Introduction
Traumatic brain injury (TBI), occurs due to primary
damage to the brain induced by mechanical forces on the head and
brain through various mechanisms. Secondary injury due to TBI
involves a variety of complex biochemical cascades, such as
oxidative stress, glutamate excitotoxicity and neuroinflammation,
leading to neuronal cell death (1). Mitochondrial dysfunction has been
reported to be a key pathway for neuroinflammation, resulting in
the accumulation of reactive oxygen species (ROS) (2). Another manifestation of TBI is an
impairment in fine motor control and coordination, and difficulty
with balance. TBI often leads to disability, with motor weakness as
its main symptom (3).
Brain-derived neurotrophic factor (BDNF) comprises a
family of neurotrophic proteins secreted by autocrine factors,
which are essential for the survival, differentiation and
regeneration of neuronal cells. BDNF has been reported to reduce
secondary brain injury through neuroprotection mechanisms and to
restore connectivity (4). Previous
studies have reported clinical improvement with BDNF-related
treatment in TBI. Studies increasing BDNF expression using protein
kinase-like endoplasmic reticulum kinase antagonist (5), the direct use of collagen-domain bond
BDNF (6) and the activation of
BDNF receptor using 7,8-dihydroxyflavone have demonstrated the
promotion of neuron growth and an improvement in overall
neurological function (7,8).
There are three types of superoxide dismutase (SOD)
in humans; SOD1 is present in various parts of mammalian cells,
while SOD2 is localized to the mitochondria and efficiently
eliminates superoxide, and has been reported to be induced in
several inflammatory conditions in the central nervous system
(9). Neurological and clinical
improvements have been reported with SOD-related treatment in
previous studies. Previous studies treating animals with TBI with
SOD mimetic (10), exogenous
lecithinized SOD (11) and the
transgenic increment of SOD activity have revealed an improvement
in hemodynamic recovery, more viable neurons, and an improvement in
neurological function following TBI (12).
c-Jun N-terminal kinase (JNK)3 has been found to
play a key role in the etiology of brain damage. JNK3 is activated
in the brain in response to TBI, stroke and ischemia, resulting in
the promotion of apoptosis, inflammation and oxidative stress. JNK3
activation can induce the production of pro-inflammatory cytokines
and chemokines, which can aggravate brain damage by boosting immune
cell recruitment and the activation of microglia. JNK3 can also
trigger the generation of ROS, which can further harm neurons and
other brain cells (13).
Citrus sinensis (C. sinensis) has
various compounds, such as flavonoids, that have been identified in
citrus. Flavonoids have a variety of biological benefits, such as
antioxidant and anti-inflammatory effects (14). A previous study by the authors
demonstrated the potential antioxidant effects of C.
sinensis extracts using molecular docking analyses and dynamic
simulations (15). However, the
neurotrophic, antioxidant and anti-apoptotic potential of C.
sinensis extract has not been investigated to date, at least to
the best of our knowledge. The present study thus aimed to
investigate the neurotrophic, antioxidant and anti-apoptotic
effects of C. sinensis peel extract (CSPE) using in
vivo and in silico analyses.
Materials and methods
Phytochemical extraction
CSPE was used according to a previous study by the
authors (15). The metabolite
compound of CSPE was published at MetaboLights (https://www.ebi.ac.uk/metabolights/) with the
identifier no. MTBLS5785.
Molecular docking analysis
Molecular docking analysis was performed using Pyrx
9.5 (https://pyrx.sourceforge.io/) as
previously described (15). In the
present study, JNK3 (PDB ID: 7KSK) was targeted using several
compounds, such as linoleic acid (CID: 5280450), tangeretin (CID:
68077), nootkatone (CID: 1268142), alminoprofen (CID: 2097),
chanoclavine (CID: 5281381) and scoparone (CID: 8417) that have
been previously pharmacokinetically analyzed (15). THE 2D visualization of compounds
and protein target interaction was performed using Discovery Studio
(https://discover.3ds.com/discovery-studio-visualizer-download).
The strongest interaction was analyzed using molecular dynamics
(16,17).
Molecular dynamic analysis
We performed molecular dynamic simulations of the
complex interaction of chanoclavine and nootkatone with JNK using
the OpenMM engine and the AMBER force field for protein and ligand
systems in the Google Collaboratory (https://colab.research.google.com/?utm_source=scs-index).
The model was thoroughly solvated with TIP3P water models108 in a
12x12x12 water box and neutralized by the addition of NaCl at a
concentration of 150 mM. The simulation was performed using the
force field ff19SB. The system was minimized for 1,000 steps and
equilibrated in the isothermal-isobaric (NPT) ensemble for 20 nsec
with a timestep of 2 fsec to achieve temperatures and pressures in
the 310 K and 1 atm ranges, respectively. The temperature was
controlled using the modified Berendsen thermostat with a coupling
time of 0.1 psec, while the pressure was controlled by
Parrinello-Rahman with a reference pressure of 1.0 bar and a
compressibility of 4.5Xe-5 bar -1. The periodic electrostatic
interactions were computed using particle mesh Ewald (PME)
summation with a grid spacing <1 Å. The protein was
unconstrained during the MD simulation (18). In the present study, the Radius
gyration (Rg), root mean square distance (RMSD) and root mean
square fluctuation (RMSF) were evaluated.
