Introduction
Hepatoblastoma (HB) is the most common malignant
liver tumor in children, arising from hepatic progenitors or
hepatoblast cells and accounting for 1% of all pediatric cancer
cases in the USA (1). Potentially
due to the improvement in the survival rate of premature infants,
its incidence has increased by 2.7% per year over the last few
decades (2). Currently,
international clinical studies have been focusing on risk-adapted
therapy for patients at a standard risk with potentially resectable
tumors and for those at a high risk with unresectable tumors due to
extrahepatic extensions, large hepatic vein or main portal vein
invasion, and high serum α-fetoprotein levels (3). Most standard-risk patients have been
reported to respond to cisplatin (CDDP) treatment. In addition,
according to a report from SIOPEL-4, the combination of CDDP and
doxorubicin has improved outcomes for high-risk patients, with a
3-year overall survival rate of 83% and a 3-year event-free
survival rate of 76% (3). However,
these results are still unsatisfactory.
Molecular-targeted therapies intended to attack
specific molecules in signaling pathways responsible for tumor
development are currently a hot topic in the field of oncology
research. Among those molecules, inhibitors of apoptosis proteins
(IAPs), such as cellular IAP1 (cIAP1), cIAP2 and X-linked IAP
(XIAP), work as critical regulators of apoptosis. IAPs exert their
effects by directly binding to caspase-3, −7 and −9 (4–10).
IAPs are also involved in the activation of signaling pathways
responsible for tumorigenesis (11). At present, several clinical and
preclinical trials investigating the effects of interfering with
cIAP1, cIAP2 and XIAP function in tumor cells as a type of
biological approach to cancer therapy are in progress (12,13).
Birinapant is a divalent second mitochondria-derived activator of
caspase-mimetic particle specifically designed to target cIAP1,
cIAP2 and XIAP (6,12–14).
However, the clinical importance of cIAP1, cIAP2 and XIAP in HB is
yet to be elucidated.
The present study focused on the antitumor effect of
cIAP1 inhibition in HB cells. To the best of our knowledge, this
study is the first to focus on this topic. The study aimed to
provide insights into a new therapeutic strategy for HB by using
birinapant as a molecular effector of IAPs. Standard chemotherapy
for patients with HB in the standard-risk group is based on CDDP
monotherapy, but the late side effects of CDDP are urgent issues
that need to be addressed for children with HB. Therefore, the
study also investigated whether the CDDP dosage could be decreased
by combining it with birinapant. Furthermore, CDDP-resistant Huh6
(Huh6-CDDPR) cells were established by repeatedly
treating Huh6 wild-type (Huh6-WT) cells with CDDP in order to
investigate the potential of birinapant as a salvage drug for
patients with HB who do not respond to CDDP.
Materials and methods
Cell lines and cell culture
The human HB Huh6 and HepG2 cell lines were
purchased from the Japanese Collection of Research Bioresources
Cell Bank. The cells were maintained in high-glucose Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% fetal bovine serum, 100 U/ml penicillin
and 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.),
and were incubated at 37°C in a humidified atmosphere of 95% air
and 5% CO2. Both cell lines were correctly authenticated
by short tandem repeat profiling (cell numbers: KBN0819-01 and
KBN0819-02).
Reagents
Birinapant was obtained from Selleck Chemicals and
CDDP was obtained from Wako Pure Chemical Industries, Ltd.
Establishment of CDDP-resistant HB
cells
Huh6-WT cells were continuously exposed to
increasing concentrations of CDDP (up to 4.5 µM over 6 months), and
the surviving cells were determined to be Huh6-CDDPR
cells. To ensure persisting resistance, the cells were maintained
in DMEM containing CDDP (4.5 µM) for 72 h, and cell survival was
confirmed using Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cultured Huh6 cells 16
h after exposure to 100 µM birinapant and from excised tumor
tissues of Huh6-xenograft in mice using RA1, RAW2 and RA3 buffers
in a NucleoSpin RNA kit (Takara Bio, Inc.). Premix Taq kit (Takara
Bio, Inc.) was used for RT-PCR to synthesize cDNA following the
manufacture's protocol. Human cIAP1 and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) were amplified by using SYBR Premix Ex Taq II
(Takara Bio, Inc.). qPCR, consisting of annealing at 95°C for 30
sec, followed by 40 cycles of PCR at 95°C for 5 sec and 60°C for 34
sec, and melt curve analysis, was performed using an Applied
Biosystems 7900HT Fast Real-Time PCR System (Thermo Fisher
Scientific, Inc.). All the aforementioned procedures were performed
following the manufacturer's instructions. Primers (purchased from
Invitrogen; Thermo Fisher Scientific, Inc.) were as follows: cIAP1
forward, 5′-TGTTGTCAACTTCAGATACCACTGG and reverse,
5′-CATCATGACAGCATCTTCTGAAGA; and GAPDH forward,
5′-ATCTTCCAGGAGCGAGATCC and reverse, 5′-ACCACTGACACGTTGGCAGT. The
results from each sample were analyzed and compared using the
2−∆∆Cq method (15).
Experiments were performed in triplicate.
Western blot analysis
Rabbit anti-cIAP1 antibody (catalog no. 7065) and
rabbit anti-β-actin antibody (catalog no. 4967) were purchased from
Cell Signaling Technology, Inc., and were used as primary
antibodies at a 1:1,000 dilution. Horseradish peroxidase
(HRP)-conjugated anti-rabbit IgG (catalog no. 7074), also obtained
from Cell Signaling Technology, Inc., was used as the secondary
antibody at a 1:1,000 dilution. At 16 h post-exposure to 50 or 100
µM birinapant, protein samples were extracted from cell lysates of
Huh6 or HepG2 by RIPA buffer (catalog no. 16488-34; Nacalai Tesque,
Inc.). After protein determination by bicinchoninic acid assay, 40
µg of each protein sample was loaded per lane, separated using
5–20% gradient polyacrylamide gels and then transferred onto
polyvinylidene difluoride membranes. The membrane was blocked using
blocking buffer consisting of Tris-buffered saline with 0.1%
Tween-20 (TBS-T) and 5% skimmed milk for 1 h at room temperature,
followed by incubation with the primary antibodies overnight at
4°C. The membranes were then washed with TBS-T and incubated with a
secondary antibody for 1 h at room temperature. Immunoreactive
signals were detected using Immunostar Zeta (Wako Pure Chemical
Industries, Ltd.). Each protein of interest was visualized using
ImageQuant LAS 4000 Mini (Cytiva). The intensity of each band was
quantified using the downloaded ImageJ software (ImageJ bundled
with 64-bit java version 1.8.0_172).
