Introduction
In older-aged patients with COVID 19, pre-existing
dementia is a major risk factor for the severity of COVID-19
infection (1,2). Apolipoprotein E (APOE) e4 genotype,
which increases the risk of Alzheimer's disease (AD), has predicted
severe COVID-19 infection in one UK Biobank (UKB) cohort (3).
Single nucleotide polymorphisms (SNPs) in the
bridging integrator 1 (BIN1) gene exhibit the second highest odds
ratios for sporadic AD, exceeded only by APOE variants. The most
frequently reported BIN1 AD risk variant is the SNP rs744373, which
has a global allele frequency of 37% and is associated with an
increase in AD risk with odds-ratio of 1.17-1.19(4).
In the present study, UK Biobank (UKB)-derived data
were used to evaluate the effects of BIN1 and the SNP rs744373 on
the survival of patients with COVID-19(5). In addition, the interaction and
alignment of BIN1 and SARS-Cov-2 were evaluated.
Data and methods
UKB
The UKB databases consist of data obtained from
>500,000 community volunteers between 40-70 years of age at
baseline (2006-2010), residing close to 22 assessment centers in
the UK, Scotland and Wales. Baseline assessments on the database
include demographics, the lifestyle of the subjects and the disease
history of the subjects, with links available leading to electronic
medical records. The UKB application for the present study was
approved as UKB project 57245 (S.L. and P.H.R.).
Electronic links regarding age between the UKB
records and the COVID-19 laboratory test results from the National
Health Service in the UK are available from March 16 to April 26,
2020, including the peak of daily COVID-19 laboratory-confirmed
cases due to the current pandemic. During this time period, the
testing of subjects in older-aged groups was largely restricted to
hospital inpatients exhibiting any clinical indications of
infection; thus, test positivity is considered a good marker of
severe COVID-19 infection (5).
The present study analyzed baseline (2006-2010)
demographic characteristics and data for pre-existing diagnoses
during follow-up available on the UKB database. No data from
COVID-19 testing were available for the UKB assessment centers in
Scotland and Wales. The present study included data from subjects
who were alive and who tested positive for COVID-19, as well as
data from and deceased subjects who had tested positive for
COVID-19.
Data of medical diagnoses were available from
baseline questionnaires (2006-2010) that recorded reports of the
disease of participants diagnosed by doctors/medical professionals.
Data for diagnoses of new disease since baseline were from linked
electronic medical records to hospital inpatient routine data to
March 2017, which were coded according to the International
Classification of Diseases 10th revision (ICD-10; https://www.cdc.gov/nchs/icd/icd10.htm).
The diagnoses which were considered were coronary heart disease
(CHD) and AD. The effects of hypertension could not be evaluated,
as only one hypertensive subject was available in the patient
sample examined. Autopsy data were not available and parts of the
respiratory system affected could not be evaluated. In addition,
the expression of BIN1 rs744373 in the lung tissue of
newly-infected patients with CHD could not be evaluated.
Data analysis
Data processing was performed on Minerva, a Linux
mainframe with Centos 7.6, at the Icahn School of Medicine at Mount
Sinai. The present study used PLINK, a whole-genome association
analysis toolset, to process the UKB chromosome 2 files (6). The present study also used The UK
Biobank Data Parser (ukbb_parser), a python-based package that
allows easy interfacing with the large UK Biobank dataset (7). The major (non-Alzheimer's) BIN1
allele was designated as BIN and the SNP rs744373 minor
(Alzheimer's) allele as RS7.
