Introduction
Renal cell carcinoma (RCC) is a commonly encountered
and lethal malignancy, accounting for ~2% of all cancer cases and
related deaths worldwide (1), its
major subtypes being clear cell RCC (ccRCC), papillary PCC (pRCC)
and chromophobe RCC (chRCC). Of all the RCC subtypes, ccRCC is the
most common histological manifestation and its pathogenesis is
characterized by the constitutive activation of hypoxia-inducible
factors (HIFs) due to the loss-of-function in the von Hippel-Lindau
(VHL) tumor suppressor gene (1).
Various targeted therapies against vascular endothelial growth
factor (VEGF) or mammalian target of rapamycin (mTOR) signaling and
immune check point inhibitors for metastatic disease have been
developed. However, disease progression is inevitable in the
majority of patients (1,2).
A hallmark of ccRCC is the abundance of
intracellular lipid droplets (LDs), consisting of a neutral lipid
core containing triglycerides (TGs) and cholesterol-esters (CEs)
surrounded by a phospholipid monolayer (3). LDs are dynamic organelles responsible
for lipid uptake and storage, and are used to maintain homeostasis,
facilitate energy production and protect against various types of
stress or to sustain membrane biogenesis during rapid tumor cell
growth in several types of cancer (4). In fact, previous studies have
demonstrated that LDs contribute to ccRCC progression (5,6).
The fatty acids (FAs) which comprise the main
component of lipids have been reported to serve as substrates for
energy storage, membrane synthesis and the production of signaling
molecules. Elongation and desaturation are central steps of the
de novo synthesis of long-chain FAs (LC-FAs), the length and
degree of unsaturation being determinants of FA function and
metabolic fate (7). Stearoyl-CoA
desaturase 1 (SCD1), a member of the fatty acyl desaturase family,
has been extensively studied in ccRCC (6,8,9). It
has been demonstrated that increased SCD1 expression supports
viability, while a SCD1 small molecule inhibitor (A939572) has been
shown to suppress cellular proliferation in ccRCC (8,9).
Moreover, Yang et al (10)
demonstrated that sterol regulatory element-binding protein 1
(SREBP1) promoted lipid desaturation through FA acid desaturase 1
(FADS1) prior to the activation of NF-κB signaling for the
promotion of cellular proliferation in ccRCC. However, any
potential roles of elongases in RCC remain unclear.
It has been reported that, in mammals, the initial
and rate-controlling FA condensation reactions are catalyzed by a
family of elongase enzymes referred to as elongation of
very-long-chain FAs (ELOVL). To date, seven ELOVL members have been
identified, which can be generally divided into those specific for
saturated and monounsaturated FAs (ELOVL1, ELOVL3, ELOVL6 and
ELOVL7) or for polyunsaturated FAs (PUFAs; ELOVL2, ELOVL4 and
ELOVL5). Lucarelli et al (9) revealed a significant accumulation of
PUFAs and an increased expression of ELOVL2 and ELOVL5 in ccRCC. Of
note, it has been demonstrated that systemic docosahexaenoic acid
(DHA) is endogenously produced and controls de novo
lipogenesis, while also regulating lipid storage in a sterol
regulatory element-binding transcription factor 1
(SREBPF1)-independent manner, due to ELOVL2 ablation in mice
(11). Moreover, ELOVL2
overexpression has been shown to promote LD accumulation with an
enhanced FA uptake in both the preadipocyte cell line 3T3-L1 and
F442A cells (12). Previous in
vitro studies, with DHA exogenously administered for cancer
cells, have demonstrated that DHA exerts anti-tumorigenic,
anti-inflammatory and pro-apoptotic effects on cancer cells
(13–15). However, any potential role of
ELOVL2 in ccRCC progression through the modulation of lipid
metabolism remains largely unexplored.
In the present study, the roles of ELOVL2 in ccRCC
progression were explored by performing the transcriptional
profiling of primary ccRCC and normal kidney samples, revealing
that ELOVL2 is overexpressed in ccRCC and is overrepresented among
ELOVL isozymes. Of note, ELOVL2 was also overexpressed in ccRCC, as
well as in pRCC and chRCC, this being significantly associated with
a poor prognosis of patients with ccRCC or pRCC. Moreover, it was
demonstrated that ELOVL2 inhibition affects lipid metabolism and
suppresses cellular proliferation via the promotion of apoptosis,
at least in part through the disruption of endoplasmic reticulum
(ER) homeostasis in renal cancer cells. The results of the present
study suggested that ELOVL2 may be a potential novel therapeutic
target for RCC treatment.
Materials and methods
Cell lines and cultures
293T (RCB2202) and OS-RC-2 (RCB0735) cell lines were
purchased from RIKEN BioResource Center while 786-O and ACHN cell
lines were purchased from ATCC; SK-RC-52 cells were a kind gift
from Dr J.G. Old (Memorial Sloan Kettering Cancer Center). RPTEC
cell line, which is derived from epithelial cells of the human
renal proximal tubule, was purchased from Lonza Group, Ltd.
(CC-2553). The ACHN, 786-O, SK-RC-52 and OS-RC-2 cells were
cultured in RPMI-1640 medium supplemented with 10% fetal bovine
serum (FBS) at 37°C with a 5% humidified CO2 atmosphere.
The 293T cells were cultured in Dulbecco's modified Eagle's medium
supplemented with 10% fetal bovine serum (FBS) at 37°C with a 5%
humidified CO2 atmosphere. The RPTEC cells were cultured
in renal epithelial growth medium (REGM™) Bulletkit™ (CC-3190;
Lonza Bioscience) at 37°C with a 5% humidified CO2
atmosphere.
Patients and ccRCC samples
RCC tissues and adjacent normal kidney tissues from
46 patients who received radical or partial nephrectomies were
obtained from the University of Tsukuba Hospital, between 2006 and
2015 according to the protocols approved by the Ethics Committee of
the University of Tsukuba (approval no. H28-104). All patients
provided written informed consent prior to undergoing surgery.
Tumor stages were assigned according to the TNM staging of the
Union for International Cancer Control (16). Pathological grades were classified,
according to the four-tiered Fuhrman grading system (17). All patient characteristics are
summarized in Table SI.
RNA extraction and cDNA synthesis
Total RNA was extracted from the frozen tissues and
renal cancer cells using TRIzol® Reagent (Thermo Fisher
Scientific, Inc.). Following RNA purification, RNA was then reverse
transcribed into cDNA using a High-Capacity cDNA Reverse
Transcription kit (cat. no. 4368814; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Gene expression levels were quantified using a 7500
Fast Real-Time PCR machine with Fast SYBR-Green Master Mix (Thermo
Fisher Scientific, Inc.). The cycling conditions were as follows:
Initial hold at 95°C for 20 sec, 40 cycles at 95°C for 3 sec and
60°C for 30 sec, followed by a melting curve ranging from 95°C for
15 sec to 60°C for 1 min. Hypoxanthine phosphoribosyltransferase 1
(HPRT1) was used as an internal control. All primer sequences are
listed in Table SII. In order to
evaluate apoptosis-related genes, the relative expression levels of
the pro-apoptotic genes, Bcl-2-associated X protein (BAX), Bcl-2
homologous antagonist/killer (BAK),
phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1/NOXA) and
p53 upregulated modulator of apoptosis (BBC3/PUMA) and of the
anti-apoptotic gene BCL2 and induced myeloid leukemia cell
differentiation protein gene (MCL1) were analyzed. The relative
gene expression levels were quantified using the 2−ΔΔCq
method (18).
Western blot analysis
Western blot analysis was carried out as previously
described (19). Cells were lysed
in lysis buffer (20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM EDTA,
1% SDS and protease inhibitor) and sonicated on ice. The lysates
were centrifuged at 1,000 × g for 20 min at 4°C and the
supernatants were collected as samples. The protein quantification
of samples was performed using Quick Start Bradford Protein assay
(cat. no. 5000202JA; Bio-Rad Laboratories, Inc.). Samples were
subjected to 10% SDS-PAGE and separated products were subsequently
transferred to PVDF membranes. The membranes were blocked with ECL
Prime blocking agent (cat. no. RPN418V; Cytiva) for 60 min at room
temperature. The membranes were then incubated overnight at 4°C
with the following antibodies: Anti-serine/threonine-protein
kinase/endoribonuclease inositol-requiring enzyme 1α (anti-IRE1α;
1:200; #3294, Cell Signaling Technology, Inc.), anti-phosphorylated
IRE1α (anti-p-IRE1α; 1:200; NB100-2323, Novus Biologicals, LCC),
anti-C/EBP homologous protein (anti-CHOP; 1:200; MA1-250, Thermo
Fisher Scientific, Inc.). Anti-rabbit or anti-mouse immunoglobulin
G HRP-linked, whole donkey Ab (cat. no. NA934V, NA931VS; GE
Healthcare; Cytiva) were used at 1:10,000 as secondary antibodies.