Experimental and animal design
A total of 30 Rattus norvegicus rats aged
8-12 weeks, weighing 300-450 g, were sourced from the Animal
Research Laboratory, Medical Faculty, Brawijaya University, Malang,
Indonesia. The weight and age data of the rats are presented in
Table SI. The animals were housed
in a controlled environment and acclimatized at room temperature
(21-25˚C) and humidity (45-50%) with 12-h light/dark
cycle for 2 weeks (19). The rats
were monitored once daily and were provided with standard food and
water ad libitum. The rats were randomly divided into 5
groups using simple randomization, as shown in Table I. The minimum sample size was 20
rats in total (at least 4 rats in each group), calculated using the
Federer formula. CSPE was orally administered for 7 days following
the induction of TBI. The animals were subjected to wire grip and
Barnes maze tests on the 1st, 3rd and 7th days following the
induction of TBI. Following treatment, the rats were sacrificed
using ketamine (40-100 mg/kg) and xylazine (5-13 mg/kg) as
anesthetics, followed by decapitation. Various criteria, such as
self-mutilation, amputation or crush injury of the limbs and tail,
generalized dermatitis, sepsis and severe dehydration, were also
used to determine the indication of euthanasia. The death of the
animals was verified by the absence of respiration, the absence of
a heartbeat, pupillary dilation, the absence of reflexes, the poor
color of extremities and mucous membranes, and dry and opaque
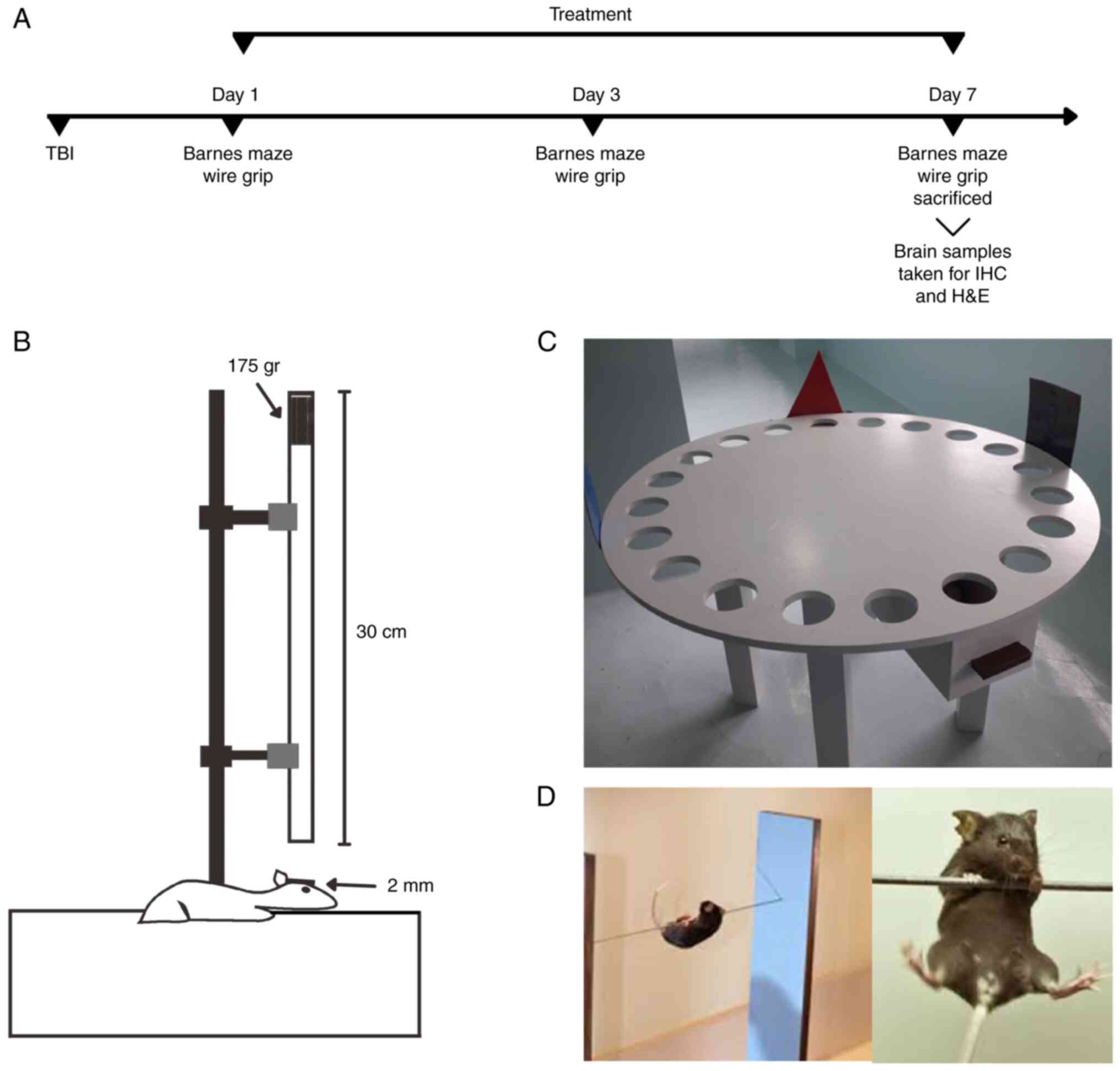
corneas. The study experimental design is illustrated in Fig. 1A. Brain samples were stored and
placed in a buffered formalin solution for fixation. Further
examinations were performed using histological analysis and
immunohistochemistry (IHC). The present study was conducted under
international ethical principles and the National Institutes of
Health's Guide for the Care and Use of Laboratory Animals of
Medical Faculty, Brawijaya University.
 | Table IExperimental animal groups in the
present study. |
Table I
Experimental animal groups in the
present study.
| Group | Treatment |
|---|
| Sham | Normal diet +
normal saline for 7 days |
| TBI | TBI induction +
normal diet + normal saline for 7 days |
| Treatment group
1 | TBI induction +
normal diet for 7 days + CSPE administered orally at a dose of 50
mg/kg body weight for 7 days |
| Treatment group
2 | TBI induction +
normal diet for 7 days + CSPE administered orally at a dose of 250
mg/kg body weight for 7 days |
| Treatment group
3 | TBI induction +
normal diet for 7 days + CSPE administered orally at a dose of 500
mg/kg body weight for 7 days |
Induction of TBI
TBI was induced according to a previously developed
weight drop model to produce mild brain injury (16). Briefly, the rats were anesthetized
using ketamine (40-100 mg/kg) and xylazine (5-13 mg/kg). The scalp
was cleaned with 10% povidone-iodine, and 10% lidocaine HCl was
then injected as a local anesthetic. The scalp was incised
longitudinally at the midline of the skull with a diameter of 5 mm
to expose the skull underneath, between the lambda and bregma
sutures. A steel plate (2-mm-thick) was fixed on the exposed skull.
Subsequently, 175 g of weight was dropped onto the steel plate
through a 30-cm-long cylinder, and the scalp of the rats was
stitched back, as shown in Fig.
1B, and as previously described (20,21).
The tats in the sham-operated (sham) group only underwent scalp
incision and stitching.
Spatial memory and motor test
Barnes maze test, adapted from Krishna et al
(7), was performed to analyze the
rat's spatial memory, as illustrated in Fig. 1C. The diameter of the circular
platform was 160 cm, with 20 circular holes equally sawed 5.5 cm
from the platform's edge. The holes were each 8 cm in diameter and
were equally distanced at 16 cm from each other. The escape box was
made from plexiglass (30x15x15 cm) and positioned under the
selected escape hole (7).
The wire grip test was performed as illustrated in
Fig. 1D. A wire grip apparatus
with a large plastic box (55x40x35 cm) and a 2-mm-thick wire
attached to two vertical stands. Bedding material with a distance
of 35 cm below the wire was used to prevent injury to the animal
(22). The investigator allowed
the rat to hang from the wire using only its forelimbs. As soon as
the rat held the wire properly, the timer began. The time taken for
the rat to keep its limbs on the wire was recorded. If the rat
fell, the timer stopped.
Surviving neuron cell count using
hematoxylin and eosin staining (H&E)
The rat brains were fixed in 4% paraformaldehyde
(Path Chem, BBC Biochemical) for 24 h at 24˚C. The fixed brain
tissues were embedded in paraffin and sliced coronally. The sliced
brain tissues were then stained with hematoxylin for 20 min and
eosin for 3 sec at 24˚C (Leica Biosystems Inc.). Stained slides
were analyzed under a light microscope at x400 magnification (BX51,
Olympus Corp.). The numbers of surviving neurons per high power
field (hpf) were counted. Non-surviving neurons were identified by
their shrunken and thickened nuclei (23).
IHC
The brain tissues were deparaffinized and stored for
24 s at room temperature prior to use. The brain tissues were then
washed using phosphate-buffered saline (PBS) and then incubated
with normal rabbit serum at room temperature. The incubated brain
tissues were washed with PBS and subsequently incubated at room
temperature for 30 min with a secondary antibody (AMF080, ScyTek
Laboratories, Inc.), washed with PBS, and developed with 0.05%
diaminobenzidine tetrahydrochloride. The primary antibodies used
for IHC were anti-BDNF (cat. no. sc-65514,), anti-SOD1 (cat. no.
sc-101523), and anti-SOD2 antibody (cat. no. sc-137254) (1:50; all
from Santa Cruz Biotechnology, Inc.). The expression levels of
BDNF, SOD1, and SOD2 were observed under a light microscope at
x1,000 magnification in 20 fields of view (BX51, Olympus Corp.), as
previously described (24).