Molecular structure analysis
To evaluate the interaction and alignment of BIN1
and SARS-Cov-2, Protein Data Bank (pdb) entries were searched on
the RCSB Protein Data Bank. In total, 4 entries were identified
that allowed us to examine the association between BIN1 and
SARS-Cov2 as follows: i) 6YUN: Crystal Structure of C-terminal
Dimerization Domain of Nucleocapsid Phosphoprotein from SARS-CoV-2
(deposited, April 27, 2020; released, May 20, 2020; method, X-ray
diffraction; resolution, 1.44 Å) (8). ii) 7K7P: Structure of SARS-CoV-2
non-stuctural protein 1 (NSP1) (deposited September 23, 2020;
released, September 30, 2020; method, X-ray diffraction;
resolution, 1.77 Å). iii) 6WEY: High-resolution structure of the
SARS-CoV-2 NSP3 Macro X domain (deposited, April 3, 2020; released,
April 29, 2020; method, X-ray diffraction; resolution, 0.95 Å
(9). iv) 2FIC: Crystal structure
of the BAR domain from human Bin1/Amphiphysin II (deposited,
December 29, 2005; released, November 14, 2006; method, X-ray
diffraction; resolution, 1.99 Å (10). BAR domains are found in proteins
that bind and remodel membranes and participate in cytoskeletal and
nuclear processes. Amphiphysin II is a protein that in humans is
encoded by the BIN1 gene. The protein structures were superimposed
and aligned with Pymol v 2.3.4.
Statistical analysis
Statistical analysis was performed using Fisher's
exact test and logistic regression analysis and using SPSS v22
software. Data are from UK Biobank. Values of P<0.05 were
considered to indicate statistically significant differences.
Results
The present study analyzed data from 619 subjects
from the UKB database (49% female, 51% male). The mean age was
54±9.2 years (means ± SD). In total, 115 (18.57%) patients
succumbed to COVID-19 infection. A total of 10 subjects (1.6%) had
AD, 33 (5.33%) had CHD (CHD) and 1 subject had hypertension
(Table I).
 | Table IDemographics of the patient
sample. |
Table I
Demographics of the patient
sample.
| Demographic | No./percentage of
subjects |
|---|
| Total no. of
subjects | 619 |
| Female | 49% |
| Male | 51% |
| Age, years (mean ±
SD) | 54±9.2 |
| Deceased due to
COVID-19 | 115 (18.5%) |
| Alzheimer's
disease | 10 (1.6%) |
| Coronary heart
disease | 33 (5.3%) |
| Hypertension | 1 |
Data pertaining to COVID-19-related survival vs. the
BIN1 genotype are presented in Table
II. The effect of genotype on survival was significant
(P=0.004, two-tailed Fisher's exact test). The BIN RS7 heterozygote
was associated with the lowest mortality rate (11.7%), followed by
the BIN BIN homozygote (17.2%). The RS7 RS7 homozygote was
associated with the highest mortality rate (28.1%).
| Table IICOVID-19 survival versus BIN1
genotype. |
Table II
COVID-19 survival versus BIN1
genotype.
| Genotype | Alive | Deceased | Total | % Deceased |
|---|
| BIN BIN | 308 | 64 | 372 | 17.2 |
| BIN RS7 | 250 | 33 | 283 | 11.7 |
| RS7 RS7 | 46 | 18 | 64 | 28.1 |
A total of 2 COVID-19 outcomes were analyzed: i)
Alive with test positivity; and ii) deceased with test-confirmed
positivity for COVID-19. The COVID-19 outcome by multiple factors
is presented in Table III.
Logistic regression analysis was performed using survival (alive)
or mortality (deceased) as dependent variables, and sex, age,
genotype, AD and CHD as independent variables. The effects of sex,
age and genotype were significant at the 95% level. Males and
older-aged subjects with test-confirmed COVID-19 positivity were
more likely to not survive than females and younger subjects. The
effects of AD and CHD were insignificant.
| Table IIICOVID-19-related survival. |
Table III
COVID-19-related survival.
