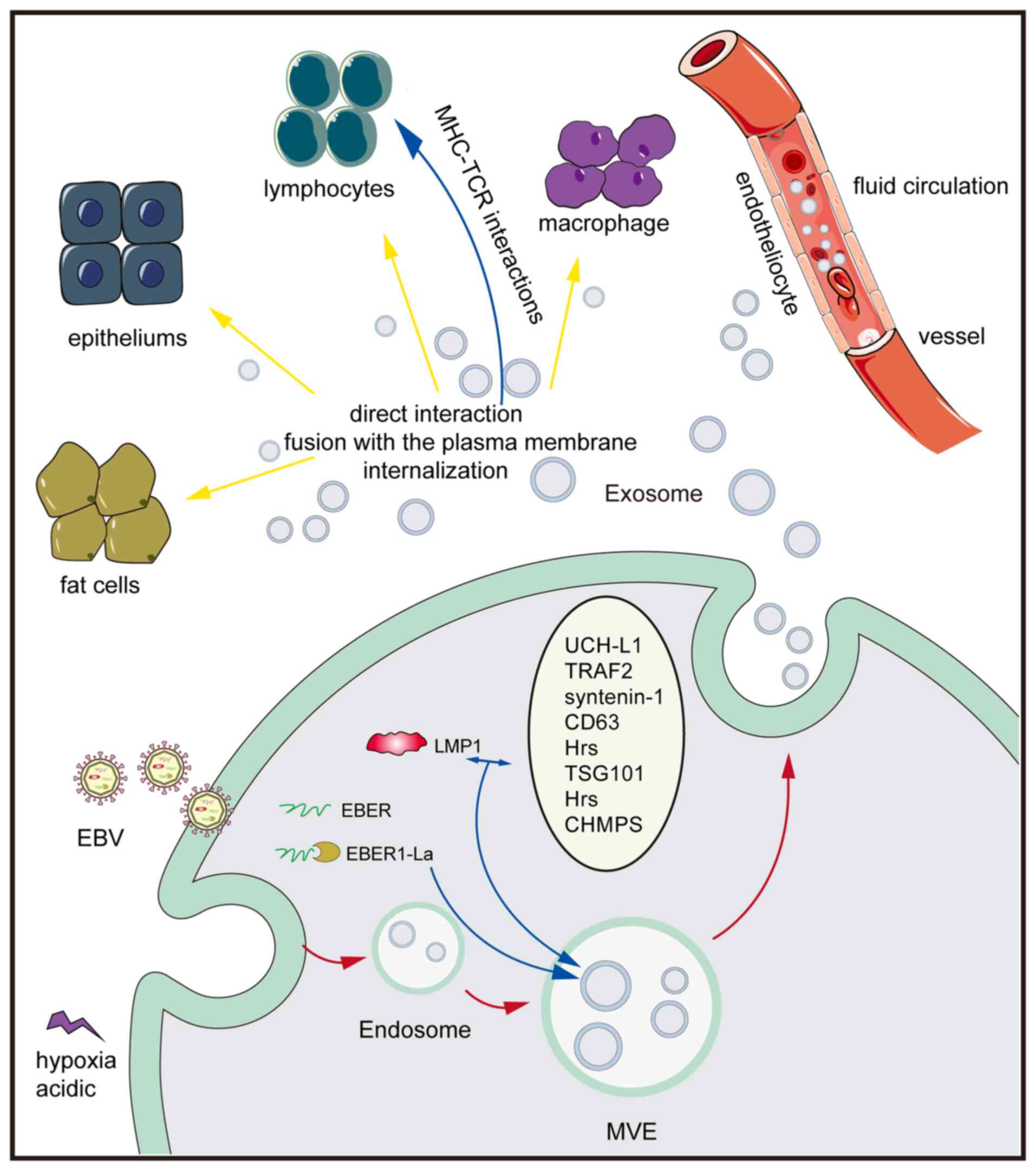
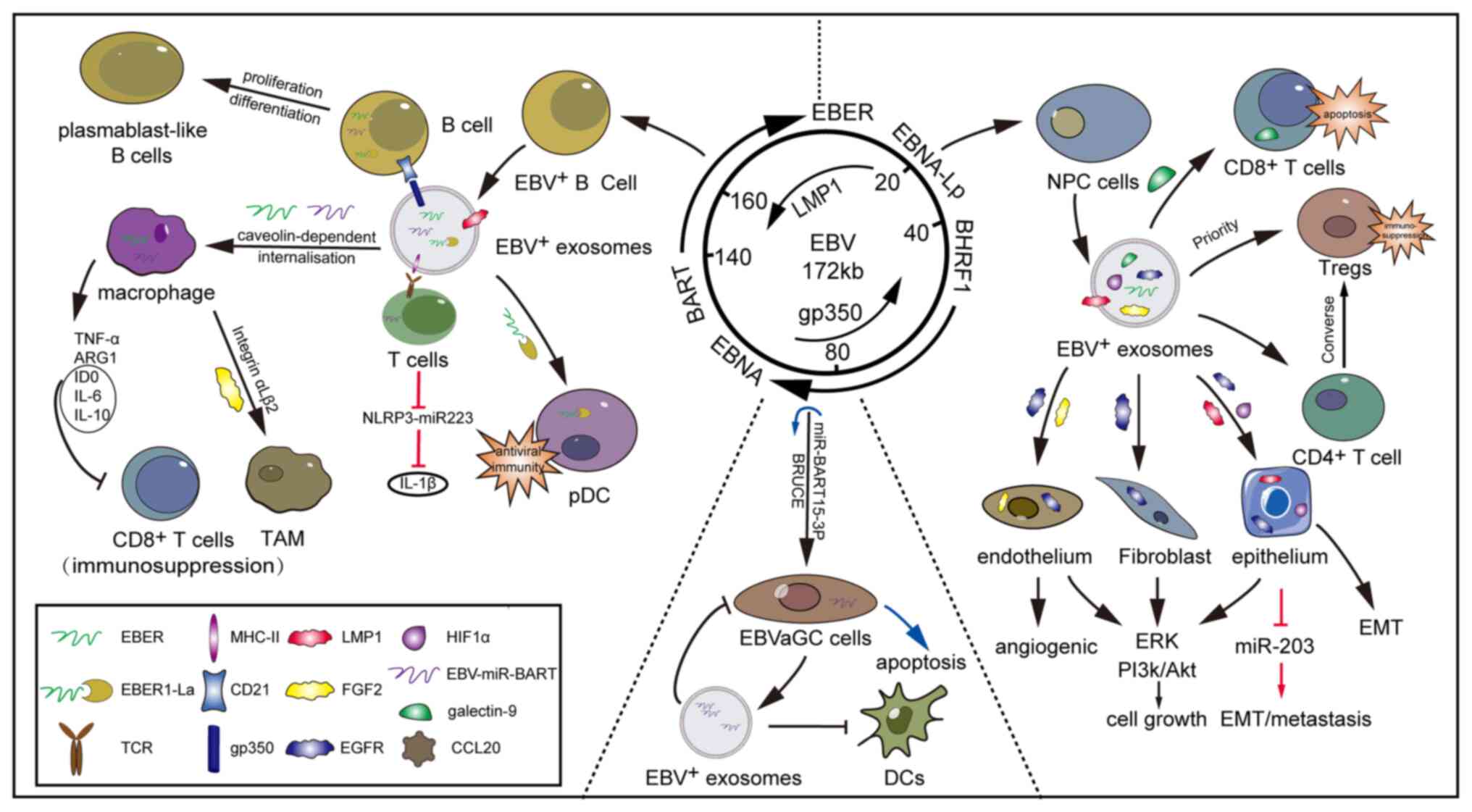
|
1
|
Epstein MA, Achong BG and Barr YM: Virus particles in cultured lymphoblasts from Burkitt's lymphoma. Lancet. 1:702–703. 1964. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vockerodt M, Yap LF, Shannon-Lowe C, Curley H, Wei W, Vrzalikova K and Murray PG: The Epstein-Barr virus and the pathogenesis of lymphoma. J Pathol. 235:312–322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsao SW, Tsang CM, To KF and Lo KW: The role of Epstein-Barr virus in epithelial malignancies. J Pathol. 235:323–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Niller HH, Banati F, Salamon D and Minarovits J: Epigenetic alterations in Epstein-Barr virus-associated diseases. Adv Exp Med Biol. 879:39–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lyu X, Wang J, Guo X, Wu G, Jiao Y, Faleti OD, Liu P, Liu T, Long Y, Chong T, et al: EBV-miR-BART1-5P activates AMPK/mTOR/HIF1 pathway via a PTEN independent manner to promote glycolysis and angiogenesis in nasopharyngeal carcinoma. PLoS Pathog. 14:e10074842018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hulse M, Johnson SM, Boyle S, Caruso LB and Tempera I: Epstein-Barr virus-encoded latent membrane protein 1 and B-cell growth transformation induce lipogenesis through fatty acid synthase. J Virol. 95:e01857–20. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang J, Qin Y, Yang C, Wan C, Dai X, Sun Y, Meng J, Lu Y, Li Y, Zhang Z, et al: Downregulation of ABI2 expression by EBV-miR-BART13-3p induces epithelial-mesenchymal transition of nasopharyngeal carcinoma cells through upregulation of c-JUN/SLUG signaling. Aging (Albany NY). 12:340–358. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cai L, Ye Y, Jiang Q, Chen Y, Lyu X, Li J, Wang S, Liu T, Cai H, Yao K, et al: Epstein-Barr virus-encoded microRNA BART1 induces tumour metastasis by regulating PTEN-dependent pathways in nasopharyngeal carcinoma. Nat Commun. 6:73532015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wong Y, Meehan MT, Burrows SR, Doolan DL and Miles JJ: Estimating the global burden of Epstein-Barr virus-related cancers. J Cancer Res Clin Oncol. Oct 27–2021.(Epub ahead of print). View Article : Google Scholar
|
|
10
|
AbuSalah MAH, Gan SH, Al-Hatamleh MAI, Irekeola AA, Shueb RH and Yean Yean C: Recent advances in diagnostic approaches for Epstein-Barr virus. Pathogens. 9:2262020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Théry C, Zitvogel L and Amigorena S: Exosomes: Composition, biogenesis and function. Nat Rev Immunol. 2:569–579. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Raposo G and Stoorvogel W: Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol. 200:373–383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nanbo A, Katano H, Kataoka M, Hoshina S, Sekizuka T, Kuroda M and Ohba Y: Infection of Epstein-Barr virus in type III latency modulates biogenesis of exosomes and the expression profile of exosomal miRNAs in the burkitt lymphoma mutu cell lines. Cancers (Basel). 10:2372018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kobayashi E, Aga M, Kondo S, Whitehurst C, Yoshizaki T, Pagano JS and Shackelford J: C-terminal farnesylation of UCH-L1 plays a role in transport of Epstein-Barr virus primary oncoprotein LMP1 to exosomes. mSphere. 3:e00030–18. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Villarroya-Beltri C, Baixauli F, Gutiérrez-Vázquez C, Sánchez-Madrid F and Mittelbrunn M: Sorting it out: Regulation of exosome loading. Semin Cancer Biol. 28:3–13. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ahmed W, Tariq S and Khan G: Tracking EBV-encoded RNAs (EBERs) from the nucleus to the excreted exosomes of B-lymphocytes. Sci Rep. 8:154382018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zuo L, Xie Y, Tang J, Xin S, Liu L, Zhang S, Yan Q, Zhu F and Lu J: Targeting exosomal EBV-LMP1 transfer and miR-203 expression via the NF-κB pathway: The therapeutic role of aspirin in NPC. Mol Ther Nucleic Acids. 17:175–184. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen H, Wang L, Zeng X, Schwarz H, Nanda HS, Peng X and Zhou Y: Exosomes, a new star for targeted delivery. Front Cell Dev Biol. 9:7510792021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Henne WM, Buchkovich NJ and Emr SD: The ESCRT pathway. Dev Cell. 21:77–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, van Eijndhoven MA, Hopmans ES, Lindenberg JL, de Gruijl TD, Würdinger T and Middeldorp JM: Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA. 107:6328–6333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kosaka N, Iguchi H, Hagiwara K, Yoshioka Y, Takeshita F and Ochiya T: Neutral sphingomyelinase 2 (nSMase2)-dependent exosomal transfer of angiogenic microRNAs regulate cancer cell metastasis. J Biol Chem. 288:10849–10859. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Val S, Krueger A, Poley M, Cohen A, Brown K, Panigrahi A and Preciado D: Nontypeable Haemophilus influenzae lysates increase heterogeneous nuclear ribonucleoprotein secretion and exosome release in human middle-ear epithelial cells. FASEB J. 32:1855–1867. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Villarroya-Beltri C, Gutiérrez-Vázquez C, Sánchez-Cabo F, Pérez-Hernández D, Vázquez J, Martin-Cofreces N, Martinez-Herrera DJ, Pascual-Montano A, Mittelbrunn M and Sánchez-Madrid F: Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat Commun. 4:29802013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Martín-Cófreces NB, Baixauli F and Sánchez-Madrid F: Immune synapse: Conductor of orchestrated organelle movement. Trends Cell Biol. 24:61–72. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Morelli AE, Larregina AT, Shufesky WJ, Sullivan ML, Stolz DB, Papworth GD, Zahorchak AF, Logar AJ, Wang Z, Watkins SC, et al: Endocytosis, intracellular sorting, and processing of exosomes by dendritic cells. Blood. 104:3257–3266. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blanc L and Vidal M: New insights into the function of Rab GTPases in the context of exosomal secretion. Small GTPases. 9:95–106. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jahn R and Scheller RH: SNAREs-engines for membrane fusion. Nat Rev Mol Cell Biol. 7:631–643. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tkach M, Kowal J, Zucchetti AE, Enserink L, Jouve M, Lankar D, Saitakis M, Martin-Jaular L and Théry C: Qualitative differences in T-cell activation by dendritic cell-derived extracellular vesicle subtypes. EMBO J. 36:3012–3028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Prada I and Meldolesi J: Binding and fusion of extracellular vesicles to the plasma membrane of their cell targets. Int J Mol Sci. 17:12962016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tian T, Zhu YL, Hu FH, Wang YY, Huang NP and Xiao ZD: Dynamics of exosome internalization and trafficking. J Cell Physiol. 228:1487–1495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang L, Huang X, Guo H, Wang L, Yang W, Wu W, Jing D and Shao Z: Exosomes as efficient nanocarriers in osteosarcoma: Biological functions and potential clinical applications. Front Cell Dev Biol. 9:7373142021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Koppers-Lalic D, Hackenberg M, Bijnsdorp IV, van Eijndhoven MAJ, Sadek P, Sie D, Zini N, Middeldorp JM, Ylstra B, de Menezes RX, et al: Nontemplated nucleotide additions distinguish the small RNA composition in cells from exosomes. Cell Rep. 8:1649–1658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nkosi D, Sun L, Duke LC, Patel N, Surapaneni SK, Singh M and Meckes DG Jr: Epstein-Barr virus LMP1 promotes syntenin-1- and Hrs-induced extracellular vesicle formation for its own secretion to increase cell proliferation and migration. mBio. 11:e00589–20. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Friedl P and Alexander S: Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell. 147:992–1009. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao TJ, Zhu N, Shi YN, Wang YX, Zhang CJ, Deng CF, Liao DF and Qin L: Targeting HDL in tumor microenvironment: New hope for cancer therapy. J Cell Physiol. 236:7853–7873. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Quail DF and Joyce JA: Microenvironmental regulation of tumor progression and metastasis. Nat Med. 19:1423–1437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