Statistical analyses
Statistical analyses were performed using SPSS 26.0
software (IBM Corp.). All results are presented as the mean ± SD.
The normality of the data was assessed using the Shapiro-Wilk test.
The analyses of BDNF, SOD1, SOD2, Barnes maze latency and
wire-closure time data were performed using one-way ANOVA with the
Bonferroni post-hoc test.
Results
Molecular docking analysis of CSPE
against JNK
Previously, the authors analyzed various compounds
in CSPE that can penetrate the brain blood-barrier (BBB) (15). Herein, the binding affinity of the
compounds with JNK protein was analyzed. It was found that
chanoclavine and nootkatone had a stronger binding affinity
compared to
1-(trans-4-{[7-oxo-8-(propan-2-yl)-7,8-dihydropyrido[2,3-d]pyrimidin-2-yl]amino}cyclohexyl)-3-propan-2-ylurea
as a control, as shown in Table
II. Molecular docking analysis identified that chanoclavine
bound to JNK3 with the highest affinity compared to the control
ligand and other ligands. Chanoclavine exhibited a binding affinity
of -6.8 Kcal/mol, whereas nootkatone had a binding affinity of -6.6
Kcal/mol. The molecule with the lowest bond energy will have a
constant temperature and pressure, and this is known as a stable
molecule (19,23,24).
The amino acid residues affect the binding domain of the target
protein, as well as the sort of chemical interplays in the binding
domain.
| Table IIBinding affinity interaction of CSPE
compounds and c-Jun N-terminal kinase. |
Table II
Binding affinity interaction of CSPE
compounds and c-Jun N-terminal kinase.
| Compound | Target protein (PDB
ID) | Binding affinity
energy (kcal/mol) | Interacting
residues | Type of bond | Grid box |
|---|
| Chanoclavine | JNK (4Y5H) | -6.8a | Ile70 | Hydrophobic | Coordinates |
| | | | Val78 | Hydrophobic | X: 1.161 |
| | | | Ala91 | Hydrophobic | Y: -29.628 |
| | | | Met146 | Hydrophobic | Z: -30.545 |
| | | | Val196 | Hydrophobic | |
| | | | Leu206 | Hydrophobic | Radius |
| | | | | | X:10 |
| | | | | | Y: 10 |
| | | | | | Z: 10 |
| Nootkatone | | -6.6a | Ile70 | Hydrophobic | |
| | | | Val78 | Hydrophobic | |
| | | | Ala91 | Hydrophobic | |
| | | | Met146 | Hydrophobic | |
| | | | Met149 | Hydrophobic | |
| | | | Asn152 | Hydrogen | |
| | | | Val196 | Hydrophobic | |
| | | | Leu206 | Hydrophobic | |
| Tangeretin | | -6.3 | Ile70 | Hydrophobic | |
| | | | Val78 | Hydrophobic | |
| | | | Ala91 | Hydrophobic | |
| | | | Lys93 | Hydrophobic | |
| | | | Met146 | Hydrophobic | |
| | | | Asn152 | Hydrogen | |
| | | | Val196 | Hydrophobic | |
| Alminoprofen | | -5.5 | Ile70 | Hydrophobic | |
| | | | Ala80 | Hydrophobic | |
| | | | Leu148 | Hydrophobic | |
| | | | Met149 | Hydrogen | |
| | | | Asn152 | Hydrogen | |
| | | | Val196 | Hydrophobic | |
| | | | Leu206 | Hydrophobic | |
| Scoparone | | -5.8 | Val78 | Hydrophobic | |
| | | | Ala91 | Hydrophobic | |
| | | | Gln155 | Hydrogen | |
| | | | Val196 | Hydrophobic | |
| | | | Leu206 | Hydrophobic | |
| Linoleic acid | | -5.5 | Ile70 | Hydrophobic | |
| | | | Val78 | Hydrophobic | |
| | | | Ala91 | Hydrophobic | |
| | | | Leu148 | Hydrophobic | |
| | | | Met149 | Hydrophobic | |
| | | | Asp150 | Hydrogen | |
| | | | Gln155 | Hydrogen | |
| | | | Val196 | Hydrophobic | |
| | | | Leu206 | Hydrophobic | |
|
1-(trans-4-{[7-oxo-8- | | -6.5 | Ile70 | Hydrophobic | |
|
(propan-2-yl)-7, | | | Val78 | Hydrophobic | |
|
8-dihydropyrido[2,3-d] | | | Ala91 | Hydrophobic | |
|
pyrimidin-2-yl]amino} | | | Met146 | Hydrogen | |
|
cyclohexyl)-3-propan- | | | Glu147 | Hydrogen | |
| 2-ylurea
(control) | | | Met149 | Hydrophobic | |
| | | | GLn155 | Hydrogen | |
| | | | Val196 | Hydrophobic | |
| | | | Leu206 | Hydrophobic | |
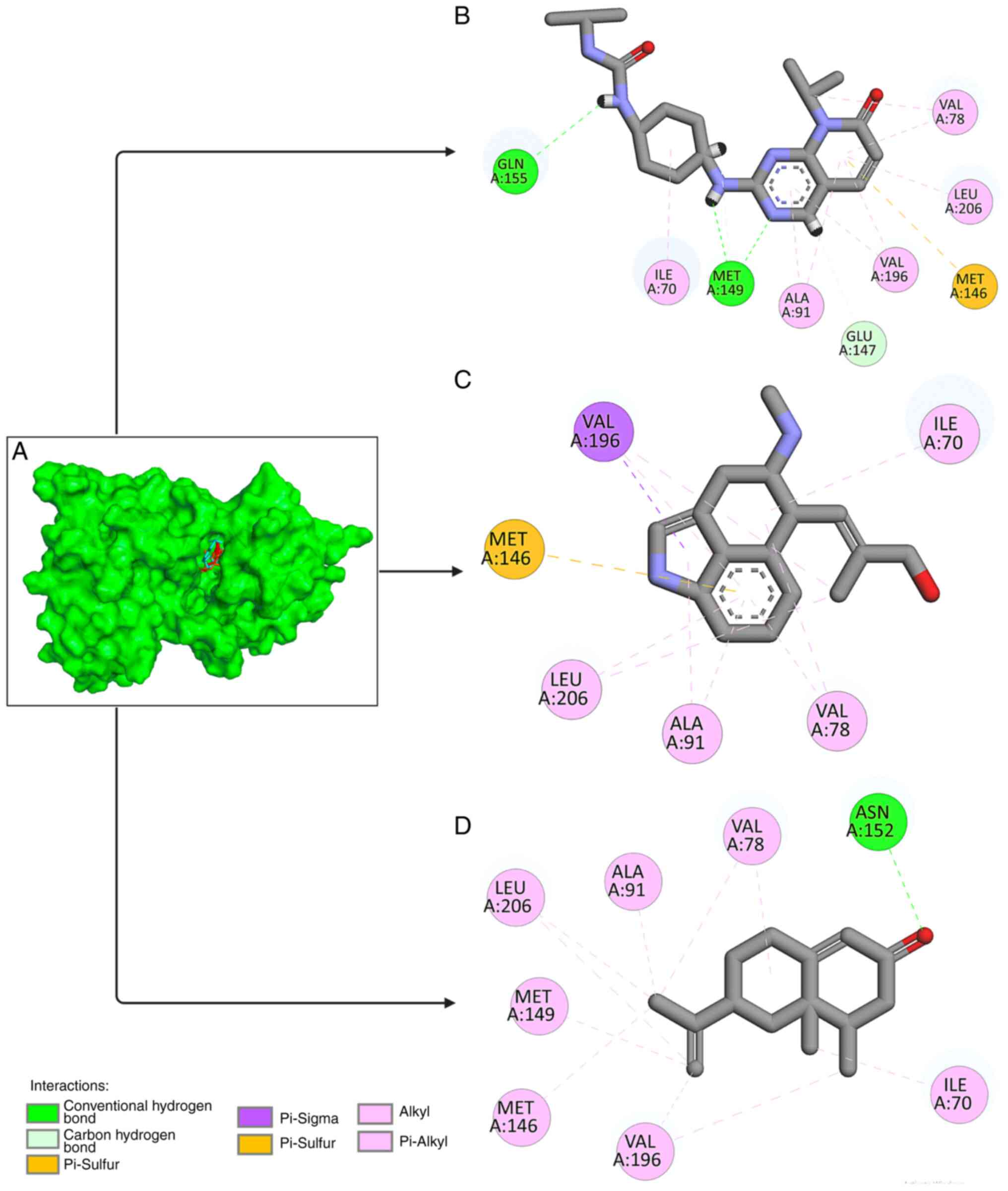
The different types of bonds found between
chanoclavine, nootkatone and the target protein were analyzed. The
type of interaction between the ligand and protein target is a
critical parameter in predicting the effect of interactions
(25,26). Chanoclavine attaches to JNK via six
hydrophobic bond interactions, (ILE70, VAL78, ALA91, MET146, VAL196
and LEU206). Furthermore, nootkatone interacts with eight amino
acid residues. Nootkatone created hydrogen bonds with ASN152 and
hydrophobic bonds with the amino acids ILE70, ALA80, LEU148,
MET149, VAL196, and LEU206, as shown in Fig. 2. The present study investigated the
interaction with the control ligand and found similar amino acid
residue interaction, as shown in Table II.