The western blots were visualized with ImmunoStar® Zeta
(FUJIFILM Wako Pure Chemical Corporation) using a Fujifilm LAS-4000
imager and LAS400IR (FUJIFILM Wako Pure Chemical Corporation).
β-actin was used as the internal control (1:10,000, cat. no. A5316;
Sigma-Aldrich).
Bioinformatics analysis of gene
expression
Clinical and RNA-sequencing (RNA-seq) data of
primary tumors collected from patients with RCC in The Cancer
Genome Atlas (TCGA) database were downloaded from the Genomic Data
Commons (GDC) Data Portal (20). A
total of 1,005 (880 diseased and 125 control) samples were
examined, including 530 diseased and 71 control samples for the
kidney renal clear cell carcinoma cohort (KIRC; ccRCC), 286
diseased and 31 healthy samples for the cervical kidney renal
papillary cell carcinoma cohort (KIRP; pRCC), and 64 diseased and
23 healthy samples for the kidney chromophobe cohort (KICH;
chRCC).
ELOVL2 gene expression in the genitourinary cancer
samples was analyzed using the Gene Expression database of Normal
and Tumor tissues (GENT2). Gene expression data were downloaded
from the gene expression omnibus (GEO) public repository of the
U133 Plus 2 (GPL570) platform (http://gent2.appex.kr/gent2/) (21).
Plasmids and lentiviral
transduction
Small hairpin RNAs (shRNAs) for ELOVL2 characterized
in previous studies (22,23) were subcloned into the lentiviral
vector pLKO.1 (plasmid #8453; Addgene, Inc.). The oligonucleotide
sequences used in the construction of the shRNA vector are listed
in Table SIII. Lentiviruses were
generated in 293T cells by co-transfecting four plasmids in
trans, including the lentiviral vector (pLKO-shControl or
pLKO-shELOVL2), pMDLg/pRRE (plasmid #12251; Addgene, Inc.),
pRSV-Rev (plasmid #12253; Addgene, Inc.), and pMD2.G (plasmid
#12259; Addgene, Inc.) using Lipofectamine 2000®
transfection reagent (Thermo Fisher Scientific, Inc.). At 48 h
post-transfection, virus-containing supernatants were collected and
filtered through a 0.45-µm filter for infection. For viral
transductions, pLKO-shControl or pLKO-shELOVL2 lentiviruses were
incubated with the 786-O, ACHN and SK-RC-52 cells overnight at 37°C
in a humidified cell culture incubator. Cells were selected in the
presence of puromycin at 24 h post-infection. To stabilize
subclones, cells were cultured in medium with puromycin for 1 month
at 37°C in a humidified cell culture incubator and surviving pools
were utilized for express ion and proliferation analyses.
For overexpression experiments, the OSRC-2 cells
were transfected with pCMV6 empty vector (plasmid #PS100001;
Origene Technologies, Inc.) or pCMV6-ELOVL2 (plasmid #RC209232;
Origene Technologies, Inc.) at 37°C in a humidified cell culture
incubator, using Lipofectamine 3000® transfection
reagent (Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. The cells were used for the indicated
assays at 48 h post-transfection.
CRISPR/Cas9 design and cloning
The pX330-U6-Chimeric_BB-CBh-hSpCas9 (pX330) plasmid
was a gift from Feng Zhang (Addgene, Inc.; plasmid #42230)
(24). CRISPR single-guide (sg)
sequences for ELOVL2 were specifically targeted to exon 2 (sgELOVL2
#1) and exon 3 (sgELOVL2 #2 and #3) of ELOVL2 using the
CRISPR Design Tool (GE Healthcare Dharmacon, Inc.; http://dharmacon.horizondiscovery.com/gene-editing/crispr-cas9/crispr-design-tool/),
prior to cloning into pX330. The CRISPR guide sequence for
single-guide control (sgControl) was previously designed (25). The oligonucleotide sequences of the
single-guide RNAs (sgRNAs) are listed in Table SIII.
The ACHN cells were then seeded into 6-well plates
(1.4×105 cells/well) and co-transfected 2 h later with 2
µg of pX330-expresed sgRNAs and 0.2 µg pCI-neo vector (Promega
Corporation; cat. no. E1841) at 37°C in a humidified cell culture
incubator, using FuGENE HD (Promega Corporation; cat. no. E2312).
At 24 h post-transfection, 400 µg/ml G418 were applied for 7 to 10
days and the cells were allowed to recover for 2 or 3 days. When
the cells were approaching confluency, they were reseeded sparsely
in 10 cm dishes. After 2 or 3 weeks, discernible colonies were
isolated by using cloning discs (MilliporeSigma; cat. no.
Z374431-100EA). Individual clones were expanded and evaluated for
knockout status by Sanger sequencing for the target area.
Cellular proliferation assays
Cellular proliferation was assessed using MTT assay
with a Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies,
Inc.) according to the manufacturer's instructions. Briefly, the
cells were seeded into 96-well plates and 10 µl of WST-8 were added
after 72 h. OD460 was measured following a 1-h incubation at
37°C.
Subcutaneous xenografting
BALB/c nude (nu/nu) female mice (n=12; 6–8 weeks
old) were purchased from Charles River Laboratories, Inc.. The mice
were housed under specific pathogen-free conditions, under a 12-h
light/dark cycle, with ad libitum access to food and water.
For subcutaneous xenograft assays, 1×107 cells suspended
in 100 µl PBS were injected subcutaneously into the right flank by
using a 24G needle and the tumor volumes were measured once a week.
Tumor volume was calculated by the formula: mm3 = length
× width × height × 0.52 (26).
After 90 days, all animals were sacrificed, and the xenograft
tumors were excised. Animals were anesthetized with 2% isoflurane
before being euthanized by cervical dislocation.
Formalin-fixed and paraffin-embedded (FFPE)
specimens of the xenograft tumors were cut into 4-µm-thick
sections, prior to deparaffinization and rehydration. For antigen
retrieval, the sections were pretreated by microwave for 21 min in
a citric acid buffer. After the antigen retrieval procedure,
endogenous peroxidase activity was blocked with 3%
H2O2 for 25 min and the slides were incubated
with Ki-67 antibody (1:200; Dako; Agilent Technologies, Inc.;
M7240) at 4°C overnight. The immunohistochemical reaction was
visualized using the secondary antibody Histofine Simple Stain MAX
PO® (Nichirei Biosciences, Inc,; cat. no. 424151) by
using diaminobenzidine as the chromogen.
All animal studies were approved by the Animal
Experiment Committee of the University of Tsukuba and all
experiments were performed in accordance with the guidelines of the
University of Tsukuba's Regulations of Animal Experiments (approval
no. 20–370).
Apoptosis assays
Apoptosis was assessed using the
Caspase-Glo® 3/7 Assay System (Promega Corporation; cat.
no. G8090), Annexin-V-FLUOS staining kit (Roche Diagnostics; cat.
no. 11858777001), and JC-1 Mitochondrial Membrane Potential Assay
kit (Cayman Chemical Company; cat. no. 10009172), according to the
manufacturer's instructions.
Staining of LDs
Cells were seeded on glass coverslips in 60mm dishes
and cultured with oleic acid (200 µM)-containing medium at 37°C
overnight in a 5% CO2 incubator. The medium was then
aspirated, and the cells were washed twice with PBS before fixation
with 4% formaldehyde (FUJIFILM Wako Pure Chemical Corporation) for
5 min at room temperature. The cells were then washed twice with
PBS, and incubated with 0.5 µM Lipi-Green (Dojindo Molecular
Technologies, Inc.) for 30 min in the dark at 37°C. Thereafter, the
cells were washed twice with PBS and the coverslips were mounted on
glass slides using VECTASHIELD® Antifade Mounting Medium
with DAPI (Vector Laboratories, Inc.; cat. no. H-1200). Images of
the stained cells were acquired with a BZ-X710 fluorescence
microscope (Keyence Corporation).
Live cells were incubated with 0.5 µM Lipi-Green in
Hanks' Balanced Salt Solution (HBSS) for 30 min before washing
twice in HBSS, resuspended in 1 mM EDTA/PBS (pH 8.0) with 0.5% BSA,
and passed through a cell strainer. Cells were analyzed on a
Gallios flow cytometer (Beckman-Coulter, Inc.) and data were
analyzed using FlowJo v10 software (FlowJo LLC).