Cell viability assay
Viable Huh6 or HepG2 cells were seeded at a density
of ~5×103 cells per well in a 96-well plate and cultured
in medium with or without various concentrations of birinapant
(3.2, 6.3, 12.5, 25, 50 and 100 µM) and/or CDDP (3, 3.2, 6, 6.3,
12.5, 25, 50 and 100 µM) for 24 and 72 h. Dimethyl sulfoxide (DMSO)
(0.1%) was used as a control. The relative viability of cells in
each well was examined using CCK-8 (Dojindo Molecular Technologies,
Inc.) at 24 and 72 h after exposure to these reagents. The duration
of incubation with the CCK-8 reagent was 3 h.
In vivo tumor xenotransplantation
experiments
A total of 20 female SCID mice (4 weeks old; body
weight, 17–19 g; Japan SLC, Inc.) were used in this experiment
according to the Osaka University Protocol and were maintained at
25°C and in 50% humidity, with a 12-h light/12-h dark cycle in an
aseptic room. The animals had free access to sterile water and
food. Viable Huh6 cells (~2×106) suspended in 100 ml PBS
were injected into the subcutaneous space on the backs of
4-week-old female SCID mice (n=3). The body weight of each mouse on
the day of tumor inoculation was around 18 g. When the tumor
diameter reached 15 mm, tumors were excised and cut into
3-mm3 sections followed by the implantation into the
subcutaneous space on the hind flank of other 4-week-old female
SCID mice. The body weight of each mouse on the day of tumor
implantation was around 18 g. After the tumor volume grew to ~100
mm3, 30 mg/kg birinapant (n=4), 2.5 mg/kg CDDP (n=4) and
CDDP + birinapant (n=4) were administered intraperitoneally twice a
week for 4 weeks, with PBS as the control (n=5), and then the mice
were carefully observed for an additional 2 weeks without
treatment. Tumor diameters were measured three times a week in a
blinded manner using electric calipers. The maximum tumor size
among them throughout this experiment was 11.6×9.6 mm (534.5
mm3) in the control group. Tumor volumes were calculated
using the following formula: Tumor volume (mm3)=length
x(width)2/2. Due to the differences in mean tumor size
between groups on day 0 (Fig. S1A and
B), the tumor volume of each mouse after day 0 was defined as
the ratio to the tumor volume of day 0. All mice were euthanized 42
days after the treatment initiation by stepwise escalation (30–50%
of chamber volume, 0.02 MPa) of carbon dioxide inhalation. In order
to minimize suffering, all animal experiments were reviewed and
approved by the Animal Experimentation Committee of Osaka
University (Suita, Japan; approval no. 01-006-004) and were
conducted according to the institutional guidelines and national
and international guidelines and laws for humane animal care and
research ethics (including ARRIVE guidelines).
Immunohistochemistry of HB
specimens
Paraffin-embedded tumor tissues and normal liver
samples (≥15 mm from the tumor) were obtained from patients with HB
in the form of biopsies before chemotherapy and resection after
chemotherapy at Osaka University Hospital. A total of 3 patients (a
10-month-old male, a 2-year-old female and a 4-year-old male) were
enrolled in this study between June 2015 and March 2020. Written
informed consent was obtained from the legal guardians of the
patients for the use of the tissues in laboratory analysis
according to the institutional requirements at the time of
admission. This study was reviewed and approved by the
Institutional Review Board of Osaka University Hospital (approval
no. 15022) and was conducted in accordance with institutional,
national and international guidelines. The expression of cIAP1 was
evaluated using immunohistochemical staining. Formalin-fixed and
paraffin-embedded sections were stained using the
ImmPACT™ DAB Substrate kit (catalog no. SK-4105; Vector
Laboratories, Inc.). Tissues were fixed with 4% paraformaldehyde at
4°C for 72 h, followed by paraffin-embedding. Paraffin-embedded
samples were sliced into 6-µm-thick sections. Dewaxing of the
sections were performed by heating followed by soaking in 100, 90,
80 and 70% alcohol, respectively. In order to avoid endogenous
peroxidase/phosphatase activity, slides were treated with 3%
hydrogen peroxide. After this quenching step, slides were incubated
with 1.5% skimmed milk at 25°C for 20 min. An anti-cIAP1 rabbit
polyclonal antibody (catalog no. ab2399; Abcam) was used as the
primary antibody. Slides were incubated overnight at 4°C with a
1:500 dilution of cIAP1 antibodies in phosphate-buffered saline.
Biotin-conjugated secondary antibodies (as aforementioned) were
then applied and the slides were incubated with a 1:200 dilution of
avidin-biotin complex using VECTASTAIN Elite ABC Kit (catalog no.
PK-6101; Vector Laboratories, Inc.) at 25°C for 30 min. As a
chromogen detection step, the expression of cIAP1 was visualized
using an ImmPACT™ DAB Substrate Kit and counter-stained
with hematoxylin at 25°C for 30 sec, and then imaged under a light
microscope.
Statistical analysis
Data are shown as the mean ± standard deviation.
One-way or two-way analysis of variance (ANOVA) followed by the
Tukey-Kramer post hoc test was used for determining the statistical
significance of three or more between-group differences. Student's
t-test was used for determining the statistical significance of two
between-group differences. P<0.05 was used to indicate a
statistically significant difference.
