1. Introduction
Changes in diet composition in aquaculture have
brought about concerns regarding some negative effects at the
gastrointestinal levels. The majority of cultured fish species
encounter issues when a fish meal is replaced with a plant
protein-based source in a significant proportion of their diet. The
proliferative and inflammatory responses in the intestines of
various (functionally) monogastric animals, which include the
Atlantic salmon Salmo salar (1), common carp Cyprinus carpio
(2), and rainbow trout
Oncorhynchus mykiss (3) are
among the several concerns which have arisen.
The importance of dietary acid supplementation to
fish and feed production supports expanding fish production for
sustainable feed production. Products commonly contain plant
by-products, oilseeds, legumes, pulses, lupins and cereal to
replace fishmeal, including an exogenous enzyme applied to enhance
the utilization of plant nutrients in aquaculture diets (4).
The concentration of hydrochloric acid in the
stomach decreases with meal consumption, increasing pH levels.
However, this increase in pH levels exerts a detrimental effect on
the activation of pepsin and pancreatic enzyme secretions,
decreasing the digestive capacity and affecting growth performance.
As a result, acidifiers, such as organic acids and their salts
provide a viable option. As a result, these have attracted
increasing attention as a potential antibiotic substitute for
improving fish development and health. Furthermore, organic acids
are involved in several energy-generating metabolic pathways
(5).
The present review article focuses on the effects of
acidifiers on growth performance and feed efficiency in different
mechanisms, such as acid source in the gastrointestinal tract (GI
tract), its effect on the metabolism and the physiological strategy
of acidification, and specifically on the effects of dietary pH
levels.
2. Physiology of fish
Digestive morphology
Fish rely on a broad array of food sources in
nature. Carnivorous fish that eat meat and other more digestible
feeds vary in their physical and behavioral functions from
herbivorous fish that eat fibrous meals, such as phytoplankton and
macrophytes. Carnivorous fish have a short and simple stomach with
a thick mucosa for absorption. Herbivorous fish have an additional
masticatory apparatus or other physiological adaptation to aid in
the breakdown of plant cell walls before digestion begins and a
long, thin stomach to extend gut retention time and improve
digestion and absorption.
Plant elements in diets can expose fish to a cascade
of anti-nutritional agents, culminating in pathological disorders
later in life (6). Soybean meal
has exhibited substantial negative alterations in the intestinal
architecture of Nile tilapia, such as the expansion of the
submucosa (SM) and lamina propia (LP), and an increase in the
number of goblet cells, as compared to other studied ingredients.
Soybean meal has been reported to include anti-nutritional
chemicals that lead to digestive issues in Atlantic salmon
(Salmo salar) and summer flounder (Paralichthys
dentatus) (7). The observed
changes in the intestinal morphology of the Nile tilapia were less
severe than those in salmonids and were mostly located in the
proximal region. In addition, the mucosa of the proximal part of
the intestinal tract is longer in the tilapia. It also possesses
more branched villi than the middle and distal regions in the
tilapia (8), suggesting more prone
to intestinal disorders.
The digestive tract of Oreochromis niloticus
(O. niloticus; Nile tilapia) is characterized by a sequence
of loops arranged in a consistent and detailed pattern that is both
unique among species and one of the most complex patterns ever
observed in fish. The intestine leaves the stomach and enters the
spiral portion of the intestine, following the elongated margins of
the liver. The spiral intestine consists of two primary coils
(proximal and distal) with a centripetal and centrifugal loop in
each. Between two large coils, a short gastric loop is inserted.
Finally, the terminal segment of the intestine leaves the spiral
region and follows a straight path to the anus (9). The possession of an intestine with a
length far surpassing that of the body cavity, as well as the
ability to arrange the elongated gut into loops or coils of some
type, as O. niloticus, in general, are found in adult
herbivorous fish. With a total intestinal length of 0.8 to 15 times
the body length, the Nile tilapia is on the shorter end of the
range of herbivorous fish (8).
This trait may be due to the adaptable fish diet, which can easily
be changed from its standard diet.
The digestive mechanisms of the Nile tilapia differ
from those of carnivores and herbivores. It can eat a wide variety
of foods. A series of loops and coils comprise the macroscopic
morphology of the digestive tract. The hepatic loop, proximal major
coil, gastric loop, distal major coil, distal major coil and
terminal segment are all found in the caudal stomach (9). The proximal gut has received
increasing attention in morphological and functional responses to
fasting and famine; no variations have been found between Tilapia
intestinal sections and those of higher vertebrates (10,11).
The Nile tilapia has a well-developed GI tract linked to its
feeding behaviors and food preferences. The distal intestines of
the Nile tilapia are known to have active microbial fermentation
and short-chain fatty acid absorption (12,13).
When the tilapia is fed a low-protein diet, intestinal bacteria
release a greater amount of necessary amino acids (14).
The gastrointestinal system (GI tract) is the site
of food digestion and nutritional absorption in fish and the first
line of defense against hazardous chemicals (1,15).
Different feed components have been shown to alter intestinal
morphology (2,16). Fish are exposed to several foreign
components, such as carbohydrates and anti-nutritional factors when
plant-based foods substitute animal protein sources in their diet.
These foreign components can interfere with the normal processes
occurring in the gut (17). Some
researchers have replaced fishmeal with plant protein-based diets
for the tilapia (18-20).
The intestinal morphology of the Nile tilapia is negatively
affected by soybean meal when paired with an environmental
influence, according to Tran-Ngoc et al (21). However, there is still a scarcity
of data available on the mechanisms through which other plant-based
substances alter the intestinal shape.
Histology of the GI tract
The GI tracts of cultivated omnivore fish, such as
the Nile tilapia are well-developed due to their feeding patterns
and food types. The esophagus, Y-shaped stomach and lengthy
intestine comprise the GI tract of the Nile tilapia. The GI tract
wall has a variety of cell types that are related to the anatomy
and physiology of each fish. The mucosa, SM, muscularis propria and
serosa are the four layers of the gut wall that comprise the basic
histological anatomy of the GI tract in vertebrates (22). These were examined using optical
and electron microscopes based on histological structures (23).
GI tract epithelial cells, goblet cells and certain
gland cells comprise the mucous cells in the fish GI tract
(24,25). Mucous cells in the stomach
epithelium appear as compacted columnar mucous cells (26). The mucosal layer has piqued the
interest of several researchers as it is a mucous membrane
containing mucous cells that are crucial for lubrication,
absorption and the transportation of macromolecules, increasing
digestive capacity, and preventing acidity and bacteria at the
epithelial level (27,28). Mucous-secreting cells, termed
goblet cells, are compacted in the esophageal epithelium and are
scattered in the intestinal epithelium. The mucous in the GI tract
comprises various mucosubstances or mucins, such as neutral mucin
and acid mucin; however, the mucins in the GI tracts of fish vary
depending on the species, age and location (29). Neutral mucin is abundant in all
three regions of the intestines of the Nile tilapia, crucial for
enzymatic digestion and absorption. The Nile tilapia has little
acid mucin in the early section of the gut and no sulfated acid
mucin. As a result, acid mucin does not appear well in the
intestine of the tilapia, which excretes soft meal remnants.
According to histochemical research, the esophagus mucus of the
Nile tilapia contains neutral and acid mucin. The epithelium mucus
of the stomach of the tilapia on the other hand, has a large
quantity of neutral mucin and low content of acid mucin (26). Acid mucin has been detected in the
produced mucus from the gastric glands, found in the stomach
epithelium of herbivorous fish, such as the tilapia. Using acidic
lysis, plant cell walls have been successfully disrupted and
triturated (30). This
histochemical analysis of the Nile tilapia is critical to the
understanding of the GI tract for the formulation of sustainable
feed. Concerning this literature, the addition of organic acids or
decreasing the dietary pH of the diet have significantly improved
the digestibility and intestinal morphology of the tilapia. Hence,
histological analysis is crucial for determining which cells are
increased. In the study by Huan et al (31), the cross-section of the intestinal
mucosa morphology of the tilapia was measured, such as villus
height and width, to determine the growth and digestibility based
on the intestinal structure. Some of the following terms are
important for determining the different parts of histological
cells: i) Enterocytes are simple columnar epithelial cells that
line the inner surface of small and large intestines; ii)
eosinophilic granulocytes are inflammatory cells that occasionally
migrate into the LP; goblet cells are unicellular intraepithelial
mucin-secreting glands scattered within the simple epithelium, such
as cuboidal, columnar and pseudostratified cells; goblet cells are
mucus-secreting, and they are compacted in the esophageal
epithelium and dispersed in the intestinal epithelium (26); iii) the LP is the thin and delicate
core of connective tissue in simple folds; iv) the lumen is the
inner space of a tubular structure, such as an artery or intestine;
v) microvilli are found on the top of villi; vi) the SM is a thin
layer of connective tissue between the base of folds and the
stratum compactum; vii) villi (height/width) are small, finger-like
structures in the small intestine; viii) intestine.
The intestinal functional physiology of fish is
influenced by various factors and differs by species. The function
of the fish intestine is critical for ensuring cost-effective
production and low waste output (32). The presence of potentially toxic
components in food, such as anti-nutritional substances (33) and oxidized components (34), as well as production techniques,
such as feeding regimes (35) and
diet composition (36), can alter
digestive functions. As a result, it is critical to continually
assess the effects of acid on digestive physiology to guarantee
that the raw material is safe and effective.
Tilapia fed supplemented organic acids, such as
potassium diformate and calcium butyrate diets exhibit more
pronounced improvements in intestinal morphology under hypoxic
conditions than normal conditions. In the distal intestine, the
tilapia have a thinner SM and LP, and fewer goblet cells, which is
considered an improvement in the intestinal epithelium (37). The capacity of organic acid to
strengthen intestinal morphology is strongly dependent on the
conditions of upbringing (38).
Butyric acid and butyrate also affect cellular functions that are
crucial for intestinal health, such as reducing mucosal
inflammation and oxidative stress and increasing the barrier
function of the intestinal epithelia (39). Protease and organic acid salts in
combination improve nutrient digestion and intestinal architecture
(31). Different proximo-distal
gradients of distinct digestive enzymes may differ within the same
fish species. Lipid membrane hydrolysis, for example, occurs
primarily in the proximal regions of the intestine. The hydrolysis
of carbohydrates and protein components, on the other hand, appears
in the medial and distal regions of the intestine (40).
While gastric glands are found in the front section
of the fish stomach (41), the
stomach of the Nile tilapia contains three regions (cardiac, fundic
and pyloric); gastric glands are found in the cardiac and fundic
regions. The fish stomach can be split into two main parts
histologically: The anterior cardiac and fundic region with gastric
glands and the posterior pyloric region without gastric glands
(42). In addition, tubular and
acinar mucous glands have been discovered in the posterior region
of the stomach of the tilapia (26).
Digestive enzyme regulation
Based on the weight gain results from a previous
study, it appears that acidifiers, particularly citric acid, at a
dose of 1.5% of the meal, increase the activity of digestive
enzymes in the red drum. When organic acids were added to the diet,
pepsin activity, pancreatic enzyme activities (trypsin, lipase, and
amylase) and intestinal enzymes increased (43).
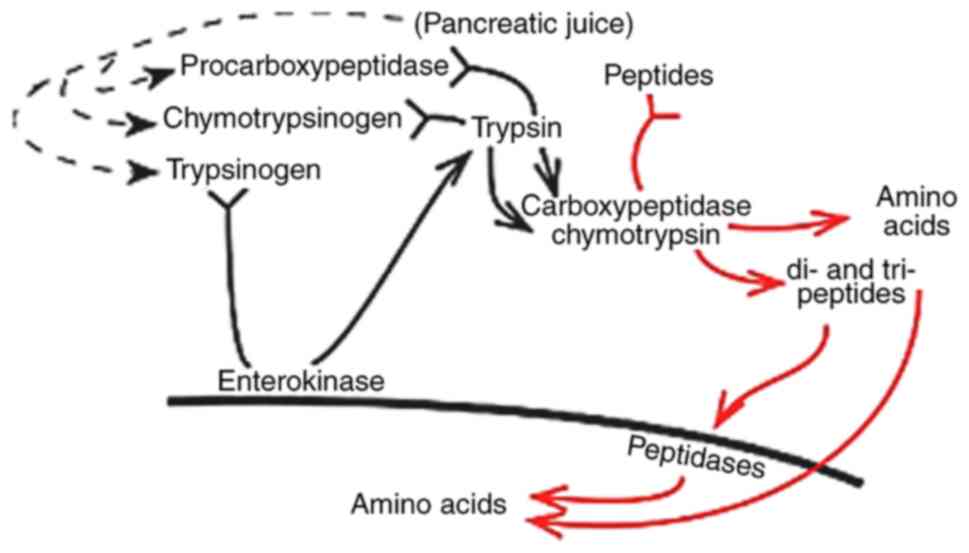
i) Stomach enzymes. Hydrochloric acid
concentrations in the stomach are decreased during periods of
high-feed consumption when the animals are young or the meals are
high in protein, for example. This reduction negatively affects
pepsin activation and pancreatic enzyme secretion and impairs
digestion. Therefore, acidifiers have been added to the feed to
address this issue and aid feed digestion. In addition, organic
acids have been found to aid in the hydrolysis of proteins
(44).