de Jong OG, Verhaar MC, Chen Y, Vader P, Gremmels H, Posthuma G, Schiffelers RM, Gucek M and van Balkom BW: Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J Extracell Vesicles. 1:2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao H, Yang L, Baddour J, Achreja A, Bernard V, Moss T, Marini JC, Tudawe T, Seviour EG, San Lucas FA, et al: Tumor microenvironment derived exosomes pleiotropically modulate cancer cell metabolism. Elife. 5:e102502016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Parolini I, Federici C, Raggi C, Lugini L, Palleschi S, De Milito A, Coscia C, Iessi E, Logozzi M, Molinari A, et al: Microenvironmental pH is a key factor for exosome traffic in tumor cells. J Biol Chem. 284:34211–34222. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Park JE, Tan HS, Datta A, Lai RC, Zhang H, Meng W, Lim SK and Sze SK: Hypoxic tumor cell modulates its microenvironment to enhance angiogenic and metastatic potential by secretion of proteins and exosomes. Mol Cell Proteomics. 9:1085–1099. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hsu YL, Hung JY, Chang WA, Lin YS, Pan YC, Tsai PH, Wu CY and Kuo PL: Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene. 36:4929–4942. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
King HW, Michael MZ and Gleadle JM: Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer. 12:4212012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xue M, Chen W, Xiang A, Wang R, Chen H, Pan J, Pang H, An H, Wang X, Hou H and Li X: Hypoxic exosomes facilitate bladder tumor growth and development through transferring long non-coding RNA-UCA1. Mol Cancer. 16:1432017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ramteke A, Ting H, Agarwal C, Mateen S, Somasagara R, Hussain A, Graner M, Frederick B, Agarwal R and Deep G: Exosomes secreted under hypoxia enhance invasiveness and stemness of prostate cancer cells by targeting adherens junction molecules. Mol Carcinog. 54:554–565. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dorayappan KDP, Wanner R, Wallbillich JJ, Saini U, Zingarelli R, Suarez AA, Cohn DE and Selvendiran K: Hypoxia-induced exosomes contribute to a more aggressive and chemoresistant ovarian cancer phenotype: A novel mechanism linking STAT3/Rab proteins. Oncogene. 37:3806–3821. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Farrell PJ: Epstein-Barr virus and cancer. Annu Rev Pathol. 14:29–53. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pfeffer S, Zavolan M, Grässer FA, Chien M, Russo JJ, Ju J, John B, Enright AJ, Marks D, Sander C and Tuschl T: Identification of virus-encoded microRNAs. Science. 304:734–736. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Meister G: miRNAs get an early start on translational silencing. Cell. 131:25–28. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hadinoto V, Shapiro M, Sun CC and Thorley-Lawson DA: The dynamics of EBV shedding implicate a central role for epithelial cells in amplifying viral output. PLoS Pathog. 5:e10004962009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Münz C: Epstein-Barr Virus-specific immune control by innate lymphocytes. Front Immunol. 8:16582017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cohen JI: Epstein-Barr virus infection. N Engl J Med. 343:481–492. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Crombie JL and LaCasce AS: Epstein Barr Virus associated B-cell lymphomas and iatrogenic lymphoproliferative disorders. Front Oncol. 9:1092019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chiu YF and Sugden B: Epstein-Barr Virus: The path from latent to productive infection. Annu Rev Virol. 3:359–372. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kanda T: EBV-encoded latent genes. Adv Exp Med Biol. 1045:377–394. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yang JH, Sun XS, Xiao BB, Liu LT, Guo SS, Liang JD, Jia GD, Tang LQ, Chen QY and Mai HQ: Subdivision of de-novo metastatic nasopharyngeal carcinoma based on tumor burden and pretreatment EBV DNA for therapeutic guidance of locoregional radiotherapy. BMC Cancer. 21:5342021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