Results
Antitumor effects of birinapant in
vitro
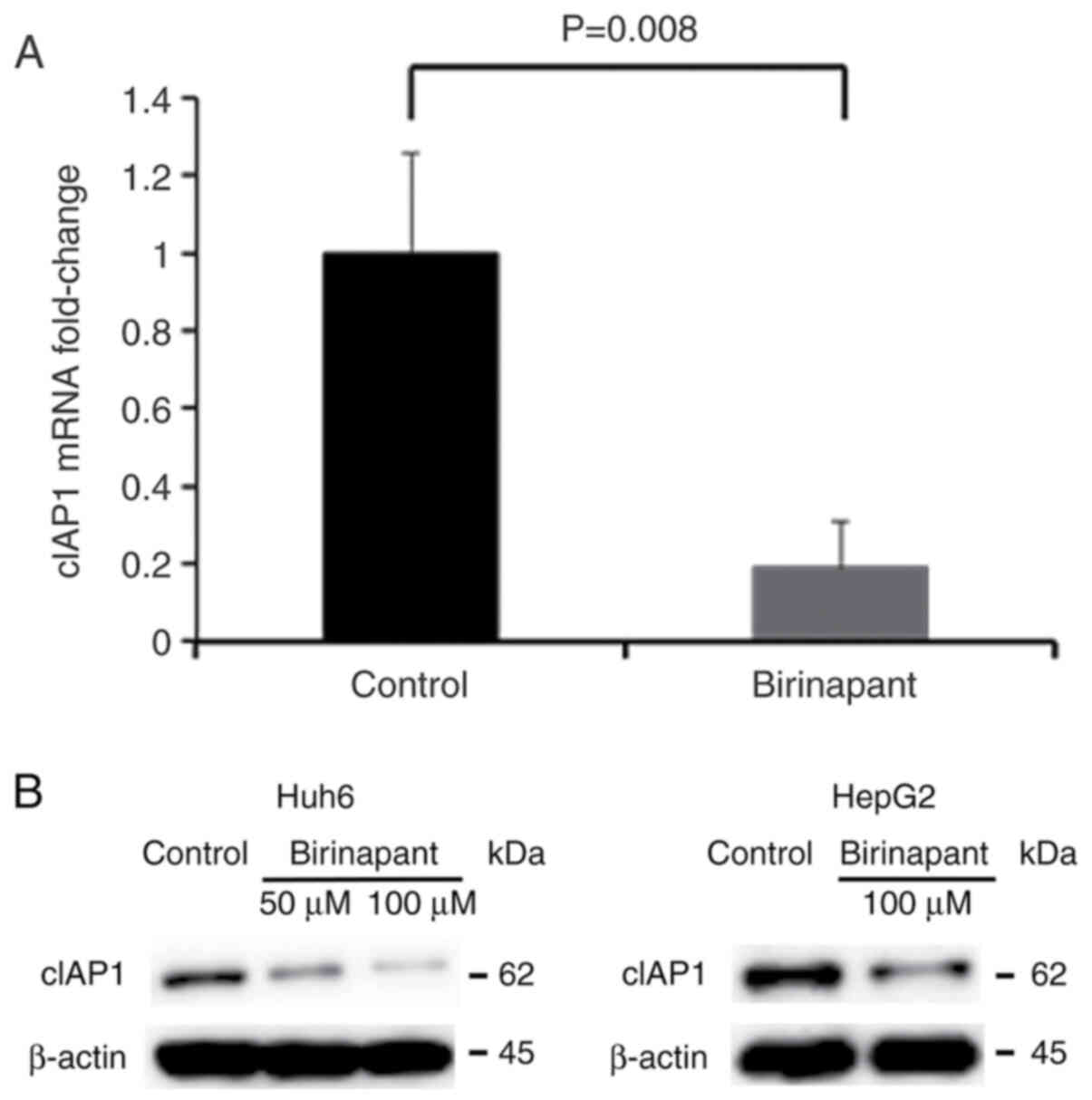
At 16 h post-exposure to 50 or 100 µM birinapant,
the results of RT-qPCR and western blot analysis confirmed the
suppressive effect of birinapant on cIAP1 expression in Huh6 and
HepG2 cells (Fig. 1A and B). The
mRNA expression of cIAP1 was not suppressed in HepG2 cells (data
not shown) and thus only the higher concentration (100 µM) of
birinapant was shown in the western blot images of HepG2 cells
(Fig. 1B).
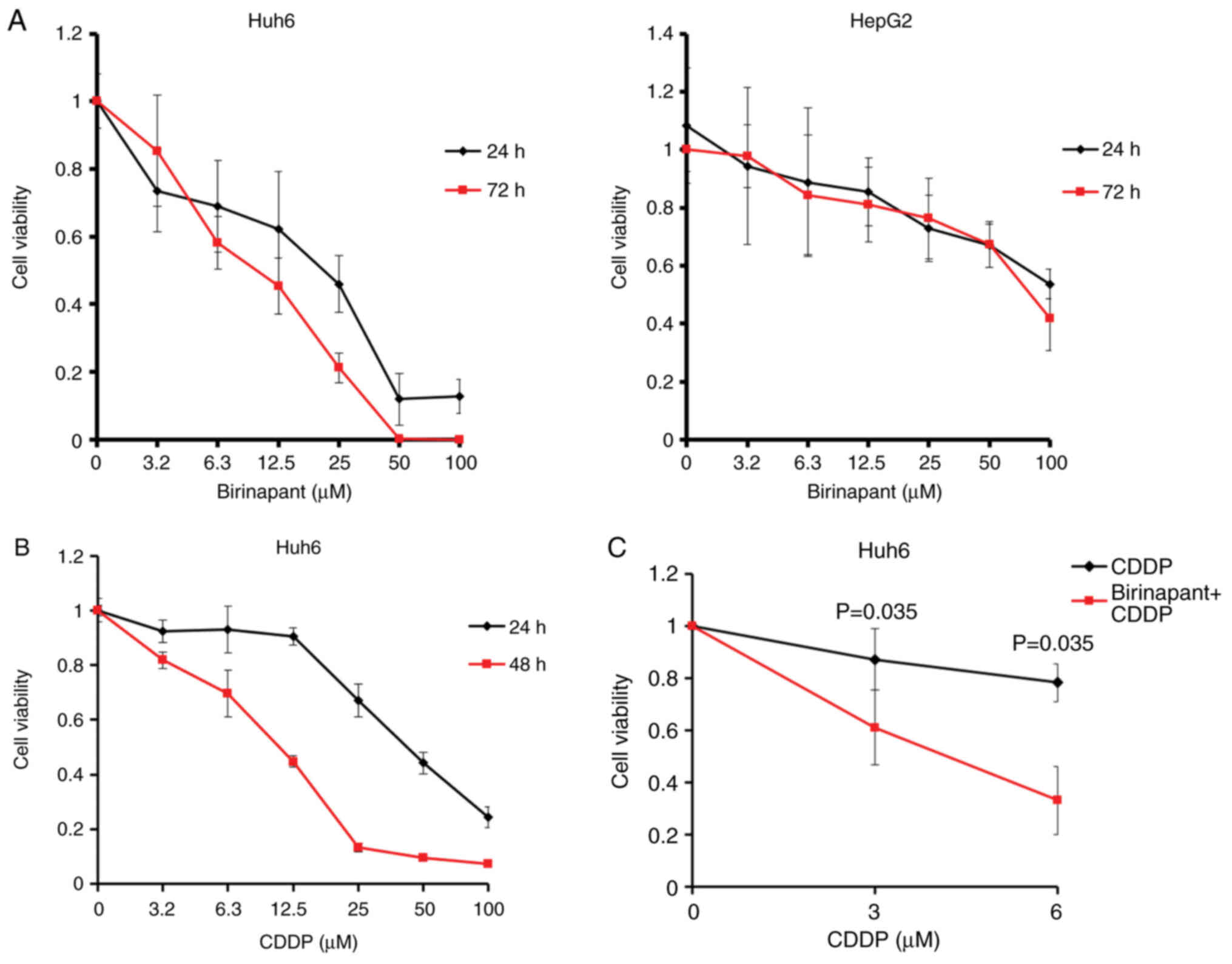
Consistent with these results, it was demonstrated
that treatment with birinapant for 24 and 72 h decreased the
viability of the Huh6 cells and HepG2 cells in a dose-dependent
manner (Fig. 2A). Based on these
results, the half maximal inhibitory value (IC50) values
of birinapant against Huh6 cells were 21 µM (24 h) and 9.8 µM (72
h), and those against HepG2 cells were over 100 µM (24 h) and 80.3
