Introduction
According to global cancer statistics published by
the International Agency for Research on Cancer of the World Health
Organization, breast cancer surpassed lung cancer in 2020 to become
the most diagnosed cancer in the world (1). Triple-negative breast cancer (TNBC) is
the most aggressive subtype of breast cancer, characterized by
negative expression of estrogen receptor, progesterone receptor and
human epidermal growth factor receptor-2 (2). Although this subtype accounts for
15–20% of all types of breast cancer, it lacks effective
therapeutic targets and treatment options are limited (3). Therefore, it is critical to find novel
therapeutic targets for the treatment of TNBC.
Pseudolaric acid B (PAB) is a diterpene acid derived
from the cortex of Pseudolarix kaempferia (golden larch)
that exhibits diverse properties, including anti-inflammatory
(4), antifungal (5), antiangiogenic (6), pro-apoptotic (7) and microtubule-destabilizing (8) effects. In previous years, researchers
have shown that PAB has antitumor effects in liver cancer (9), gastric cancer (10), medulloblastoma (7), lung cancer (11) and leukemia (12). However, the mechanisms underlying
these antitumor effects remain unclear.
Apoptosis, also known as programmed cell death, is a
form of self-cleaning in which the body removes damaged cells in an
orderly and efficient manner. The dysregulation of apoptosis is a
major reason for the unlimited proliferation exhibited by tumor
cells (13). Thus, dysregulation of
apoptosis is considered one of the hallmarks of cancer. The
mitochondrial apoptotic pathway comprises the core apoptotic
signaling pathway in vertebrates and is triggered by a change in
mitochondrial outer membrane permeabilization (14). Subsequently, cytochrome c is
released from the mitochondria into the cytoplasm, stimulating
caspase-3 and other mechanisms of apoptosis (15). Previous studies demonstrated that
PAB induces apoptosis through the mitochondrial and death receptor
pathway in numerous types of cancer, including hepatocellular
(9), cervical (16), head and neck (17) and colorectal cancer (18). However, to the best of our
knowledge, the apoptotic mechanism of PAB in TNBC has not been
studied.
The PI3K/AKT/mTOR signaling pathway plays a key role
in several processes of tumor development, such as apoptosis,
proliferation, metabolism and metastasis (19). It can also enhance the malignancy of
various types of tumor cells (20).
Previous studies have demonstrated that PI3K/AKT/mTOR signaling
affects the mitochondrial apoptosis pathway by regulating proteins
in the Bcl-2 family (21–23).
The present study aimed to explore the anticancer
activity and related mechanisms of PAB in the TNBC MDA-MB-231 cell
line. Effects of PAB on migration and invasion were also
investigated.
Materials and methods
Reagents and antibodies
PAB was purchased from Beijing Solarbio Science
& Technology Co., Ltd. Primary antibodies for caspase-3
(1:1,000; cat. no. 19677-1-AP), caspase-9 (1:1,000; cat. no.
10380-1-AP), Bax (1:2,000; cat. no. 50599-2-Ig), Bcl-2 (1:2,000;
cat. no. 60178-1-Ig), p53 (1:5,000; cat. no. 80077-1-RR), p21
(1:2,000; cat. no. 10355-1-AP) and GAPDH (1:8,000; cat. no.
10494-1-AP), and HRP-conjugated Affinipure goat anti-rabbit (cat.
no. SA00001-2; 1:8,000) and anti-mouse (cat. no. SA00001-1;
1:8,000) IgG secondary antibodies were purchased from ProteinTech
Group, Inc. In addition, the primary antibodies for PARP (cat. no.
T40050); Bcl-xl (cat. no. T40057), Cytochrome c (cat. no. T55734),
PI3K (cat. no. T55224), AKT (cat. no. T55561), p-AKT (cat. no.
T40067), mTOR (cat. no. T55306), p-mTOR (cat. no. T56571) (all
1:1,000) were purchased from Abmart Pharmaceutical Technology Co.,
Ltd. Antibodies for CDK1 (1:1,000; cat. no. PTM-6521), cyclin B1
(1:1,000; cat. no. PTM6659), Vimentin (1:1,000; cat. no. PTM5376),
E-cadherin (1:2,000; cat. no. PTM6222) and N-cadherin (1:1,000;
cat. no. PTM5221) were purchased from PTM Biolabs, Inc. Annexin
V-FITC/PI Apoptosis Detection kit was purchased from Vazyme Biotech
Co., Ltd. Cell Counting Kit-8 (CCK-8) was purchased from
MedChemExpress. Mitochondrial membrane potential (MMP) assay kit
with JC-1 (cat. no. C2006) and western stripping buffer (cat. no.
P0025N) were purchased from Beyotime Institute of
Biotechnology.
Cell culture
The TNBC cell line MDA-MB-231 and human breast cell
line MCF-10A were donated by the Department of Oncology, Shengjing
Hospital of China Medical University (Shenyang, China). MDA-MB-231
cells were cultured in Leibovitz's L-15 medium (Procell Life
Science & Technology Co., Ltd.) containing 5% fetal bovine
serum (FBS; Procell Life Science & Technology Co., Ltd.) and 1%
antibiotics (100 U/ml penicillin and 100 U/ml streptomycin). MCF10A
cells were cultured in DMEM/F12 (Procell Life Science &
Technology Co., Ltd.) containing 5% HS, 20 ng/ml epidermal growth
factor, 0.5 µg/ml Hydrocortisone, 10 µg/ml insulin, 1%
non-essential amino acid and 1% antibiotics (100 U/ml penicillin
and 100 U/ml streptomycin). MDA-MB-231 cells were grown free of
CO2 in a cell culture incubator at 37°C. MCF-10A cells
were cultured at 37°C with 5% CO2.
Cell viability assay
The effect of PAB on cellular viability was measured
using a CCK-8 assay. Cells (5×103/well) were cultured in
a 96-well plate overnight, then treated with different
concentrations of PAB (0, 2.5, 5, 7.5, 10, 12.5 and 15 µM) at 37°C
for 24, 48 and 72 h. After treatment, the medium was removed and a
mixture of 90 µl medium (Leibovitz's L-15 or DMEM/F12) and 10 µl
CCK-8 reagent was added to each well (24). After 2 h of incubation at 37°C, the
absorbance at 450 nm was measured using a microplate reader.
Colony formation assay
To explore the effect of PAB on cellular
proliferation, the colony formation assay was conducted. MDA-MB-231
and MCF-10A cells (1×103/well) were seeded in six-well
plates overnight, then treated with different concentrations of PAB
(5, 7.5 and 10 µM) at 37°C for 48 h before changing back to a
drug-free medium (Leibovitz's L-15 or DMEM/F12). Cells were
cultured for an additional 14 days, then fixed with 4%
polyoxymethylene at room temperature for 15 min and stained with
0.1% crystal violet at room temperature for 15 min. After removing
the crystal violet, plates were washed twice with
phosphate-buffered saline (PBS) and the number of colonies was
counted and analyzed by Image J 1.8.0 software (National Institutes
of Health). A colony with more than 50 cells is defined as a
colony.
EdU staining assay
The BeyoClick™ EdU-488 cell proliferation kit
(Beyotime Institute of Biotechnology) was used to evaluate the
effect of PAB on cellular proliferation. MDA-MB-231 cells
(2×104/well) were seeded in 12-well plates and cultured
at 37°C overnight. The kit was used according to manufacturer's
instructions. Briefly, cells were incubated with medium containing
EdU (10 µM) at 37°C for 2 h, then fixed with 4% paraformaldehyde
for 15 min at room temperature. Cells were then stained with the
Click reaction solution at room temperature for 30 min in darkness.
Next, cell nuclei were stained using Hoechst 33342 and images were
captured using a fluorescence microscope (Nikon Corporation) at a
magnification of 100.
Cell cycle analysis
Cells were treated with 5, 7.5 and 10 µM PAB at 37°C
for 48 h. After harvesting and washing with pre-cold PBS, cells
(1×106/ml) were resuspended with cold 70% ethanol and
fixed at 4°C overnight. Cells were washed twice with PBS and
centrifuged at 1,000 × g at 4°C for 5 min to remove residual
ethanol, then stained with 50 µg/ml propidium iodide (PI) and 100
µg/ml RNase solution (Beijing Solarbio Science & Technology
Co., Ltd.) at 37°C for 20 min (25). The cell cycle arrest in
G2-M phase was measured using flow cytometry (FACS
Calibur; BD Biosciences) and analyzed with FlowJo 7.6 software
(FlowJo LLC).