He Y, Yang D, Zhou T, Xue W, Zhang J, Li F, Wang F, Wang T, Wu Z, Liao Y, et al: Epstein-Barr virus DNA loads in the peripheral blood cells predict the survival of locoregionally-advanced nasopharyngeal carcinoma patients. Cancer Biol Med. 18:888–899. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Qin JQ, Yin H, Wu JZ, Chen RZ, Xia Y, Wang L, Zhu HY, Fan L, Li JY, Liang JH and Xu W: Pretreatment whole blood Epstein-Barr virus DNA predicts prognosis in Hodgkin lymphoma. Leuk Res. 107:1066072021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim JH, Cho H, Sung H, Jung AR, Lee YS, Lee SW, Ryu JS, Chae EJ, Kim KW, Huh J, et al: Reappraisal of the prognostic value of Epstein-Barr virus status in monomorphic post-transplantation lymphoproliferative disorders-diffuse large B-cell lymphoma. Sci Rep. 11:28802021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Xie Y, Wang T and Wang L: Hydroa vacciniforme-like lymphoproliferative disorder: A study of clinicopathology and whole-exome sequencing in Chinese patients. J Dermatol Sci. 99:128–134. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Miyake T, Yamamoto T, Hirai Y, Otsuka M, Hamada T, Tsuji K, Morizane S, Suzuki D, Aoyama Y and Iwatsuki K: Survival rates and prognostic factors of Epstein-Barr virus-associated hydroa vacciniforme and hypersensitivity to mosquito bites. Br J Dermatol. 172:56–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Thorley-Lawson DA: EBV persistence-introducing the virus. Curr Top Microbiol Immunol. 390:151–209. 2015.PubMed/NCBI
|
|
62
|
Gutzeit C, Nagy N, Gentile M, Lyberg K, Gumz J, Vallhov H, Puga I, Klein E, Gabrielsson S, Cerutti A and Scheynius A: Exosomes derived from Burkitt's lymphoma cell lines induce proliferation, differentiation, and class-switch recombination in B cells. J Immunol. 192:5852–5862. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ito M, Kudo K, Higuchi H, Otsuka H, Tanaka M, Fukunishi N, Araki T, Takamatsu M, Ino Y, Kimura Y and Kotani A: Proteomic and phospholipidomic characterization of extracellular vesicles inducing tumor microenvironment in Epstein-Barr virus-associated lymphomas. FASEB J. 35:e215052021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Young LS and Rickinson AB: Epstein-Barr virus: 40 Years on. Nat Rev Cancer. 4:757–768. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hurwitz SN, Nkosi D, Conlon MM, York SB, Liu X, Tremblay DC and Meckes DG Jr: CD63 regulates Epstein-Barr Virus LMP1 exosomal packaging, enhancement of vesicle production, and noncanonical NF-κB signaling. J Virol. 91:e02251–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Middeldorp JM and Pegtel DM: Multiple roles of LMP1 in Epstein-Barr virus induced immune escape. Semin Cancer Biol. 18:388–396. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kaye KM, Izumi KM and Kieff E: Epstein-Barr virus latent membrane protein 1 is essential for B-lymphocyte growth transformation. Proc Natl Acad Sci USA. 90:9150–9154. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Nkosi D, Howell LA, Cheerathodi MR, Hurwitz SN, Tremblay DC, Liu X and Meckes DG Jr: Transmembrane domains mediate intra- and extracellular trafficking of Epstein-Barr virus latent membrane protein 1. J Virol. 92:e00280–18. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Rialland P, Lankar D, Raposo G, Bonnerot C and Hubert P: BCR-bound antigen is targeted to exosomes in human follicular lymphoma B-cells. Biol Cell. 98:491–501. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Vallhov H, Gutzeit C, Johansson SM, Nagy N, Paul M, Li Q, Friend S, George TC, Klein E, Scheynius A and Gabrielsson S: Exosomes containing glycoprotein 350 released by EBV-transformed B cells selectively target B cells through CD21 and block EBV infection in vitro. J Immunol. 