Molecular dynamic simulations analysis
of CSPE against JNK
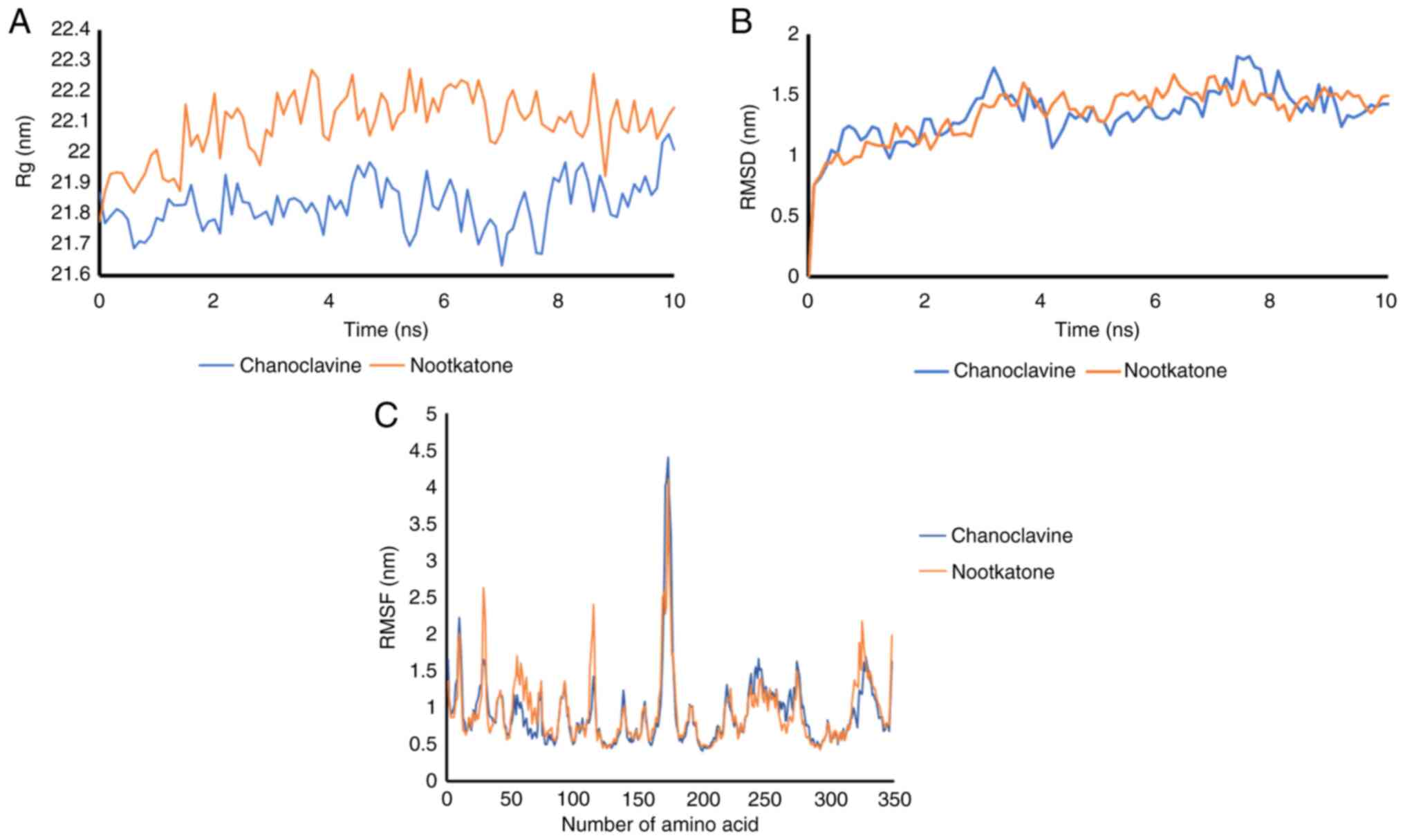
The Rg (radius of gyration), which is defined as the
root mean square distance of the collection of atoms from their
shared center of gravity, defines the general dispersion of the
molecule. The radius of gyration values for protein-ligand
complexes, illustrated in Fig. 3A,
reveal that while chanoclavine was found to affect the secondary
structure of the proteins, the radius of gyration of JNK remained
constant following contact. The nootkatone Rg value was higher than
that of chanoclavine, indicating that chanoclavine is more compact
than nootkatone. A lower radius of gyration indicates that the
polymer is relatively compact, implying that the polymer spends the
majority of its time folded along its course.
The JNK protein complex with chanoclavine had
average RMSD fluctuations of ~1.2 nm, with an equilibrium of 6
nsec. The RMSD of he nootkatone and JNK complex had an unstable
fluctuation from 0 to 10 nsec. Similarly, substantial RMSD
deviations were reported for the nootkatone-Kelch like ECH
associated protein 1 (Keap1) complex, indicating that the complex
generated was unstable, as shown in Fig. 3B.
Subsequently, the RMSF was investigated. RMSF was
monitored at 10-nsec intervals to calculate residual flexibility.
The complex interaction combination had a wavelength ~0.75 nm for
each residue, whereas it had the most unstable residue at residue
number 170-177 with a RMSF value of >2.5 nsec, as shown in
Fig. 3C.