The mechanism of dietary acid to feed is to lower
the pH level of the feed for the increase in pepsin levels. Pepsin
is an important acidic aspartic protease widely applied in protein
hydrolysis. It is found in fish viscera, primarily in the gastric
juice of the stomach lumen (45-48),
constituting 5% of fish weight (49). The peptide linkages are easily
broken, allowing proteins to degrade in acidic environments
(50). It is termed pepsinogen as
it is generated and released in an inactive condition in the
stomach membrane (SM). This SM is stable in neutral and weak
alkaline settings and contains 44 amino acids. When exposed to the
hydrochloric acid (HCl) present in gastric juice (pH 1.5-2.0), the
44 amino acids are proteolytically eliminated in an autocatalytic
method, resulting in the activation of pepsin (51), the pepsin activity reported in
Sparus aurata has only been found in the stomach (52).
The main product of peptic cells is pepsinogen, and
it is found in this form in the gastric mucosa (53), blood, urine and other body fluids
(54,55). Pepsinogen can only be measured
after its irreversible conversion to the active enzyme, pepsin, by
an autocatalytic process at pH <6.046, following its
secretion into the stomach. Pepsinogen rapidly converted to pepsin
at pH 2.0, but extremely slowly at pH 5.0 to 6.0. Furthermore,
pepsin functions optimally in an acidic environment with a pH of
≤2.0, whereas it functions at a slow rate at pH 5.0 to 6.0.
Furthermore, pepsin functions optimally in an acidic environment
(pH 2.0 to 3.5) and rapidly degrades above this pH (56). As a result, the optimal pH (the pH
value that provides the highest enzymatic activity) and pH
stability (the pH range that provides adequate enzyme stability)
substantially affect fish pepsin activity. Pepsin activity
diminishes when the pH decreases below optimal levels (57). In the study by Castillo et
al (43), the pepsin activity
was shown to be greater in the homogenized stomachs of juvenile red
drum from 1.5% citric acid, 0.75 potassium diformate and 1.5%
potassium diformate treatment at 2 h after feeding. However, only
treatment with 0.75% potassium diformate treatment led to
statistically significant results. Since the optimum pH
significantly affects the activity of fish pepsin (57), it would be of interest to determine
whether the pH of the stomach contents is connected to protein
digestibility.
Exocytosis releases pepsinogen as a proenzyme
(zymogen) from main cells. The acidic stomach juice reversibly
activates some pepsinogen molecules. These activated pepsinogen
molecules permanently activate themselves and the remaining
inactive pepsinogen molecules to create Pepsin by intramolecular
and intermolecular cleavage. Catalysis exposes the catalytic domain
by removing an autoinhibitory region (activation peptide) (Fig. 1).
ii) Intestinal enzymes. Enzyme activities in
the digestive tract of gilthead sea bream were investigated by
Deguara et al (52). The
results of their study revealed that pepsin activity was found
exclusively in the stomach. By contrast, other enzymes such as
trypsin, chymotrypsin, carboxypeptidase A, carboxypeptidase B and
amylase were found in all gut regions, including the stomach.
Pepsin activity has been found to be absent in all other sections
of the digestive system save the stomach in white sturgeon and
striped snakehead (58). The
activity of other enzymes, such as trypsin and amylase, was much
lower in the stomach (52).
Trypsin, chymotrypsin, carboxypeptidases A and B, and amylase have
all been found in the stomach of fish (59-61).
Chakrabarti et al (58)
hypothesized that fish intestines are still at an evolutionary
stage when most areas can generate all of the major enzymes before
the emergence of site-specific enzyme synthesis found in higher
invertebrates.
Inorganic acid inclusion, trypsin activity, lipase
and amylase levels have been shown to be higher in juvenile red
drum fed acidified diets, such as citric acid and potassium
diformate. Increasing secretin levels leads to a lower pH, possibly
stimulating pancreatic secretions (43). In addition, digestive enzymes, such
as leucine-aminopeptidases and phosphatases increase the activity
of the intestine. As acid and alkaline phosphatases are involved in
the hydrolysis of phosphorus, the stimulation of digestive enzymes
by organic acids may be one of the reasons for enhanced mineral
digestion by fish. However, organic acids may indirectly affect the
activity of the intestine digesting enzymes.
Haemato-immunological responses
Malic acid (5 g/kg) added to the Nile tilapia diet
has been shown to increase the hematological values (62). On the other hand, a positive result
on organic acid basal meal enriched with commercial formic and
propionic acids at a 1 or 2 g/kg feed dosage was fed to Nile
tilapia fingerlings. This improved growth, hemogram parameters,
white blood cell counts, lymphocyte and neutrophil counts, and the
body chemical composition, and also reduced microbial contamination
in intestine in a dose-dependent manner in the Nile tilapia
fingerlings (63). The amount of
uric acid in the blood has also been shown to be decreased when an
organic acid is added to the diet (64), which results in the improved
utilization of amino acid digestibility and proteins due to the
protein metabolism in the urea as the major end product.
Gut microbiota and blood
parameter
With the supplementation of acid to fish, the
microbial balance and proper pH in the digestive tract eliminate
pathogenic microorganisms and maintain fish health satisfactorily.
Therefore, the use of organic acids is a preventive alternative in
maintaining the health of cultured fish. Its actions in the GI
tract inhibit the growth of pathogenic bacteria, mainly
Gram-negative bacteria, aid in the digestion and absorption of
nutrients, as well as exert beneficial effects on animal
performance (65). However, some
organic acids, particularly citric, metacetonic and acetic acids,
are added to pellets for storage effects or the improvement of feed
utilization, or both (22). Other
studies have not found a growth-promoting effect of organic acids
(66,67). This is dependent on the fish
species, type of organic acid and dosage used (5). As a result, it is critical to
recognize that the potential benefits of organic acids may vary
depending on the species and dose used.
Acidifiers improve feed performance; they decrease
the uptake of pathogenic organisms and toxic metabolites. In the
intestinal tract, the acidifiers reduce the pH level in the
stomach, particularly in the small intestine, through the delivery
of H+ ions. On the other hand, they inhibit the growth
of gram-negative bacteria by dissociating the acids and producing
anions inside bacterial cells (68).
At a pH <5, the growth of several Gram-negative
bacteria is reduced. A low pH also creates a natural barrier
against germs rising from the ileum and large intestine. Low
molecular weight acids are lipophilic and penetrate past the cell
membrane of Gram-negative bacteria. Organic acid supplementation
has been shown in several trials to reduce pathogenic bacterial
counts in the digestive tract, while increasing the amount of
acid-tolerant, ‘good’ bacteria, such as Lactobacilli (5).
Another effect of the supplementation of organic
acids is antimicrobial activity. Owen et al (69) demonstrated that the proportion of
Gram-positive bacteria in Clarias gariepinus treated with
sodium butyrate tended to increase. Using organic acids/salts in
aquafeeds for commercial purposes can improve growth and disease
management and exert antimicrobial effects as it releases protons
into the cytoplasm and penetrates the cell wall of Gram-negative
bacteria (68). The bacteria
require a large amount of adenosine triphosphate to maintain a
balanced intracellular pH, resulting in cellular energy depletion
with eventual cell death (65).
Lactic acid bacteria is one of the most common probiotic bacteria
used in aquafeed (70). Lowering
the gut pH with dietary potassium diformate exerts a eubiotic
effect on the allochthonous, beneficial lactic acid bacteria.
Lactic acid bacteria can grow at a relatively low pH, exhibiting
further resistance to organic acids and salts than Gram-negative
bacteria. These indigenous probiotic bacteria can colonize the
intestinal surface and form a barrier, serving as the first defense
to limit the direct attachment or interaction of pathogenic fish
bacteria to the gut mucosa (70).
Organic acids reduce the pH level of the gastrointestinal content
in the red hybrid tilapia. They could lead to an enhanced
reproduction of beneficial bacteria, the inhibition of pathogenic
microorganisms and subsequently, in an improved micro-ecosystem
(71,72).
The improvement of protein digestibility and
nitrogen retention is contributed to the lower gastric pH
associated with a higher pepsin activity (73). For example, in red hybrid tilapia,
dietary potassium diformate at 2 g/kg was shown to reduce the diet
and digesta pH of the fish stomach and gut; it also markedly
decreased the total bacterial counts in the feces (67).
As low molecular weight lipophilic organic acids can
infiltrate through the cell membrane of Gram-negative bacteria, the
acidification of their metabolisms can cause bacterial cell death.
In the hybrid tilapia, dietary potassium diformate promotes the
colonization of certain gut bacteria, while inhibiting the growth
of others (74). Another
explanation for improved growth performance could be the spread of
indigenous probionts. These Gram-positive bacteria aid in
fermenting some non-digestible carbohydrates, hence increasing
nutritional availability (70).
The acid anions of the dissociated organic acids are counteracted
by the high intracellular potassium concentrations in Gram-positive
bacteria (75). The organic acid
can acidify the cytoplasm of Gram-negative bacteria, resulting in
eventual cell death. Potassium diformate can modify microbial
communities in tilapia guts, accounting for its ability to initiate
an immune response. The continuous vitalization provided by the
endemic intestinal flora affects the quality and quantity of immune
cells in the gut mucosa (76).
Elala and Ragaa (77) examined the
eubiotic effect of a dietary acidifier (potassium diformate) on the
health status of cultured O. niloticus. Tilapia were fed
graded levels of potassium diformate (control diet, 0%; 0.1, 0.2
and 0.3%) for 60 days at an initial body weight of 6.15 g. The
addition of potassium diformate to the diet improved growth and
apparent protein digestibility. It also exerted a eubiotic effect
on the proliferation of indigenous bacteria, which is crucial for
stimulating the immunological response to diseases.
Nutrients and mineral absorption
Acidifiers improve feed performance by enhancing
nutrient absorption, the proliferation of cells in the mucosal
epithelium of the intestine, preventing aquatic pollution and
reducing phosphorus discharge in water (78). For example, a 0.922 g/kg organic
acid blend (calcium propionate, calcium formate and sodium acetate)
supplemented protease in meat and bone meal diet has been shown to
improve nutrient digestibility and nutrient retention in the Nile
tilapia (31). In addition,
supplementing diets with organic acid and their salts has been
found to exert beneficial effects on mineral absorption (79) and nutrient digestibility (80).
Organic acid supplementation decreases duodenal pH
levels, improves nitrogen retention and improves nutritional
digestibility (81). In
aquaculture, dietary acidification using organic acids or their
salts in aquatic animals has been used (82). Furthermore, organic acids or their
salts inclusions in the diet of aquatic animals have been observed
to increase the nutritional value of the diets of aquatic animals
and their growth (82). Several
studies have been performed on various aquatic species, including
carnivorous species, such as rainbow trout, Atlantic salmon and
Arctic charr, herbivorous filter feeders, such as carp and tilapia,
and omnivorous species, such as catfish and shrimp (5,83-89).
In addition, organic acids improve phosphorus absorption (90). Thus, organic acids, salts, or
mixtures are promising feed additives for aquatic animals to
enhance some fish species' growth performance and feed utilization.
Furthermore, they inhibit bacteria and contribute to nutrition,
which is involved in several metabolic pathways for generating
energy and improving major nutrient digestibility (82).
The addition of 0.2% formic and propionic acid/salt
to the diet of the Nile tilapia has been found to increase the
amount of retained protein and fat. In addition, supplementation
with acidifiers improves nutrient digestibility (91). A high dose of formic and propionic
acid/salt is required to improve the immune status in tilapia feed.
Fish extracts and other aquatic organisms are rich in organic acids
(92). Organic acid compounds
improve weight gain and the food conversion ratio by improving dry
matter, crude protein digestibility and mineral absorption
(67,73,93).