ER tracker staining
The ACHN cells (ACHN/sgControl, ACHN/sgELOVL2-1 and
ACHN/sgELOVL2-2) were seeded on glass coverslips in 60-mm dishes
and cultured at 37°C for 48 h in a 5 % CO2 incubator.
The medium was then aspirated, and the cells were washed twice with
HBSS before fixation with 4% formaldehyde (FUJIFILM Wako Pure
Chemical Corporation) for 5 min at room temperature. The cells were
then washed twice with HBSS and incubated with 500 nM ER Tracker
Red (Thermo Fisher Scientific, Inc.; cat. no. E34250) for 2 h in
the dark at 37°C. Subsequently, the cells were washed twice with
PBS and the coverslips were mounted on glass slides. Images of the
stained cells were acquired with a BZ-X710 fluorescence microscope
(Keyence Corporation).
Live cells were incubated with 500 nM ER Tracker in
HBSS for 30 min before washing twice in HBSS, re-suspended in 1 mM
EDTA/PBS (pH 8.0) with 0.5% BSA, and passed through a cell
strainer. Cells were analyzed on a Gallios flow cytometer and data
were analyzed using FlowJo v10 software.
Gas chromatography-mass spectrometry
(GC-MS)
The extraction of total lipids from cell pellets was
performed as previously described (27). The hydrolysis and derivatization of
the extracts to FA methyl esters (FAMEs) was performed. For the
internal standards of conjugated FAs, C20:4 (ω-6), C20:5 (ω-3),
C22:5 (ω-6), C22:5 (ω-3) and C22:6 (ω-3) FAMEs were purchased from
MilliporeSigma or Cayman Chemical Company.
GC-MS analysis was performed using a GCMS-TQ8040
(Shimadzu Corporation) with an Omegawax® Capillary GC
Column (MilliporeSigma; cat. no. 24136). The temperature gradient
was initially increased from 70 to 150°C at a rate of 20°C per min
and from 150 to 280°C at a rate of 8°C per min before decreasing
from 280 to 200°C at a rate of 15°C per min. Sample injection was
performed in splitless mode with 1.0 µl of sample injected. For MS
analysis, the ion source and interface temperature were set to 200°
and 270°C, respectively. The data were acquired in full scan mode
(30–600 m/z) under 70 eV of ionization voltage. FAMEs were
identified according to retention time and electron ionization-MS
(EI-MS) spectrum matching with reference FAME standards. The
relative amount of FAMEs was calculated by comparing average peak
area per sample mass. Each sample was independently measured three
times.
Statistical analysis
Data are expressed as the mean ± SD. All statistical
analyses were performed using JMP 10 software (SAS Institute,
Inc.), GraphPad Prism8 (GraphPad Software, Inc.) or R package
(version 4.0.2; RStudio, Inc.). The significance of the differences
between the two groups was assessed using the unpaired Student's
t-test or Mann-Whitney test. The significance of the differences
between the three or more groups was assessed using one-way ANOVA
with post hoc comparisons using Dunnett's test or Kruskal-Wallis
test followed by a Dunn's post hoc test with Bonferroni correction.
Survival curves were constructed using the Kaplan-Meier method and
the difference between the curves was evaluated using the log-rank
test. Patients were divided into two groups, a low expression or
high expression group, using the cutoff of median expression
values. P<0.05 was considered to indicate a statistically
significant difference.
Results
ELOVL2 is highly overexpressed in
RCC
The abundance of seven ELOVL isozymes in
corresponding normal and tumor tissues was first examined in the
present cohort, revealing that the ELOVL1, ELOVL2, ELOVL5 and
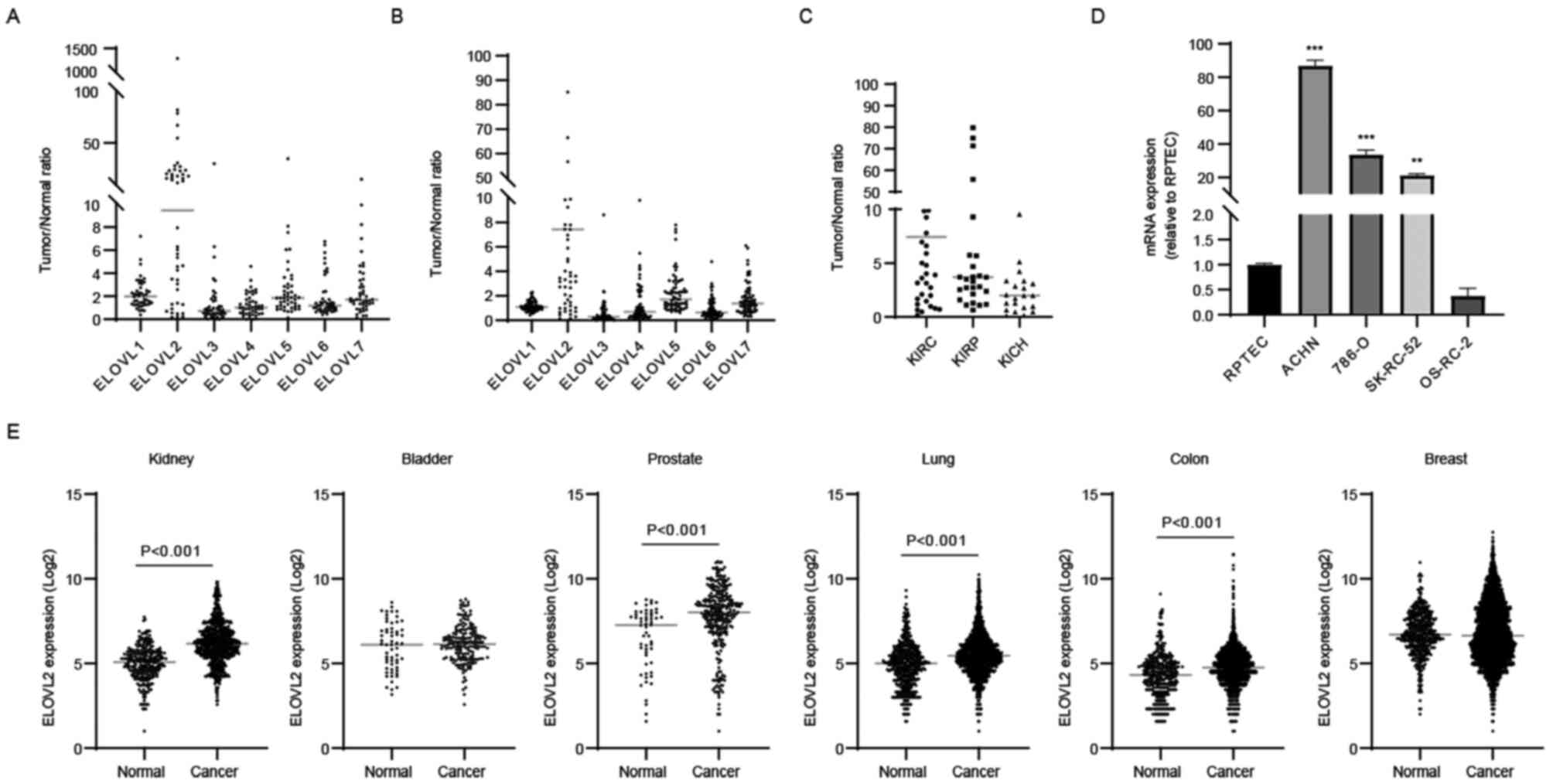
ELOVL7 expression levels were elevated in ccRCC tissues (Fig. 1A). To validate these results, ELOVL
isozyme gene expression in corresponding normal and tumor tissues
was examined using TCGA database (KIRC cohort). It was confirmed
that ELOVL2 mRNA expression was most highly and significantly
elevated in ccRCC tissues (Fig.
1B; P<0.0001). Subsequently, ELOVL2 mRNA expression was
investigated further among three major histological subtypes of RCC
using TCGA database (KIRC, KIRP and KICH cohorts). This revealed
that ELOVL2 was overexpressed in the pRCC and chRCC tissues;
however, its expression in ccRCC tissues was the highest among the
histological subtypes (Fig. 1C).
Moreover, the ELOVL2 mRNA expression levels were markedly elevated
in the ACHN, 786-O and SK-RC-52 cell lines, although the mRNA
expression levels in OS-RC-2 cell line were lower as compared with
those in the RPTEC cell line (Fig.
1D).
Subsequently, the cancer-associated alteration of
ELOVL2 gene expression in various types of cancer was investigated
using the GENT2 database (Fig.