186:73–82. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Nanbo A, Kawanishi E, Yoshida R and Yoshiyama H: Exosomes derived from Epstein-Barr virus-infected cells are internalized via caveola-dependent endocytosis and promote phenotypic modulation in target cells. J Virol. 87:10334–10347. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Samanta M, Iwakiri D, Kanda T, Imaizumi T and Takada K: EB virus-encoded RNAs are recognized by RIG-I and activate signaling to induce type I IFN. EMBO J. 25:4207–4214. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Nanbo A, Inoue K, Adachi-Takasawa K and Takada K: Epstein-Barr virus RNA confers resistance to interferon-alpha-induced apoptosis in Burkitt's lymphoma. EMBO J. 21:954–965. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fok V, Friend K and Steitz JA: Epstein-Barr virus noncoding RNAs are confined to the nucleus, whereas their partner, the human La protein, undergoes nucleocytoplasmic shuttling. J Cell Biol. 173:319–325. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Baglio SR, van Eijndhoven MA, Koppers-Lalic D, Berenguer J, Lougheed SM, Gibbs S, Léveillé N, Rinkel RN, Hopmans ES, Swaminathan S, et al: Sensing of latent EBV infection through exosomal transfer of 5′pppRNA. Proc Natl Acad Sci USA. 113:E587–E596. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Burassakarn A, Srisathaporn S, Pientong C, Wongjampa W, Vatanasapt P, Patarapadungkit N and Ekalaksananan T: Exosomes-carrying Epstein-Barr virus-encoded small RNA-1 induces indoleamine 2,3-dioxygenase expression in tumor-infiltrating macrophages of oral squamous-cell carcinomas and suppresses T-cell activity by activating RIG-I/IL-6/TNF-α pathway. Oral Oncol. 117:1052792021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Pratt ZL, Kuzembayeva M, Sengupta S and Sugden B: The microRNAs of Epstein-Barr Virus are expressed at dramatically differing levels among cell lines. Virology. 386:387–397. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Higuchi H, Yamakawa N, Imadome KI, Yahata T, Kotaki R, Ogata J, Kakizaki M, Fujita K, Lu J, Yokoyama K, et al: Role of exosomes as a proinflammatory mediator in the development of EBV-associated lymphoma. Blood. 131:2552–2567. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Haneklaus M, Gerlic M, Kurowska-Stolarska M, Rainey AA, Pich D, McInnes IB, Hammerschmidt W, O'Neill LA and Masters SL: Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1β production. J Immunol. 189:3795–3799. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Bauernfeind FG, Horvath G, Stutz A, Alnemri ES, MacDonald K, Speert D, Fernandes-Alnemri T, Wu J, Monks BG, Fitzgerald KA, et al: Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol. 183:787–791. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Johnnidis JB, Harris MH, Wheeler RT, Stehling-Sun S, Lam MH, Kirak O, Brummelkamp TR, Fleming MD and Camargo FD: Regulation of progenitor cell proliferation and granulocyte function by microRNA-223. Nature. 451:1125–1129. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Chim CS, Ma SY, Au WY, Choy C, Lie AK, Liang R, Yau CC and Kwong YL: Primary nasal natural killer cell lymphoma: Long-term treatment outcome and relationship with the international prognostic index. Blood. 103:216–221. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Lee JH, Choi J, Ahn YO, Kim TM and Heo DS: CD21-independent Epstein-Barr virus entry into NK cells. Cell Immunol. 327:21–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ahmed W, Philip PS, Attoub S and Khan G: Epstein-Barr virus-infected cells release Fas ligand in exosomal fractions and induce apoptosis in recipient cells via the extrinsic pathway. J Gen Virol. 96:3646–3659. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Chuang YC, Hsieh MC, Lin CC, Lo YS, Ho HY, Hsieh MJ and Lin JT: Pinosylvin inhibits migration and invasion of nasopharyngeal carcinoma cancer cells via regulation of epithelial-mesenchymal transition and inhibition of MMP2. Oncol Rep. 46:1432021. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Keryer-Bibens C, Pioche-Durieu C, Villemant C, Souquère S, Nishi N, Hirashima M, Middeldorp J and Busson P: Exosomes released by EBV-infected nasopharyngeal carcinoma cells convey the viral latent membrane protein 1 and the immunomodulatory protein galectin 9. BMC Cancer. 6:2832006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Hirashima M, Kashio Y, Nishi N, Yamauchi A, Imaizumi TA, Kageshita T, Saita N and Nakamura T: Galectin-9 in physiological and pathological conditions. Glycoconj J. 19:593–600. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Miller WE and Raab-Traub N: The EGFR as a target for viral oncoproteins. Trends Microbiol. 7:453–458. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Meckes DG Jr, Shair KH, Marquitz AR, Kung CP, Edwards RH and Raab-Traub N: Human tumor virus utilizes exosomes for intercellular communication. Proc Natl Acad Sci USA. 107:20370–20375. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yang MH and Wu KJ: TWIST activation by hypoxia inducible factor-1 (HIF-1): Implications in metastasis and development. Cell Cycle. 7:2090–2096. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Aga M, Bentz GL, Raffa S, Torrisi MR, Kondo S, Wakisaka N, Yoshizaki T, Pagano JS and Shackelford J: Exosomal HIF1α supports invasive potential of nasopharyngeal carcinoma-associated LMP1-positive exosomes. Oncogene. 33:4613–4622. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Basilico C and Moscatelli D: The FGF family of growth factors and oncogenes. Adv Cancer Res. 59:115–165. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ceccarelli S, Visco V, Raffa S, Wakisaka N, Pagano JS and Torrisi MR: Epstein-Barr virus latent membrane protein 1 promotes concentration in multivesicular bodies of fibroblast growth factor 2 and its release through exosomes. Int J Cancer. 121:1494–1506. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Yu H, Lu J, Zuo L, Yan Q, Yu Z, Li X, Huang J, Zhao L, Tang H, Luo Z, et al: Epstein-Barr virus downregulates microRNA 203 through the oncoprotein latent membrane protein 1: A contribution to increased tumor incidence in epithelial cells. J Virol. 86:3088–3099. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Mrizak D, Martin N, Barjon C, Jimenez-Pailhes AS, Mustapha R, Niki T, Guigay J, Pancré V, de Launoit Y, Busson P, et al: Effect of nasopharyngeal carcinoma-derived exosomes on human regulatory T cells. J Natl Cancer Inst. 107:3632014.PubMed/NCBI
|
|
96
|
Yogev O, Henderson S, Hayes MJ, Marelli SS, Ofir-Birin Y, Regev-Rudzki N, Herrero J and Enver T: Herpesviruses shape tumour microenvironment through exosomal transfer of viral microRNAs. PLoS Pathog. 13:e10065242017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Fan C, Tang Y, Wang J, Xiong F, Guo C, Wang Y, Xiang B, Zhou M, Li X, Wu X, et al: The emerging role of Epstein-Barr virus encoded microRNAs in nasopharyngeal carcinoma. J Cancer. 9:2852–2864. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lei T, Yuen KS, Xu R, Tsao SW, Chen H, Li M, Kok KH and Jin DY: Targeting of DICE1 tumor suppressor by Epstein-Barr virus-encoded miR-BART3* microRNA in nasopharyngeal carcinoma. Int J Cancer. 133:79–87. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Wang J, Zheng X, Qin Z, Wei L, Lu Y, Peng Q, Gao Y, Zhang X, Zhang X, Li Z, et al: Epstein-Barr virus miR-BART3-3p promotes tumorigenesis by regulating the senescence pathway in gastric cancer. J Biol Chem. 294:4854–4866. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Murphy G, Pfeiffer R, Camargo MC and Rabkin CS: Meta-analysis shows that prevalence of Epstein-Barr virus-positive gastric cancer differs based on sex and anatomic location. Gastroenterology. 137:824–833. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Burke AP, Yen TS, Shekitka KM and Sobin LH: Lymphoepithelial carcinoma of the stomach with Epstein-Barr virus demonstrated by polymerase chain reaction. Mod Pathol. 3:377–380. 1990.PubMed/NCBI
|
|
102
|