RNA upregulation
In the study by Busti et al (94), the effects of dietary organic acids
on the immune response of European sea bass resulted in the
upregulation of target genes, such as IL-8, IL-10 and TGF-β. Thus,
the organic acids exhibit prebiotic properties in the gut
microbiota, promoting beneficial bacteria taxa, such as
Lactobacillus, Leuconostoc and Bacillus.
Furthermore, these acids appear to induce a potential functional
reconfiguration of the gut microbiota, enabling a significant
decrease in several inflammation-promoting and homeostatic
functions. Again, for the first time in the seabass study, the
exposure to suboptimal rearing conditions was shown to modify the
gut microbiota structure, reducing lactic acid bacteria and
increasing proteobacteria. These findings were consistent with the
inflammatory process observed at the mRNA level (94).
The feeding frequency in the two feeding trials in
the juvenile red drum (Sciaenops occelatus) revealed a
balance in the timeframe of feeding, such as 08:00, 12:00, 16:00,
to digest the eaten feeds from the stomach and intestine in an
appropriate, timely manner. It was reiterated in the illumination
pattern, and feeding time is involved at a different level in
regulating the secretion of digestive juices. There is a
significant change in stomach pH levels after feeding (95). The addition of acid to lower the
dietary pH in the red hybrid Tilapia and rainbow Trout
(Oncorhynchus mykiss) studies may increase the pepsin
activation and mineral absorption (67,96).
It may explain why growth and feed efficiency have improved. The
concentration of hydrochloric acid in the stomach is reduced during
periods of high-feed intake, and the pH level increases. The
increase in pH levels negatively affects pepsin activation,
potentially attenuating protein digestion in the stomach (5,57).
This may be the reason why the pH 8.00 diet-initiated mortality
indicates that growth is declining in Nile tilapia fry. The dietary
pH adjustment is a major solution that best fits the required
dosage level to the Nile tilapia dietary acidification. The stomach
converts pepsinogen to pepsin rapidly at pH 2.0 to 4.0; however,
this conversion is attenuated at pH 5.0 to 6.0 and declines rapidly
above this pH (56); this
coincides with the red hybrid Tilapia and rainbow Trout
(Oncorhynchus mykiss) (67,96).
In the case of pigs, the optimum pH value has the highest enzymatic
activity, significantly affecting pepsin activity. Pepsin activity
decreases when the pH levels fall below the ideal level (97). Thus, perhaps the pH 4.6 diet
significantly affects pepsin activity.
3. Dietary acidification
Acidifiers present a promising alternative for
improving the performance and health of livestock. The effects of
dietary acidification and acid sources on the growth performance of
fish and feed efficiency are presented in Table I. Acidifiers can also improve feed
performance; they can improve growth, feed utilization and disease
resistance in fish (5), improve
the shelf life of pellets (98),
and improvement in pig growth performance as a result of increased
food consumption due to increased diet palatability, a more
efficient conversion of the food to live weight due to a reduction
in bacterial competition for nutrients, or increased enzymic
activity due to a reduction in gastric pH due to the acidification
of the diet (99). These organic
acids, such as acetic, butyric, citric and malic acids, and their
salts are found in plant and animal tissue. They can be used as
acidifiers in the livestock feed industry, including aquaculture.
When added in sufficient amounts, acidifiers and their salts can
stimulate growth and feed efficiency and enhance feed quality. The
acids in the aquaculture diet should be standardized to respond to
gastric and agastric species (78).
 | Table IEffects of dietary acidification and
acid sources on different fish growth performances and feed
efficiency. |
Table I
Effects of dietary acidification and
acid sources on different fish growth performances and feed
efficiency.
| Fish species | Acid/acid salt
sources | Dose (%) pH | Parameters
increased in SGR (%), FCR, and WG (g) | (Refs.) |
|---|
| Nile tilapia
(Oreochromis niloticus) | Hydrochloric
acid | pH 4.60 | WG: 2.6; FCR: 1.3;
SGR: 4.9 | (86) |
| Common carp
(Labeo rohita) | Citric acid | 3.00% | FCR: 1.09;
SGR:1.58 | (87) |
| Red drum
(Sciaenops ocellatus) | | 1.50% | WG: 1547; FCR:
0.98 | (43) |
| Tilapia
(Oreochromis sp.) | | 2.00% | WG: 193.05;
FCR:1.82; SGR:2.10 | (105) |
| | | | WG: 1060.2 FCR:1.30
SGR: 2.35 | (67) |
| Nile tilapia
(Oreochromis niloticus) | | pH 4.60 | WG: 2.6; FCR: 1.5;
SGR: 4.7 | (86) |
| Beluga (Huso
huso) | | 3.00% | WG: 688.8 FCR: 1.08
SGR:3.75 | (90) |
| Rohu (Labeo
rohita) | | 3.00% | | (140) |
| Carp (Cyprinus
carpio) | | 3.00% | WG: 112.56 FCR:
1.17 SGR:1.20 | (141) |
| Sea bream Pagrus
major | | 1.00% | WG: 69.32 FCR: 1.00
SGR:2.20 | (146) |
| Nile tilapia
(Oreochromis niloticus) | Acetic acid | pH 4.60 | WG: 2.6; FCR: 1.5;
SGR: 4.7 | (86) |
| Nile tilapia
(Oreochromis niloticus) | Malic acid | 0.800 or 3.20% | WG:94.10 FCR:
1.48 | (139) |
| Nile tilapia
(Oreochromis niloticus) | | 5.00 or 10.0
g/kg | WG:48.5 FCR: 1.4
SGR: 2.40 | (108) |
| | Formic and
diformic | 1.00 or 2.00
g/kg | WG: 15.33 FCR: 5.20
SGR: 0.70 | (63) |
| Nile tilapia
(Oreochromis niloticus) | Calcium propionate,
calcium formate and sodium acetate | 0.922 g/kg | WG: 945.2; FCR:
1.20 | (31) |
| Red drum (Sparus
aurata) | Sodium
butyrate | 6.25 g/kg | WG: 9.83%; FCR:
1.18 SGR: 1.67 | (135) |
| Nile tilapia
(Oreochromis niloticus) | Potassium
diformate | 0.3% | WG:24.0 FCR: 1.45
SGR: 2.07 | (113) |
| Nile tilapia
(Oreochromis niloticus) | | 2 g/kg | WG: 1060.2; FCR:
1.30; SGR: 2.35 | (67) |
| | | 0.2 or 0.3% | 6.75% increased
apparent protein digestibility (APR) | (77) |
The use of acids to preserve fish and fish viscera
in the preparation of fish silage is a popular procedure with
widespread use in fish feed and recorded benefits (100,101). According to Batista (102), fish silage production was
initiated in the 1930s, initially with sulphuric and the
hydrochloric acid preservation of fish waste. The advantages of
acid-preserved products have drawn the attention of the scientific
community, leading to the investigation of the effects of these
short-chain acids on fish feed.
Inorganic acid
In all monogastric species, stomach acid is
involved, such as fishes are hydrochloric acid, a very strong
inorganic acid produced by gastric glands. This acid reduces the pH
of the stomach to a level of 2 to 3, depending on the species and
diet. Hydrochloric acid generation is minimal at birth/hatching,
but increases as animals mature. The pH of the stomach decreases as
more acid is formed. Therefore, pepsin, a proteolytic enzyme
required for protein digestion, must be activated at a low pH.
Pepsin activity is optimal at a pH of 2. Its effectiveness is
greatly limited at higher levels (103).
Organic acid
Organic acids are commonly used as an additive as
they function as chelating agents to bind cations along the gut
resulting in improved mineral absorption (104). In aquaculture species, organic
acids are common acidifiers. Organic acids have been found to exert
positive mineral absorption effects (90) and nutrient digestibility (80) by reducing the pH levels in the
digestive tract, specifically in the stomach and small intestine,
through H+ ion deposition (5).
Romano et al (105) (Table
I) demonstrated that when the citric acid level was increased
by 2%, there were leukocyte infiltrations and more excessive
necrosis and hemorrhaging that affected the growth of tilapia.
Citric acid and its salts, and formic acid and its salts are the
most extensively studied organic acids in aquaculture, according to
the review article by Ng and Koh (106). These short-chain organic acids
are ingested primarily by passive diffusion through the intestinal
epithelia, supplying energy for intestinal epithelia renewal and
gut health (93). In addition,
citric and lactic effectively enhance feeding behavior when applied
individually or with other extractive compounds (107).
There is an improvement in growth and feed
efficiency in the induced optimization of intestinal pH by malic
acid in the Nile tilapia (108).
However, no clear association has been found between the pH of the
diets and pepsin activity, implying that other factors may be
involved in the synthesis of the enzyme. A recent study determined
the optimum levels of acid supplementation at 6.25 g/kg (sodium
butyrate), providing growth performance immunity against
Aeromonas hydrophila (109). Furthermore, these lactic acid
bacteria have increased digestive enzyme activities in fish
(110).
Dietary pH levels
The resulting pH of the diet due to a combination of
ingredients in the formulation also affects gastrointestinal pH
levels with related consequences on protein digestion and gut
microbiota in monogastric vertebrates (111). A high proportion of fish meal
with a high acid-binding capacity in the diet ensures the nutrition
of fish. Compared to most alternative vegetable aquafeed
components, including sunflower, soybean, rapeseed and gluten, fish
meal has one of the greatest buffering capabilities (112). Decreasing the fish meal content
in modern aquaculture diets may affect the ideal gastrointestinal
pH for digestive enzyme action and gut bacterial community
(111). These observations also
suggest that fish nutritionists could manipulate the final dietary
pH to elicit positive growth performance, ultimately increasing
profitability in the aquaculture industry. A limited number of
studies have addressed the effects of dietary pH on the conditions
of aquatic animals, e.g., shrimp (83) and channel catfish (84).
The pH value of animal feed is crucial as it can
affect digestion following ingestion. For example, the increase in
pH levels in the stomach negatively affects the activation of
pepsin and possibly decreases the protein digestion capability in
fish (57). Therefore,
supplementing diets with organic acid and their salts exerts
beneficial effects on the growth performance of fish (113). On the other hand, it decreases
the digestive pH levels of the GI tract by accumulating
H+ ions, thus reducing the pH levels in the stomach and
stimulating the activation of pepsinogen to pepsin, increasing
protein digestibility and reducing gastric emptying rate. In
addition, the acids lower the pH of the stomach and foregut, which
stimulates pepsin activity and improves protein digestibility and
mineral absorption (114).
Furthermore, the acids further enhance protein digestion by
increasing the rate of proteolysis of large protein molecules
(115).
The buffering capacity of feed ingredients plays a
main role in reducing pH levels in the feed and stomach. Animal
protein (e.g., fishmeal) has a 15-fold higher buffering capacity
than cereals, which is why it is widely used in aquaculture diets.
Due to the low hydrochloric acid output of young animals, these
effects are of particular importance (116); the concentration of hydrochloric
acid in the stomach decreases during food consumption, increasing
pH levels. The activation of pepsin and pancreatic enzyme
secretions are inhibited by this increase in pH levels, reducing
the digestive capacity. Munilla-Morán and Saborido-Rey (117) determined that the pH optima for
pepsin activity in the stomach of Sparus aurata were 2.0.
Similar to all enzymes, the digestive enzymes markedly affect pH
levels. Distinct enzymes have different pH optimums when activity
is at its peak; activity decreases on either side of this optimum
rapidly and significantly. A pH adjustment can produce a 50%
reduction in inactivity in certain enzymes. As a result, it
significantly affects the rate and scope of digestion. Normal
digestive processes can explain the changes in pH levels in the
stomach of fish. The pH levels decrease as acids are secreted in
response to feed entering the stomach, followed by increases in pH
levels as acid secretion is terminated and digestion is evacuated.
The acidity decreases somewhat in the upper intestine before
increasing when bicarbonate ions are released into the gut lumen.
The effect of bicarbonate secretion becomes evident when going down
the length of the intestine. There is an ever-increasing trend in
the average pH from 6.8 in the upper intestine to 7.9 in the lower
intestine. The optimum pH in the different gut regions would affect
the digestibility of some dietary ingredients, leading to better
feeding and the performance of fish (117).
Decreasing the pH value in feed leads to a lower
buffer capacity. It thus promotes digestion in the animal since
less hydrochloric acid has to be produced in the stomach to
activate pepsin, and therefore, optimal protein digestion is
ensured (118). Furthermore, the
acid anion is complex with calcium (Ca), phosphorus (P), magnesium
(Mg), and zinc (Zn) results in the improved digestibility of these
minerals (119). On the other
hand, different acidifiers do not affect the contents of the
intestine 6 h following ingestion. Therefore, organic acids are
probably mostly metabolized by that time. Furthermore, pancreatic
secretions may act as a buffer against the effects of the acidifier
(43).