µM (72 h), suggesting that the Huh6 cells were more sensitive to
birinapant than the HepG2 cells (Fig.
2A).
In addition, the study confirmed the antitumor
efficacy of CDDP against Huh6 cells in a dose-dependent manner
(Fig. 2B). Based on this result,
the IC50 of CDDP against Huh6 cells was 41.9 µM at 24 h
and 10.7 µM at 48 h following exposure to CDDP. The combination of
birinapant (80 µM) and CDDP (ranging from 3–6 µM) significantly
decreased the cell viability of the Huh6 cells compared with use of
CDDP monotherapy (P=0.035; Fig.
2C), implying that combination treatment with birinapant
reduces the therapeutic dose of CDDP.
Antitumor effects of birinapant and
CDDP on HB xenografts
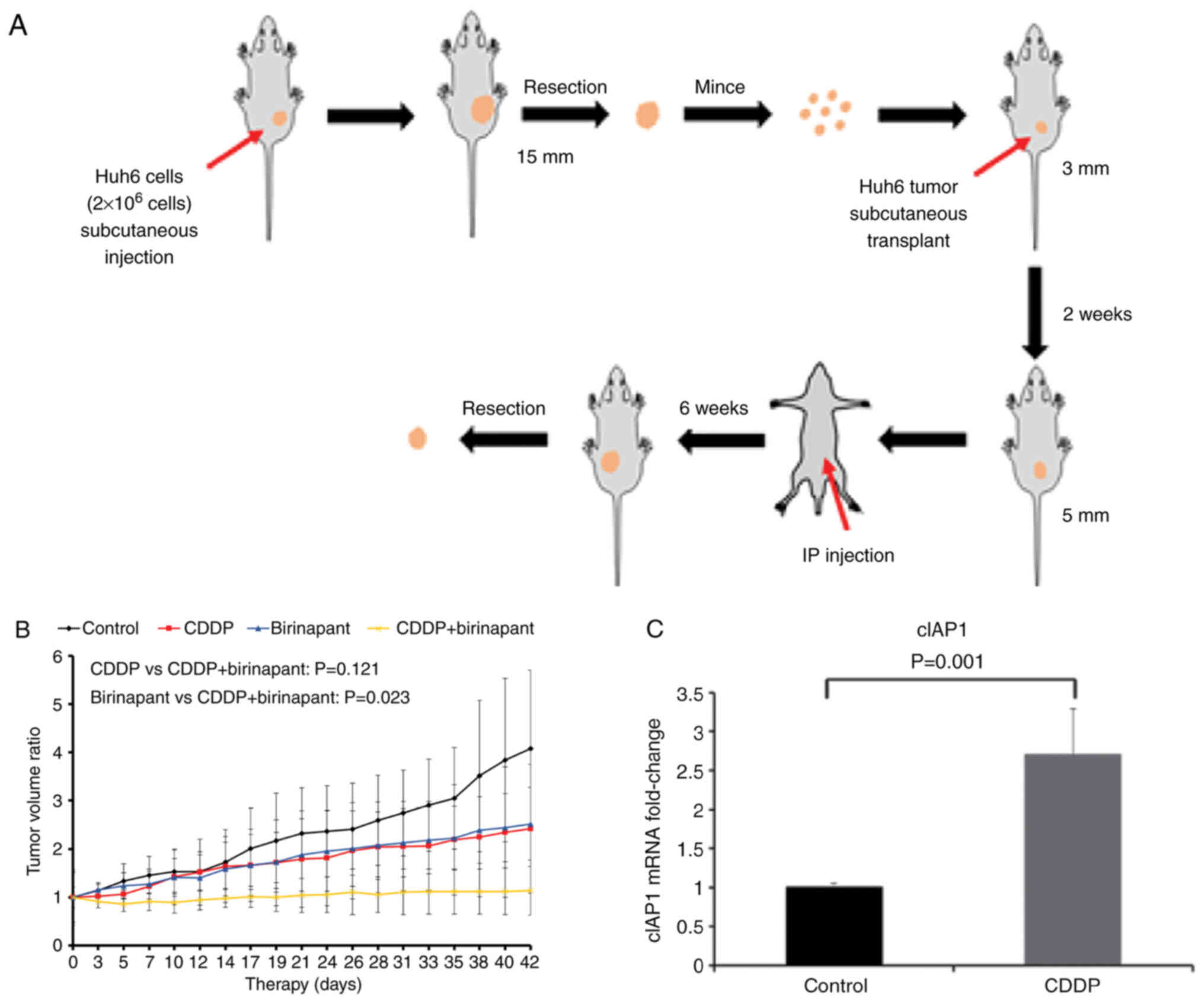
Next, the in vivo antitumor activity of
birinapant and CDDP was evaluated using Huh6 cells and the
antitumor efficacy between the control group (0.1% DMSO), the CDDP
group, the birinapant group and the CDDP + birinapant group was
compared (Fig. 3A and B).