1E). Compared with normal tissues, ELOVL2 expression was found
to be significantly higher in kidney, prostate, lung and colon
cancers, whereas ELOVL2 expression was similar in bladder or breast
cancers. These results suggested that the roles of ELOVL isozymes
differed by cancer type and that ELOVL2 overexpression may be
associated with the carcinogenesis or disease progression of RCC,
particularly ccRCC.
ELOVL2 overexpression is associated
with the progression of RCC
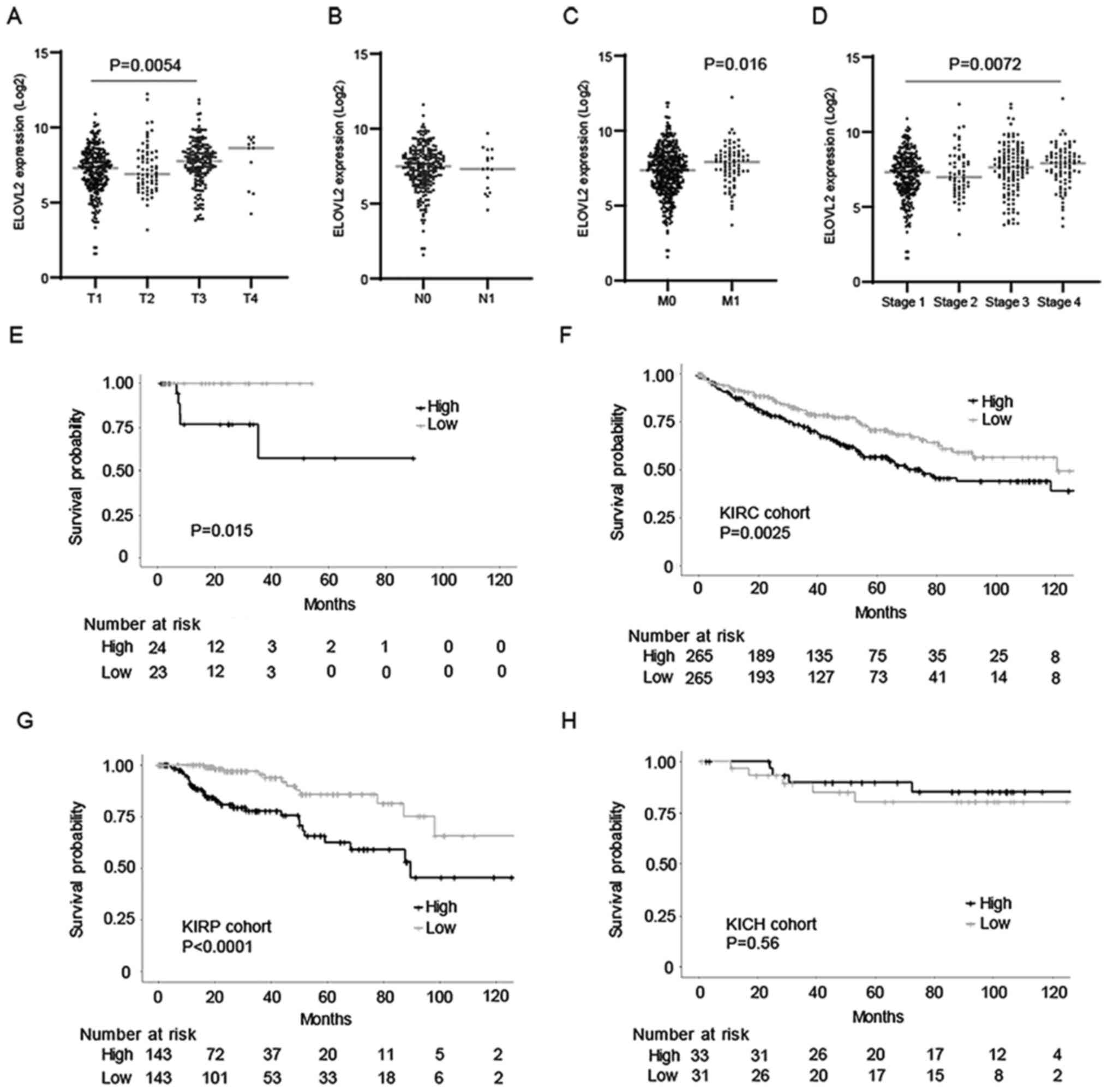
The association between the gene expression of
ELOVL2 and tumor-node-metastasis (TNM) stages in the KIRC cohort
was examined, in order to clarify the clinical significance of
ELOVL2 in ccRCC. ELOVL2 expression was higher in locally advanced
and metastatic disease, although its expression was not associated
with the lymph node metastasis status (Fig. 2A-C). Collectively, ELOVL2
expression was increased according to clinical stage (Fig. 2D).
Subsequently, the association between the expression
of ELOVL2 and the prognosis of patients with RCC was evaluated, in
order to elucidate the clinical impact of ccRCC. According to
Kaplan-Meier analysis, it was revealed that the high expression of
ELOVL2 was significantly associated with a poor prognosis in the
present cohort (P=0.0015, Fig.
2E). To validate these results, the association between the
expression of ELOVL2 and the prognosis of patients with ccRCC was
examined further in the KIRC cohort and it was demonstrated that,
similarly, the high expression of ELOVL2 was significantly
associated with a poor prognosis (P<0,01, Fig. 2F). Moreover, a high expression of
ELOVL2 was significantly associated with a poor prognosis of
patients with pRCC (P<0.0001, Fig.
2G). However, this was not observed in patients with chRCC
(Fig. 2H). In total, the
aforementioned results indicated that ELOVL2 may mediate ccRCC and
pRCC disease progression, at least partially.
ELOVL2 promotes the proliferation of
renal cancer cells
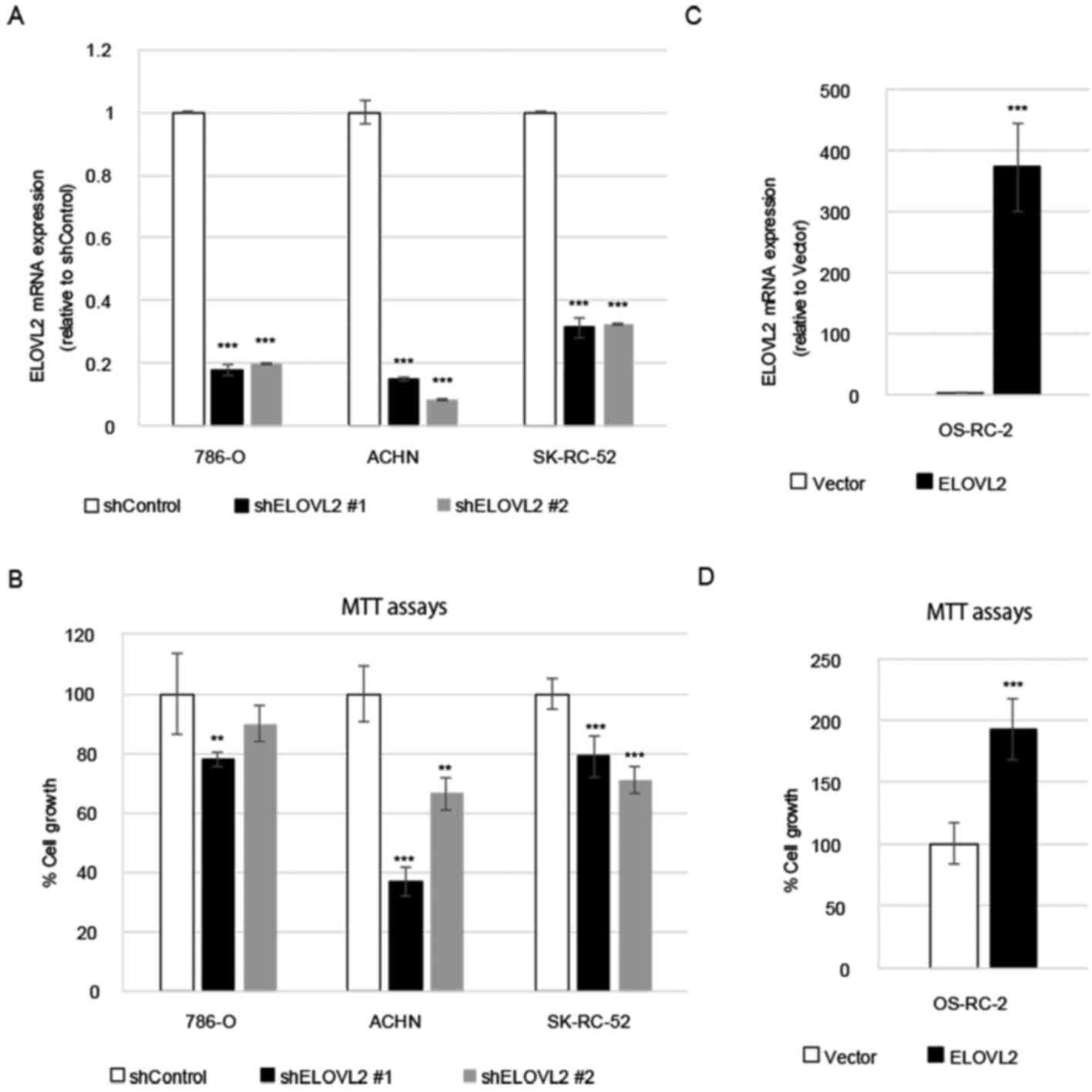
To assess the function of ELOVL2 in the
proliferation of RCC cells, the shRNA knockdown of ELOVL2 in 786-O,
ACHN and SK-RC-52 cells was performed. The effects of each shRNA
were evaluated using RT-qPCR and a significant decrease in ELOVL2
expression was confirmed (Fig.