In another study by Yúfera et al (120), the pH levels in the stomach of
juvenile gilthead seabream were measured after they were fed via
three different strategies: Once, twice, or continuously. Feeding
was performed at the time points of 09:00, 09:00 and 17:00, or
continuously between 9:00 or 17:00 and 21:00. Under the three
feeding regimes tested, the gastric pH levels exhibited significant
daily rhythms. The constant supply feeding regimen ensured that the
stomach pH levels remained in the optimal pepsin range for a long
period of time. It may be one of the reasons for the significantly
greater weight of the fish following the feeding regimen. It may
also be explained by higher gastric activity in fish with a gastric
pH <4.5, as described by Márquez et al (57).
Yúfera et al (120) also investigated the association
between stomach pH levels and pepsin activity in juvenile marine
fish. Fish were fed either a single meal, twice, or the same diet
continuously at the same time. The stomach pH of fish fed only once
was around 4.5, and the highest pepsin activity was reported before
the feeding, with 30 pepsin activity units per fish. The fish fed
continuously had a stomach pH of 5.25 and a pepsin activity of
almost 280 units per fish in the late afternoon. This demonstrates
how low pH levels may affect pepsin activation. Ringø et al
(85) examined the effects of 1%
sodium lactate on the growth of Arctic charr fish and discovered a
significant increase. On the other hand, these results were not
observed in another study in Atlantic salmon using the same dosage
(66).
Fabay et al (86) (Table
I) focused on the contribution of dietary pH influencing the
gut pH levels, which presumably affects the capability and
efficiency to utilize dietary nutrients that convert to the flesh
(i.e., growth and feed efficiency). Therefore, the evaluation of
the growth rates and feed efficiency of the Nile tilapia as regards
dietary pH in that study was an indirect comparison of the general
physiological conditions in which the total digestive enzyme
activities operate on their corresponding substrates, stomach and
intestine. The inclusion of hydrochloric acid in the diet was more
effective in the tilapia fry with a pH range of 4.6.
4. Acid sources of fish
The condition that possibly affects the growth
performance and feed utilization may be the gastric or
gastrointestinal pH mechanisms. The gastrointestinal luminal
digestive disorders are very important in optimizing the
utilization of modern aquafeeds. Gut pH seems to be influenced by
the pH of the diet (121). The pH
of the stomach and intestine creates a medium in which the adequate
digestion of dietary proteins and lipids is affected by an optimal
environment for the activation and activity of the digestive
enzymes. A suitable pH level in the intestine may also be the
perfect habitat for some gut flora and fauna to thrive, while
others do not survive.
Stomach acid and intestinal acid
The majority of fish have a low acid secretion in
their lumen compared with mammals. The inclusion of dietary
acidifiers reduces the pH levels in the GI tract, increases phytic
acid breakdown and eliminates GI pathogenic microorganisms. It also
decreases the emptying time of the GI tract, improves nitrogen
retention, increases nutrient digestibility, and improves mineral
absorption and transportation (81,122). The stomach acid secretion of
vertebrates, such as fish exhibits continuous acid secretion and
low gastric pH levels during fasting. The presence acid is
maintained neutral gastric pH during fasting hydrochloric acid
released only after a meal's ingestion. Fish such as tilapia and
catfish can digest feeds due to the presence of a stomach and
intestine. It is important to acknowledge that dietary acids may
provide beneficial effects. The optimal pH level in the gut has a
significant effect on the activity of fish pepsin (57).
The main reason for the addition of acid to the diet
is to lower the pH level of feeds, which may benefit the digestive
functions of the Nile tilapia towards an increased growth rate and
better feed efficiency. The H+ ion from the acid
stimulates the activation of pepsinogen to pepsin in the stomach,
thus improving protein digestibility (94). In addition, during feed intake, the
hydrochloric acid concentration in the gut is reduced. In the study
by Moriarty (123), the stomach
pH level of tilapia was found to be ≤1.6, enabling it to digest a
high protein content (~49%), lysing in a high concentration of
amino acid as the feed remains for longer periods of time in the
stomach.
Similarly, in that study, the protein composition of
the diet was 47-49%, and the diet with a pH 4.6 led to maximal
growth rate and an optimal feed efficiency. The stomach does not
secrete acid when it is empty; thus, acid secretion begins upon
ingestion between pH 2.0 to 3.0(123). Therefore, if the acid requirement
of the tilapia was at pH 1.6, it required a pH 0.4-1.5 to convert
it into pH 2.0-3.0, which meant that approximately pH 1.0 was
sufficient to maintain the acid requirement of the tilapia. The pH
of the control diet was ~5.7, which requires pH 1.0 to convert it
into 4.6; the acid requirement is sufficient for in the stomach to
digest the high protein content. In another study on Indian carp
fed a diet supplemented with 3.0% citric acid, Baruah et al
(87) (Table I) reported a decrease in the pH of
feed from 5.87 to 4.85, with a subsequent reduction in the pH of
gut digesta from 6.62 to 5.65, increasing growth; their results
directly agree with dietary pH study (86).
Chyme pH and buffering capacity
The pH value influences the digestion and
availability of nutritional matters in the digestive tract of
animals in the gut (116). The
digestion of feeds creates a dynamic alteration in the resting pH
of each GI tract section which influences the dissolution and
precipitation of the dietary ions. In addition, the chemical
characteristics of the chyme are also altered during digestion, as
it passes along the GI tract with protein and carbohydrate
degradation occurring in the stomach and intestine that affect the
binding of ions to the solid phase (124).
Buffer capacity is defined as the HCl secretion in
ml or mmol required to lower the pH levels to pH 3.0 in the stomach
following feed intake. Different nutrients in animal feed increase
the buffer capacity of the feed, which is crucial for fish. A feed
with a high buffer capacity leads to a higher mortality than a feed
with a lower buffer capacity. Therefore, it is important to
consider the buffer capacity of the feed for the dosage rate. The
concentration of hydrochloric acid in the stomach is reduced during
periods of high feed intake, and the pH rises. The increase in pH
negatively affects pepsin activation, potentially reducing the
ability of the stomach to digest proteins (57). The stomach pH of fish given only
one feed decreased significantly to 2.6 after 8 h in a study by
Deguara et al (52);
however, when fish receive two feeds, the pH drops to 2.5
immediately after feeding and increases to 5.1 after 12 h.
Plant ingredients, such as extracted sunflower
(ESF), pea protein concentrate and soy protein concentrate exert
various and differing effects on the digestive physiology and
metabolism of the Atlantic salmon (1). This involves a decrease in
chyme-associated leucine aminopeptidases in fish fed plant
ingredients compared to fishmeal (88).
In addition, cellular sloughing has been observed in
at least some intestinal regions, which could be due to lower
enterocyte turnover and leucine aminopeptidase activity in rainbow
trout (Oncorhynchus mykiss) fed at least some plant
ingredients. When fed soybean protein, the fish have been found to
exhibit an increase in the height and thickness of their intestinal
villi (125).
ESF does not affect the brush border
membrane-leucine aminopeptidase activities, resulting in a
significant increase in the nitrogen concentration in all
intestinal compartments. Therefore, this suggests a reduced protein
digestibility compared to the fishmeal diets. In salmon and other
fish species, ESF has a relatively high digestibility (126). The high levels of nitrogen in
chyme in the ESF diet, on the other hand, suggest that the protein
digestibility of the ESF is low due to high inclusion levels (20%).
The high nitrogen levels indicate that the stomach secretion of
pepsin or other proteins is stimulated, as evidenced by the 30%
increase in nitrogen in the stomach, which is significantly higher
than that of salmon fed the other alternative feed ingredients.
Furthermore, the ESF significantly increases the dry matter content
of chyme in the mid-intestine, suggesting that other components
other than pancreatic enzymes and bile acids may be secreted or
present in the intestinal chyme, such as the higher number of
bacteria found in this diet group (1).
The optimum dietary pH in the study by Fabay et
al (86) (Table I) was slightly lower than that
observed another study in the stomach chyme of the Atlantic salmon
at pH 4.8(89) and that in rainbow
trout (Oncorhynchus mykiss) at pH 4.9(125). The effects of dietary pH on chyme
pH vary between fish species. Nikolopoulou et al (127) demonstrated that 2 h after
feeding, the sea bass (Dicentrarchus labrax L.) and
gilthead seabream (Sparus aurata) pH levels in the stomach
were 4.7 and 5.7, respectively. The former exhibited dietary
effects on stomach pH, while only the latter exhibited changes in
the chyme of the proximal and mid-intestine. The study by Fabay
et al (86) indicated
indirect evidence that there was an effect on chyme pH. This effect
was manifested in the growth and feed efficiency performance of the
Nile tilapia.
The adjustment of the pH level to 4.6 in diets may
have induced some enzymes. For example, carboxypeptidases A and B
levels in stomach peptic digestion cannot increase without acid in
the diet (123). Citric,
hydrochloric and acetic acid directly lead to a lower dietary pH.
This in turn presumably results in a lower gut pH level in fish.
This may also activate and enhance pepsin and other digestive
enzyme activities, and improve the solubility of minerals. The
stomach is an organ that requires high acid levels, while intestine
pH levels vary. In the case of dietary treatments with a higher
dietary pH, diets with 7.0 and 8.0 may have different digestive
physiology. They may result in the alkalinization of the chyme
entering from the stomach. Alkalinization is necessary for the
intestine to maintain intestinal epithelial integrity and
pancreatic and intestinal activity (128). This innate alkalinization may not
be sufficient to increase digestion efficiency; thus, this suggests
a lower growth performance than the diet with a pH 4.6(86).
Gastric acidification strategies
Two gastric acidification strategies have been
reported in vertebrates. First, species that maintain a permanent
acidic environment in the stomach are not affected by the presence
or absence of acids (e.g., mammals and birds). Second, species
maintain a neutral pH in the stomach lumen between meals and become
slightly acidic following a meal, e.g., sharks (129).
The first strategy has been observed in cobia
juveniles (121), rainbow trout
(Oncorhynchus mykiss) (124), southern bluefin tuna (Thunnus
maccoyii) (130), and in some
Elasmobranchii species (131,132). It appears that these species are
strictly carnivorous, and the observations were from studies that
involved a comparison of fed and fasted fish; by contrast, the Nile
tilapia is omnivorous and requires a daily feeding habit. In
gilthead seabream (Sparus aurata), erratic daily feeding by
changing the time of feed delivery at random each day may affect
the daily pattern from neutral/acid alternation to permanent
acidification (133). This
voracious species can always activate pepsinogen to begin the
hydrolysis of the ingested prey thanks to a constant acidic gastric
pH. Continuously feeding Nile tilapia acidic diets of various pH
resulted in irreversible lumen acidification (i.e., the first
technique of gastric acidification). Thus, the atmosphere in which
the production of digestive enzymes or the activation of
established digestive enzymes occur is related to increased
nutrient utilization.
The adjustment of dietary pH levels in feed may
reflect the acid requirement in the gastrointestinal lumen of the
tilapia. Studies on the effects of organic acids on the growth
performance of fish have yielded conflicting results. Some
researchers have not found growth-promoting effects of organic
acids (66), which vary from
species to species, type of organic acid and the dosage used
(5). Several studies have
evaluated the effects of dietary supplementation of specific
organic acid on the diet of tilapia (Table I). Still, the optimal dietary pH
requirement has not been directly investigated to date. Some
results of studies on specific dietary acid supplementation in the
tilapia include the following: Improved growth performance upon the
addition of 2 g/kg of potassium diformate (67); increased activity of intestinal
protease upon the supplementation of citric acid (134); increased growth performance when
potassium-diformate at ≥5 g/kg was added to the diet (73); it reduced the total bacteria count
in the gut, and increased body weight and improved feed utilization
when the diet contained a combination of malic acid at 10 g/kg and
Bacillus subtilis at 1.1x105 led to an increased
in growth performance (112). In
other species such as sea bream, the supplementation of coated
sodium butyrate increased weight gain by 9.83% (135), supplementing 1.5% citric acids to
the diet juvenile red drum (Sciaenops ocellatus), improving
growth performance in terms of weight gain (43).