Effect of CSPE on spatial memory and
motor performance
A total of 30 rats were used in the present study,
and no deaths occurred during the study period. The results
revealed a significant difference in wire grip duration within
group 3 (Table III). The
difference was observed on the 3rd day following TBI compared to
the 7th day following TBI. The mean latency in all groups treated
with CSPE improved progressively from the 1st to the 7th day
following the induction of TBI.
| Table IIIEffects of traumatic brain injury and
treatment with Citrus sinensis peel extract on wire grip
duration and Barnes maze latency. |
Table III
Effects of traumatic brain injury and
treatment with Citrus sinensis peel extract on wire grip
duration and Barnes maze latency.
| | Groups (mean ±
SD) |
|---|
| Parameter | Sham | TBI | Group 1 | Group 2 | Group 3 |
|---|
| Wire grip duration
(sec) | | | | | |
|
D1 | 22.83±11.64 | 5.33±2.50 | 5.83±2.32 | 7.67±4.46 | 5.17±2.79 |
|
D3 | 26.50±12.82 | 12.00±4.00 | 16.67±8.82 | 13.67±4.97 | 14.33±6.65 |
|
D7 | 28.33±11.91 | 14.67±6.22 | 21.17±3.97 | 26.00±11.82 |
27.83±10.72a |
| Barnes maze latency
(sec) | | | | | |
|
D1 | 47.67±43.50 | 172.50±8.39 | 173.17±8.95 | 173.33±5.24 | 170.33±8.17 |
|
D3 | 42.67±38.90 |
114.00±38.58a |
87.50±46.45a |
83.00±27.49a |
88.67±39.05a |
|
D7 | 40.67±37.30 | 73.50±54.17 |
49.00±65.07a |
37.17±24.91a |
33.17±23.24a |
A downward pattern in the mean Barnes maze latency
values was observed in all groups, gradually decreasing from the
1st to the 7th day following TBI. In the sham group, there was no
significant difference in the Barnes maze latency between each test
within the group. Significant differences were found in all groups
treated with CSPE, with significantly lower Barnes maze latency
values on the 3rd day compared to the 1st day following TBI, and on
the 7th day compared to the 3rd day following TBI. The results of
the Barnes maze test are presented in Table III.
Surviving neuron cell count
Neuron survival was assessed by cell counting using
H&E-stained brain samples. Non-surviving neurons were
identified by shrunken and thickened nuclei. The number of
surviving neurons in groups 2 and 3 was significantly higher than
those in the TBI group (P<0.05; Table IV). Staining images of the
surviving neurons are presented in Fig. S1.
| Table IVEffects of traumatic brain injury and
treatment with Citrus sinensis peel extract on surviving
neurons, and on SOD1, SOD2 and BDNF expression. |
Table IV
Effects of traumatic brain injury and
treatment with Citrus sinensis peel extract on surviving
neurons, and on SOD1, SOD2 and BDNF expression.
| | Groups (mean ±
SD) | |
|---|
| Parameters | Sham | TBI | Group 1 | Group 2 | Group 3 | P-value |
|---|
| Surviving
neurons | 42.50±20.95 | 17.67±11.09 | 25.00±13.84 |
30.67±5.85b |
41.00±14.04b | 0.024 |
| SOD1 | 49.67±15.31 | 60.67±11.20 | 64.33±5.39 |
72.33±7.92a |
82.00±17.55a,b | 0.002 |
| SOD2 | 56.17±13.18 | 60.67±9.67 |
76.33±11.22a,b |
77.50±11.08a,b |
93.17±19.95a,b | 0.001 |
| BDNF | 53.00±13.33 | 68.83±9.75 |
75.33±8.50a |
86.67±13.10a,b |
113.83±11.27a,b | <0.001 |
BDNF and SOD expression
The results of BDNF and SOD expression analyses are
shown in Table IV. An increasing
pattern in BDNF expression was observed with the increasing CSPE
dosage. The results revealed a significantly higher BDNF expression
in groups 2 and 3 compared to the TBI group (P<0.05). There was
an increasing pattern of SOD1 and SOD2 expression with increasing
CSPE dosage. The results revealed a significantly higher SOD1
expression in group 3 compared to the TBI group, and a
significantly higher SOD2 expression in all groups treated with
CSPE compared to the TBI group (P<0.05). Staining images of
BDNF, SOD1 and SOD2 expression are presented in Figs. S2, S3 and S4.
Discussion
The present study demonstrated the potential of CSPE
to attenuate various neurological injuries following TBI. A
significant increase in BDNF expression was found following
treatment with CSPE. This is consistent with the findings of
previous studies, as indicated below. Several compounds contained
in CSPE have been examined for their effects in inducing
neurotrophins, such as BDNF. Auraptene, a compound found in citrus,
has been proven to activate the cyclic adenosine monophosphate
(cAMP) response element-binding protein (CREB) pathway, resulting
in the increased production of various neurotrophic factors,
including BDNF (27). A
significant increase in BDNF mRNA levels has been found in mouse
neuroblastoma neuro2a cells (27).
Sawamoto et al revealed a promoting effect of the
heptamethoxyflavone (HMF) active compound in citrus on BDNF via the
cAMP/ERK/CREB pathway. In their study, treatment of C6 glioma cells
with 10 µM HMF resulted in an increase in cAMP levels, CREB
phosphorylation and BDNF expression (28). Another citrus flavonoid,
hesperidin, was previously found to stimulate BDNF (29). Hesperidin was used to treat
zebrafish in a pentylenetetrazole-induced convulsion paradigm,
which significantly increased seizure latency and decreased
hyperactive responses. Hesperidin was found to have a good
N-methyl-D-aspartate (NMDA) receptor binding affinity,
according to an in silico investigation. NMDA receptor
activation stimulated the CREB/BDNF pathway (29).
In the present study, a significant increase in SOD1
and SOD2 expression was found following treatment with CSPE.
Previous studies have also shown the antioxidant effects of CSPE
via increments in the levels of antioxidant enzymes. For example, a
significantly higher SOD level was previously reported in rats with
alcohol-induced peptic ulcers pre-treated with 200 and 400 mg/kg
CSPE (30). The levels of other
antioxidant enzymes, such as glutathione peroxidase and catalase
were significantly increased in the rats pre-treated with similar
dosages. A significant decrease in the levels of oxidative stress,
indicated by a decrease in the levels of malondialdehyde and
hydrogen peroxide was also observed in rats pre-treated with 100,
200 and 400 mg/kg CSPE (30). An
increase in the levels of antioxidant enzymes has also been found
in rats treated with CSPE in their diet. Erukainure et al
(31) examined rats fed a diet
consisting of 35% CSPE for 6 weeks. A significant increase in the
levels of SOD and glutathione was observed following 6 weeks of
treatment (31). In another study,
in Wistar rats treated orally with a combination of Citrus
aurantifolia and Cinnamomum burmannii at 100, 300 and
500 mg/kg, exhibited an increase in the levels of SOD following
treatment (32).
In addition to inducing antioxidative enzymes,
previous studies have shown that CSPE exerts antioxidant effects
via various other mechanisms, such as oxidative enzyme inhibition,
radical scavenging and metal chelating activity. The study by
Malterud and Rydland (33)
demonstrated an inhibitory effect on lipoxygenase (LOX) by various
flavones of CSPE. In their study, various flavones that inhibited
LOX could penetrate the BBB, such as sinensetin and tangeretin
(33). Another study by Pepe et
al (34) demonstrated that
CSPE exerted an inhibitory effect on nitric oxide synthase.