Apoptosis assay
The rate of apoptosis was detected using the annexin
V-FITC/PI apoptosis kit (Vazyme Biotech Co., Ltd.) according to the
manufacturer's instructions. Briefly, MDA-MB-231 cells
(2×105) were seeded in six-well plates overnight, then
cultured with PAB (5, 7.5 and 10 µM), LY294002 (40 µM) or both (7.5
µM PAB + 40 µM LY294002) at 37°C for 48 h. Next, cells were
harvested with EDTA-free trypsin and washed twice with chilled PBS.
Cell precipitates were resuspended with 1X binding buffer and then
stained with 5 µl annexin V-FITC and 5 µl PI staining solution in
the dark for 10 min at room temperature (24,26).
Finally, the rate of apoptosis was analyzed by flow cytometry (FACS
Calibur; BD Biosciences).
Assessment of MMP and (reactive oxygen
species) ROS
Considering that a decline in MMP is a key trigger
to activate the mitochondrial apoptosis pathway, MMP assay kit with
JC-1 was used to detect the MMP level of MDA-MB-231 cells. JC-1
forms aggregates in the mitochondrial matrix when the MMP is high,
resulting in red fluorescence. When the MMP is low, JC-1 cannot
aggregate in the mitochondrial matrix and cells fluoresce is green.
Briefly, after treatment with PAB (5, 7.5 and 10 µM) at 37°C for 48
h, cells were harvested and resuspended in 500 µl JC-1 working
solution, in the darkness at 37°C for 20 min. Cells were then
washed twice with chilled JC-1 staining buffer (1X) and immediately
examined using flow cytometry (FACS Calibur; BD Biosciences).
To assess ROS levels, cells were cultured with PAB
(5, 7.5 and 10 µM) at 37°C for 48 h, then collected and incubated
with 10 µM DCFH-DA fluorescent probe for 20 min at 37°C in the
darkness. The cells were washed twice with FBS-free Leibovitz's
L-15 medium (Procell Life Science & Technology Co., Ltd.) and
the cellular ROS level was detected using flow cytometry (FACS
Calibur; BD Biosciences).
DAPI staining
Cells (1×104/well) were seeded on glass
coverslips in 24-well plates overnight, then treated with PAB (5,
7.5 and 10 µM) at 37°C for 48 h. The cells were fixed with 4%
paraformaldehyde at room temperature for 15 min and permeabilized
with 0.5% Triton X-100 at room temperature for 20 min. After
washing three times with PBS, the cells were stained with
fluorescent dye DAPI in a darkroom at room temperature for 15 min.
The nuclear morphology was observed using a fluorescence microscope
at a magnification of 200.
Transwell migration and invasion
assays
Cell migration and invasion assays were performed
using Transwell chambers with a pore size of 8 µm (27). Matrigel (Corning Biocoat; Corning
Life Sciences) was used for the cell invasion assay but not for the
cell migration assay. A mixture of Martrigel and FBS-free
Leibovitz's L-15 medium was added to the upper chamber and placed
at 37°C for 1 h. Cells were cultured with serum-free Leibovitz's
L-15 medium overnight, then suspended and diluted to a density of
1×105 with various concentrations of PAB (5, 7.5 and 10
µM). A 200 µl cell suspension containing 2×104 cells was
added to the upper chamber on the 24-well plate. The lower chamber
was filled with 500 µl Leibovitz's L-15 medium supplemented with
10% FBS. After incubation at 37°C for 48 h, the cells that passed
through the chambers were fixed with 4% paraformaldehyde at room
temperature for 20 min, stained with 0.1% crystal violet at room
temperature for 30 min and then non-penetrating cells were wiped
off with a cotton swab. Cells were observed under light microscope
at a magnification of 100.
Wound healing assays
MDA-MB-231 cells were grown in six-well plates until
the cell confluence reached 100%. A wound was made by scratching
the adherent cell layer with a 200-µl pipette tip (25). Shed cells were washed off with PBS
and the remaining cells were treated with serum-free Leibovitz's
L-15 medium containing different concentrations of PAB (5, 7.5 and
10 µM) for 48 h. Cells were observed under light microscope at a
magnification of 200. Data analysis was performed using ImageJ
1.8.0 software (National Institutes of Health). The rate of wound
healing=[(the wound width of 0–48 h)/0 h wound width] ×100%.
Western blotting
Protein expression levels were evaluated using
western blotting as previously described (26,28).
Cells were treated with 5, 7.5 and 10 µM PAB at 37°C for 48 h, then
lysed with RIPA buffer (Beyotime Institute of Biotechnology) for
protein extraction. Cells were centrifuged at 10,000 × g for 15 min
at 4°C, and protein concentrations were measured using a BCA
Protein Assay kit (Beyotime Institute of Biotechnology). Protein
samples (30 µg/lane) were separated using 10–12.5% SDS-PAGE gels
before being transferred to the PVDF membrane (MilliporeSigma).
PVDF membranes were horizontally cut to probe proteins with
different molecular weights. After blocking with 5% skim milk
diluted with Tris-buffered saline with Tween (TBST) containing 0.1%
Tween at room temperature for 2 h, the PVDF membranes were
incubated with primary antibodies at 4°C overnight. Blots were
washed three times with TBST and incubated with secondary
antibodies for 2 h at room temperature. Stripping buffer was used
to remove the antigen-antibody complex from the PVDF membrane in
order to re-probe other antibodies on the same membrane. The blots
were measured using SuperFemto ECL Chemiluminescence kit (Vazyme
Biotech Co., Ltd) through the chemiluminescence detection system of
the Amersham Imager 600 (GE Healthcare Production).
Statistical analysis
All data were obtained from three independently
replicated experiments and presented as mean ± standard deviation.