Similarly, sodium butyrate has been used to
supplement the diet of Carassius auratus (136). The effects of different organic
acids on the intestine vary substantially and depend on the dose of
the respective organic acid in the feed (103). It provides clear evidence that
the use of any of the three acids used to adjust the pH to 4.6
resulted in maximal growth and survival, while a markedly high (pH
8.0) and low pH (pH 2.5) may cause growth deprivation in the Nile
tilapia and mortality (86). This
result was supported by Romano et al (105) (Table
I) with the dietary supplementation of citric acid to the diet
of tilapia resulting in reduced growth as the acid concentration
increased and significantly decreased intestinal short-chain fatty
acid. Thus, at a dietary pH of 4.6, the following mechanisms may
improve nutrient utilization: Reducing gastric pH levels,
increasing digestive enzymes, increasing mineral solubilization
during digestion processes, or altered intestinal microbial
behavior, which may produce valuable nutrient contents.
5. Growth performance
The growth rate is a key indicator for determining
the economic efficiency of commercial fish farming, and it is
influenced by a variety of factors. Therefore, the impact of
various acidifiers on the growth of a variety of fish species has
been investigated. Organic acids, salts, or mixtures, according to
some findings, can improve the growth and feed utilization of
certain fish species (137).
Nonetheless, other studies have not found growth-promoting effects
of organic acids (67), which
appear to depend on the fish species, type of organic acid and
dosage used (5). Based on this
information, the possible beneficial effects of organic acids may
be species- and dosage-dependent. For example, a dose of 6.25 g/kg
sodium butyrate has been shown to improve the growth performance,
e.g., weight gain and specific growth rate in the tilapia (109). Likewise, a dose of 1% oxalic acid
+ malic acid and calcium lactate + sodium acetate has been found to
enhance growth and feed utilization (138).
Studies on the effects of citric acidified diets on
growth and feed performance have yielded positive results. A
mixture of organic acids (acetic, lactic and citric acid) was used
in a previous study to assess rainbow trout growth. Chen et
al (139) (Table I) examined the effect of dietary
L-malic acid on the growth and feed utilization of the genetically
improved farmed Tilapia. Various factors, such as the experimental
fish species and physiological age, the type and the level of
organic acids, the diet composition, and the culture conditions,
may all influence the growth-promoting effects of dietary organic
acids (72).
The findings in the study of Fabay et al
(86) re consistent with earlier
findings in the literature in the used of organic acid in the
aquaculture diet (Table I).
Optimum levels that have been shown to elicit maximal growth
performance are as follows: 1 or 2 g/kg of formic and propionic
acid (63), 0.3% potassium
diformate (115), 2 g/kg
potassium diformate (67), and
increased feed efficiency at 0.2 or 0.3% of potassium diformate
(80), 0.2% formic and propionic
acid (100). The observation that
a dietary pH 4.6 resulted in the optimal growth among the
treatments was supported by the study of Elala and Ragaa (77) (3 g/kg potassium diformate), Chen
et al (139) (Table I) (0.8 or 3.2% L-malic acid;
increased fish body weight and weight gain), Huan et al
(31) (organic acid blend;
increased weight gain), and Hassaan et al (108) (malic acid at 5; or 10 g/kg rose
weight gain and specific growth rate).
While the use organic acid in aquaculture can
improved growth rate, feed efficiency and fish health have been
reported in several studies (Table
I), Reda et al (63)
also reported the supplementation of the Nile tilapia diet with a
mixture of formic acid, propionic acid, and calcium propionate at
concentrations of 0.1 and 0.2%. The optimal dietary pH value has
been shown to be 4.6(86), which
affects the growth performance in the final average body weight.
Other dietary pH values higher or lower than pH 4.6 have resulted
in lower growth rates. Thus, the pH of feeds may affect fish
gastrointestinal or chyme pH.
A 0.922 g/kg organic acid blend (calcium
propionate, calcium formate and sodium acetate) supplemented
protease in meat and bone meal diet has been shown to improve
weight gain in the Nile tilapia (31). The optimal dietary pH value has
been found to be 4.6(86), which
affects optimal growth performance, manifesting in weight gain.
Other dietary pH values higher or lower than pH 4.6 result in lower
growth rates. The pH of feeds may affect fish gastrointestinal or
chyme pH. Citric acid (3%) has been found to induce weight gain in
the Beluga (90), Rohu (140) and common carp (141). In addition, a significant
increase in the final weight, weight gain and daily growth rate has
been observed in a growth trial experiment on Carassius
auratus fed a diet supplemented with apple cider vinegar
(142). In another study, 1%
citric acid improved weight gain in the yellowtail (143). In the case of the red drum, based
on the weight gain results from the study it appears that
acidifiers at a dose of 1.5% of the diet, especially citric acid,
may improve growth performance (weight gain). Chen et al
(139) (Table I) examined the effect of dietary
L-malic acid and demonstrated that both 0.8 and 3.2% L-malic acid
improved the weight gain compared to the control at 0.0%. A study
on the red hybrid tilapia (Oreochromis sp.) fed a diet
supplemented with 0.2% potassium diformate revealed a significant
decrease in the mortality rate following a challenge with
Streptococcus bacteria (from 58.3 to 16.6%) (74). Lim et al (144) also observed that graded dietary
potassium diformate up to 10 k/g improved weight gain and feed
efficiency in O. niloticus.
An increased protein digestibility has been
observed with the use of 0.2 and 0.3% potassium diformate compared
with the control diet by almost 6.75% (77). Furthermore, red hybrid tilapia fed
diets supplemented with 2 k/g potassium diformate have been shown
to exhibit a tendency towards an increased body weight, feed
utilization and nutrient digestibility (67).
A previous study on the South African abalone
(145) reported an increase
significantly in the specific growth rate compared with the control
when the abalone was fed a diet containing a mixture of sodium
benzoate and sodium sorbate. Citric acid (3%) has been found to
enhance the specific growth rate of the Beluga (90), Rohu (140), and common carp (141). A significant increase in the
specific growth rate has also been observed in Carassius
auratus fed a diet supplemented with apple cider vinegar
(142). The best dietary pH in
the Nile tilapia diet was pH 4.6(86), which affected the best growth
performance, which manifested a specific growth rate. Other dietary
pH values higher or lower than pH 4.6 resulted in lower growth
rates. Thus, the pH of feeds may have affected fish
gastrointestinal or chyme pH.
6. Feed efficiency
In a previous study, 1% citric acid was found to
improve weight and the feed conversion ratio in the red sea bream
(146). A higher feed conversion
ratio, total length, and feed intake were also found in an organic
acid-treated group (137).
Cuvin-Aralar et al (113)
reported an improved growth and feed conversion ratio (FCR) in the
juvenile Nile tilapia fed a diet supplemented with 0.3% potassium
diformate compared with the control diet. The primary explanation
for the improved growth efficiency and protein digestibility of
fish diet supplemented with potassium diformate is the pH levels in
the stomach and upper gut. In tilapia grow-out in Indonesia, Ramli
et al (147) studied
potassium diformate (potassium salt of formic acid) as a growth
promoter. In this sample, fish were fed six times a day with a diet
containing various percentages (0, 0.2 0.3, and 0.5) of potassium
diformate over a period of 85 days. The diets included 32% crude
protein, 25% carbohydrates, 6% lipids and 10% fiber. Potassium
diformate markedly increased feed consumption over the whole
feeding period, from day 1 to 85, and weight gain significantly
improved the feed conversion ratio.
Furthermore, the protein efficiency ratio was also
improved considerably due to the addition of the formic acid salt.
The potassium diformate additions of 0.2 and 0.5% yielded optimal
results. The authors concluded that applying the potassium
diformate at 0.2% was an efficient tool to control bacterial
infections in tropical cultures (74).
At the same concentration, citric acid was shown to
improve feed performance in the red sea bream (148), and the protein efficiency ratio
in the Beluga (90) and Rohu
(140). In the study by Ryan
et al (118), pH affects
the optimal feed efficiency banifested by the protein efficiency
ratio. Other dietary pH values higher or lower than pH 4.6 result
in lower growth rates. Thus, the pH of feeds may affect fish
gastrointestinal or chyme pH.
It has been demonstrated that the shrimp diet
supplemented with acidifiers improves digestibility (149-151).
However, the evaluation is limited to Litopenaeus vannamei
diets supplemented with butyrate and propionate (149). In another study, formic, lactic,
malic and citric were incorporated into shrimp diets. The results
revealed that with addition of acid, increased the growth of
Litopenaeus vannamei (148) and Penaeus monodon
(150). The diets of
Litopenaeus vannamei were supplemented with 0.5%, potassium
diformate increased productivity over the control diet. In
addition, these organic acids and their salts increase nitrogen and
phosphorus retention and bioavailability of calcium and phosphorus
(79,90,152). The effects of vinegar and sodium
acetate on the growth performance of Pacific white shrimp
(Penaeus vannamei) were examined by Serrano et al
(151). The results revealed that
the supplementation of organic vinegar to the diet improved the
growth performance (final average body weight, weight gain and
specific growth rate) and feed utilization (feed conversion ratio
and protein efficiency ratio).
7. Conclusions
Feed researchers, developers and manufacturers are
being encouraged to use plant protein ingredients to formulate
feeds due to a global shortage of fish meals. However, the presence
of anti-nutritional factors and imbalanced amino acid profiles in
plant protein sources are obstacles they face. Their digestibility
and growth performance are poor due to these two factors, and their
use in aquafeeds is restricted. In conclusion, with the addition of
acid to the diet, pepsin activation and mineral absorption
increase, resulting in improved growth and feed utilization. In the
dietary acidification of tilapia, the growth performance induced by
the use of acetic, citric and hydrochloric acids in the feed has
revealed that dietary pH and not a specific acid were the most
crucial improvements in the fish stomach, specifically in
monogastric species. This is the basis for vertebrates; two gastric
acidification strategies have been discovered. The first are those
that keep the stomach acidic at all times, regardless of whether
food is present (e.g., mammals and birds). The second is to
maintain the pH of the stomach lumen at a neutral level between
meals and to subsequently become slightly acidic. However, the
majority of studies on teleost fish studied thus far have used this
second strategy. On the other hand, certain types of fish, such as
cobia juveniles, rainbow trout, southern bluefin tuna and some
Elasmobranchii species, have been observed using the first
strategy, strictly carnivorous. Studies have compared fed and
fasted fish feeding in the gilthead seabream; erratic daily feeding
by changing the time of feed delivery at random can change the
daily pattern from neutral/acid alternation to permanent
acidification.
Following feeding in fish, pepsinogen is activated
to begin the hydrolysis of the ingested prey to a constant acidic
gastric pH. In the case of dietary acidification, the continuous
feeding of acidic diets at various pH levels or amounts to certain
types of fish, such as the tilapia and carp, may lead to a
condition of permanent acidification of the lumen (i.e., the first
strategy of gastric acidification). Thus, this dietary
acidification follows the first strategy. This refers to a
situation in which the production of digestive enzymes or the
activation of existing digestive enzymes was easily achieved to
improve nutrient utilization. Thus, the data on the fish growth
rate and feed utilization efficiency were reviewed on dietary acid
sources, such as acetic acid, citric acid hydrochloric acid. In
addition, the attractability of the diets at various pH levels and
dietary acid sources is another factor to determine the efficiency
of the diet. The survival rates of cultured fish were increased
based on the various dietary acids used that match the requirements
of the fishes. Any acids at an optimum pH level, as for example pH
4.6 in the diet of the tilapia fry, which increased attractability,
growth and feed efficiency warrant further attention. In general,
the addition of acid to the diet, specifically in monogastric fish,
such as the carp (no stomach) and tilapia (herbivorous, less acid
produced in the stomach), can improve growth, and feed efficiency
digestibility and mineral absorption in a culture system. Thus, the
inclusion of acid in aquaculture diets may have promising results
for feed manufacturers.
Furthermore, these dietary acidifications can
eliminate the impact of bacterial infections to prevent diseases
and result in high survival. Acids in the diet can efficiently
achieve a sustainable, economic and safe fish production. Further
studies are required to focus on cultured fish with different
dietary pH levels in herbivorous and omnivorous species, such as
milkfish and carp.
Acknowledgements
The authors would like to thank the DOST-SEI for
the scholarship of RVF (ASTHRDP) and the collaborative project of
the DOST-PCAARRD and SEI for the scholarship of MSA (GREAT
Program). The authors are also grateful to the Mindanao State
University-Maguindanao for granting permission RVF to perform the
study under the faculty development program.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
All authors (RVF, AER, MSA and JVF) contributed to
the conception and design of the study. In addition, all authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hartviksen M, Bakke AM, Vecino JG, Ringø E
and Krogdahl Å: Evaluation of the effect of commercially available
plant and animal protein sources in diets for Atlantic salmon
(Salmo salar L.): Digestive and metabolic investigations.
