Introduction
A diverse microbial community of the human gut
microbiome has been shown to metabolize aromatic amino acids
(1,2). The secondary metabolites produced by
these reactions can affect multiple aspects of human well-being.
Among others, the intestinal bacterium, Clostridium
difficile (C. difficile), produces para-cresol
(p-cresol) by metabolizing tyrosine via p-hydroxyphenylacetate. For
a number of years, the final phenolic product, p-cresol, was known
as a uremic toxin, since, at high concentrations, this compound can
induce chronic renal disease symptoms (3). Other p-cresol-producing bacteria also
are present in the human gut, between them are different strains of
and Clostridiaceae, Bifidobacteriaceae, Enterobacteriaceae,
Coriobacteriaceae, Bacteroidaceae, Fusobacteriaceae and
Lactobacillaceae families (4). Still, compared with these, C.
difficile produces 10-1,000-fold more p-cresol and can be
regarded as a potent source of this metabolite (1,4). Due
to its hydrophobicity, this compound easily penetrates the
intestinal mucosa and reaches the bloodstream and all organs,
including the brain (5). The
p-cresol level is often significantly higher in the urine of
patients with autism spectrum disorder (ASD) and epilepsy. It can
be used as a possible biomarker of disease severity (3,6,7). In
previous studies, the inhibitory effects of p-cresol on the enzyme,
dopamine beta-hydroxylase (DBH), were shown to result in an
increase in dopamine levels and the diminished production of
norepinephrine (8,9). Thus, the inhibition of DBH possibly
dysregulates the activities of dopaminergic neurons and can cause
the aggravation of certain diseases, including major depressive
disorder and bipolar disorder (10). DBH gene mutations and decreased
levels of DHB in maternal serum have been shown to be associated
with autism in children (11,12),
suggesting the involvement of alterations in the dopaminergic
signaling in C. difficile-induced precipitation and aggravation of
autism spectrum disorder.
The majority of clinical laboratories assume that
all C. difficile isolates are sensitive to vancomycin, the
medication most frequently used in the treatment of C.
difficile-related illnesses (13). Recurrent C. difficile
infections are often treated with vancomycin taper regimens
(14). Previous studies have
demonstrated that oral vancomycin treatment causes a considerable
improvement in the symptoms of patients with ASD (15,16).
In a recent study, the authors found that a low concentration of
p-cresol, compared with a high concentration, potentiates nerve
growth factor-induced differentiation via the secretion of
brain-derived neurotrophic factor (BDNF) in cultured PC-12 cells,
demonstrating that low doses of p-cresol, in contrast to a high
toxic dose, can affect neuronal cell structural remodeling
(17). The present study examined
the effects of vancomycin on the behavioral responses of rats in
open-field tests and novel object recognition tasks. The results
presented herein demonstrate that the treatment of rats with
vancomycin affects the behavioral activity of the animals. These
changes occur in parallel with the decreases in p-cresol levels in
the brain. These data may indicate a direct effect of p-cresol on
the behavioral responses of rats.
Materials and methods
Animals
Animal care during the experimental procedures was
carried out by the recommendation of the Ilia State University
Research Projects Ethics Commission (Decision no. R/266-23) and by
the Council of Europe Directive 2010/63/EU for animal experiments.
Young adult (weight, 250-300 g) male Wistar rats obtained from
breeding colony of the vivarium of I. Beritashvili Center of
Experimental Biomedicine (Tbilisi, Georgia) were housed in cages (6
rats per cage) and provided with food and water available ad
libitum and maintained under conditions at a temperature of
20-22˚C and 40-55% humidity on a 12-h light/dark cycle (lights on
at 7:00 a.m.). The rats were subjected to an enforced swimming test
in order to exclude endogenic depression. Prior to commencing
treatment, all rats were marked and separated for 24 h. A total of
2 g of fecal specimens from a 12-h period were randomly collected
from each animal and stored at -80˚C. Following these procedures, a
total of 12 male rats (10 weeks old) were randomly assigned into
the following subgroups: i) The control group; and ii) the
vancomycin-treated group. Behavioral procedures were conducted
between 9:00 a.m. and 11: 00 a.m.
Following behavioral testing, all animals were
separated again for 24 h. A total of 2 g of fecal specimens from a
12-h period were randomly collected from each animal and stored at
-80˚C. Before and after placebo/vancomycin treatment, the fecal
specimens were subjected to the qualitative detection of C.
difficile. After performing behavioral tests, the rats were
euthanized by rapid decapitation using a hand-operated guillotine.