One-way analysis of variance was used to analyze statistical
significance for multiple comparisons, followed by Tukey's multiple
comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
PAB inhibits the proliferation of TNBC
cells
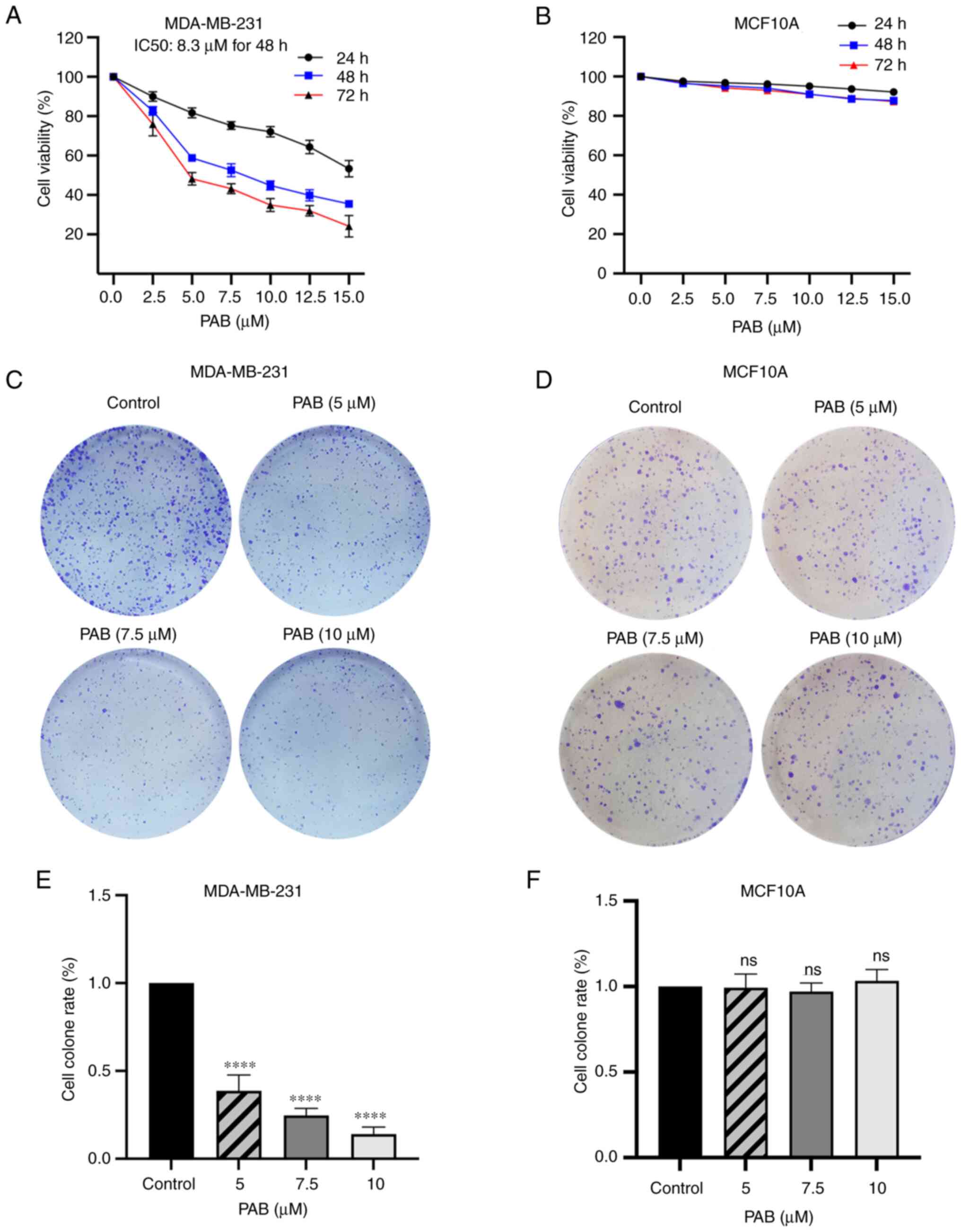
To investigate the effect of PAB on the viability of
TNBC cells, the CCK-8 assay was performed to evaluate the influence
of various concentrations of PAB for 24, 48 and 72 h. PAB inhibited
the proliferation of MDA-MB-231 cells in dose- and time-dependent
manners, with IC50 values of 19.3, 8.3 and 5.76 µM at 24, 48 and 72
h respectively (Fig. 1A). However,
the result of CCK-8 showed that PAB had no obvious side-effects on
normal cells line MCF10A (Fig. 1B).
Colony formation assays further confirmed that PAB suppressed the
proliferation of MDA-MB-231 cells in a dose-dependent manner, but
not MCF10A cells (Fig. 1C-F).
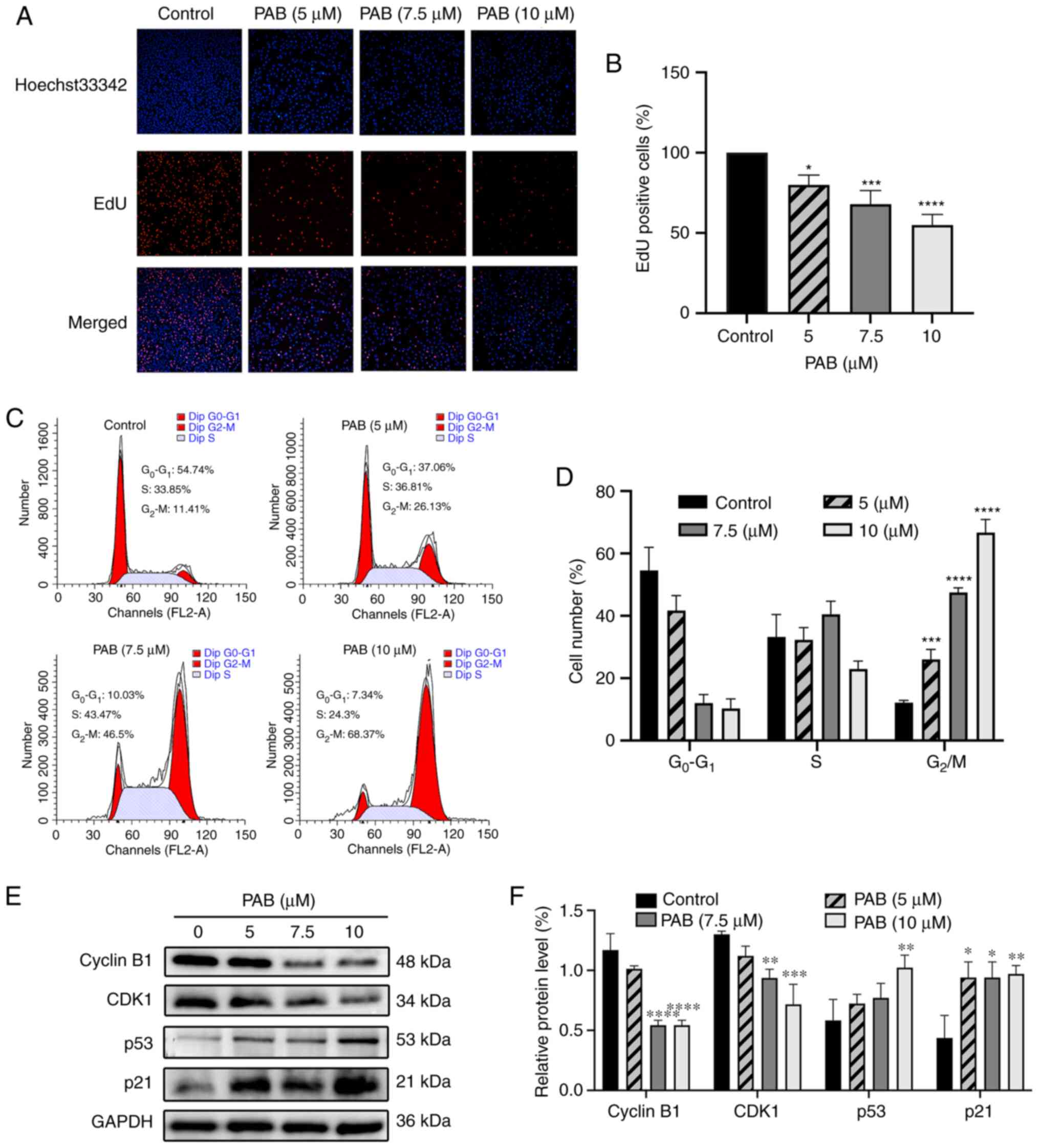
Furthermore, according to the EdU assay, compared with the control,
all three different concentrations (5, 7.5 and 10 µM) of PAB can
significantly reduce the number of EDU positive MDA-MB-231 cells
(Fig. 2A), indicating that PAB
could suppress the proliferative capacity of these cells (Fig. 2B).
G2/M cell cycle arrest of
MDA-MB-231 cells is induced by PAB
The cell cycle distribution was analyzed by flow
cytometry after propidium iodide staining. MDA-MB-231 cells treated
with various concentration of PAB for 48 h were harvested for cell
cycle analysis. As shown in Fig. 2C and
D, it induced a significant increase in the number of cells in
G2/M phase. Meanwhile, western blotting showed that the
protein expression levels of CDK1 and cyclin B1 were significantly
reduced compared with the control, while those of p53 and p21 were
increased after PAB treatment (Fig. 2E
and F). These results indicate that PAB may induce cell cycle
arrest by altering the expression of cell recycle regulators.
PAB induces apoptosis via the
mitochondrial pathway
After treatment with various concentration of PAB
for 48 h, flow cytometry showed that PAB induced apoptosis in
MDA-MB-231 cells in a dose-dependent manner (Fig. 3A and B). The effect of PAB on the
nuclear status of MDA-MB-231 cells was tested using DAPI staining.
Apoptotic cells presented with nuclear condensation and DNA
fragmentation (Fig. 3C).