Fish Physiol Biochem. 40:1621–1637. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Urán PA, Schrama JW, Rombout JH,
Taverne-Thiele JJ, Obach A, Koppe W and Verreth JA: Time-related
changes of the intestinal morphology of Atlantic salmon, Salmo
salar L., at two different soybean meal inclusion levels. J
Fish Dis. 32:733–744. 2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yamamoto T, Suzuki N, Furuita H, Sugita T,
Tanaka N and Goto T: Supplemental effect of bile salts to soybean
meal-based diet on growth and feed utilization of rainbow trout
Oncorhynchus mykiss. Fish Sci. 73:123–131. 2007.
|
|
4
|
Castillo S and Gatlin DM: Dietary
supplementation of exogenous carbohydrase enzymes in fish
nutrition: A review. Aquaculture. 435:286–292. 2015.
|
|
5
|
Lückstädt C: The use of acidifiers in fish
nutrition. CAB Rev Perspect Agric Veterinary. 3:1–8. 2008.
|
|
6
|
Krogdahl Å, Penn M, Thorsen J, Refstie S
and Bakke AM: Important antinutrients in plant feedstuffs for
aquaculture: An update on recent findings regarding responses in
salmonids. Aquac Res. 41:333–344. 2010.
|
|
7
|
Bone RM: Pathological εffects of soybean
anti-nutritional factors on summer flounder (Paralichthys
Dentatus) tissues. 2013. Available from: https://digitalcommons.uri.edu/theses/57.
|
|
8
|
Gargiulo AM, Ceccarelli P, Dall'Aglio C
and Pedini V: Histology and ultrastructure of the gut of the
tilapia (Tilapia spp.), a hybrid teleost. Anat Histol
Embryol. 27:89–94. 1998.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Smith BJ, Smith SA, Tengjaroenkul B and
Lawrence TA: Gross morphology and topography of the adult
intestinal tract of the tilapian fish, Oreochromis niloticus
L. Cells Tissues Organs. 166:294–303. 2000.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ferraris RP and Ahearn GA: Sugar and amino
acid transport in fish intestine. Comp Biochem Physiol A Mol Integr
Physiol. 77:397–413. 1984.
|
|
11
|
German DP, Neuberger DT, Callahan MN,
Lizardo NR and Evans DH: Feast to famine: The effects of food
quality and quantity on the gut structure and function of a
detritivorous catfish (Teleostei: Loricariidae). Comp Biochem
Physiol Part A Mol Integr Physiol. 155:281–293. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Titus E and Ahearn GA: Short-chain fatty
acid transport in the intestine of a herbivorous teleost. J Exp
Biol. 135:77–94. 1988.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Titus E and Ahearn GA: Transintestinal
acetate transport in a herbivorous teleost: Anion exchange at the
basolateral membrane. J Exp Biol. 156:41–61. 1991.
|
|
14
|
Newsome SD, Fogel ML, Kelly L and del Rio
CM: Contributions of direct incorporation from diet and microbial
amino acids to protein synthesis in Nile tilapia. Funct Ecol.
25:1051–1062. 2011.
|
|
15
|
Niklasson L: Intestinal mucosal immunology
of Salmonids-response to stress and infection and crosstalk with
the physical barrier. Gothenburg University, Gothenburg, 2013.
|
|
16
|
Baeverfjord G and Krogdahl A: Development
and regression of soybean meal induced enteritis in Atlantic
salmon, Salmo salar L., distal intestine: A comparison with
the intestines of fasted fish. J Fish Dis. 19:375–387. 1996.
|
|
17
|
Steiner T and Encarnacão P: Latest trends
in gut health management. Aquafeed Advances in Processing and
Formulation. Kailua, Hawaii, pp9-10, 2010.
|
|
18
|
Tram NDQ, Ngoan LD, Hung LT and Lindberg
JE: A comparative study on the apparent digestibility of selected
feedstuffs in hybrid catfish (Clarias macrocephalus x
Clarias gariepinus) and Nile tilapia (Oreochromis
niloticus). Aquac Nutr. 17:636–643. 2011.
|
|
19
|
Vidal L, Xavier TO, Michelato M, Martins
EN, Pezzato LE and Furuya WM: Apparent protein and energy
digestibility and amino acid availability of corn and co-products
in extruded diets for Nile Tilapia, Oreochromis niloticus. J
World Aquac Soc. 46:183–190. 2015.
|
|
20
|
Figueiredo-Silva C, Lemme A, Sangsue D and
Kiriratnikom S: Effect of DL-methionine supplementation on the
success of almost total replacement of fish meal with soybean meal
in diets for hybrid tilapia (Oreochromis niloticus x
Oreochromis mossambicus). Aquac Nutr. 21:234–241. 2015.
|
|
21
|
Tran-Ngoc KT, Huynh ST, Sendão J, Nguyen
TH, Roem AJ, Verreth JAJ and Schrama JW: Environmental conditions
alter the effect of organic acid salts on digestibility and
intestinal morphology in Nile tilapia (Oreochromis
niloticus). Aquac Nutr. 25:134–144. 2019.
|
|
22
|
Kumar S and Tembhre M: Anatomy and
physiology of Fishes. New Delhi, India, Vikas Pub. House Pvt, 1996,
1998.
|
|
23
|
Ikpegbu E, Ezeasor DN, Nlebedum UC and
Nnadozie O: Morphological and histochemical observations on the
oesogaster of the domesticated African catfish (Clarias
Gariepinus Burchell, 1822). Bulg J Vet Med. 16:88–95. 2013.
|
|
24
|
Grau A, Crespo S, Sarasquete MC and De
Canales MG: The digestive tract of the amberjack Seriola
dumerili, Risso: A light and scanning electron microscope
study. J Fish Biol. 41:287–303. 1992.
|
|
25
|
Cao XJ and Wang WM: Histology and mucin
histochemistry of the digestive tract of yellow catfish,
Pelteobagrus fulvidraco. Anat Histol Embryol. 38:254–261.
2009.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Phrompanya P, Saenphet K and Saenphet S:
Comparative histochemical study of the gastrointestinal tracts of
the Nile tilapia (Oreochromis niloticus) and the hybrid
catfish (Clarias batrachus x Clarias gariepinus).
Acta Histochem. 121:261–267. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Murray HM, Wright GM and Goff GP: A
comparative histological and histochemical study of the
post-gastric alimentary canal from three species of pleuronectid,
the Atlantic halibut, the yellowtail flounder and the winter
flounder. J Fish Biol. 48:187–206. 1996.
|
|
28
|
Neuhaus H, Van der Marel M, Caspari N,
Meyer W, Enss ML and Steinhagen D: Biochemical and histochemical
study on the intestinal mucosa of the common carp Cyprinus
carpio L., with special consideration of mucin glycoproteins. J
Fish Biol. 70:1523–1534. 2007.
|
|
29
|
Sarasquete C, Gisbert E, Ribeiro L, Vieira
L and Dinis MT: Glyconjugates in epidermal, branchial and digestive
mucous cells and gastric glands of gilthead sea bream, Sparus
aurata, Senegal sole, Solea senegalensis and
Siberian sturgeon, Acipenser baeri development. Eur J
Histochem. 45:267–278. 2001.PubMed/NCBI View
Article : Google Scholar
|
|
30
|
Lobel PS: Trophic biology of herbivorous
reef fishes: Alimentary pH and digestive capabilities. J Fish Biol.
19:365–397. 1981.
|
|
31
|
Huan D, Li X, Chowdhury MAK, Yang H, Liang
G and Leng X: Organic acid salts, protease and their combination in
fish meal-free diets improved growth, nutrient retention and
digestibility of tilapia (Oreochromis niloticus x O.
aureus). Aquac Nutr. 24:1813–1821. 2018.
|
|
32
|
Cho CY and Bureau DP: Reduction of waste
output from salmonid aquaculture through feeds and feeding.
Progress Fish Cult. 59:155–160. 1997.
|
|
33
|
Francis G, Makkar HP and Becker K:
Antinutritional factors present in plant-derived alternate fish
feed ingredients and their effects in fish. Aquaculture.
199:197–227. 2001.
|
|
34
|
Koshio S, Ackman RG and Lall SP: Effects
of oxidized herring and canola oils in diets on growth, survival,
and flavor of Atlantic salmon, Salmo salar. J Agric Food
Chem. 42:1164–1169. 1994.
|
|
35
|
Krogdahl Å, Hemre GI and Mommsen TP:
Carbohydrates in fish nutrition: Digestion and absorption in
postlarval stages. Aquac Nutr. 11:103–122. 2005.
|
|
36
|
Kraugerud OF, Penn M, Storebakken T,
Refstie S, Krogdahl Å and Svihus B: Nutrient digestibilities and
gut function in Atlantic salmon (Salmo salar) fed diets with
cellulose or non-starch polysaccharides from soy. Aquaculture.
273:96–107. 2007.
|
|
37
|
Tran-Ngoc KT, Dinh NT, Nguyen TH, Roem AJ,
Schrama JW and Verreth JAJ: Interaction between dissolved oxygen
concentration and diet composition on growth, digestibility and
intestinal health of Nile tilapia (Oreochromis niloticus).
Aquaculture. 462:101–108. 2016.
|
|
38
|
Gao Y, Storebakken T, Shearer KD, Penn M
and Øverland M: Supplementation of fishmeal and plant protein-based
diets for rainbow trout with a mixture of sodium formate and
butyrate. Aquaculture. 311:233–240. 2011.
|
|
39
|
Hamer HM, Jonkers D, Venema K, Vanhoutvin
S, Troost F and Brummer RJ: The role of butyrate on colonic
function. Aliment Pharmacol Ther. 27:104–119. 2008.
|
|
40
|
Kuz'mina V: Classical and modern concepts
in fish digestion. In: Feeding and Digestive Functions in Fishes,
1st Edition. CRC Press, p70, 2008.
|
|
41
|
Caceci T, El-Habback HA, Smith SA and
Smith BJ: The stomach of Oreochromis niloticus has three
regions. J Fish Biol. 50:939–952. 1997.
|
|
42
|
Wilson JM and Castro LFC: Morphological
diversity of the gastrointestinal tract in fishes. Fish Physiol.
30:1–55. 2010.
|
|
43
|
Castillo S, Rosales M, Pohlenz C and
Gatlin DM: Effects of organic acids on growth performance and
digestive enzyme activities of juvenile red drum Sciaenops
ocellatus. Aquaculture. 433:6–12. 2014.
|
|
44
|
Mroz Z, Jongbloed AW, Partanen KH, Vreman
K, Kemme PA and Kogut J: The effects of calcium benzoate in diets
with or without organic acids on dietary buffering capacity,
apparent digestibility, retention of nutrients, and manure
characteristics in swine. J Anim Sci. 78:2622–2632. 2000.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Fruton JS: Hydrolysis: Peptide bonds.
Academic Press Inc. New York. 3:120–152. 1971.
|
|
46
|
Effront J, Prescott SC and Venable CS:
Biochemical catalysts in life and industry: Proteolytic enzymes.
Kessinger Publishing. 151–289. 2007.
|
|
47
|
Bobe J and William Goetz F: An ovarian
progastricsin is present in the trout coelomic fluid after
ovulation. Biol Reprod. 64:1048–1055. 2001.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Kurokawa T, Uji S and Suzuki T:
Identification of pepsinogen gene in the genome of stomachless
fish, Takifugu rubripes. Comp Biochem Physiol B Biochem Mol
Biol. 140:133–140. 2005.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Gildberg A: Recovery of proteinases and
protein hydrolysates from fish viscera. Bioresour Technol.
39:271–276. 1992.
|
|
50
|
Simpson BK: Digestive proteinases from
marine animals. Food Science and Technology-New York-Marcel Dekker.
191–214. 2000.
|
|
51
|
Raufman JP: Pepsin, In: Encyclopedia of
Gastroenterology. Academic Press, San Diego, 2004.
|
|
52
|
Deguara S, Jauncey K and Agius C: Enzyme
activities and pH variations in the digestive tract of gilthead sea
bream. J Fish Biol. 62:1033–1043. 2003.
|
|
53
|
Langley JN: On the changes in serous
glands during secretion. J Physiol. 2(261)1879.
|
|
54
|
Hirschowitz BI: Urinary excretion of
pepsinogen in gastroduodenal ulceration. Lancet. 261:66–69.