Several compounds, such as vitamin E and carotenoid
contained in CSPE are also known as singlet oxygen
(1O2) scavengers (35). CSPE has the most radical scavenging
ability compared to other citrus species, such as Citrus
aurantifolia and Citrus limonum (36). In addition to its antioxidant
effects, neuronal growth promoted by neurotrophic agents has been
proven to be beneficial in TBI. Various studies have shown that
treatment with SOD and its related pathways in TBI can lead to
better neurological and clinical recovery, demonstrating the
importance of oxidative stress control in TBI (10-12).
In addition, previous studies using BDNF treatment or treatment
with its related pathways have demonstrated that this treatment
leads to an improvement in neurological recovery and better
clinical function, highlighting the importance of neurological
recovery accelerated by neurotrophic agents (5,7,8).
In the present study, the significant increase in
the numbers of surviving neurons in rats in treatment groups 2 and
3 may be attributed to increased levels of antioxidants and
neurotrophic expression. Significantly higher levels of antioxidant
enzymes prevent further cellular damage from oxidative stress,
while increased levels if neurotrophins lead to more rapid neuron
repair and growth. However, a previous study by Chen et al
(37) provided insight into
another pathway. Their study demonstrated that the treatment of
mice with optic nerve injury with various citrus flavones, such as
naringenin, nobiletin and hesperidin led to increased retinal
ganglion survival (37). In
addition, in the same study, in vitro experiments using 293T
cells, in which the JNK-JUN apoptotic pathway was activated by
incubation with 500 mM sorbitol, revealed that treatment with 2.5
µM of naringenin resulted in a significant decrease in JUN
phosphorylation and its subsequent apoptotic pathway (37).
In the present study, subsequent molecular docking
analysis confirmed the potential effects of CSPE on the JNK
pathway, with nookatone and chanoclavine exhibiting a strong
binding energy, and similar amino acid interactions compared to the
control. In its various isoforms, JNK is crucial for brain
development, in regulating various functions such as cell death in
early brain development, and neuronal migration and axon
maintenance in later brain development. The most critical function
of JNK in relation to TBI is its function in mediating
stress-induced neuronal cell death in the adult brain. A previous
study revealed that JNK knockout (KO) mice exhibited resistance to
neuronal cell death induced by a neurotoxin,
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (38). Other studies have also indicated
that JNK KO mice are resistant to ischemia-induced neuronal cell
death, and excitotoxicity-induced neuronal cell death by kainic
acid (39,40). The JNK-mediated apoptotic pathway
involves the nuclear translocation of JNK, activating various
transcription factors such as c-Jun, resulting in the activation of
various pro-apoptotic genes, and in the subsequent release of
apoptotic proteins (41).
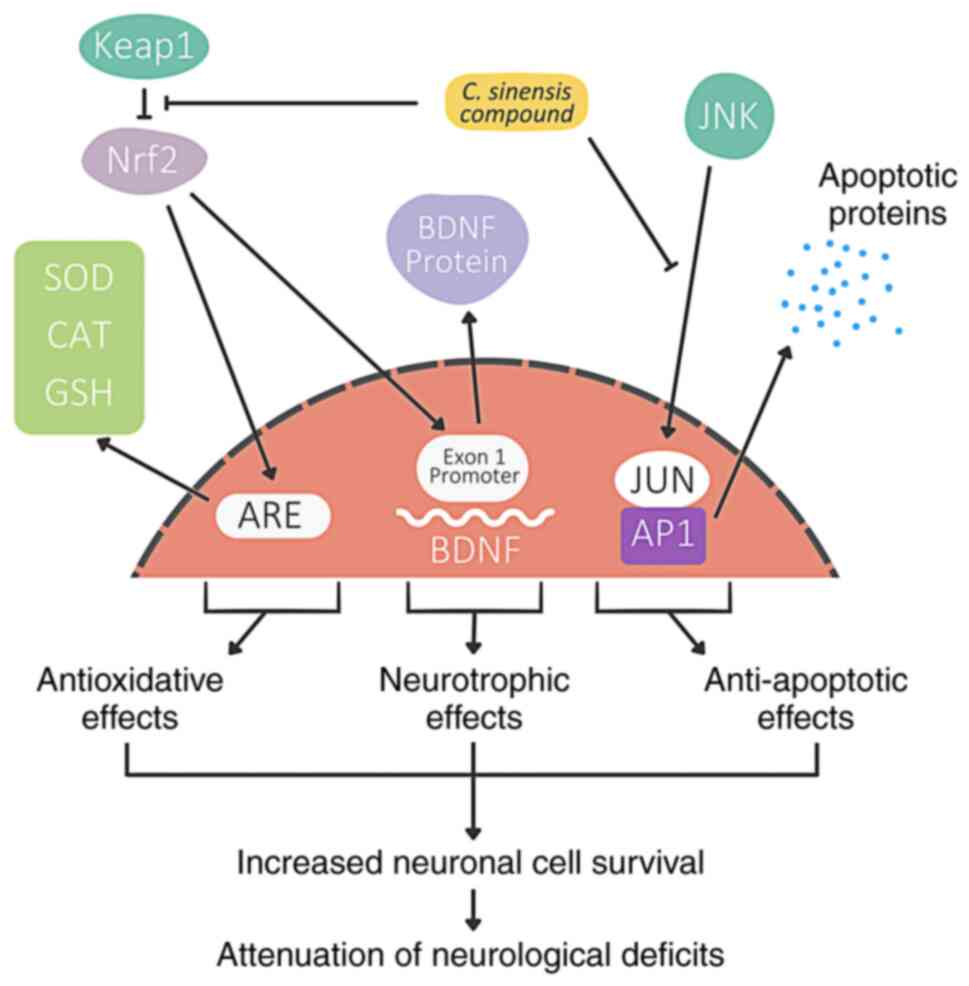
A previous study by the authors found that CSPE has
potential to inhibit Keap1 activity (15). In the present study, the pathways
suggested to be involved in the various mechanisms of the effects
of CSPE on TBI are illustrated in Fig.
4. CSPE exerts antioxidant effects by inducing an increase in
the levels of antioxidant enzymes, radical scavenging and metal
chelating activity. CSPE also exerts neurotrophic activity via the
activation of exon 1 promoter in the BDNF gene and CREB-BDNF
pathway. Its antioxidant activity, combined with neurotrophic
activity, and potential anti-apoptotic activity, lead to an
increased neuronal cell survival and the consequent attenuation in
motor and visuospatial memory deficits, as revealed herein.
In the presents study, motor and cognitive aspects
assessed using wire grip and Barnes maze tests revealed clinical
improvements following CSPE treatment. This finding is in
accordance with the findings of previous studies. For example, a
previous study by Liu et al (42) demonstrated a significant
improvement in a grip strength test in model rats with spinal cord
injury treated with trihydroxyethyl rutin. Another study by Zhu
et al (43) revealed a
significant improvement in Morris water maze escape latency in rats
with TBI treated with docosahexaenoic acid (43). These findings prove the beneficial
effects of neuroprotective treatment on clinical motor and
cognitive improvements.