| Demographic | OR | 95% LB | 95% UB | P-value |
|---|
| Sex | 1.681 | 1.046739 | 2.702886 | 0.032 |
| Age | 1.185 | 1.144216 | 1.227869 | <0.001 |
| Genotype | 1.628 | 1.132469 | 2.338711 | 0.008 |
| AD | 1.178 | 0.292433 | 4.76606 | 0.816 |
| CHD | 0.989 | 0.419169 | 2.333758 | 0.981 |
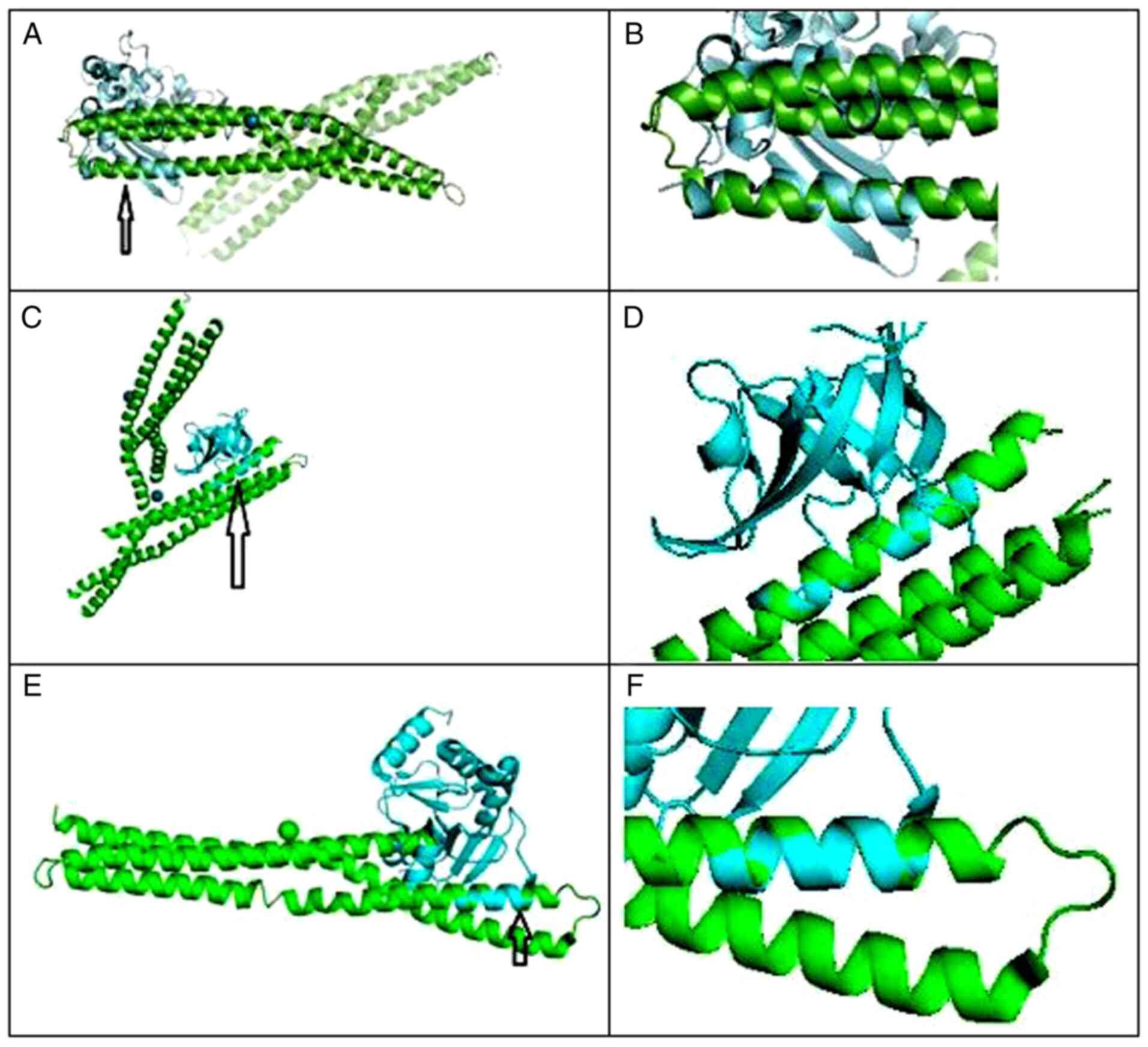
For BIN1 and Crystal Structure of C-terminal
Dimerization Domain of Nucleocapsid Phosphoprotein from SARS-CoV-2,
Pymol was used to perform 6 cycles of calculations on 119 atoms
aligned, final root mean square deviation of atomic positions
(RMSD)=1.850 Å for 101 atoms. Amino acid residues serine
81-tyrosine 100 of BIN1 aligned closely with residues asparagine
99-proline 118 of SARS-Cov2 (Fig.