Birinapant tended to enhance the antitumor activity of CDDP
(P=0.121) and CDDP significantly enhanced the antitumor activity of
birinapant (P=0.023). As is shown in Fig. 2C, birinapant could significantly
enhance the antitumor efficacy of CDDP in vitro, but this
effect was somewhat modest in the in vivo model. The
discrepancy in the antitumor efficacy of the combination treatment
with CDDP and birinapant between the in vitro and in
vivo experiments raises a new question. Specifically,
considering the in vitro model of short-term exposure to
CDDP, we hypothesized that long-term repeated administration of
CDDP may have caused genetic alterations in cIAP1 expression within
the xenograft tumors. On day 42 of therapy, all xenograft tumors in
each group were excised and the mRNA expression of cIAP1 was
quantified. It was found that the expression of cIAP1 was
significantly higher in the CDDP group compared with that in the
control group (Fig. 3C).
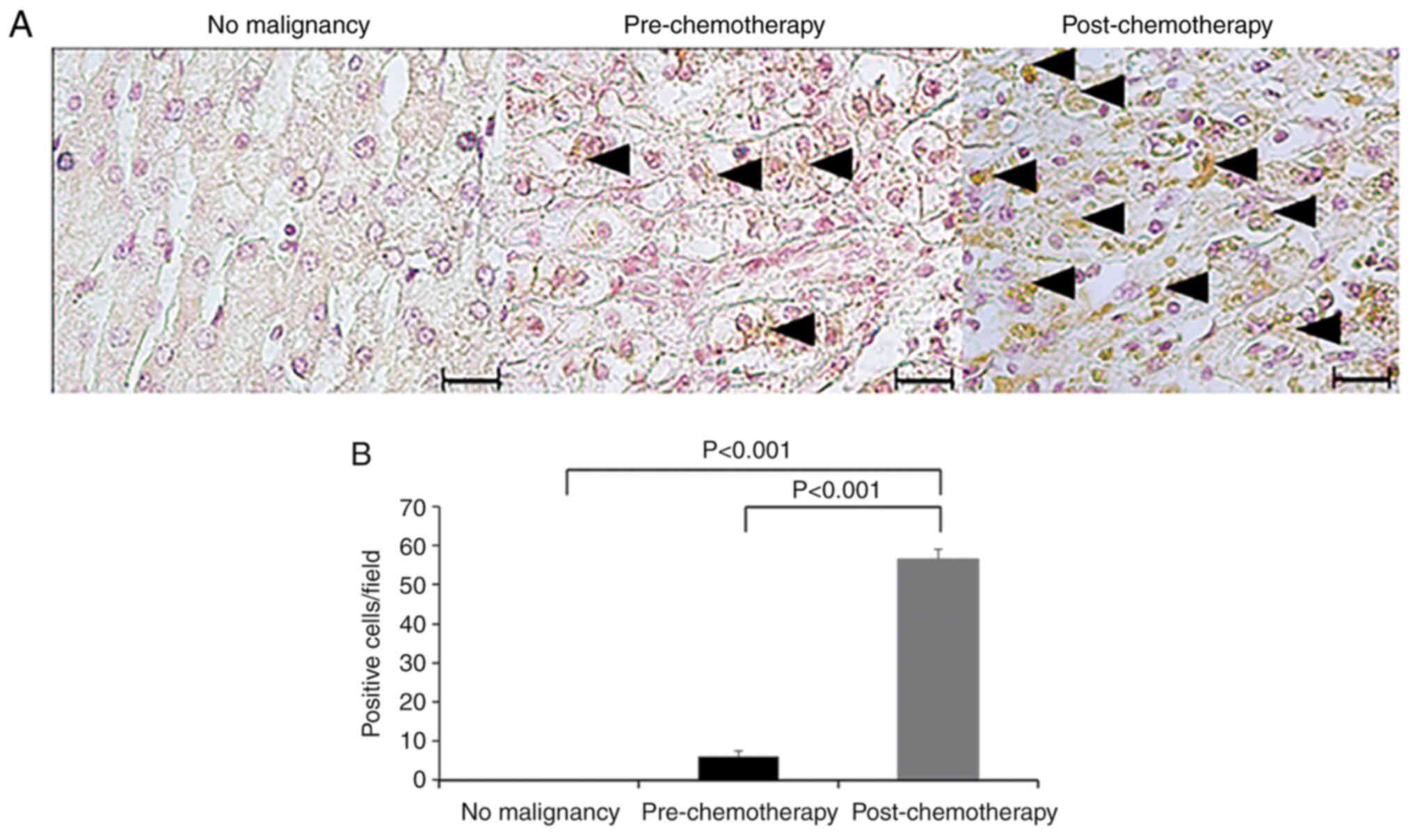
cIAP1 expression in HB specimens
cIAP1 expression was also compared in HB specimens
before and after chemotherapy (CDDP) using immunohistochemical
staining. It was found that the expression of cIAP1 in both
pre-chemotherapy and post-chemotherapy specimens was higher than
that in normal liver specimens. Furthermore, the expression of
cIAP1 in the post-chemotherapy specimens was significantly higher
than that in the pre-chemotherapy specimens (P<0.001; Fig. 4A and B).