In the present study, various effects of CSPE on TBI
were demonstrated by analyzing related components, such as SOD1,
SOD2 and BDNF, which revealed the antioxidant and neurotrophic
effects of CSPE. Molecular docking analysis and dynamic simulation
also revealed the anti-apoptotic activity of the extract. While the
present study delved into the antioxidant and neurotrophic effects
of CSPE by analyzing components such as SOD1, SOD2 and BDNF, it
should be noted that it focused on a specific subset of pathways.
There are numerous other pathways involved in antioxidant,
neurotrophic and anti-apoptotic responses. The limited focus of the
present study on these particular components suggests that other
significant factors and interactions that may also have an impact
on the effects of CSPE in TBI may have been overlooked, which
limits the findings presented herein. The analysis of other
component pathways needs to be conducted in the future in order to
better understand the additional pathways involved in the effects
of CSPE on TBI. Toxicity analysis should also be conducted in
future studies in order to elucidate the aspects of the toxicity of
different doses of CSPE.
In conclusion, the present study demonstrated the
potential antioxidant effects of CSPE on TBI by significantly
increasing the levels of SOD1 and SOD2, enhancing neurotrophic
activities by significantly increasing BDNF, and attenuating
overall neurological deficits, as evidenced by improvements in the
wire grip and Barnes maze tests. Chanoclavine and nootkatone can
also inhibit JNK to induce apoptosis, as revealed using molecular
docking and dynamic simulation analyses. Future research however,
is warranted to evaluate the efficacy and safety of CSPE in
improving motor and cognitive functions in mouse models of TBI.
Supplementary Material
Staining images of surviving neurons
in each group. (A) Sham group, (B) traumatic brain injury group,
(C) treatment group 1, (D) treatment group 2, (E) treatment group
3.
Staining images of brain-derived
neurotrophic factor in each group. (A) Sham group, (B) traumatic
brain injury group, (C) treatment group 1, (D) treatment group 2,
(E) treatment group 3.
Staining images of superoxide
dismutase 1 in each group. (A) Sham group, (B) traumatic brain
injury group, (C) treatment group 1, (D) treatment group 2, (E)
treatment group 3.
Staining images of superoxide
dismutase 2 in each group. (A) Sham group, (B) traumatic brain
injury group, (C) treatment group 1, (D) treatment group 2, (E)
treatment group 3.
Weight and age of the experimental
animals in the present study.
Acknowledgements
Not applicable.
Funding
Funding: The present study was financially supported by the
Faculty of Medicine Brawijaya University, Malang, Indonesia for its
financial support (grant no. 34/SK/ UN10.F08.06/ KS/ 2019).
Availability of data and materials
The data that support the findings of this study are
available on the MetaboLights Compound Database (https://www.ebi.ac.uk/metabolights/MTBLS5785).
Authors' contributions
WMS, HS, and GFAP were involved in the conception
and design of the study, data collection and analysis, and in the
writing, revising and reviewing of the manuscript. MFRS, RAV, JPK
and ISM were involved in the conception and design of the study,
and in the revising and reviewing of the manuscript. WMS and HS
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was conducted by following the
University of Brawijaya animal ethics guidelines and was approved
by the Ethics Committee (Approval no. 84/EC/KEPK/04/2020), Faculty
of Medicine, Brawijaya University, Malang, Indonesia.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Donkin JJ and Vink R: Mechanisms of
cerebral edema in traumatic brain injury: Therapeutic developments.
Curr Opin Neurol. 23:293–299. 2010.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Angeloni C, Prata C, Sega FV, Piperno R
and Hrelia S: Traumatic brain injury and NADPH oxidase: A deep
relationship. Oxid Med Cell Longev. 2015(370312)2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Stephens J, Salorio C, Denckla M,
Mostofsky S and Suskauer S: Subtle motor findings during recovery
from pediatric traumatic brain injury: A preliminary report. J Mot
Behav. 49:20–26. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Korley FK, Diaz-Arrastia R, Wu AHB, Yue
JK, Manley GT, Sair HI, Eyk JV and Everett AD: TRACK-TBI
investigators. Okonkwo DO, et al: Circulating brain-derived
neurotrophic factor has diagnostic and prognostic value in
traumatic brain injury. J Neurotrauma. 33:215–225. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Sen T, Gupta R, Kaiser H and Sen N:
Activation of PERK elicits memory impairment through inactivation
of CREB and downregulation of PSD95 after traumatic brain injury. J
Neurosci. 37:5900–5911. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yin R, Zhao S and Qiu C: Brain-derived
neurotrophic factor fused with a collagen-binding domain inhibits
neuroinflammation and promotes neurological recovery of traumatic
brain injury mice via TrkB signalling. J Pharm Pharmacol.
72:539–550. 2022.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Krishna G, Agrawal R, Zhuang Y, Ying Z,
Paydar A, Harris NG, Royes LFF and Gomez-Pinilla F:
7,8-dihydroxyflavone facilitates the action exercise to restore
plasticity and functionality: Implications for early brain trauma
recovery. Biochim Biophys Acta. 1863:1204–1213. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Gustafsson D, Klang A, Thams S and Rostami
E: The role of BDNF in experimental and clinical traumatic brain
injury. Int J Mol Sci. 22(3582)2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ishihara Y, Takemoto T, Itoh K, Ishida A
and Yamazaki T: Dual role of superoxide dismutase 2 induced in
activated microglia. J Biol Chem. 290:22805–22817. 2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Seno S, Wang J, Cao S, Saraswati M, Park
S, Simoni J, Ma L, Soltys B, Hsia CJC, Koehler RC, et al:
Resuscitation with macromolecular superoxide dismutase/catalase
mimetic polynitroxylated PEGylated hemoglobin offers
neuroprotection in guinea pigs after traumatic brain injury
combined with hemorrhage shock. BMC Neurosci. 21(22)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yunoki M, Kawauchi M, Ukita N, Noguchi Y,
Nishio S, Ono Y, Asari S, Ohmoto T, Asanuma M and Ogawa N: Effects
of lecithinized superoxide dismutase on traumatic brain injury in
rats. J Neurotrauma. 14:739–746. 1997.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Mikawa S, Kinouchi H, Kamii H, Gobbel GT,
Chen SF, Carlson E, Epstein CJ and Chan PH: Attenuation of acute
and chronic damage following traumatic brain injury in copper,
zinc-superoxide dismutase transgenic mice. J Neurosurg. 85:885–891.
1996.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Plotnikov MB, Chernysheva GA, Smolyakova
VI, Aliev OI, Trofimova ES, Sherstoboev EY, Osipenko AN, Khlebnikov
AI, Anfinogenova YJ, Schepetkin IA and Atochin DN: Neuroprotective
effects of a novel inhibitor of c-Jun N-terminal kinase in the rat
model of transient focal cerebral ischemia. Cells.
9(1860)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Khan R, Mallick N and Feroz Z:
Anti-inflammatory effects of Citrus sinensis L., Citrus paradisi L.
and their combinations. Pak J Pharm Sci. 29:843–852.
2016.PubMed/NCBI
|
|
15
|
Syaban M, Putra G, Vadhana R, Muhyiddin A,
Farida L, Sabila F, Haitsam M and Santoso WM: Molecular docking
analysis and dynamics simulation of ethanol extract of Citrus
sinensis as a Keap1 and NMDA inhibitor in brain injury. World Acad
Sci J. 5(14)2023.
|
|
16
|
Wang Z, Wang X, Li Y, Lei T, Wang E, Li D,
Kang Y, Zhu F and Hou T: farPPI: A webserver for accurate
prediction of protein-ligand binding structures for small-molecule
PPI inhibitors by MM/PB(GB)SA methods. Bioinformatics.