1953.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Hirschowitz BI and O'Leary DK: Dose
dependence of insulin-stimulated gastric secretion. Am J Dig
Dis. 9:379–397. 1964.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Zhao L, Budge SM, Ghaly AE and Brooks MS:
Extraction, purification and characterization of fish pepsin: A
critical review. Int J Food Proc Technol. 2(6)2011.
|
|
57
|
Márquez L, Robles R, Morales GA and Moyano
FJ: Gut pH as a limiting factor for digestive proteolysis in
cultured juveniles of the gilthead sea bream (Sparus
aurata). Fish Physiol Biochem. 38:859–869. 2012.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Chakrabarti I, Gani MA, Chaki KK, Sur R
and Misra KK: Digestive enzymes in 11 freshwater teleost fish
species in relation to food habit and niche segregation.
Comparative Biochem Physiol. 112:167–177. 1995.
|
|
59
|
Hsu YL and Wu JL: Relationship between
feeding-habits and digestive proteases of some freshwater-fishes.
Bull Inst Zool Acad Sin. 18:45–53. 1979.
|
|
60
|
Uys W and Hecht T: Assays on the digestive
enzymes of sharptooth catfish, Clarias gariepinus (Pisces:
Clariidae). Aquaculture. 63:301–313. 1987.
|
|
61
|
Sabapathy U and Teo LH: Some properties of
the intestinal proteases of the rabbitfish, Siganus
canaliculatus (Park). Fish Physiol Biochem. 14:215–221.
1995.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Hassaan MS, Soltan MA, Jarmołowicz S and
Abdo HS: Combined effects of dietary malic acid and Bacillus
subtilis on growth, gut microbiota and blood parameters of Nile
tilapia (Oreochromis niloticus). Aquac Nutr. 24:83–93.
2018.
|
|
63
|
Reda RM, Mahmoud R, Selim KM and El-Araby
IE: Effects of dietary acidifiers on growth, hematology, immune
response and disease resistance of Nile tilapia, Oreochromis
niloticus. Fish Shellfish Immunol. 50:255–262. 2016.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Sturkie PD (ed): Avian Physiology.
Springer Science and Business Media, 2012.
|
|
65
|
Defoirdt T, Boon N, Sorgeloos P,
Verstraete W and Bossier P: Short-chain fatty acids and
poly-beta-hydroxyalkanoates: (New) Biocontrol agents for a
sustainable animal production. Biotechnol Adv. 27:680–685.
2009.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Gislason G, Olsen RE and Hinge E:
Comparative effects of dietary Na+-lactate on Arctic
char, Salvelinus alpinus L., and Atlantic salmon, Salmo
salar L. Aquac Res. 27:429–435. 1996.
|
|
67
|
Ng WK, Koh CB, Sudesh K and Siti-Zahrah A:
Effects of dietary organic acids on growth, nutrient digestibility
and gut microflora of red hybrid tilapia, Oreochromis sp.,
and subsequent survival during a challenge test with
Streptococcus agalactiae. Aquac Res. 40:1490–1500. 2009.
|
|
68
|
Lückstädt C: Use of organic acids as feed
additives-sustainable aquaculture production the non-antibiotic
way. Int Aquafeed. 9:21–26. 2006.
|
|
69
|
Owen MA, Waines P, Bradley G and Davies S:
The effect of dietary supplementation of sodium butyrate on the
growth and microflora of Clarias gariepinus (Burchell 1822).
The XII International Symposium Fish Nutrition and Feeding,
2006.
|
|
70
|
Denev S, Staykov Y, Moutafchieva R and
Beev G: Microbial ecology of the gastrointestinal tract of fish and
the potential application of probiotics and prebiotics in finfish
aquaculture. Int Aquat Res. 1:1–29. 2009.
|
|
71
|
Baruah K, Pal AK, Sahu NP, Debnath D,
Yengkokpam S, Norouzitallab P and Sorgeloos P: Dietary crude
protein, citric acid and microbial phytase interacts to influence
the hemato-immunological parameters of rohu, Labeo Rohita,
Juveniles. J World Aquac Soc. 40:824–831. 2009.
|
|
72
|
Knarreborg A, Miquel N, Granli T and
Jensen BB: Establishment and application of an in vitro methodology
to study the effects of organic acids on coliform and lactic acid
bacteria in the proximal part of the gastrointestinal tract of
piglets. Anim Feed Sci Technol. 99:131–140. 2002.
|
|
73
|
Liebert F, Mohamed K and Lückstädt C:
Effects of diformates on growth and feed utilization of all male
Nile Tilapia fingerlings. (Oreochromis niloticus) reared in
tank culture, 2010.
|
|
74
|
Zhou Z, Liu Y, He S, Shi P, Gao X, Yao B
and Ringø E: Effects of dietary potassium diformate (KDF) on growth
performance, feed conversion and intestinal bacterial community of
hybrid tilapia (Oreochromis niloticus ♀ × O. aureus
♂). Aquaculture. 291:89–94. 2009.
|
|
75
|
Russell JB and Diez-Gonzalez F: The
effects of fermentation acids on bacterial growth. Adv Microb
Physiol. 39:205–234. 1997.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Balcázar JL, De Blas I, Ruiz-Zarzuela I,
Vendrell D, Gironés O and Muzquiz JL: Enhancement of the immune
response and protection induced by probiotic lactic acid bacteria
against furunculosis in rainbow trout (Oncorhynchus mykiss).
FEMS Microbiol Immunol. 51:185–193. 2007.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Abu Elala NM and Ragaa NM: Eubiotic effect
of a dietary acidifier (potassium diformate) on the health status
of cultured Oreochromis niloticus. J Adv Res. 6:621–629.
2015.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Baruah SK, Norouzitallab P, Debnath D, Pal
AK and Sahu NP: Organic acids as non-antiobiotic nutraceuticals in
fish and prawn feed. Aquac Health Int. 4–6. 2008.
|
|
79
|
Hossain MA, Pandey A and Satoh S: Effects
of organic acids on growth and phosphorus utilization in red sea
bream Pagrus major. Fish Sci. 73:1309–1317. 2007.
|
|
80
|
Morken T, Kraugerud OF, Barrows FT,
Sørensen M, Storebakken T and Øverland M: Sodium diformate and
extrusion temperature affect nutrient digestibility and physical
quality of diets with fish meal and barley protein concentrate for
rainbow trout (Oncorhynchus mykiss). Aquaculture.
317:138–145. 2011.
|
|
81
|
Øverland M, Granli T, Kjos NP, Fjetland O,
Steien SH and Stokstad M: Effect of dietary formates on growth
performance, carcass traits, sensory quality, intestinal
microflora, and stomach alterations in growing-finishing pigs. J
Anim Sci. 78:1875–1884. 2000.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Ng WK and Koh CB: Application of organic
acids in aquafeeds: Impacts on fish growth, nutrient utilisation
and disease resistance. In: Standards for Acidifiers, Principles
for the Use of Organic Acids in Animal Nutrition. Edited by
Lückstädt C, Nottingham University Press, 49-58, 2011.
|
|
83
|
Lim C: Effect of dietary pH on amino acid
utilization by shrimp (Penaeus vannamei). Aquaculture.
114:293–303. 1993.
|
|
84
|
Wilson RP, Harding DE and Garling DL:
Effect of dietary pH on amino acid utilization and the lysine
requirement of fingerling channel catfish. J Nutr. 107:166–170.
1977.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Ringø E, Olsen RE and Castell JD: Effect
of dietary lactate on growth and chemical composition of Arctic
charr Salvelinus alpinus. J World Aquac Soc. 25:483–486.
1994.
|
|
86
|
Fabay RV, Tumbokon BL and Serrano AE Jr:
Effects of dietary pH and acid source on growth and feed efficiency
of the Nile Tilapia, Oreochromis niloticus fry. Isr J Aquac.
72(1114685)2020.
|
|
87
|
Baruah K, Pal AK, Sahu NP, Jain KK,
Mukherjee SC and Debnath D: Dietary protein level, microbial
phytase, citric acid and their interactions on bone mineralization
of Labeo rohita (Hamilton) juveniles. Aquac Res. 36:803–812.
2005.
|
|
88
|
Chikwati EM, Sahlmann C, Holm H, Penn MH,
Krogdahl Å and Bakke AM: Alterations in digestive enzyme activities
during the development of diet-induced enteritis in Atlantic
salmon, Salmo salar L. Aquaculture. 402:28–37. 2013.
|
|
89
|
Krogdahl Å, Gajardo K, Kortner TM, Penn M,
Gu M, Berge GM and Bakke AM: Soya saponins induce enteritis in
atlantic salmon (Salmo salar L.). J Agric Food Chem.
63:3887–3902. 2015.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Khajepour F and Hosseini SA: Citric acid
improves growth performance and phosphorus digestibility in Beluga
(Huso huso) fed diets where soybean meal partly replaced
fish meal. Anim Feed Sci Technol. 171:68–73. 2012.
|
|
91
|
Nuez-Ortin WG: Gustor-Aqua: An effective
solution to optimize health status and nutrient utilization. Int
Aquafeed. 18–20. 2011.
|
|
92
|
Hidaka I, Zeng C and Kohbara J: Gustatory
responses to organic acids in the yellowtail Seriola
quinqueradiata. Nippon Suisan Gakk. 58:1179–1187. 1992.
|
|
93
|
Vielma J and Lall SP: Dietary formic acid
enhances apparent digestibility of minerals in rainbow trout,
Oncorhynchus mykiss (Walbaum). Aquac Nutr. 3:265–268.
1997.
|
|
94
|
Busti S, Rossi B, Volpe E, Ciulli S, Piva
A, D'Amico F and Parma L: Effects of dietary organic acids and
nature identical compounds on growth, immune parameters and gut
microbiota of European sea bass. Sci Rep. 10:1–14. 2020.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Hlophe SN, Moyo NAG and Ncube I:
Postprandial changes in pH and enzyme activity from the stomach and
intestines of Tilapia rendalli (B oulenger, 1897),
Oreochromis mossambicus (Peters, 1852) and Clarias
gariepinus (Burchell, 1822). J Appl Ichthyol. 30:35–41.
2014.
|
|
96
|
De Wet L: Can organic acid effectively
replace antibiotic growth promotants in diets for rainbow trout,
Oncorhynchus mykiss raised under suboptimal water
temperatures. WAS Conference, Bali, Indonesia, 2005.
|
|
97
|
Blank R, Mosenthin R, Sauer WC and Huang
S: Effect of fumaric acid and dietary buffering capacity on ileal
and fecal amino acid digestibilities in early weaned pigs. J Anim
Sci. 77:2974–2984. 1999.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Sugiura SH, Dong FM, Rathbone CK and Hardy
RW: Apparent protein digestibility and mineral availabilities in
various feed ingredients for salmonid feeds. Aquaculture.
159:177–202. 1998.
|
|
99
|
Kirchgessner M: Fumaric acid as a feed
additive in pig nutrition. Pig News Inform. 3:259–264. 1982.
|
|
100
|
Gildberg A and Raa J: Properties of a
propionic acid/formic acid preserved silage of cod viscera. J Sci
Food Agric. 28:647–653. 1977.
|
|
101
|
Asgard T and Austreng E: Fish silage for
salmonids: A cheap way of utilizing waste as feed. Feedstuffs.
53:22–24. 1981.
|
|
102
|
Batista I: Fish silage: Preparation and
uses. In: Nutrition in Marine Aquaculture. Bruno A. (ed).
FAO/UNDP/MEDRAP, 227-48, 1987.
|
|
103
|
Lückstädt C: Dietary Acidification in
Aquaculture. Global Aquaculture Advocate, 2015.
|
|
104
|
Partanen KH and Mroz Z: Organic acids for
performance enhancement in pig diets. Nutr Res Rev. 12:117–145.
1999.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Romano N, Simon W, Ebrahimi M, Fadel AH,
Chong CM and Kamarudin MS: Dietary sodium citrate improved
oxidative stability in red hybrid tilapia (Oreochromis sp.)
but reduced growth, health status, intestinal short chain fatty
acids and induced liver damage. Aquaculture. 458:170–176. 2016.
|
|
106
|
Ng WK and Koh CB: The utilization and mode
of action of organic acids in the feeds of cultured aquatic
animals. Rev Aquac. 9:342–368. 2017.
|
|
107
|
Mackie AM: Identification of the gustatory
feeding stimulants. In Chemoreception in fishes. Edited by Hara TJ,
Elsevier, 275-291, 1982.
|
|
108
|
Hassaan MS, Mohammady EY, Adnan AM, Abd
Elnabi HE, Ayman MF, Magdy AM and El-Haroun ER: Effect of dietary
protease at different levels of malic acid on growth, digestive
enzymes and haemato-immunological responses of Nile tilapia, fed
fish meal free diets. Aquaculture: 522, 2020.
|
|
109
|
El-Naby ASA, Khattaby AE, Samir F, Awad SM
and Abdel-Tawwab M: Stimulatory effect of dietary butyrate on
growth, immune response, and resistance of Nile tilapia,
Oreochromis niloticus against Aeromonas hydrophila
infection. Anim Feed Sci Technol: Jul, 2019 (Epub ahead of print).
doi: 10.1016/j.anifeedsci.2019.114212.
|
|
110
|
Askarian F, Kousha A, Salma W and Ringø E:
The effect of lactic acid bacteria administration on growth,
digestive enzyme activity and gut microbiota in Persian sturgeon
(Acipenser persicus) and beluga (Huso huso) fry.