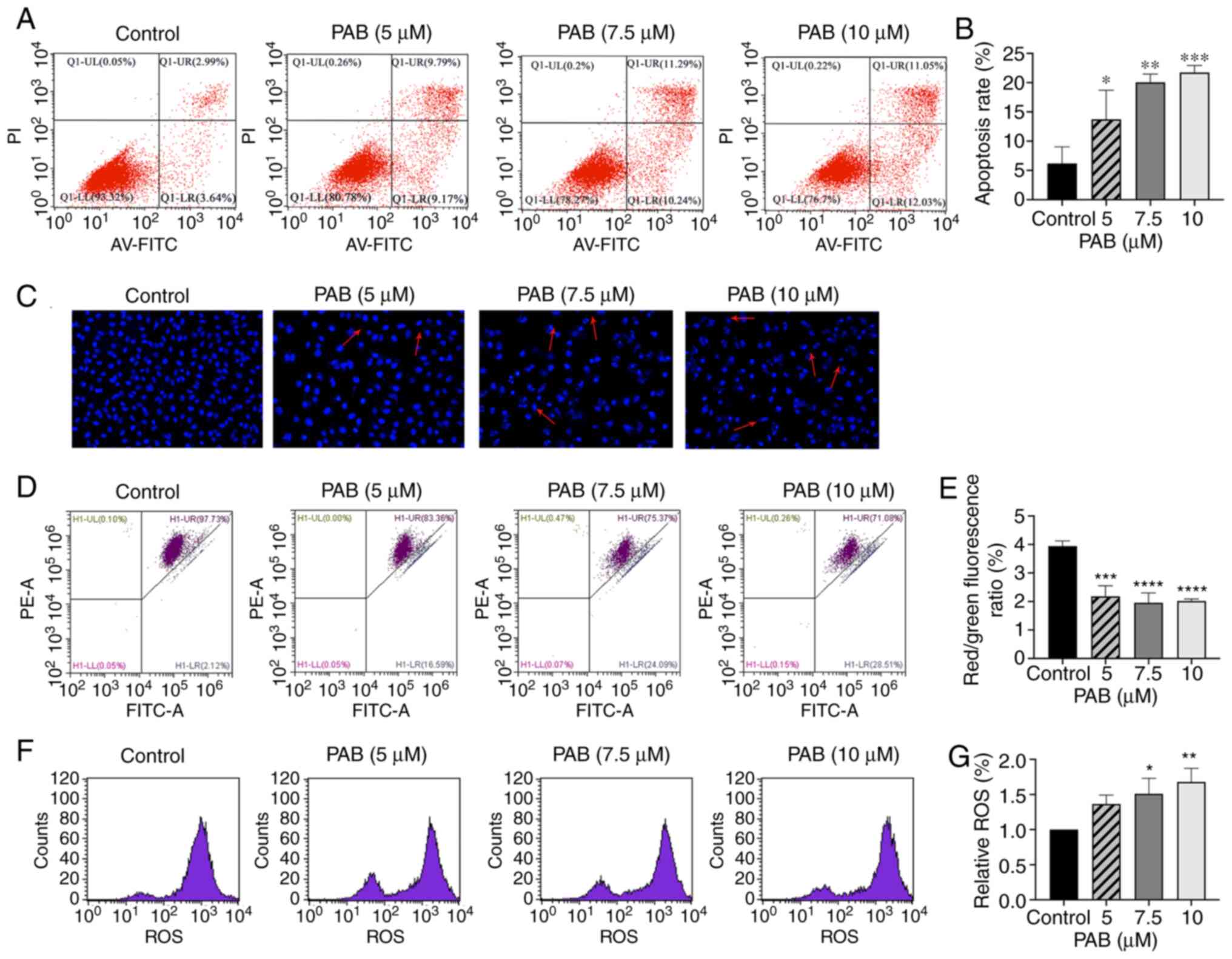 | Figure 3.PAB induces apoptosis in MDA-MB-231
cells. (A) Cells were pretreated with PAB (5, 7.5, 10 µM) for 48 h.
Annexin V-FITC and PI staining were used to identify the apoptosis,
and the data were analyzed by flow cytometry. (B) Quantitative data
of PAB-induced apoptosis. (C) Cell nuclei were observed by confocal
microscope (magnification, 400×) after 48 h of PAB treatment by
DAPI staining. Typical apoptosis morphological changes were shown
in treated cells including chromatin condensation and DNA
fragmentation. (D) Cell mitochondrial membrane potential was
determined using flow cytometry with JC-1 staining after 48 h of
treatment with 0, 5, 7.5 and 10 µM PAB, respectively. (E) Red/green
fluorescence ratio. The ratio of red to green fluorescence
represents the percentage of decreased MMP. (F) Fluorescence in the
cell is represented as the percentage of ROS production (G)
analyzed using flow cytometry. *P<0.05, **P<0.01,
***P<0.001, ****P<0.0001 vs. control. PAB, Pseudolaric acid
B; MMP, mitochondrial membrane potential; ROS, reactive oxygen
species. |
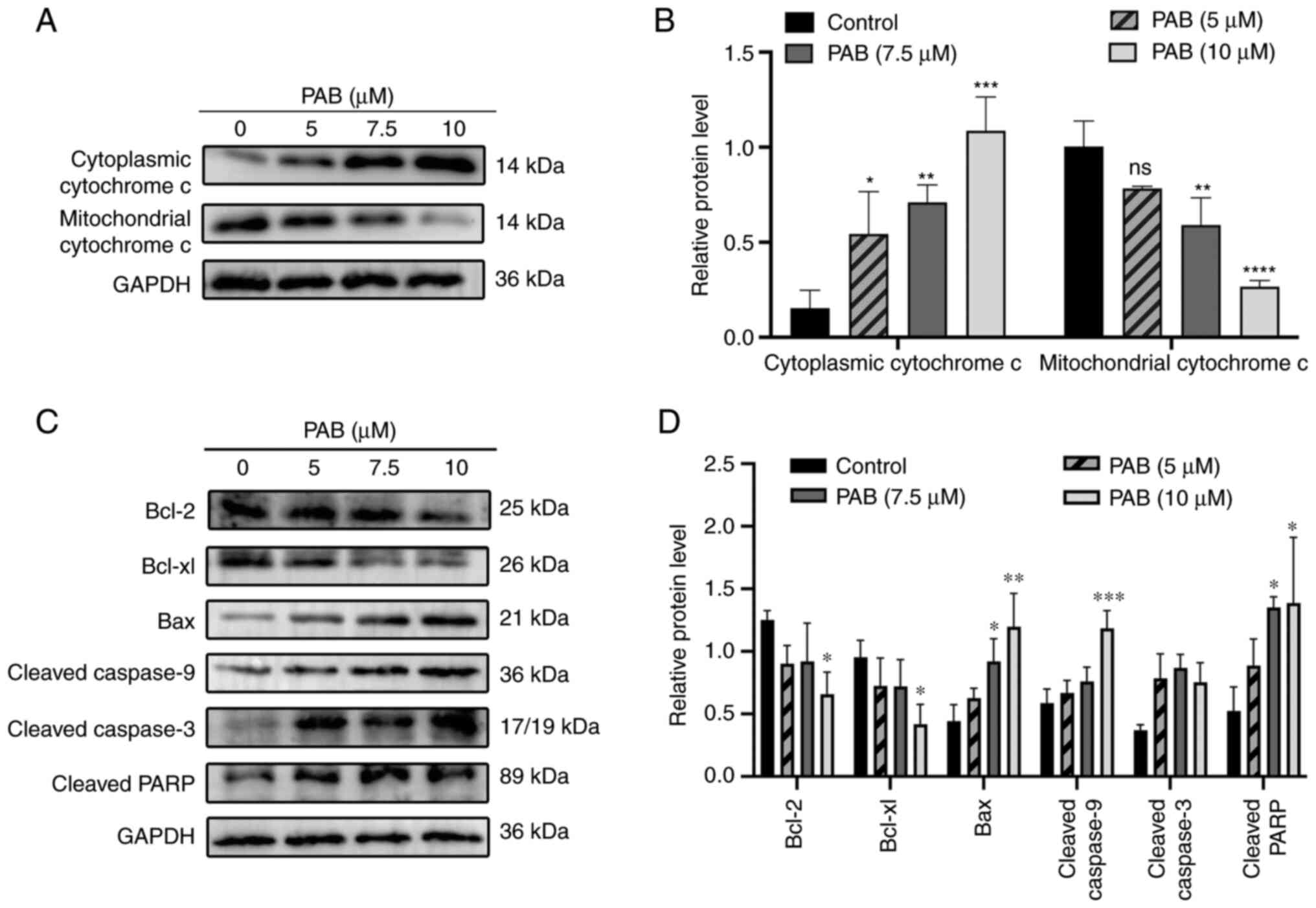
A collapse of MMP is an important factor leading to
apoptosis mediated by the mitochondrial apoptosis pathway. JC-1
staining showed that PAB induced a dose-dependent loss of MMP in
MDA-MB-231 cells (Fig. 3D and E).