35:1777–1779. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Arviana S, Yueniwati Y, Rahayu M and
Syaban M: 7,8-dihydroxyflavone as a neuroprotective agent in
ischemic stroke through the regulation of HIF-1α protein. Res J
Pharm Technol. 15:3980–3986. 2022.
|
|
18
|
Arantes PR, Polêto MD, Pedebos C and
Ligabue-Braun R: Making it rain: Cloud-based molecular simulations
for everyone. J Chem Inf Model. 61:4852–4856. 2021.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Kurniawan DB, Syaban MFR, Mufidah A,
Zulfikri MUR and Riawan W: Protective effect of Saccharomyces
cerevisiae in Rattus norvegicus ischemic stroke model.
Res J Pharm Technol. 14:5785–5789. 2021.
|
|
20
|
Marmarou A, Foda MA, van den Brink W,
Campbell J, Kita H and Demetriadou K: A new model of diffuse brain
injury in rats: Part I: Pathophysiology and biomechanics. J
Neurosurg. 80:291–300. 1994.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kim HJ and Han SJ: A simple rat model of
mild traumatic brain injury: A device to reproduce anatomical and
neurological changes of mild traumatic brain injury. PeerJ.
5(e2818)2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Hoffman E and Winder SJ: A modified wire
hanging apparatus for small animal muscle function testing. PLoS
Curr.
8(ecurrents.md.1e2bec4e78697b7b0ff80ea25a1d38be)2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Du G, Zhao Z, Chen Y, Li Z, Tian Y, Liu Z,
Liu B and Song J: Quercetin protects rat cortical neurons against
traumatic brain injury. Mol Med Rep. 17:7859–7865. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Tandean S, Japardi I, Loe ML, Riawan W and
July J: Protective effects of propolis extract in a rat model of
traumatic brain injury via Hsp70 induction. Open Access Maced J Med
Sci. 7:2763–2766. 2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Syaban MFR, Faratisha IFD, Yunita KC,
Erwan E, Kurniawan DB and Putra GFA: Molecular docking and
interaction analysis of propolis compounds against SARS-CoV-2
receptor. J Tropical Life Sci. 12:219–230. 2022.
|
|
26
|
Yueniwati Y, Syaban MFR, Faratisha IFD,
Yunita KC, Putra GFA, Kurniawan DB, Putra GFA and Erwan NE:
Molecular docking approach of natural compound from herbal medicine
in java against severe acute respiratory syndrome coronavirus-2
receptor. Open Access Maced J Med Sci. 9:1181–1186. 2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Furukawa Y, Washimi YS, Hara RI, Yamaoka
M, Okuyama S, Sawamoto A and Nakajima M: Citrus Auraptene
induces expression of brain-derived neurotrophic factor in Neuro2a
cells. Molecules. 25(1117)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sawamoto A, Okuyama S, Nakajima M and
Furukawa Y: Citrus flavonoid 3,5,6,7,8,3',4'-heptamethoxyflavone
induces BDNF via cAMP/ERK/CREB signaling and reduces
phosphodiesterase activity in C6 cells. Pharmacol Rep. 71:653–658.
2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Sharma P, Kumari S, Sharma J, Purohit R
and Singh D: Hesperidin interacts with CREB-BDNF signaling pathway
to suppress pentylenetetrazole-induced convulsions in zebrafish.
Front Pharmacol. 11(607797)2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Selmi S, Rtibi K, Grami D, Sebai H and
Marzouki L: Protective effects of orange (Citrus sinensis
L.) peel aqueous extract and hesperidin on oxidative stress and
peptic ulcer induced by alcohol in rat. Lipids Health Dis.
16(152)2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Erukainure OL, Ajiboye JA, Davis FF,
Obabire K, Okoro EE, Adenekan SO, Adegbola M, Awogbemi BJ, Odjobo
BO and Zaruwa MZ: Effect of soy oil, orange (Citrus
sinensis) peel oil and their blends on total phospholipid,
lipid peroxidation, and antioxidant defense system in brain tissues
of normo rats. Grasas Y Aceites. 67(e113)2016.
|
|
32
|
Mawarti H, Khotimah MZA and Rajin M:
Ameliorative effect of Citrus aurantifolia and Cinnamomum
burmannii extracts on diabetic complications in a hyperglycemic
rat model. Trop J Pharm Res. 17(823)2018.
|
|
33
|
Malterud KE and Rydland KM: Inhibitors of
15-lipoxygenase from orange peel. J Agric Food Chem. 48:5576–5580.
2000.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Pepe G, Sommella E, Cianciarulo D,
Ostacolo C, Manfra M, Di Sarno V, Musella S, Russo M, Messore A,
Parrino B, et al: Polyphenolic extract from tarocco (Citrus
sinensis L. Osbeck) Clone ‘Lempso’ exerts anti-inflammatory and
antioxidant effects via NF-kB and Nrf-2 activation in murine
macrophages. Nutrients. 10(1961)2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Dasgupta A and Klein K: Antioxidant
Vitamins and Minerals. In: Antioxidants in Food, Vitamins and
Supplements: Prevention and Treatment of Disease. Elsevier,
Amsterdam, 277-294, 2014.
|
|
36
|
Rauf A, Uddin G and Ali J: Phytochemical
analysis and radical scavenging profile of juices of Citrus
sinensis, Citrus anrantifolia, and Citrus limonum. Org
Med Chem Lett. 4(5)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chen J, Li H, Yang C, He Y, Arai T, Huang
Q, Liu X and Miao L: Citrus naringenin increases neuron survival in
optic nerve crush injury model by inhibiting JNK-JUN pathway. Int J
Mol Sci. 23(385)2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Hunot S, Vila M, Teismann P, Davis RJ,
Hirsch EC, Przedborski S, Rakic P and Flavell RA: JNK-mediated
induction of cyclooxygenase 2 is required for neurodegeneration in
a mouse model of Parkinson's disease. Proc Natl Acad Sci USA.
101:665–670. 2004.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Kuan CY, Whitmarsh AJ, Yang DD, Liao G,
Schloemer AJ, Dong C, Bao J, Banasiak KJ, Haddad GG, Flavell RA, et
al: A critical role of neural-specific JNK3 for ischemic apoptosis.
Proc Natl Acad Sci USA. 100:15184–15189. 2003.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Pirianov G, Brywe KG, Mallard C, Edwards
AD, Flavell RA, Hagberg H and Mehmet H: Deletion of the c-Jun
N-terminal kinase 3 gene protects neonatal mice against cerebral
hypoxic-ischaemic injury. J Cereb Blood Flow Metab. 27:1022–1032.
2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Dhanasekaran DN and Reddy EP: JNK
signaling in apoptosis. Oncogene. 27:6245–6251. 2008.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Liu Y, Liu Q, Yang Z, Li R, Huang Z, Huang
Z, Liu J, Wu X, Lin J, Wu X and Zhu Q: Trihydroxyethyl rutin
provides neuroprotection in rats with cervical spinal cord
hemi-contusion. Front Neurosci. 15(759325)2021.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zhu W, Chi N, Zou P, Chen H, Tang G and
Zhao W: Effect of docosahexaenoic acid on traumatic brain injury in
rats. Exp Ther Med. 14:4411–4416. 2017.PubMed/NCBI View Article : Google Scholar
|