3A). MTT assays were then performed to examine the effects of
ELOVL2 knockdown on cellular proliferation. It was observed that
the knockdown of ELOVL2 inhibited the proliferation of all cells
compared with the cells transfected with control shRNA (Fig. 3B).
By contrast, ELOVL2 overexpression was induced in
the OS-RC-2 cells, a cell line with a lower basal expression of
ELOVL2. The transfection efficacy was evaluated using RT-qPCR and
significantly increased expression of ELOVL2 was confirmed
(Fig. 3C). MTT assays revealed
that the proliferation of ELOVL2-overexpressing cells was
significantly promoted, in comparison with the control group
(Fig. 3D), indicating that ELOVL2
may be a promoter of the proliferation of renal cancer cells.
ELOVL2 ablation suppresses tumor
growth in vivo, the synthesis of PUFAs and the production of LDs in
renal cancer cells
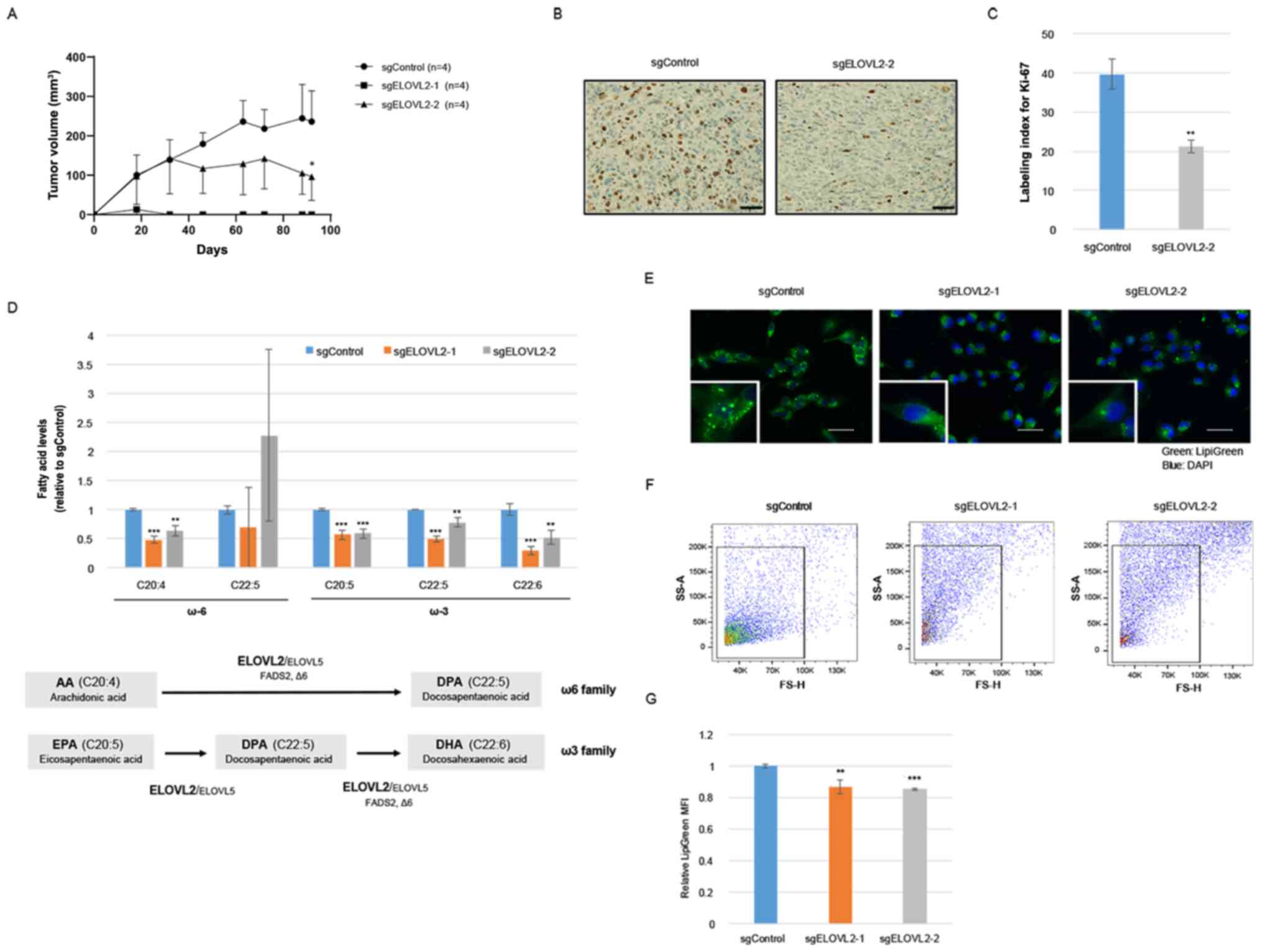
To further clarify the roles of ELOVL2 in RCC
progression, an ELOVL2-knockout ACHN cell line was established, by
using a CRISPR/Cas9 system (sgELOVL2-1 and sgELOVL2-2) (Fig. S1). In vitro, cellular
proliferation was significantly decreased by ELVOVL2 ablation in
the ACHN cells (Fig. S1). More
importantly, the CRISPR/Cas9-mediated ablation of ELOVL2 suppressed
tumor growth when the cells were implanted subcutaneously into mice
(Fig. 4A). The maximum volumes of
the subcutaneous xenograft tumors in the ACHN/sgControl and
ACHN/sgELOVL2-2 at the day of sacrifice were 241 and 109
mm3, respectively. Of note, all xenograft tumors in the
ACHN/sgELOVL2-1 group spontaneously regressed at 14 days following
transplantation and were not detected at the time of extirpation.
The Ki-67 index was also examined in the xenograft tumors
transfected with sgControl or sgELOVL2 and it was observed that the
ACHN/sgELOVL2-2 cells exhibited a significantly lower Ki-67 index
than the ACHN/sgControl cells (Fig. 4B
and C). These results further supported the possibility that
ELOVL2 augments tumor growth in vivo.
As ELOVL2 is located on chromosome 6p24.2 and
encodes for an endoplasmic reticulum transmembrane protein
controlling the elongation of C20–C24 PUFAs (7), total FA levels were then evaluated in
the ACHN/sgControl and ACHN/sgELOVL2 cells using GC-MS analysis
(Fig. S2). The alteration of
LC-PUFA levels is demonstrated in Fig.
4D. ELOVL2 is an essential enzyme in the endogenous production
of docosahexaenoic acid (DHA, C22:6 n-3) (28,29)
and, as expected, the DHA levels were significantly decreased by
the ablation of ELOVL2 in the renal cancer cells. Moreover, the
amounts of other LC-PUFAs species, including arachidonic acid (AA,
C20:4 n-6), eicosapentaenoic acid (EPA) and docosapentaenoic acid
(DPA), were also significantly decreased. On the other hand, the
production of DPA was not consistently altered in the ACHN/sgELOVL2
cells. The amount of DPA in the ACHN cells was very low and was not
detected in several samples (Fig.
S2).
Since previous studies have demonstrated that ELOVL2
promotes the accumulation of LDs through the production of DHA
(11,12), the storage of LDs was further
assessed using neutral lipid staining. Of note, the abundance of
LDs in the ACHN/sgELOVL2 cells was significantly decreased compared
with the ACHN/sgControl cells (Fig.
4E-4G), suggesting that the alteration of lipid metabolism by
ELOVL2 overexpression may affect RCC cell proliferation.