To further explore the mechanisms of PAB-induced apoptosis, the
related protein expression was measured and it was revealed that
PAB significantly induced Cytochrome c release from the
mitochondria into the cytosol (Fig. 4A
and B), upregulated the expression of cleaved-caspase3,
cleaved-caspase9, cleaved-PARP and Bax, and downregulated the
expression of Bcl-2 and Bcl-xl (Fig. 4C
and D). All these changes were statistically significant
(P<0.05). Therefore, the results demonstrated that PAB induced
apoptosis mediated by mitochondrial apoptosis pathway in TNBC.
PAB increases ROS levels
It is well known that ROS production is related to
mitochondrial pathway-associated apoptosis. Therefore, ROS
production was detected by the fluorescent probe DCFH-DA. As shown
in Fig. 3F and G, flow cytometry
demonstrated that the ROS accumulation was directly related to PAB
concentration.
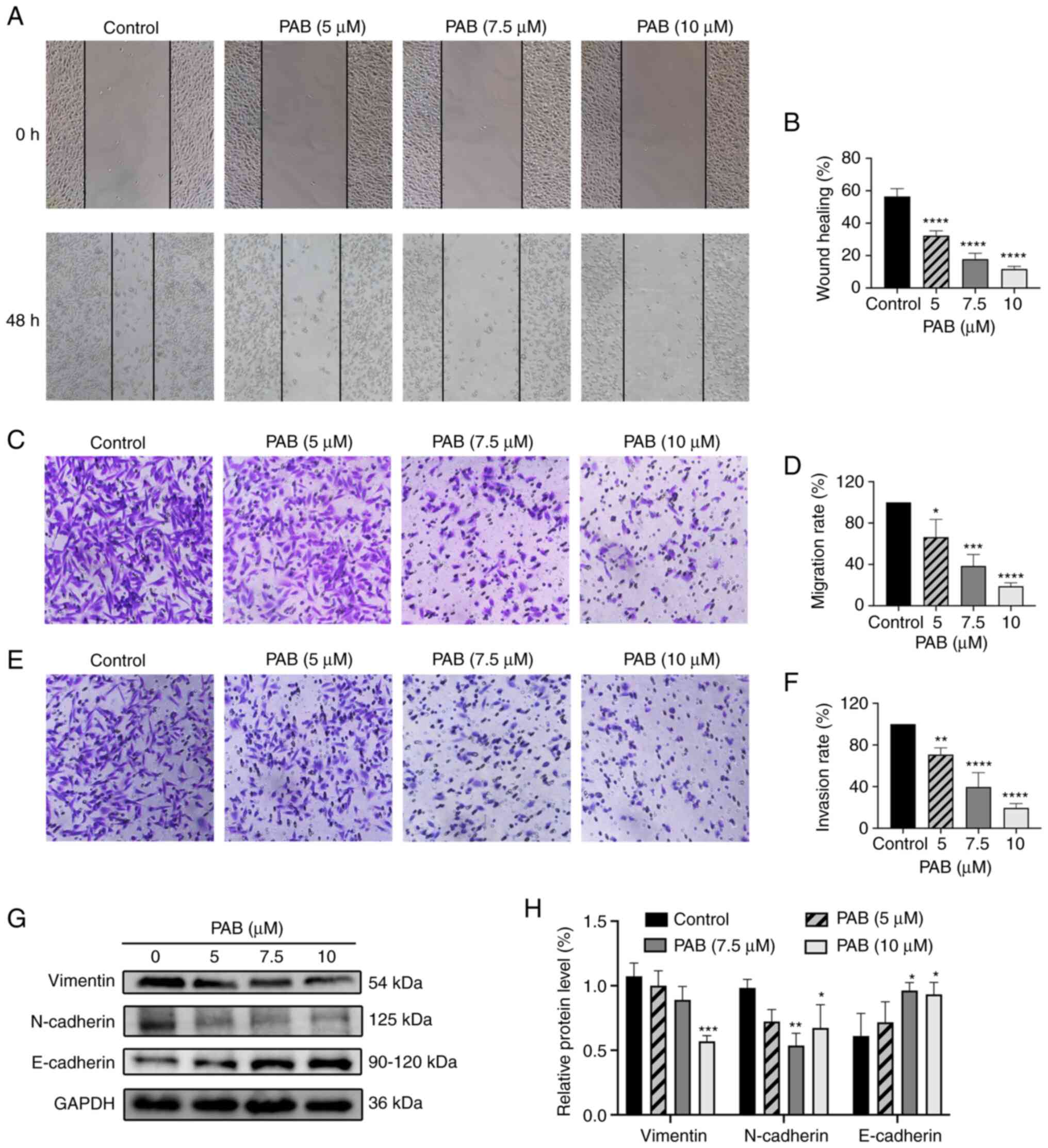
PAB inhibits migration and invasion by
regulating the epithelial-mesenchymal transition (EMT) pathway
Migration of MDA-MB-231 cells was measured by the
wound healing and Transwell migration assays. PAB significantly
inhibited wound healing ability (Fig.
5A and B) and Transwell migration ability (Fig. 5C and D) in a dose-dependent manner.
As shown in Fig. 5E and F, PAB
significantly inhibited cell invasion. Protein levels of N-cadherin
and vimentin were reduced by PAB, while the protein level of
E-cadherin was increased (Fig. 5G and
H). Overall, these data suggested that PAB inhibited migration,
invasion and EMT in MDA-MB-231 cells.
PAB inhibits the PI3K/AKT/mTOR
signaling pathway
To investigate the role of the PI3K/AKT/mTOR
signaling pathway in the anticancer effect of PAB on MDA-MB-231
cells, the activation of PI3K, AKT and mTOR were evaluated by
western blotting. PAB inhibited PI3K (p110β), the phosphorylation
of AKT and the phosphorylation of mTOR in a dose-dependent manner
and PAB did not influence total levels of AKT and mTOR (Fig. 6A and B). Furthermore, the apoptotic
rate of PAB and LY294002 co-treated cells exceeded that of PAB or
LY294002 alone (Fig. 6C and D).
These results suggested that the PI3K/AKT/mTOR signaling pathway
was involved in PAB-induced apoptosis in MDA-MB-231 cells.