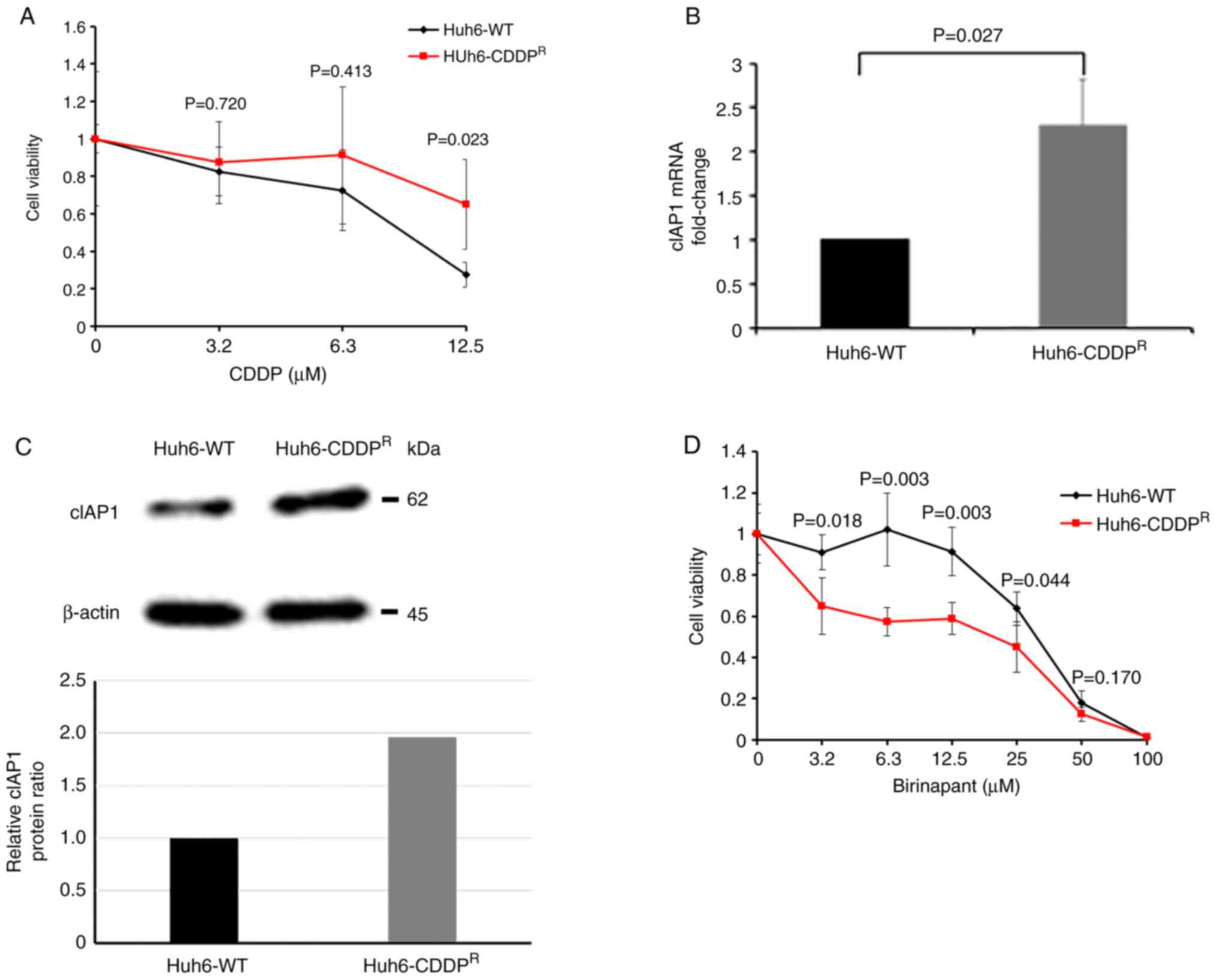
Establishment of CDDP-resistant cells
and cytotoxic effects of birinapant
The genetic alterations in CDDP-resistant HB cells
in vitro were examined by mimicking the clinical course of
chemotherapy using CDDP. First, the cytotoxic effects of CDDP were
compared in Huh6-WT and Huh6-CDDPR cells. The cell
viability of the Huh6-CDDPR cells was significantly
higher than that of the Huh6-WT cells 72 h after exposure to 12.5
µM CDDP (P=0.023); this finding demonstrated the CDDP resistance of
the Huh6-CDDPR cells (Fig.
5A). Next, the mRNA expression of cIAP1 was compared in Huh6-WT
cells and Huh6-CDDPR cells, and a significant increase
in cIAP1 expression was found in the Huh6-CDDPR cells
(Fig. 5B). Furthermore, via
western blot analysis, it was confirmed that the expression of
cIAP1 at the protein level was increased in the
Huh6-CDDPR cells. The intensity of each band was
quantified using Image J software and the expression of cIAP1 was
defined as the ratio to the expression of β-actin (Fig. 5C). Finally, the antitumor effects
of birinapant on Huh6-WT and Huh6-CDDPR cells were
evaluated 72 h after treating these cells with various
concentrations of birinapant, and it was demonstrated that
birinapant could significantly decrease the viability of the
Huh6-CDDPR cells even at low concentrations (i.e.,
ranging from 3.2 to 12.5 µM) (Fig.
5D).
Discussion
The present study examined the potential of
birinapant for treating patients with HB and whether it exhibits
sufficient cytotoxicity against Huh6 cells both in vitro and
in vivo. To the best of our knowledge, the present study is
the first to evaluate this research question. The findings imply
that the combination of birinapant and CDDP is a promising
therapeutic modality for HB, and that birinapant could be a
candidate therapeutic drug for patients with HB who have undergone
chemotherapy and acquired chemotherapeutic resistance.