ELOVL2 ablation induces the apoptosis
of renal cancer cells
Previous studies have revealed that the production
of intracellular LDs promotes apoptosis under stressful tumor
microenvironments (4). Therefore,
in the present study, apoptosis was evaluated in ACHN/sgControl or
ACHN/sgELOVL2 cells and a significantly elevated activity of
caspase 3/7 was detected in the ACHN/sgELOVL2 cells (Fig. 5A). Similarly, a greater number of
apoptotic ACHN/sgELOVL2 cells compared with ACHN/sgControl cells
was identified, as evidenced by Annexin V/PI staining (Fig. 5B). Depending on cellular stress,
intrinsic apoptosis (30) leads to
the loss of mitochondrial membrane potential; thus, the present
study further assessed the mitochondrial transmembrane electric
potential of ACHN/sgControl or ACHN/sgELOVL2 cells using
fluorescent JC-1. The aggregate/monomer ratio was significantly
decreased in the ACHN/sgELOVL2 cells (Fig. 5C), indicating the promotion of
mitochondrial transmembrane polarization and the activation of the
intrinsic apoptotic pathway by ELOVL2 ablation. More precisely,
pro-apoptotic gene (BAX, BAK, PUMA and NOXA) expression levels were
significantly increased, while anti-apoptotic gene (BCL2 and MCL1)
expression levels were significantly decreased due to ELOVL2
ablation (Fig. 5D). These results
indicated that ELOVL2 contributes to cellular survival through the
inhibition of theapoptosis of renal cancer cells.
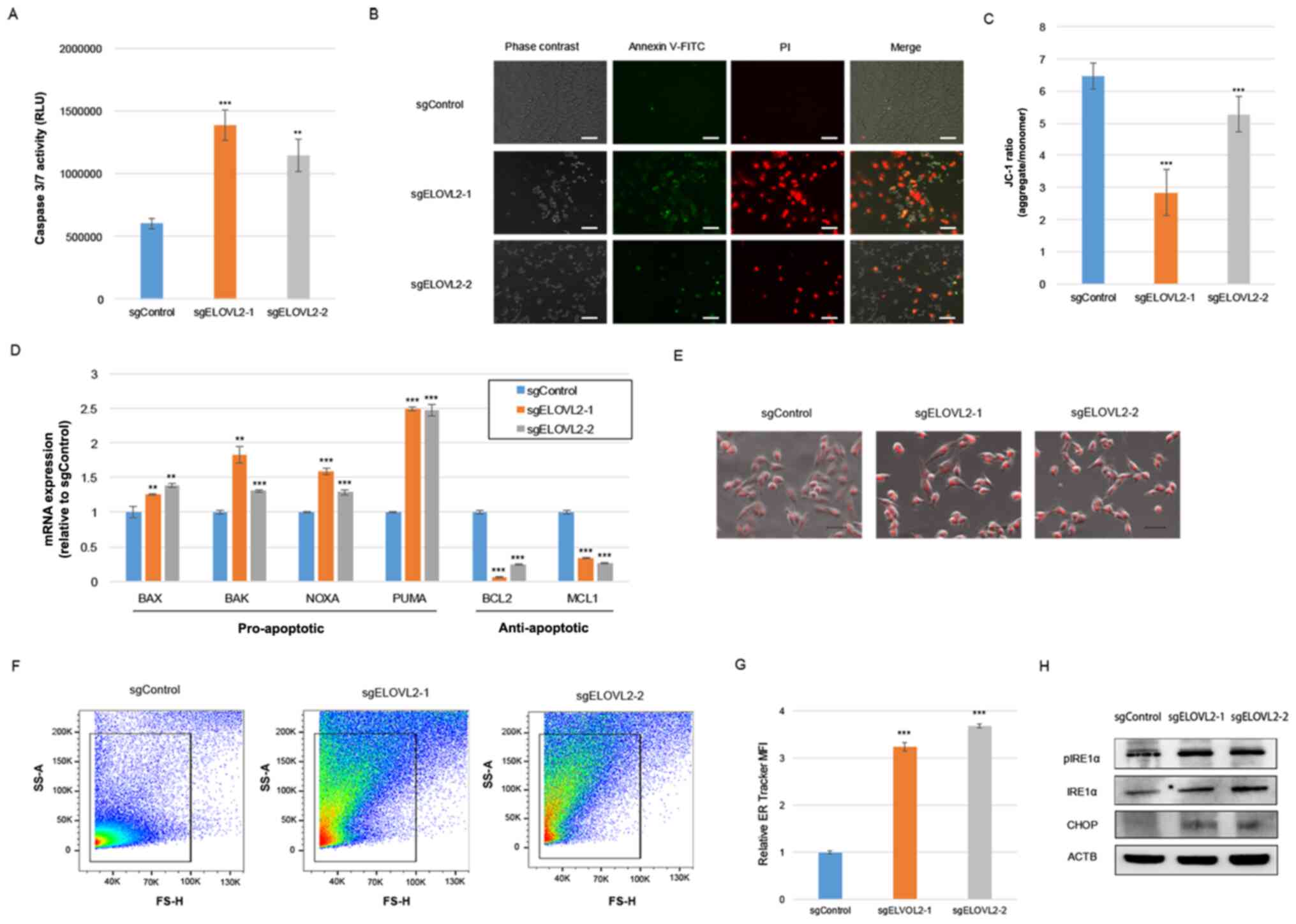 | Figure 5.CRISPR/Cas9-mediated knockdown of
ELOVL2 promotes the loss of mitochondrial transmembrane potential
and apoptosis in ACHN cells. (A) Analysis of cell apoptosis by
caspase 3/7 assay in ACHN/sgControl or ACHN/sgELOVL2 cells. (B)
Representative images of Annexin V-FITC staining in ACHN/sgControl
or ACHN/sgELOVL2 cells. Scale bar, 100 µm. (C) Aggregates/monomers
relative ratio evaluation in ACHN/sgControl or ACHN/sgELOVL2 cells
by using JC-1 assay. (D) Apoptosis-related gene expression levels
in ACHN/sgControl or ACHN/sgELOVL2 cells. (E) Representative images
of ER tracker staining in ACHN/sgControl or ACHN/sgELOVL2 cells.
Scale bar, 50 µm. (F) Representative flow cytometry dot plots of ER
tracker staining in ACHN/sgControl or ACHN/sgELOVL2 cells. (G) ER
tracker fluorescence quantification in ACHN/sgControl or
ACHN/sgELOVL2 cells. (H) Western blot analysis of pIRE1α, IRE1α,
and CHOP in ACHN/sgControl or ACHN/sgELOVL2 cells. P-values were
assessed using ANOVA with post hoc comparisons using Dunnett's test
(A, C, D and G). **P<0.01 and ***P<0.001, vs. control. ELOVL,
very-long-chain fatty acid; sg, single guided; IRE1α,
serine/threonine-protein kinase/endoribonuclease inositol-requiring
enzyme 1α; pIRE1α, phosphorylated-IRE1α; CHOP, C/EBP homologous
protein. |
The degradation of lipid storage capacity in LD form
can lead to the collapse of ER homeostasis, triggering the unfolded
protein response (UPR) and cellular apoptosis (4,5).
Therefore, in order to support the hypothesis of the involvement of
ELOVL2 expression in ER homeostasis, ER tracker staining was
performed in ACHN/sgControl or ACHN/sgELOVL2 cells. Subsequent ER
imaging (Fig. 5E) and
quantification using flow cytometry (Fig. 5F and G) indicated ER expansion in
the ACHN/sgELOVL2 cells, due to ER stress. Additionally, ELOVL2
ablation promoted the phosphorylation of IRE1α, an activator of UPR
sensors (Fig. 5H), while the
expression of CHOP, which plays a main role in ER stress-induced
apoptosis, was upregulated by ELOVL2 ablation (Fig. 5H). Collectively, these results
demonstrated that ELOVL2 mediated-lipid metabolism suppresses cell
apoptosis in renal cancer cells and suggested that disruption of ER
homeostasis may be a potential mechanism of induction.
Discussion
The present study demonstrated that ELVOL2 may alter
lipid metabolism and promote tumor growth via the inhibition of the
apoptosis of renal cancer cells. Moreover, it was observed that
ELOVL2 expression in RCC tissues was elevated and that the higher
expression of ELOVL2 was significantly associated with a poor
prognosis of patients with RCC.
There are limited studies available on the roles of
ELOVL2 in cancer progression (22,31,32)
and their results are controversial. Gimple et al (22) demonstrated that ELOVL2 promoted
LC-PUFA synthesis, supporting efficient EGFR signaling and the
proliferation of glioblastoma stem cells. By contrast, Kang et
al (31) reported that ELOVL2
ablation may promote cell migration and the colony formation of
breast cancer cells and revealed that a decreased ELOVL2 expression
was associated with a poorer prognosis of patients with breast
cancer. Furthermore, Ding et al (32) reported that ELOVL2 suppresses the
proliferation of neuroblastoma cells and was associated with
favorable survival rates. According to the conflicting findings
reported in previous studies, a multi-role function for ELOVL2 in
cancer progression has been demonstrated, that clearly varies by
cancer type.