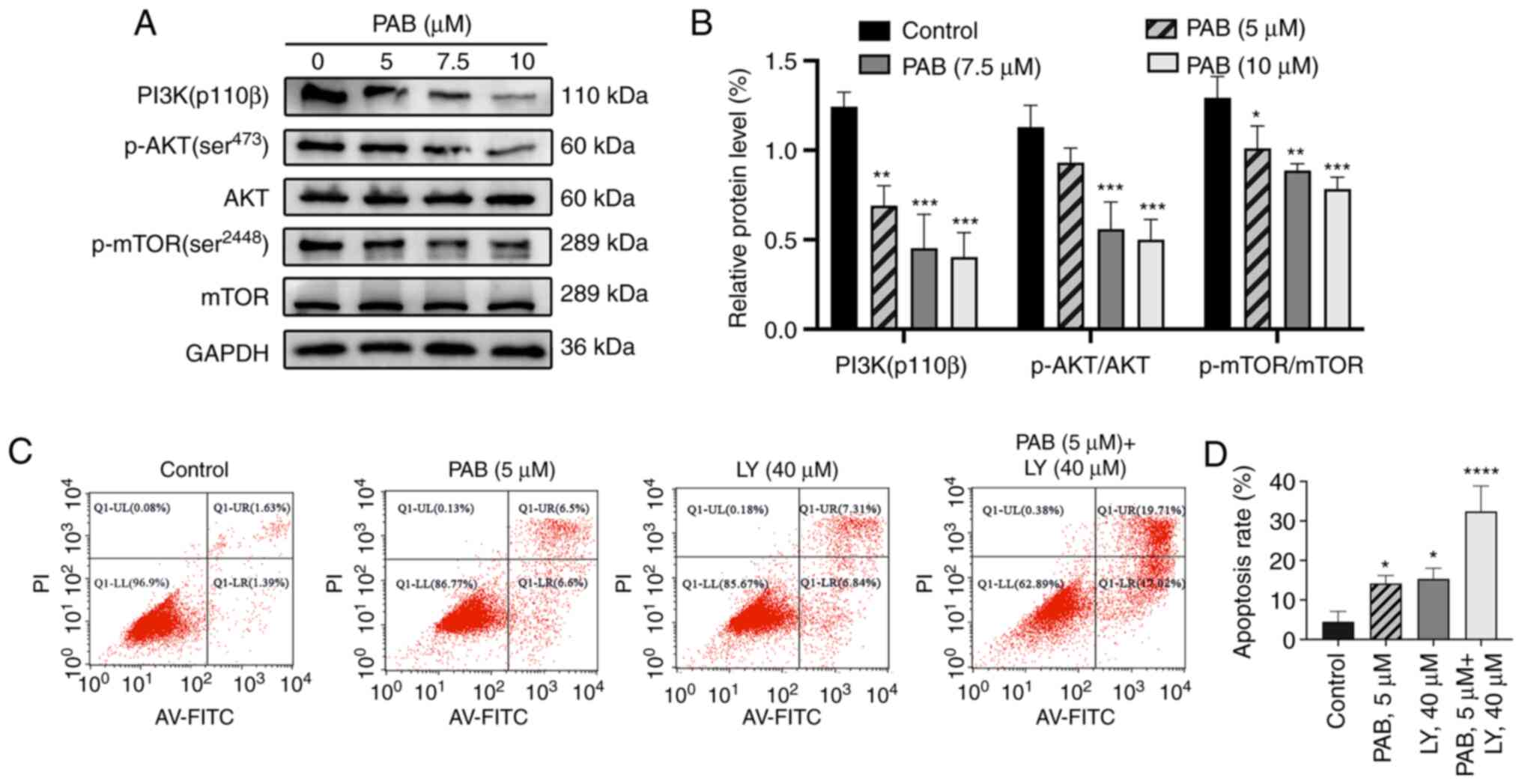 | Figure 6.PAB inhibits the PI3K/mTOR/AKT
signaling pathway. (A) MDA-MB-231 cells were exposed to the
indicated doses of PAB for 48 h. The expression levels of PI3K,
p-AKT (ser473), AKT, p-mTOR (ser3448) and mTOR were assessed by
western blotting. (B) Quantitative data of the relative protein
expression. (C) Cells were pretreated with PAB (5 µM), LY294002 (40
µM) or both for 48 h. Annexin V-FITC and PI staining were used to
identify apoptosis rate, and (D) the data were analyzed by flow
cytometry. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001
vs. control. PAB, Pseudolaric acid B; p-, phosphorylated. |
Discussion
PAB, the main medicinal component of Cortex
pseudolaricis, is a natural plant product with potential
antifungal, immunosuppressive and anticancer properties. Previous
studies have shown that PAB has multi-target anticancer effects in
different types of tumors (9,29–31).
To investigate the anticancer properties and mechanisms of PAB on
TNBC, the present study investigated the effects of PAB on the
proliferation, apoptosis, invasion and migration of MDA-MB-231
cells. CCK-8, EdU and colony formation assays demonstrated that PAB
inhibited the proliferation of MDA-MB-231 cells in a dose- and
time-dependent manner.
Excessive cell division is an important reason for
the continuous proliferation of tumor cells (32). Recently, researchers have indicated
that numerous natural drugs exert their anticancer effects by
regulating the cycle checkpoints, which can lead to arrest of the
tumor cell cycle (33–36). Among these checkpoints, cyclin B1
and CDK1 are necessary for the cell cycle to switch from the S to
the G2/M phase (37).
The tumor suppressor p53 induces G2/M phase arrest by
activating the downstream transcriptional target p21, which is a
CDK inhibitor (38,39). The present results indicated that
PAB induced G2/M phase arrest of MDA-MB-231 cells by
upregulating the expression of p53 and p21, resulting in the
downregulation of cyclin B1 and CDK1 proteins.
Flow cytometry showed that PAB induced apoptosis in
MDA-MB-231 cells in a concentration-dependent manner. Mitochondria
are the hubs for cellular energy metabolism, and mitochondrial
dysfunction is the main cause for activation of the mitochondrial
apoptosis pathway (40). ROS are
mainly produced in mitochondria, and its excessive accumulation can
damage mitochondria and activate the mitochondria-mediated
intrinsic apoptotic pathway (41).
To further investigate the mechanism of apoptosis, nuclear
morphology, MMP level, ROS levels and protein expression were
evaluated. DAPI nuclear staining indicated that PAB led to
chromatin condensation, cellular shrinkage and DNA fragmentation.
It also caused a decrease of MMP and an increase of ROS.
Furthermore, western blotting showed that it upregulated the
proapoptotic protein Bax and downregulated the antiapoptotic
proteins Bcl-2 and Bcl-xl. Cytochrome c is released from
mitochondria into the cytosol and subsequently activates caspase-9
and caspase-3, which help to initiate the mitochondrial apoptosis
pathway (42). The present
experiments showed that PAB could promote the release of cytochrome
c from mitochondria into the cytoplasm. In addition, levels of
cleaved caspase-3 and caspase-9, as well as PARP, were increased.
Western blotting further demonstrated that it could downregulate
the anti-apoptosis proteins Bcl-2 and Bcl-xl and increased the
pro-apoptosis protein Bax. These results suggested that PAB induced
apoptosis of MDA-MB-231 cells via the mitochondrial apoptosis
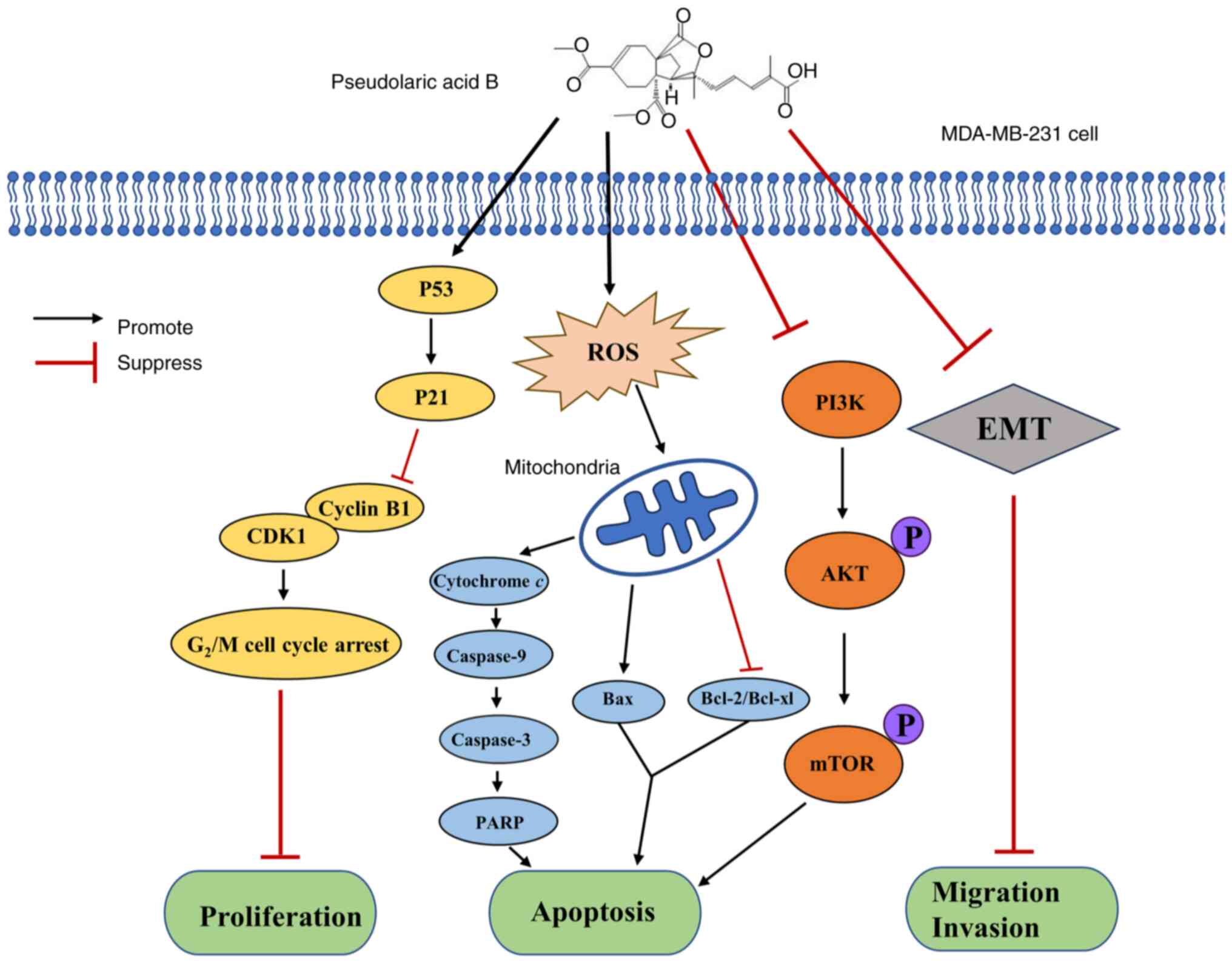
pathway (Fig. S1).