1A and B).
For BIN1 and Structure of SARS-CoV-2 NSP1, Pymol was
used to perform 6 cycles of calculations on 76 atoms aligned, final
root mean square deviation of atomic positions (RMSD)=0.208 Å for
64 atoms. Amino acid residues glutamic acid 229-asparagine 243 of
BIN1 aligned closely with residues valine 26-glycine 40 of
SARS-Cov2 NSP1 (Fig. 1C and
D).
For BIN1 and Structure of SARS-CoV-2 NSP3, Pymol was
used to perform 5 cycles of calculations on 119 atoms aligned,
final root mean square deviation of atomic positions (RMSD)=0.795 Å
for 79 atoms. Amino acid residues glutamic acid 168-valine 158 of
BIN1 aligned closely with residues valine 267-arginine 277 of
SARS-Cov2 NSP3 (Fig. 1E and
F).
Discussion
BIN1 was first identified as a tumor suppressor and
a mediator of muscle development (4). BIN1 has been shown to exert
protective effects against small cell lung cancer (11), and is moderately expressed in
pneumocytes. The interaction between BIN1 and SARs-Cov2 could be
related in part to the alignment of α helices shown in Fig. 1 and may explain the increased
survival of the BIN RS7 heterozygotes.
The decreased mortality of the heterozygotes (11.7%)
compared to the BIN BIN homozygotes (17.2%) (Table II) may be due to heterozygote
advantage (12). The
heterozygous genotype has a higher relative fitness than either the
homozygous dominant or homozygous recessive genotypes. In humans,
heterozygotes for sickle cell trait are resistant to malaria.
Heterozygotes for cystic fibrosis may be more resistant to
tuberculosis and diarrheal diseases. A significantly lower
proportion of HLA-DRB1 heterozygosity exists among Hepatitis C
Virus infected cases than uninfected cases (13).
In reference to BIN1 and SARs-Cov2, the BIN allele
may interfere with the replication of the SARs-Cov2 virus in 2
ways. By binding to the SARs-Cov2 nucleocapsid phosphoprotein, BIN1
could impede viral infection via the following: i) By conjugating
to SARs-Cov2 NSP1, BIN1 could interfere with viral infection
(14); and ii) the SARs-Cov2 NSP3
folds into a tunnel, which then plugs itself into cell membranes,
allowing newly produced viral RNA to exit (15). By conjugating to NSP3, BIN1 could
interfere with this process.
The findings of the present study demonstrate that
risks for COVID-19 mortality are not simply related to an advanced
chronological age or the comorbidities commonly observed in aging,
such as CHD and AD, but to AD genetics. Further investigation of
the possible SARs-Cov2 antiviral properties of BIN1 may lead to the
development of novel treatments for COVID-19.
Acknowledgements
The present study was supported in part through the
computational resources and staff expertise provided by Scientific
Computing at the Icahn School of Medicine at Mount Sinai.
Funding
The research reported in the present study was
supported by the Office of Research Infrastructure of the National
Institutes of Health under award numbers S10OD018522 and
S10OD026880. The content is solely the responsibility of the
authors and does not necessarily represent the official views of
the National Institutes of Health.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from UK
Biobank.
Authors' contributions
SL and PHR contributed equally to the conception,
writing, and data analysis of the present study.
Ethics approval and consent to
participate
The UK Biobank application for the present study was
approved as UKB project 57245 (SL and PHR).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Docherty AB, Harrison EM, Green CA,
Hardwick HE, Pius R, Norman L, Holden KA, Read JM, Dondelinger F,
Carson G, et al: Features of 20 133 UK patients in hospital with
covid-19 using the ISARIC WHO Clinical Characterisation Protocol:
Prospective observational cohort study. BMJ.