An altered lipid metabolism greatly affects cancer
progression, an effect observed in the results of the present
study, being in line with research depicting the role of ELOVL2 as
a primary controller of the elongation process of LC-PUFAs and a
critical enzyme in DHA biosynthesis (7). In the present study, it was
demonstrated that endogenous LC-PUFAs, including AA and DHA, were
decreased due to ELOVL2 ablation in renal cancer cells. In relation
to this, it has been previously reported that the inhibition of the
AA pathway by lipoxygenase (LOX) inhibitors induces the apoptosis
and reduces the viability of renal cancer cells in vitro
(33,34). Although the ELOVL2 overexpression
results of the present study are in line with these mechanistic
findings, it has been previously reported, however, that DHA
administration may inhibit the proliferation and invasion of renal
cancer cells in vitro (35,36).
However, LC-PUFAs are known to have distinct and contrasting
effects on cancer progression. Therefore, the alteration of a
delicate balance among a variety of LC-PUFAs, due to the inhibition
of ELOVL2 may be associated with the differing phenotypes observed
in various cancer types.
Previous studies have demonstrated that LDs are
increased through the endogenous production of DHA by ELOVL2 in
mice (11,12). Similarly, it was revealed that
ELOVL2 ablation may suppress the production of DHA and LDs in renal
cancer cells, suggesting that ELOVL2 overexpression may promote LD
production through endogenous DHA production in RCC. Cancer cells
are often found in harsher environmental (macro and micro)
conditions, including hypoxia and nutrient deprivation, yet still
rapidly grow under such severe stressors. The functions of LDs
under conditions of cellular stress, such as the maintenance of
energy and redox homeostasis, the regulation of autophagy, the
maintenance of ER homeostasis and protection against lipotoxicity,
have been demonstrated (4).
Ackerman et al (6) revealed that LDs prevent the
accumulation of toxic, saturated lipids during hypoxia by buffering
of cellular lipid saturation through exchange of TG-resident
unsaturated FAs in ccRCC. Cellular lipotoxicity due to saturated
FAs (SFA), including palmitate (C16:0), may cause apoptosis,
leading to cancer cell death (37,38).
Recently, ELOVL2 was designated as a critical pro-survival enzyme,
assisting in the prevention of glucolipotoxicity-induced β-cell
apoptosis (39). Of note,
ELOVL2/DHA axis-modified lipid partitioning results in a non-toxic
utilization of SFA palmitate by favoring its transport into
mitochondria for FA oxidation (FAO) through a CPT1-dependent
mechanism. Additionally, Balaban et al demonstrated that
C4-2B prostate cancer cells and MCF-7 breast cancer cells are
protected from palmitate-induced apoptosis by mitochondrial FAO
(37,38). During palmitate-induced
lipotoxicity, cellular levels of anti-apoptotic proteins, including
BCL-2 or MCL-1, have been reported to be decreased (40,41)
while levels of pro-apoptotic proteins, including PUMA or NOXA,
have been reported to be increased (42,43).
It was revealed in the present study that ELOVL2 ablation may
promote renal cancer cell apoptosis through the alteration of pro-
and anti-apoptotic genes in a similar manner. These findings
suggest that ELOVL2 may protect cells against lipotoxicity-driven
apoptosis to promote tumor growth in RCC.
Additionally, it was demonstrated that ELOVL2
ablation in RCC may promote ER stress and CHOP upregulation,
downregulating BCL-2 and MCL-1, while upregulating BAK and BAX, via
a mitochondria-dependent pathway (44). Indeed, the results of the present
study demonstrated that pro- and anti-apoptotic genes were
similarly altered by ELOVL2 ablation. In line with the findings in
the study by Qui et al (5),
who reported that HIF2α/PLIN2-dependent LDs promote resistance
against ER stress and cell survival in ccRCC, these collective
findings are in support of ER stress promoting cellular apoptosis
by ELOVL2 ablation in renal cancer cells, decreasing LDs and
removing the cellular buffer against toxic lipids.
In conclusion, the present study demonstrated for
the first time, to the best of our knowledge, that ELOVL2 promotes
tumor growth by the inhibition of apoptosis via the elongation of
PUFAs, at least partly by maintaining ER homeostasis in renal
cancer cells. Taken together, it was suggested that ELOVL2-induced
LDs may contribute to cancer progression due to multiple protective
effects against cellular stress in RCC. Moreover, it was suggested
that ELOVL2 may be an attractive potential therapeutic target for
patients with RCC.
Supplementary Material
Supporting Data
Acknowledgements
The authors are grateful for the skillful technical
assistance of Mr. Minoru Suzuki, Mrs. Naoko Ueki, Mrs. Noriko
Kunita (University of Tsukuba).
Funding
The present study was supported by the Japan Society for the
Promotion of Science KAKENHI grant (nos. 17K16774 and 19K09664) and
the COI-NEXT grant (no. JPMJPF2017).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SK, TK and TMa contributed to the conceptualization
of the present study. HNi, HS and TAS contributed to the design of
the present study. SS, KeT, KoT, MS, YN, SN, MW and TMi contributed
to performing the experiments and the data analyses. KeT, SK,YN and
BJM wrote the original draft. KeT, SK, TK, HNe, BJM and TS
contributed to the interpretation of the results in the present
study. KeT and SK confirmed the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All protocols have been approved by the Ethics
Committee of the University of Tsukuba (approval no. H28-104). All
patients provided written informed consent prior to undergoing
surgery. All animal studies were approved by the Animal Experiment
Committee of the University of Tsukuba and all experiments were
performed in accordance with the guidelines of the University of
Tsukuba's Regulations of Animal Experiments (approval no.
20–370).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hsieh JJ, Purdue MP, Signoretti S, Swanton
C, Albiges L, Schmidinger M, Heng DY, Larkin J and Ficarra V: Renal
cell carcinoma. Nat Rev Dis Primers. 3:170092017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rini BI, Battle D, Figlin RA, George DJ,
Hammers H, Hutson T, Jonasch E, Joseph RW, McDermott DF, Motzer RJ,
et al: The society for immunotherapy of cancer consensus statement
on immunotherapy for the treatment of advanced renal cell carcinoma
(RCC). J Immunother Cancer. 7:3542019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Walther TC and Farese RV Jr: Lipid
droplets and cellular lipid metabolism. Annu Rev Biochem.
81:687–714. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Petan T, Jarc E and Jusović M: Lipid
Droplets in Cancer: Guardians of Fat in a Stressful World.
Molecules. 23:19412018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qiu B, Ackerman D, Sanchez DJ, Li B,
Ochocki JD, Grazioli A, Bobrovnikova-Marjon E, Diehl JA, Keith B
and Simon MC: HIF2α-Dependent Lipid Storage Promotes Endoplasmic
Reticulum Homeostasis in Clear-Cell Renal Cell Carcinoma. Cancer
Discov. 5:652–667. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ackerman D, Tumanov S, Qiu B,
Michalopoulou E, Spata M, Azzam A, Xie H, Simon MC and Kamphorst
JJ: Triglycerides promote lipid homeostasis during hypoxic stress
by balancing fatty acid saturation. Cell Rep. 24:2596–2605.e5.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guillou H, Zadravec D, Martin PG and
Jacobsson A: The key roles of elongases and desaturases in
mammalian fatty acid metabolism: Insights from transgenic mice.
Prog Lipid Res. 49:186–199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
von Roemeling CA, Marlow LA, Wei JJ,
Cooper SJ, Caulfield TR, Wu K, Tan WW, Tun HW and Copland JA:
Stearoyl-CoA desaturase 1 is a novel molecular therapeutic target
for clear cell renal cell carcinoma. Clin Cancer Res. 19:2368–2380.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lucarelli G, Ferro M, Loizzo D, Bianchi C,
Terracciano D, Cantiello F, Bell LN, Battaglia S, Porta C, Gernone
A, et al: Integration of Lipidomics and Transcriptomics Reveals
Reprogramming of the Lipid Metabolism and Composition in Clear Cell
Renal Cell Carcinoma. Metabolites. 10:5092020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang H, Zhang X, Liu F, Fan J, Wang B and
Dong C: SREBP1-driven lipid desaturation supports clear cell renal
cell carcinoma growth through regulation of NF-κB signaling.