The PI3K/AKT/mTOR signaling pathway, an important
regulator of tumor proliferation, apoptosis, invasion and
migration, is closely related to the expression of Bcl-2 family
proteins in the mitochondrial apoptosis pathway (22). Western blotting showed that
treatment of TNBC cells with PAB significantly decreased levels of
PI3K (p110β), p-AKT and p-mTOR. Its apoptosis effect was also
significantly elevated when combined with LY294002, a PI3K
inhibitor. These data demonstrated that PI3K/AKT/mTOR signaling may
be the target of PAB-induced apoptosis in MDA-MB-231 cells.
TNBC is the most aggressive and malignant type of
breast cancer, and is more likely to develop lung and brain
metastases (43–45). The present study revealed that EMT
is critical for regulating the proliferation, invasion and
metastasis of carcinoma cells (46). Activation of the EMT mechanism
depends on loss of the epithelial marker E-cadherin and the
upregulation of mesenchymal markers N-cadherin and vimentin,
thereby prompting tumor cells through multiple steps in the process
of invasion and metastasis (47,48).
Results of the present wound-healing and Transwell assays suggested
that PAB inhibited the ability of cells to migrate and invade.
Further experiments demonstrated that it decreased the levels of
N-cadherin and vimentin and increased the level of E-cadherin.
These results suggested that EMT in MDA-MB-231 cells could be
inhibited by PAB treatment.
There are several limitations and challenges to the
present research. First, it was only verified that the mechanism of
PAB-induced apoptosis in MDA-MB-231 cells was related to the
mitochondrial apoptosis pathway, but it was not verified whether it
was dependent on the mitochondrial apoptosis pathway. Second, the
present study demonstrated that the PI3K/AKT/mTOR signaling pathway
had a superpositioned effect on PAB-induced apoptosis, but the
specific mechanism of action has not been clearly explored. Future
studies will hopefully explore the specific role of PI3K/AKT/mTOR
signaling pathway in PAB-induced apoptosis. Third, although the aim
of the present study was to demonstrate how PAB plays an anticancer
role in TNBC, the specific mechanism of PAB in vivo studies
remains to be determined. In addition, a previous study has
revealed that PAB-induced autophagy of breast cancer cells line
MCF-7 inhibits apoptosis and promotes cell survival, which
indicates that the combination of autophagy inhibitors may improve
the anticancer effect of PAB (49).
In conclusion, the present results demonstrated that
PAB exhibited anticancer effects against TNBC and that the
mechanism was related to multiple pathways (Fig. 7). The present study revealed that
PAB significantly inhibited the proliferative ability of MDA-MB-231
cells by arresting the cell at the G2/M phase. The
pro-apoptotic activity of PAB in TNBC was demonstrated through
activation of the mitochondrial apoptosis pathway and inhibition of
the PI3K/AKT/mTOR signaling pathway. PAB also demonstrated an
anticancer effect on TNBC by inhibiting cell migration and
invasion, through a mechanism related to the suppression of EMT.
Overall, these results provided evidence that PAB exerted multiple
anticancer activities through multiple targets in TNBC.
Supplementary Material
Supporting Data
Acknowledgements
The group would like to thank Professor Caigang Liu
from Shengjing Hospital Cancer Research Center (Shenyang, China)
for the cell support. The group would like to thank Professor Yuxin
Tong, Medical Research Center, Shengjing Hospital of China Medical
University (Shenyang, China) for technical guidance.
Funding
This work was supported by a grant from the Science and
Technology Project of Liaoning Province (grant no. 2014226033).
Availability of data and materials
The data used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
FY, SNC and KL designed the study and revised the
manuscript. KY and JQW performed the experiments and drafted the
manuscript together. All authors read and approved the final
manuscript. KY and JQW confirm the authenticity of all the raw
data.
Ethical approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yin L, Duan JJ, Bian XW and Yu SC:
Triple-negative breast cancer molecular subtyping and treatment
progress. Breast Cancer Res. 22:612020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Garrido-Castro AC, Lin NU and Polyak K:
Insights into molecular classifications of triple-negative breast
cancer: Improving patient selection for treatment. Cancer Discov.
9:176–198. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu J, Guan H, Wu D, Hu Z, Zhang H, Jiang
H, Yu J, Zeng K, Li H, Zhang H, et al: Pseudolaric acid B
ameliorates synovial inflammation and vessel formation by
stabilizing PPARγ to inhibit NF-κB signalling pathway. J Cell Mol
Med. 25:6664–6678. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li Z, Yin H, Chen W, Jiang C, Hu J, Xue Y,
Yao D, Peng Y and Hu X: Synergistic effect of pseudolaric Acid B
with fluconazole against resistant isolates and biofilm of candida
tropicalis. Infect Drug Resist. 13:2733–2743. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miao ZH, Feng JM and Ding J: Newly
discovered angiogenesis inhibitors and their mechanisms of action.
Acta Pharmacol Sin. 33:1103–11. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wei SF, He DH, Zhang SB, Lu Y, Ye X, Fan
XZ, Wang H, Wang Q and Liu YQ: Identification of pseudolaric acid B
as a novel Hedgehog pathway inhibitor in medulloblastoma. Biochem
Pharmacol. 190:1145932021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mafu S, Karunanithi PS, Palazzo TA, Harrod
BL, Rodriguez SM, Mollhoff IN, O'Brien TE, Tong S, Fiehn O,
Tantillo DJ, et al: Biosynthesis of the microtubule-destabilizing
diterpene pseudolaric acid B from golden larch involves an unusual
diterpene synthase. Proc Natl Acad Sci USA. 114:974–979. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang H, Li JC, Luo H, Zhao L, Zhang ZD
and Shen XF: Pseudolaric acid B exhibits anti-cancer activity on
human hepatocellular carcinoma through inhibition of multiple
carcinogenic signaling pathways. Phytomedicine. 59:1527592019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang D, Xin Y, Tian Y, Li W, Sun D and
Yang Y: Pseudolaric acid B inhibits gastric cancer cell metastasis
in vitro and in haematogenous dissemination model through PI3K/AKT,
ERK1/2 and mitochondria-mediated apoptosis pathways. Exp Cell Res.
352:34–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yao GD, Yang J, Li XX, Song XY, Hayashi T,
Tashiro SI, Onodera S, Song SJ and Ikejima T: Blocking the
utilization of glucose induces the switch from senescence to
apoptosis in pseudolaric acid B-treated human lung cancer cells in
vitro. Acta Pharmacol Sin. 38:1401–1411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang L, Wen C, He Q, Sun Y, Wang J, Lan
X, Rohondia S, Dou QP, Shi X and Liu J: Pseudolaric acid B induces
mitotic arrest and apoptosis in both imatinib-sensitive and
-resistant chronic myeloid leukaemia cells. Eur J Pharmacol.