369(m1985)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Perry G: Alzheimer's disease patients in
the crosshairs of COVID-19. J Alzheimers Dis. 76(1)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kuo CL, Pilling LC, Atkins JL, Masoli JAH,
Delgado J, Kuchel GA and Melzer D: APOE e4 genotype predicts severe
COVID-19 in the UK Biobank community cohort. J Gerontol A Biol Sci
Med Sci. 75:2231–2232. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Franzmeier N, Rubinski A, Neitzel J and
Ewers M: Alzheimer's Disease Neuroimaging Initiative (ADNI). The
BIN1 rs744373 SNP is associated with increased tau-PET levels and
impaired memory. Nat Commun. 10(1766)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Atkins JL, Masoli JA, Delgado J, Pilling
LC, Kuo CL, Kuchel GA and Melzer D: Preexisting comorbidities
predicting COVID-19 and mortality in the UK Biobank community
cohort. J Gerontol A Biol Sci Med Sci. 75:2224–2230.
2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Chang CC, Chow CC, Tellier LC, Vattikuti
S, Purcell SM and Lee JJ: Second-generation PLINK: Rising to the
challenge of larger and richer datasets. Gigascience.
4(7)2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhu A, Salminen LE, Thompson PM and
Jahanshad N: The UK Biobank Data Parser: A tool with built in and
customizable filters for brain studies. Organization for Human
Brain Mapping Rome, Italy, June 9-13, 2019, 2019.
|
|
8
|
Zinzula L, Basquin J, Bohn S, Beck F,
Klumpe S, Pfeifer G, Nagy I, Bracher A, Hartl FU and Baumeister W:
High-resolution structure and biophysical characterization of the
nucleocapsid phosphoprotein dimerization domain from the Covid-19
severe acute respiratory syndrome coronavirus 2. Biochem Biophys
Res Commun: Oct 3, 2020 (Epub ahead of print). doi:
10.1016/j.bbrc.2020.09.131.
|
|
9
|
Frick DN, Virdi RS, Vuksanovic N, Dahal N
and Silvaggi NR: Molecular basis for ADP-Ribose binding to the Mac1
domain of SARS-CoV-2 nsp3. Biochemistry. 59:2608–2615.
2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Casal E, Federici L, Zhang W,
Fernandez-Recio J, Priego EM, Miguel RN, DuHadaway JB, Prendergast
GC, Luisi BF and Laue ED: The crystal structure of the BAR domain
from human Bin1/amphiphysin II and its implications for molecular
recognition. Biochemistry. 45:12917–12928. 2006.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang J, Jia Y, Zhao S, Zhang X, Wang X,
Han X, Wang Y, Ma M, Shi J and Liu L: BIN1 reverses PD-L1-mediated
immune escape by inactivating the c-MYC and EGFR/MAPK signaling
pathways in non-small cell lung cancer. Oncogene. 36:6235–6243.
2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Abramovs N, Brass A and Tassabehji M:
Hardy-Weinberg equilibrium in the large scale genomic sequencing
era. Front Genet. 11(210)2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hraber P, Kuiken C and Yusim K: Evidence
for human leukocyte antigen heterozygote advantage against
hepatitis C virus infection. Hepatology. 46:1713–1721.
2007.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lapointe CP, Grosely R, Johnson AG, Wang
J, Fernández IS and Puglisi JD: Dynamic competition between
SARS-CoV-2 NSP1 and mRNA on the human ribosome inhibits translation
initiation. bioRxiv: 2020.2008.2020.259770, 2020.
|
|
15
|
Wolff G, Limpens R, Zevenhoven-Dobbe JC,
Laugks U, Zheng S, de Jong AWM, Koning RI, Agard DA, Grünewald K,
Koster AJ, Snijder EJ and Bárcena M: A molecular pore spans the
double membrane of the coronavirus replication organelle. Science.
369:1395–1398. 2020.PubMed/NCBI View Article : Google Scholar
|