IAP proteins, such as cIAP1 and melanoma IAP,
restrain apoptosis and upregulate the tumor necrosis
factor-dependent pro-survival NF-κB pathway, which is closely
associated with the survival of tumor cells and infectious diseases
(16–19). The expression of IAP genes encoding
these proteins is also reported to be enhanced in numerous tumors
such as lung and colorectal cancer, and overexpressed cIAP1 can
inhibit apoptotic cell death in variable tumor cell types such as
multiple myeloma and ovarian cancer (16). In addition, it has been reported
that overexpressed IAP genes make a large contribution to
resistance to conventional cancer chemotherapeutic agents in
several tumors (19). Specially,
some reports have shown the impact of the enhanced expression of
cIAP1 or cIAP2 on chemotherapeutic resistance or poor survival rate
in patients with various cancer types such as head and neck cancer,
and prostate cancer (20,21). With regard to the overexpression of
cIAP1 or cIAP2, chemotherapeutic resistance to multiple drugs in
multiple myeloma (22), resistance
to CDDP in lung cancer (23),
resistance to fluorouracil in colorectal cancer (24) and oral squamous cell carcinoma
(25), resistance to paclitaxel,
doxorubicin and CDDP in pancreatic cancer (26), and resistance to CDDP in head and
neck cancer (27) were previously
reported.
Based on this accumulated evidence, the present
study focused on the relationship between cIAP1 expression and CDDP
resistance in HB. CDDP-resistant cells were established in
vitro and cIAP1 expression was shown to be significantly
increased in Huh6-CDDPR cells compared with that in
Huh6-WT cells. It was also confirmed that cIAP1 expression
increased after continuous treatment with CDDP in specimens
obtained by biopsy from patients with HB. These results suggest
that cIAP1 is an important factor influencing the mechanisms of
CDDP resistance in HB.
Furthermore, birinapant was administered to
CDDP-resistant HB cells and they showed a significant increase in
drug sensitivity compared with Huh6-WT cells treated in the same
manner, suggesting that birinapant may be utilized as a second-line
therapeutic modality for patients with HB who do not sufficiently
respond to CDDP.
The potential toxicity of birinapant should be also
taken into consideration prior to future clinical use. In the
present study, the body weight of all mice was monitored weekly
throughout the experimental period and no weight loss (data not
shown) or anoxia was found. It has also been reported that
birinapant could selectively target plasmodium-infected hepatocytes
and induce apoptosis, implying that birinapant has only a trivial
side effect on normal hepatocytes (28).
The major limitation of the present study is that it
demonstrated the in vitro evidence of acquired
chemotherapeutic resistance to CDDP in HB cells by using only one
pair of wild-type and CDDP-resistant HB cell lines. Therefore, it
is necessary to gather more evidence to confirm the critical role
of cIAP1 in CDDP resistance within HB cells by establishing more
CDDP-resistant HB cell lines, although it will take a long time to
prepare these cells. Another limitation of the current
investigation is that it did not evaluate the antitumor activity of
birinapant against Huh6-CDDPR in vivo due to the
difficulties in maintaining and expanding the CDDP-resistant tumor
cells available for the in vivo experiments.
In conclusion, the present study demonstrated that
cIAP1 expression was upregulated in both CDDP-resistant HB cells
and in specimens from patients with HB who underwent repeated
chemotherapy with CDDP. Additionally, birinapant in combination
with CDDP synergistically decreased HB cell proliferation in
vitro and suppressed the tumor growth of HB in a SCID mouse
model. These findings suggest that the combination of birinapant
and CDDP is a promising therapeutic modality for patients with HB.
Furthermore, birinapant may also work as a second-line therapeutic
drug for patients with HB showing resistance to CDDP, which is the
standard drug for patients with this disease. The findings provide
new insights into salvage drug therapy for patients with
chemotherapeutic-resistant cancer.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Ms. Tomoko Haneda
(Research assistant at the Department of Pediatric Surgery, Osaka
University Graduate School of Medicine, Suita, Osaka, Japan) for
kindly helping with the completion of several in vitro
experiments.
Funding
This study was supported by the Japan Society for the Promotion
of Science KAKENHI (grant no. 20K17551).
Availability of data and materials
The datasets used in the present study are available
from the corresponding author upon reasonable request.
Authors' contributions
RT, MN and TU designed the study. RT and MN
performed the experiments. MN and HO reviewed and revised the
manuscript. TU and MN confirm the authenticity of all the raw data.
All authors approved the final version of the manuscript.
Ethics approval and consent to
participate
All animal experiments were reviewed and approved by
the Animal Experimentation Committee of Osaka University (Suita,
Japan; approval no. 01-006-004). The study was also reviewed and
approved by the Institutional Review Board of Osaka University
Hospital (approval no. 15022). Written consent was obtained by from
the legal guardians of the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IAP
|
inhibitor of apoptosis protein
|
|
cIAP1
|
cellular IAP-1
|
|
XIAP
|
X-linked IAP
|
References
|
1
|
Lim IIP, Bondoc AJ, Geller JI and Tiao GM:
Hepatoblastoma-the evolution of biology, surgery, and
transplantation. Children (Basel). 6:12018.PubMed/NCBI
|
|
2
|
Aronson DC, Czauderna P, Maibach R,
Perilongo G and Morland B: The treatment of hepatoblastoma: Its
evolution and the current status as per the SIOPEL trials. J Indian
Assoc Pediatr Surg. 19:201–207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zsiros J, Brugieres L, Brock P, Roebuck D,
Maibach R, Zimmermann A, Childs M, Pariente D, Laithier V, Otte JB,
et al: Dose-dense cisplatin-based chemotherapy and surgery for
children with high-risk hepatoblastoma (SIOPEL-4): A prospective,
single-arm, feasibility study. Lancet Oncol. 14:834–842. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schimmer AD: Inhibitor of apoptosis
proteins: Translating basic knowledge into clinical practice.