Biochem Biophys Res Commun. 495:1383–1388. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pauter AM, Olsson P, Asadi A, Herslöf B,
Csikasz RI, Zadravec D and Jacobsson A: Elovl2 ablation
demonstrates that systemic DHA is endogenously produced and is
essential for lipid homeostasis in mice. J Lipid Res. 55:718–728.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi T, Zadravec D and Jacobsson A:
ELOVL2 overexpression enhances triacylglycerol synthesis in 3T3-L1
and F442A cells. FEBS Lett. 581:3157–3163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim K, Han C, Xu L, Isse K, Demetris AJ
and Wu T: Cyclooxygenase-2-derived prostaglandin E2 activates
beta-catenin in human cholangiocarcinoma cells: Evidence for
inhibition of these signaling pathways by omega 3 polyunsaturated
fatty acids. Cancer Res. 68:553–560. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yao QH, Zhang XC, Fu T, Gu JZ, Wang L,
Wang Y, Lai YB, Wang YQ and Guo Y: ω-3 polyunsaturated fatty acids
inhibit the proliferation of the lung adenocarcinoma cell line A549
in vitro. Mol Med Rep. 9:401–406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Serhan CN, Hong S, Gronert K, Colgan SP,
Devchand PR, Mirick G and Moussignac RL: Resolvins: A family of
bioactive products of omega-3 fatty acid transformation circuits
initiated by aspirin treatment that counter proinflammation
signals. J Exp Med. 196:1025–1037. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sobin LH, Gospodarowicz MK and Wittekind
CH: TNM Classification of Malignant Tumors. (7th edition).
Wiley-Blackwell. (Chichester, West Sussex). 2009.
|
|
17
|
Fuhrman SA, Lasky LC and Limas C:
Prognostic significance of morphologic parameters in renal cell
carcinoma. Am J Surg Pathol. 6:655–663. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kandori S, Kojima T, Matsuoka T, Yoshino
T, Sugiyama A, Nakamura E, Shimazui T, Funakoshi Y, Kanaho Y and
Nishiyama H: Phospholipase D2 promotes disease progression of renal
cell carcinoma through the induction of angiogenin. Cancer Sci.
109:1865–1875. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Grossman RL, Heath AP, Ferretti V, Varmus
HE, Lowy DR, Kibbe WA and Staudt LM: Toward a Shared Vision for
Cancer Genomic Data. N Engl J Med. 375:1109–1112. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park SJ, Yoon BH, Kim SK and Kim SY:
GENT2: An updated gene expression database for normal and tumor
tissues. BMC Med Genomics. 12 (Suppl 5):1012019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gimple RC, Kidwell RL, Kim LJY, Sun T,
Gromovsky AD, Wu Q, Wolf M, Lv D, Bhargava S, Jiang L, et al:
Glioma Stem Cell-Specific Superenhancer Promotes Polyunsaturated
Fatty-Acid Synthesis to Support EGFR Signaling. Cancer Discov.
9:1248–1267. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Purdy JG, Shenk T and Rabinowitz JD: Fatty
acid elongase 7 catalyzes lipidome remodeling essential for human
cytomegalovirus replication. Cell Rep. 10:1375–1385. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cong L, Ran FA, Cox D, Lin S, Barretto R,
Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA, et al: Multiplex
genome engineering using CRISPR/Cas systems. Science. 339:819–823.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Miyamoto T, Lo PHY, Saichi N, Ueda K,
Hirata M, Tanikawa C and Matsuda K: Argininosuccinate synthase 1 is
an intrinsic Akt repressor transactivated by p53. Sci Adv.
3:e16032042017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mishra A, Zennami K, Velarde E, Thorek
DLJ, Yegnasubramanian S, DeWeese TL and Lupold SE: Longitudinal
measurement of subcutaneous and intratibial human prostate cancer
xenograft growth and response to ionizing radiation by plasma Alu
and LINE-1 ctDNA: A comparison to standard methods. Prostate.
81:745–753. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Folch J, Lees M and Sloane Stanley GH: A
simple method for the isolation and purification of total lipides
from animal tissues. J Biol Chem. 226:497–509. 1957. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gregory MK and James MJ: Rainbow trout
(Oncorhynchus mykiss) Elovl5 and Elovl2 differ in
selectivity for elongation of omega-3 docosapentaenoic acid.
Biochim Biophys Acta 1656–60. 20141841.PubMed/NCBI
|
|
29
|
Gregory MK and James MJ: Functional
characterization of the duck and turkey fatty acyl elongase enzymes
ELOVL5 and ELOVL2. J Nutr. 144:1234–1239. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matsuura K, Canfield K, Feng W and
Kurokawa M: Metabolic Regulation of Apoptosis in Cancer. Int Rev
Cell Mol Biol. 327:43–87. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kang YP, Yoon JH, Long NP, Koo GB, Noh HJ,
Oh SJ, Lee SB, Kim HM, Hong JY, Lee WJ, et al: Spheroid-Induced
Epithelial-Mesenchymal Transition Provokes Global Alterations of
Breast Cancer Lipidome: A Multi-Layered Omics Analysis. Front
Oncol. 9:1452019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ding Y, Yang J, Ma Y, Yao T, Chen X, Ge S,
Wang L and Fan X: MYCN and PRC1 cooperatively repress
docosahexaenoic acid synthesis in neuroblastoma via ELOVL2. J Exp
Clin Cancer Res. 38:4982019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Matsuyama M, Yoshimura R, Mitsuhashi M,
Tsuchida K, Takemoto Y, Kawahito Y, Sano H and Nakatani T:
5-Lipoxygenase inhibitors attenuate growth of human renal cell
carcinoma and induce apoptosis through arachidonic acid pathway.
Oncol Rep. 14:73–79. 2005.PubMed/NCBI
|
|
34
|
Matsuyama M and Yoshimura R: Relationship
between arachidonic acid pathway and human renal cell carcinoma.
OncoTargets Ther. 1:41–48. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tasaki S, Horiguchi A, Asano T, Ito K,
Asano T and Asakura H: Docosahexaenoic acid inhibits the
phosphorylation of STAT3 and the growth and invasion of renal
cancer cells. Exp Ther Med. 14:1146–1152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McCabe AJ, Wallace JMW, Gilmore WS,
McGlynn H and Strain SJ: Docosahexaenoic acid reduces in vitro
invasion of renal cell carcinoma by elevated levels of tissue
inhibitor of metalloproteinase-1. J Nutr Biochem. 16:17–22. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Balaban S, Lee LS, Varney B, Aishah A, Gao
Q, Shearer RF, Saunders DN, Grewal T and Hoy AJ: Heterogeneity of
fatty acid metabolism in breast cancer cells underlies differential
sensitivity to palmitate-induced apoptosis. Mol Oncol.
12:1623–1638. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Balaban S, Nassar ZD, Zhang AY,
Hosseini-Beheshti E, Centenera MM, Schreuder M, Lin HM, Aishah A,
Varney B, Liu-Fu F, et al: Extracellular Fatty Acids Are the Major
Contributor to Lipid Synthesis in Prostate Cancer. Mol Cancer Res.
17:949–962. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bellini L, Campana M, Rouch C, Chacinska
M, Bugliani M, Meneyrol K, Hainault I, Lenoir V, Denom J, Véret J,
et al: Protective role of the ELOVL2/docosahexaenoic acid axis in
glucolipotoxicity-induced apoptosis in rodent beta cells and human
islets. Diabetologia. 61:1780–1793. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Masuoka HC, Mott J, Bronk SF, Werneburg
NW, Akazawa Y, Kaufmann SH and Gores GJ: Mcl-1 degradation during
hepatocyte lipoapoptosis. J Biol Chem. 284:30039–30048. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shimabukuro M, Wang MY, Zhou YT, Newgard
CB and Unger RH: Protection against lipoapoptosis of beta cells
through leptin-dependent maintenance of Bcl-2 expression. Proc Natl
Acad Sci USA. 95:9558–9561. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Akazawa Y, Cazanave S, Mott JL, Elmi N,
Bronk SF, Kohno S, Charlton MR and Gores GJ: Palmitoleate
attenuates palmitate-induced Bim and PUMA up-regulation and
hepatocyte lipoapoptosis. J Hepatol. 52:586–593. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cazanave SC, Wang X, Zhou H, Rahmani M,
Grant S, Durrant DE, Klaassen CD, Yamamoto M and Sanyal AJ:
Degradation of Keap1 activates BH3-only proteins Bim and PUMA
during hepatocyte lipoapoptosis. Cell Death Differ. 21:1303–1312.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hu H, Tian M, Ding C and Yu S: The C/EBP
Homologous Protein (CHOP) Transcription Factor Functions in
Endoplasmic Reticulum Stress-Induced Apoptosis and Microbial
Infection. Front Immunol. 9:30832019. View Article : Google Scholar : PubMed/NCBI
|