Aquac Nutr. 17:488–497. 2011.
|
|
111
|
Parma L, Yúfera M, Navarro-Guillén C,
Moyano FJ, Soverini M, D'Amico F and Bonaldo A: Effects of calcium
carbonate inclusion in low fishmeal diets on growth,
gastrointestinal pH, digestive enzyme activity and gut bacterial
community of European sea bass (Dicentrarchus labrax L.)
juveniles. Aquaculture. 510:283–292. 2019.
|
|
112
|
Giger-Reverdin S, Duvaux-Ponter C, Sauvant
D, Martin O, do Prado IN and Müller R: Intrinsic buffering capacity
of feedstuffs. Anim Feed Sci Technol. 96:83–102. 2002.
|
|
113
|
Cuvin-Aralar M, Kuhlmann KJ, Schroeder K
and Luckstadt C: Effect of potassium diformate (KDF) on growth
performance of male Nile tilapia (Oreochromis niloticus).
In: XIV International Symposium on Fish Nutrition and Feeding. Book
of Abstracts, Qingdao, p187, 2010.
|
|
114
|
Jun-Sheng L, Jian-Lin L and Ting-Ting W:
Ontogeny of protease, amylase and lipase in the alimentary tract of
hybrid juvenile tilapia (Oreochromis niloticus ×
Oreochromis aureus). Fish Physiol Biochem. 32:295–303.
2006.
|
|
115
|
Lückstädt C and Theobald P: Dose dependent
effects of diformate on broiler performance. In: Standards for
Acidifiers-Principles for the use of organic acids in animal
nutrition. Edited by Lückstädt C, Nottingham University Press,
pp83-87, 2011.
|
|
116
|
Lević J, Prodanović O and Sredanović S:
Understanding the buffering capacity in feedstuffs. Biotechnol
Animal Husbandry. 21:309–313. 2005.
|
|
117
|
Munilla-Morán R and Saborido-Rey F:
Digestive enzymes in marine species. I. Proteinase activities in
gut from redfish (Sebastes mentella), seabream (Sparus
aurata) and turbot (Scophthalmus maximus). Comp Biochem
Physiol B Biochem Mol Biol. 113:395–402. 1996.
|
|
118
|
Lückstädt C, Şenköylü N, Akyürek H and
Aĝma A: Acidifier-a modern alternative for antibiotic-free feeding
in livestock growth, with special focus on broiler growth. Vet Ir
Zootech. 27:91–93. 2004.
|
|
119
|
Blanchard B, Pompon D and Ducrocq C:
Nitrosation of melatonin by nitric oxide and peroxynitrite. J
Pineal Res. 29:184–192. 2000.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Yúfera M, Moyano FJ, Astola A,
Pousão-Ferreira P and Martínez-Rodríguez G: Acidic digestion in a
teleost: Postprandial and circadian pattern of gastric pH, pepsin
activity, and pepsinogen and proton pump mRNAs expression. PLoS
One. 7(e33687)2012.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Yúfera M, Nguyen MV, Navarro-Guillén C,
Moyano FJ, Jordal AO, Espe M and Rønnestad I: Effect of increased
rearing temperature on digestive function in cobia early juvenile.
Comp Biochem Physiol A Mol Integr Physiol. 230:71–80.
2019.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Lückstädt C and Schulz C: The dietary
effects of potassium diformate on the protein and fat digestibility
of Atlantic salmon (Salmo salar) reared in sea water.
Aquaculture Europe, Krakow, Poland, 391-392, 2008.
|
|
123
|
Moriarty DJW: The physiology of digestion
of blue-green algae in the cichlid fish, Tilapia nilotica. J
Zool. 171:25–39. 1973.
|
|
124
|
Bucking C and Wood CM: The effect of
postprandial changes in pH along the gastrointestinal tract on the
distribution of ions between the solid and fluid phases of chyme in
rainbow trout. Aquac Nutr. 15:282–296. 2009.
|
|
125
|
Escaffre AM, Kaushik S and Mambrini M:
Morphometric evaluation of changes in the digestive tract of
rainbow trout (Oncorhynchus mykiss) due to fish meal
replacement with soy protein concentrate. Aquaculture. 273:127–138.
2007.
|
|
126
|
Gill N, Higgs DA, Skura BJ, Rowshandeli M,
Dosanjh BS, Mann J and Gannam AL: Nutritive value of partially
dehulled and extruded sunflower meal for post-smolt Atlantic salmon
(Salmo salar L.) in sea water. Aquac Res. 37:1348–1359.
2006.
|
|
127
|
Nikolopoulou D, Moutou KA, Fountoulaki E,
Venou B, Adamidou S and Alexis MN: Patterns of gastric evacuation,
digesta characteristics and pH changes along the gastrointestinal
tract of gilthead sea bream (Sparus aurata L.) and European
sea bass (Dicentrarchus labrax L.). Comp Biochem Physiol A
Mol Integr Physiol. 158:406–414. 2011.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Boge G, Leydet M and Houvet D: The effects
of hexavalent chromium on the activity of alkaline phosphatase in
the intestine of rainbow trout (Oncorhynchus mykiss). Aquat
Toxicol. 23:247–260. 1992.
|
|
129
|
Papastamatiou YP, Purkis SJ and Holland
KN: The response of gastric pH and motility to fasting and feeding
in free swimming blacktip reef sharks, Carcharhinus
melanopterus. J Exp Mar Biol Ecol. 345:129–140. 2007.
|
|
130
|
Leef MJ, Carter CG and Nowak BF:
Assessment of nutritional status and digestive physiology in
southern bluefin tuna Thunnus maccoyii fed a modified
baitfish diet. Aquaculture. 350:162–168. 2012.
|
|
131
|
Papastamatiou YP and Lowe CG: Variations
in gastric acid secretion during periods of fasting between two
species of shark. Comp Biochem Physiol Part A Mol Integr Physiol.
141:210–214. 2005.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Papastamatiou YP, Purkis SJ and Holland
KN: The response of gastric pH and motility to fasting and feeding
in free swimming blacktip reef sharks, Carcharhinus
melanopterus. J Exp Mar Biol Ecol. 345:129–140. 2007.
|
|
133
|
Montoya A, López-Olmeda JF, Yúfera M,
Sánchez-Muros MJ and Sánchez-Vázquez FJ: Feeding time synchronises
daily rhythms of behaviour and digestive physiology in gilthead
seabream (Sparus aurata). Aquaculture. 306:315–321.
2010.
|
|
134
|
Su X, Li X, Leng X, Tan C, Liu B, Chai X
and Guo T: The improvement of growth, digestive enzyme activity and
disease resistance of white shrimp by the dietary citric acid.
Aquac Int. 22:1823–1835. 2014.
|
|
135
|
Robles R, Lozano AB, Sevilla A, Márquez L,
Nuez-Ortín W and Moyano FJ: Effect of partially protected butyrate
used as feed additive on growth and intestinal metabolism in sea
bream (Sparus aurata). Fish Physiology and Biochemistry.
39:1567–1580. 2013.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Sun L, Liu Z, Hao G, Zhou L, Lu SQ, Zhang
JS and Xiao TY: Effects of sodium butyrate on growth and intestinal
cell proliferation of Carassius auratus. Zhongguo Shui Chan
Ke Xue. 20:893–901. 2013.
|
|
137
|
Tabrizi JM, Barzeghar A, Farzampour S,
Mirzaii H and Safarmashaei S: Study of the effect of prebiotic
(Saccharomyces cerevisiae) and acidifier on growth
parameters in grower's rainbow trout (Oncorhynchus mykiss).
Ann Biol Res. 3:2053–2057. 2012.
|
|
138
|
Soltan MA, Hassaan MS and Meshrf RN:
Response of Nile tilapia (Oreochromis niloticus) to diet
acidification: Effect on growth performance and feed utilization. J
Appl Aquac. 29:207–219. 2017.
|
|
139
|
Chen YJ, Zhang TY, Luo L, Shi YQ, Bai FJ
and Jiang DN: Impact of dietary l-malic acid supplementation on
growth, feed utilization, ash deposition, and hepatic lipid
metabolism of juvenile genetically improved farmed tilapia,
Oreochromis niloticus. J World Aquac Soc. 48:563–573.
2017.
|
|
140
|
Baruah K, Sahu NP, Pal AK, Debnath D,
Yengkokpam S and Mukherjee SC: Interactions of dietary microbial
phytase, citric acid and crude protein level on mineral utilization
by Rohu, Labeo rohita (Hamilton), juveniles. J World Aquac
Soc. 38:238–249. 2007.
|
|
141
|
Khajepour F, Hosseini SA and Imanpour MR:
Dietary crude protein, citric acid and microbial phytase and their
interacts to influence growth performance, muscle proximate
composition and hematocrite of common carp, Cyprinus carpio
L, juveniles. World J Zool. 7:118–122. 2012.
|
|
142
|
Motlagh HA, Javadmanesh A and Safari O:
Improvement of non-specific immunity, growth, and activity of
digestive enzymes in Carassius auratus as a result of apple
cider vinegar administration to diet. Fish Physiol Biochem.
46:1387–1395. 2020.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Sarker MSA, Satoh S, Kamata K, Haga Y and
Yamamoto Y: Partial replacement of fish meal with plant protein
sources using organic acids to practical diets for juvenile
yellowtail, Seriola quinqueradiata. Aquac Nutr. 18:81–89.
2012.
|
|
144
|
Lim C, Klesius PH and Lückstädt C: Effects
of dietary levels of potassium diformate on growth, feed
utilization and resistance to Streptococcus iniae of Nile
tilapia, Oreochromis niloticus, 2010.
|
|
145
|
Goosen NJ, Görgens JF, De Wet LF and
Chenia H: Organic acids as potential growth promoters in the South
African abalone Haliotis midae. Aquaculture. 321:245–251.
2011.
|
|
146
|
Sarker MSA, Satoh S and Kiron V: Inclusion
of citric acid and/or amino acid-chelated trace elements in
alternate plant protein source diets affects growth and excretion
of nitrogen and phosphorus in red sea bream Pagrus major.
Aquaculture. 262:436–443. 2007.
|
|
147
|
Ramli N, Heindl U and Sunanto S: Effect of
potassium-diformate on growth performance of tilapia challenged
with Vibrio anguillarum. Bali, Indonesia. World Aquac Soc.
248:9–13. 2005.
|
|
148
|
Romana-Eguia MRR, Larona MA and Catacutan
MR: On-farm feed management practices for Nile tilapia
(Oreochromis niloticus) in the Philippines. On-farm feeding
and feed management in aquaculture. FAO Fisheries and Aquaculture,
Rome, Technical Paper. 583:131–158. 2013.
|
|
149
|
da Silva BC, Vieira FDN, Mouriño JLP,
Bolivar N and Seiffert WQ: Butyrate and propionate improve the
growth performance of Litopenaeus vannamei. Aquac Res.
47:612–623. 2016.
|
|
150
|
Ng WK, Koh CB, Teoh CY and Romano N:
Farm-raised tiger shrimp, Penaeus monodon, fed commercial
feeds with added organic acids showed enhanced nutrient
utilization, immune response and resistance to Vibrio
harveyi challenge. Aquaculture. 449:69–77. 2015.
|
|
151
|
Serrano AE Jr, Bunda MGB, Caigoy JCC,
Tumbokon BLM and Jamis JO: Effects of vinegars and sodium acetate
on the growth performance of pacific white shrimp, Penaeus
vannamei. Isr J Aquac. 70(8)2018.
|
|
152
|
Sarker SA, Satoh S and Kiron V:
Supplementation of citric acid and amino acid-chelated trace
element to develop environment-friendly feed for red sea bream,
Pagrus major. Aquaculture. 248:3–11. 2005.
|