Choi H, Lee H, Kim SR, Gho YS and Lee SK: Epstein-Barr virus-encoded microRNA BART15-3p promotes cell apoptosis partially by targeting BRUCE. J Virol. 87:8135–8144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Balkwill FR, Capasso M and Hagemann T: The tumor microenvironment at a glance. J Cell Sci. 125:5591–5596. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Hinata M, Kunita A, Abe H, Morishita Y, Sakuma K, Yamashita H, Seto Y, Ushiku T and Fukayama M: Exosomes of Epstein-Barr virus-associated gastric carcinoma suppress dendritic cell maturation. Microorganisms. 8:17762020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Camargo MC, Kim WH, Chiaravalli AM, Kim KM, Corvalan AH, Matsuo K, Yu J, Sung JJ, Herrera-Goepfert R, Meneses-Gonzalez F, et al: Improved survival of gastric cancer with tumour Epstein-Barr virus positivity: An international pooled analysis. Gut. 63:236–243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Caby MP, Lankar D, Vincendeau-Scherrer C, Raposo G and Bonnerot C: Exosomal-like vesicles are present in human blood plasma. Int Immunol. 17:879–887. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Houali K, Wang X, Shimizu Y, Djennaoui D, Nicholls J, Fiorini S, Bouguermouh A and Ooka T: A new diagnostic marker for secreted Epstein-Barr virus encoded LMP1 and BARF1 oncoproteins in the serum and saliva of patients with nasopharyngeal carcinoma. Clin Cancer Res. 13:4993–5000. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Mao Y, Zhang DW, Zhu H, Lin H, Xiong L, Cao Q, Liu Y, Li QD, Xu JR, Xu LF and Chen RJ: LMP1 and LMP2A are potential prognostic markers of extranodal NK/T-cell lymphoma, nasal type (ENKTL). Diagn Pathol. 7:1782012. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Luan X, Yang W, Bai X, Li H, Li H, Fan W, Zhang H, Liu W and Sun L: Cyclophilin A is a key positive and negative feedback regulator within interleukin-6 trans-signaling pathway. FASEB J. 35:e219582021. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Liu L, Zuo L, Yang J, Xin S, Zhang J, Zhou J, Li G, Tang J and Lu J: Exosomal cyclophilin A as a novel noninvasive biomarker for Epstein-Barr virus associated nasopharyngeal carcinoma. Cancer Med. 8:3142–3151. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhang G, Zong J, Lin S, Verhoeven RJ, Tong S, Chen Y, Ji M, Cheng W, Tsao SW, Lung M, et al: Circulating Epstein-Barr virus microRNAs miR-BART7 and miR-BART13 as biomarkers for nasopharyngeal carcinoma diagnosis and treatment. Int J Cancer. 136:E301–E312. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Wardana T, Gunawan L, Herawati C, Oktriani R, Anwar S, Astuti I, Aryandono T and Mubarika S: Circulation EBV Mir-Bart-7 relating to clinical manifestation in nasopharyngeal carcinoma. Asian Pac J Cancer Prev. 21:2777–2782. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Notarte KI, Senanayake S, Macaranas I, Albano PM, Mundo L, Fennell E, Leoncini L and Murray P: MicroRNA and other non-coding RNAs in Epstein-Barr virus-associated cancers. Cancers (Basel). 13:39092021. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
De Re V, Caggiari L, De Zorzi M, Fanotto V, Miolo G, Puglisi F, Cannizzaro R, Canzonieri V, Steffan A, Farruggia P, et al: Epstein-Barr virus BART microRNAs in EBV-associated Hodgkin lymphoma and gastric cancer. Infect Agent Cancer. 15:422020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhao Y, Liu P, Tan H, Chen X, Wang Q and Chen T: Exosomes as smart nanoplatforms for diagnosis and therapy of cancer. Front Oncol. 11:7431892021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Coulouarn C, Factor VM, Andersen JB, Durkin ME and Thorgeirsson SS: Loss of miR-122 expression in liver cancer correlates with suppression of the hepatic phenotype and gain of metastatic properties. Oncogene. 28:3526–3536. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lou G, Song X, Yang F, Wu S, Wang J, Chen Z and Liu Y: Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J Hematol Oncol. 8:1222015. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Wang J, Jiang Q, Faleti OD, Tsang CM, Zhao M, Wu G, Tsao SW, Fu M, Chen Y, Ding T, et al: Exosomal delivery of antagomirs targeting viral and cellular MicroRNAs synergistically inhibits cancer angiogenesis. Mol Ther Nucleic Acids. 22:153–165. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Pitt JM, Marabelle A, Eggermont A, Soria JC, Kroemer G and Zitvogel L: Targeting the tumor microenvironment: Removing obstruction to anticancer immune responses and immunotherapy. Ann Oncol. 27:1482–1492. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Wang X, Xiang Z, Liu Y, Huang C, Pei Y, Wang X, Zhi H, Wong WH, Wei H, Ng IO, et al: Exosomes derived from Vδ2-T cells control Epstein-Barr virus-associated tumors and induce T cell antitumor immunity. Sci Transl Med. 12:eaaz34262020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Kwon SH, Faruque HA, Kee H, Kim E and Park S: Exosome-based hybrid nanostructures for enhanced tumor targeting and hyperthermia therapy. Colloids Surf B Biointerfaces. 205:1119152021. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Babcock GJ, Decker LL, Volk M and Thorley-Lawson DA: EBV persistence in memory B cells in vivo. Immunity. 9:395–404. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Min K, Kim JY and Lee SK: Epstein-Barr virus miR-BART1-3p suppresses apoptosis and promotes migration of gastric carcinoma cells by targeting DAB2. Int J Biol Sci. 16:694–707. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Navari M, Etebari M, De Falco G, Ambrosio MR, Gibellini D, Leoncini L and Piccaluga PP: The presence of Epstein-Barr virus significantly impacts the transcriptional profile in immunodeficiency-associated Burkitt lymphoma. Front Microbiol. 6:5562015. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Westhoff Smith D and Sugden B: Potential cellular functions of Epstein-Barr nuclear antigen 1 (EBNA1) of Epstein-Barr virus. Viruses. 5:226–240. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Choy EY, Siu KL, Kok KH, Lung RW, Tsang CM, To KF, Kwong DL, Tsao SW and Jin DY: An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J Exp Med. 205:2551–2560. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Chiang AK, Tao Q, Srivastava G and Ho FC: Nasal NK- and T-cell lymphomas share the same type of Epstein-Barr virus latency as nasopharyngeal carcinoma and Hodgkin's disease. Int J Cancer. 68:285–290. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Sakamoto K, Sekizuka T, Uehara T, Hishima T, Mine S, Fukumoto H, Sato Y, Hasegawa H, Kuroda M and Katano H: Next-generation sequencing of miRNAs in clinical samples of Epstein-Barr virus-associated B-cell lymphomas. Cancer Med. 6:605–618. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Tsao SW, Tsang CM and Lo KW: Epstein-Barr virus infection and nasopharyngeal carcinoma. Philos Trans R Soc Lond B Biol Sci. 372:201602702017. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zebardast A, Tehrani SS, Latifi T and Sadeghi F: Critical review of Epstein-Barr virus microRNAs relation with EBV-associated gastric cancer. J Cell Physiol. 236:6136–6153. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Meckes DG Jr, Gunawardena HP, Dekroon RM, Heaton PR, Edwards RH, Ozgur S, Griffith JD, Damania B and Raab-Traub N: Modulation of B-cell exosome proteins by gamma herpesvirus infection. Proc Natl Acad Sci USA. 110:E2925–E2933. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Pei Y, Wong JHY and Robertson ES: Targeted therapies for Epstein-Barr virus-associated lymphomas. Cancers (Basel). 12:25652020. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Spender LC, Lucchesi W, Bodelon G, Bilancio A, Karstegl CE, Asano T, Dittrich-Breiholz O, Kracht M, Vanhaesebroeck B and Farrell PJ: Cell target genes of Epstein-Barr virus transcription factor EBNA-2: Induction of the p55alpha regulatory subunit of PI3-kinase and its role in survival of EREB2.5 cells. J Gen Virol. 87:2859–2867. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Ahmed W and Khan G: The labyrinth of interactions of Epstein-Barr virus-encoded small RNAs. Rev Med Virol. 24:3–14. 2014. View Article : Google Scholar : PubMed/NCBI
|