876:1730642020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hanahan D: Hallmarks of cancer: New
dimensions. Cancer Discov. 12:31–46. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jeong SY and Seol DW: The role of
mitochondria in apoptosis. BMB Rep. 41:11–22. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Carneiro BA and El-Deiry WS: Targeting
apoptosis in cancer therapy. Nat Rev Clin Oncol. 17:395–417. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guan D, Li C, Lv X and Yang Y: Pseudolaric
acid B inhibits PAX2 expression through Wnt signaling and induces
BAX expression, therefore promoting apoptosis in HeLa cervical
cancer cells. J Gynecol Oncol. 30:e772019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Choi SJ, Ahn CH, Yang IH, Jin B, Lee WW,
Kim JH, Ahn MH, Swarup N, Hong KO, Shin JA, et al: Pseudolaric Acid
B induces growth inhibition and caspase-dependent apoptosis on head
and neck cancer cell lines through death receptor 5. Molecules.
24:37152019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen C, Chen J, Zhang D, Wang H, Che J, Qin
Q, He L, Cai Z, Lin M, Lou Q, et al: Pseudolaric acid B induces
mitotic arrest and apoptosis in both 5-fluorouracil-sensitive and
-resistant colorectal cancer cells. Cancer Lett. 383:295–308. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Polivka J Jr and Janku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bartholomeusz C and Gonzalez-Angulo AM:
Targeting the PI3K signaling pathway in cancer therapy. Expert Opin
Ther Targets. 16:121–130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Claerhout S, Decraene D, Van Laethem A,
Van Kelst S, Agostinis P and Garmyn M: AKT delays the
early-activated apoptotic pathway in UVB-irradiated keratinocytes
via BAD translocation. J Invest Dermatol. 127:429–438. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Quan JH, Cha GH, Zhou W, Chu JQ, Nishikawa
Y and Lee YH: Involvement of PI 3 kinase/Akt-dependent Bad
phosphorylation in Toxoplasma gondii-mediated inhibition of host
cell apoptosis. Exp Parasitol. 133:462–471. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fulda S: Synthetic lethality by
co-targeting mitochondrial apoptosis and PI3K/Akt/mTOR signaling.
Mitochondrion. 19:85–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zuo A, Zhao P, Zheng Y, Hua H and Wang X:
Tripterine inhibits proliferation, migration and invasion of breast
cancer MDA-MB-231 cells by up-regulating microRNA-15a. Biol Chem.
400:1069–1078. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang W, Feng B, Meng Y, Wang J, Geng B,
Cui Q, Zhang H, Yang Y and Yang J: FAM3C-YY1 axis is essential for
TGFβ-promoted proliferation and migration of human breast cancer
MDA-MB-231 cells via the activation of HSF1. J Cell Mol Med.
23:3464–3475. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lou C, Xu X, Chen Y and Zhao H: Alisol A
suppresses proliferation, migration, and invasion in human breast
cancer MDA-MB-231 cells. Molecules. 24:36512019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kheirandish-Rostami M, Roudkenar MH,
Jahanian-Najafabadi A, Tomita K, Kuwahara Y, Sato T and Roushandeh
AM: Mitochondrial characteristics contribute to proliferation and
migration potency of MDA-MB-231 cancer cells and their response to
cisplatin treatment. Life Sci. 244:1173392020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tohkayomatee R, Reabroi S, Tungmunnithum
D, Parichatikanond W and Pinthong D: Andrographolide exhibits
anticancer activity against breast cancer cells (MCF-7 and
MDA-MB-231 cells) through suppressing cell proliferation and
inducing cell apoptosis via inactivation of ER-α receptor and
PI3K/AKT/mTOR signaling. Molecules. 27:35442022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu B, Yue DM, Shu LH, Li NJ and Wang JH:
Pseudolaric acid B induces caspase-dependent cell death in human
ovarian cancer cells. Oncol Rep. 31:849–857. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang D, Tian Y, Feng W, Zhao L, Zhao M,
Liu J and Wang Q: Pseudolaric acid B induces endometrial cancer
Ishikawa cell apoptosis and inhibits metastasis through AKT-GSK-3β
and ERK1/2 signaling pathways. Anticancer Drugs. 28:603–612. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li X, Zhao X, Song W, Tian Z, Yang L, Niu
Q, Zhang Q, Xie M, Zhou B, Xu Y, et al: Pseudolaric Acid B inhibits
proliferation, invasion and Epithelial-to-mesenchymal transition in
human pancreatic cancer cell. Yonsei Med J. 59:20–27. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Matthews HK, Bertoli C and de Bruin RAM:
Cell cycle control in cancer. Nat Rev Mol Cell Biol. 23:74–88.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dong C, Wen S, Zhao S, Sun S, Zhao S, Dong
W, Han P, Chen Q, Gong T, Chen W, et al: Salidroside inhibits
reactive astrogliosis and glial scar formation in late cerebral
ischemia via the Akt/GSK-3β pathway. Neurochem Res. 46:755–769.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bao Y, Wu X, Jin X, Kanematsu A, Nojima M,
Kakehi Y and Yamamoto S: Apigenin inhibits renal cell carcinoma
cell proliferation through G2/M phase cell cycle arrest. Oncol Rep.
47:602022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He YC, He L, Khoshaba R, Lu FG, Cai C,
Zhou FL, Liao DF and Cao D: Curcumin nicotinate selectively induces
cancer cell apoptosis and cycle arrest through a P53-mediated
mechanism. Molecules. 24:41792019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma K, Wang K, Zhou Y, Liu N, Guo W, Qi J,
Hu Z, Su S, Tang P and Zhou X: Purified vitexin compound 1 serves
as a promising antineoplastic agent in ovarian cancer. Front Oncol.
11:7347082021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xie B, Wang S, Jiang N and Li JJ: Cyclin
B1/CDK1-regulated mitochondrial bioenergetics in cell cycle
progression and tumor resistance. Cancer Lett. 443:56–66. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fischer M, Quaas M, Steiner L and Engeland
K: The p53-p21-DREAM-CDE/CHR pathway regulates G2/M cell cycle
genes. Nucleic Acids Res. 44:164–174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Engeland K: Cell cycle arrest through
indirect transcriptional repression by p53: I have a DREAM. Cell
Death Differ. 25:114–132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Abate M, Festa A, Falco M, Lombardi A,
Luce A, Grimaldi A, Zappavigna S, Sperlongano P, Irace C, Caraglia
M and Misso G: Mitochondria as playmakers of apoptosis, autophagy
and senescence. Semin Cell Dev Biol. 98:139–153. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang Y, Karakhanova S, Hartwig W, D'Haese
JG, Philippov PP, Werner J and Bazhin AV: Mitochondria and
mitochondrial ROS in cancer: Novel targets for anticancer therapy.
J Cell Physiol. 231:2570–2581. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cao K and Tait SWG: Apoptosis and cancer:
Force awakens, phantom menace, or both? Int Rev Cell Mol Biol.
337:135–152. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Deepak KGK, Vempati R, Nagaraju GP, Dasari
VR, S N, Rao DN and Malla RR: Tumor microenvironment: Challenges
and opportunities in targeting metastasis of triple negative breast
cancer. Pharmacol Res. 153:1046832020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hosonaga M, Saya H and Arima Y: Molecular
and cellular mechanisms underlying brain metastasis of breast
cancer. Cancer Metastasis Rev. 39:711–720. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rakha EA and Chan S: Metastatic
triple-negative breast cancer. Clin Oncol (R Coll Radiol).
23:587–600. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang Y and Weinberg RA:
Epithelial-to-mesenchymal transition in cancer: Complexity and
opportunities. Front Med. 12:361–373. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Aiello NM and Kang Y: Context-dependent
EMT programs in cancer metastasis. J Exp Med. 216:1016–1026. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yu J, Chen C, Xu T, Yan M, Xue B, Wang Y,
Liu C, Zhong T, Wang Z, Meng X, et al: Pseudolaric acid B activates
autophagy in MCF-7 human breast cancer cells to prevent cell death.
Oncol Lett. 11:1731–1737. 2016. View Article : Google Scholar : PubMed/NCBI
|