Cancer Res. 64:7183–7190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Finlay D, Teriete P, Vamos M, Cosford NDP
and Vuori K: Inducing death in tumor cells: Roles of the inhibitor
of apoptosis proteins. F1000Res. 6:5872017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Benetatos CA, Mitsuuchi Y, Burns JM,
Neiman EM, Condon SM, Yu G, Seipel ME, Kapoor GS, LaPorte MG,
Rippin SR, et al: Birinapant (TL32711), a bivalent SMAC mimetic,
targets TRAF2-associated cIAPs, abrogates TNF-induced NF-κB
activation, and is active in patient-derived xenograft models. Mol
Cancer Ther. 13:867–879. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dynek JN and Vucic D: Antagonists of IAP
proteins as cancer therapeutics. Cancer Lett. 332:206–214. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nachmias B, Ashhab Y and Ben-Yehuda D: The
inhibitor of apoptosis protein family (IAPs): An emerging
therapeutic target in cancer. Semin Cancer Biol. 14:231–243. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Camp JG, Sekine K, Gerber T,
Loeffler-Wirth H, Binder H, Gac M, Kanton S, Kageyama J, Damm G,
Seehofer D, et al: Multilineage communication regulates human liver
bud development from pluripotency. Nature. 546:533–538. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tamm I, Wang Y, Sausville E, Scudiero DA,
Vigna N, Oltersdorf T and Reed JC: IAP-family protein survivin
inhibits caspase activity and apoptosis induced by Fas (CD95), Bax,
caspases, and anticancer drugs. Cancer Res. 58:5315–5320.
1998.PubMed/NCBI
|
|
11
|
Hunter AM, LaCasse EC and Korneluk RG: The
inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis.
12:1543–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amaravadi RK, Schilder RJ, Martin LP,
Levin M, Graham MA, Weng DE and Adjei AA: A phase I study of the
SMAC-mimetic birinapant in adults with refractory solid tumors or
lymphoma. Mol Cancer Ther. 14:2569–2575. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Noonan AM, Bunch KP, Chen JQ, Herrmann MA,
Lee JM, Kohn EC, O'Sullivan CC, Jordan E, Houston N, Takebe N, et
al: Pharmacodynamic markers and clinical results from the phase 2
study of the SMAC mimetic birinapant in women with relapsed
platinum-resistant or -refractory epithelial ovarian cancer.
Cancer. 122:588–597. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Allensworth JL, Sauer SJ, Lyerly HK, Morse
MA and Devi GR: Smac mimetic Birinapant induces apoptosis and
enhances TRAIL potency in inflammatory breast cancer cells in an
IAP-dependent and TNF-α-independent mechanism. Breast Cancer Res
Treat. 137:359–371. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stucchi G, Battevi N, Cairoli S and
Consonni D: The prevalence of musculoskeletal disorders in the
retail sector: An Italian cross sectional study on 3,380 workers.
Med Lav. 107:251–262. 2016.PubMed/NCBI
|
|
17
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
LaCasse EC, Baird S, Korneluk RG and
MacKenzie AE: The inhibitors of apoptosis (IAPs) and their emerging
role in cancer. Oncogene. 17:3247–3259. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wallace C and Unger S: A matter of life
and death. Commun Int. 26:341999.
|
|
20
|
Krajewska M, Krajewski S, Banares S, Huang
X, Turner B, Bubendorf L, Kallioniemi OP, Shabaik A, Vitiello A,
Peehl D, et al: Elevated expression of inhibitor of apoptosis
proteins in prostate cancer. Clin Cancer Res. 9:4914–4925.
2003.PubMed/NCBI
|
|
21
|
Tanimoto T, Tsuda H, Imazeki N, Ohno Y,
Imoto I, Inazawa J and Matsubara O: Nuclear expression of cIAP-1,
an apoptosis inhibiting protein, predicts lymph node metastasis and
poor patient prognosis in head and neck squamous cell carcinomas.
Cancer Lett. 224:141–151. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakagawa Y, Abe S, Kurata M, Hasegawa M,
Yamamoto K, Inoue M, Takemura T, Suzuki K and Kitagawa M: IAP
family protein expression correlates with poor outcome of multiple
myeloma patients in association with chemotherapy-induced
overexpression of multidrug resistance genes. Am J Hematol.
81:824–831. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu HH, Wu JY, Cheng YW, Chen CY, Lee MC,
Goan YG and Lee H: cIAP2 upregulated by E6 oncoprotein via
epidermal growth factor receptor/phosphatidylinositol 3-kinase/AKT
pathway confers resistance to cisplatin in human papillomavirus
16/18-infected lung cancer. Clin Cancer Res. 16:5200–5210. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Miura K, Karasawa H and Sasaki I: cIAP2 as
a therapeutic target in colorectal cancer and other malignancies.
Expert Opin Ther Targets. 13:1333–1345. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagata M, Nakayama H, Tanaka T, Yoshida R,
Yoshitake Y, Fukuma D, Kawahara K, Nakagawa Y, Ota K, Hiraki A and
Shinohara M: Overexpression of cIAP2 contributes to 5-FU resistance
and a poor prognosis in oral squamous cell carcinoma. Br J Cancer.
105:1322–1330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lopes RB, Gangeswaran R, McNeish IA, Wang
Y and Lemoine NR: Expression of the IAP protein family is
dysregulated in pancreatic cancer cells and is important for
resistance to chemotherapy. Int J Cancer. 120:2344–2352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee SK, Kim SB, Kim JS, Moon CH, Han MS,
Lee BJ, Chung DK, Min YJ, Park JH, Choi DH, et al: Butyrate
response factor 1 enhances cisplatin sensitivity in human head and
neck squamous cell carcinoma cell lines. Int J Cancer. 117:32–40.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ebert G, Lopaticki S, O'Neill MT, Steel
RWJ, Doerflinger M, Rajasekaran P, Yang ASP, Erickson S, Ioannidis
L, Arandjelovic P, et al: Targeting the extrinsic pathway of
hepatocyte apoptosis promotes clearance of plasmodium liver
infection. Cell Rep. 30:4343–4354.e4. 2020. View Article : Google Scholar : PubMed/NCBI
|