Decapitation was selected for euthanasia to avoid chemical
contamination for further analysis (https://rsawa.research.ucla.edu/arc/euthanasia-decapitation/).
After each use, the decapitation equipment was cleaned of any
biological fluids with ethanol and water and then wiped. Rat brains
were isolated on ice and homogenized rapidly in a 5x volume
ice-cold PBS (pH 7.4) using a hand Dounce homogenizer (Kontes Glass
Company), then centrifugated for 20 min at 1,000 x g at
4˚C.
Vancomycin treatment
Treatment was commenced 5 days following a 24-h
isolation to exclude the effects of social isolation. The rats in
the vancomycin-treated group received vancomycin once a day (light
cycle) (5 mg/kg body weight, average 1 mg per animal) with bread
for 3 weeks. The dose was selected based on published data to
minimize the effects of vancomycin on other bacterial species
(18). Vancomycin inhibits cell
wall assembly by binding to the D-Ala-D-Ala termini of lipid II and
has been shown to exhibit a high activity against C.
difficile (19). The rats in
the control group received the same portion of bread at the same
time of the day. Behavioral tests were commenced on the second day
following the completion of vancomycin treatment.
Open field test
The potential effects of vancomycin on locomotion
and anxiety-like behaviors were assessed using the open field test.
The device was an opaque Plexiglas arena with a size of 100x100x50
(H) cm. To ensure that the rats spent time in either the center or
the outside of the arena, it was divided into 25 small squares. The
rats were positioned in the center of the arena, and their behavior
was monitored and examined for 10 min. For each rat, the total
distance traveled, the total time spent and distance traveled in
the center segment, and the number of grooming and raising sessions
were recorded (20,21).
Novel object recognition task
Each animal received 10-min habituation sessions in
a plastic cage (50x50x40 cm3) that was equally lit and
devoid of objects. The animals were permitted to investigate two
identical objects that were always placed in the same location
within the box for 5 min (training session) at 24 h following the
habituation session. At 60 min after the 5-min training session,
the test session was started to assess memory retention. Both
well-known and unfamiliar objects were displayed simultaneously
during the training phase throughout the 10-min test period.
However, during the testing phase, the placement of new objects was
changed fictitiously and randomly to avoid the animals' innate
preference for one site over another. All objects (which were
provided in duplicate) had the same material and size, but various
shapes. Boxes and other materials were cleaned with 70% alcohol and
allowed to air dry between each change of the animals. The camera
recorded the time spent exploring, defined as smelling or touching
the object with the nose. The time spent examining each object or
novel object divided by the total time spent exploring both objects
throughout the training and testing stages multiplied by 100 in the
training and test phases, respectively, was finally defined as the
discrimination ratio (also known as the recognition index)
(20-22).
The time spent examining known and novel objects was separately
recorded to calculate the discrimination ratio for new object
recognition. This ratio was defined as the exploration time
differential for the novel object divided by the total exploration
time. The more time spent investigating the novel object, the more
successfully the memory could distinguish it from the familiar
object (23). Therefore, an
increase in the discrimination ratio during the test phase
indicated an improvement in recognition memory, whereas a decrease
indicated the opposite. To reduce any potential item preference
that may skew the results, diverse objects were choose in order to
be easily distinguished by the rats, while having a similar
complexity level (texture, form, color patterning, brightness,
etc.) (24). It was ensured that
the rats could climb on the objects to prevent induced preference.
By contrast, the capacity to climb on an object may boost interest
in exploration (time spent simply sitting on an object is not
counted toward the exploration time). As the loss of an object due
to damage during experimentation could interfere with the
continuation of testing and possibly result in harm or injury to
the animal, objects made of non-breakable material were used. In
the experiment, two similar objects were placed horizontally in the
box (i.e., one in the north-west corner and one in the north-east
corner) during the training phase; one unfamiliar object was
substituted for one of the familiar objects during the testing
phase (23,25).
Qualitative detection of C.
difficile
The glutamate dehydrogenase (GDH)-based detection of
C. difficile in the rat fecal specimens from both groups was
made before and after placebo/vancomycin treatment using the
Abnova™ Clostridium difficile GDH ELISA kit (KA3381, Abnova).
Briefly, 2 g fecal specimens from a 12-h period obtained were
randomly collected from each animal and stored at -80˚C. Before and
after treatment, the fecal specimens were analyzed simultaneously
to avoid the technical error of the results. The fecal specimens of
each animal were mixed and homogenized using a wooden applicator
stick to ensure adequate sampling, and 0.2 g of the obtained sample
were transferred to the test tube with 400 µl Sample Diluent for
vortex and decantation (15 min). All ensuing procedures were
performed according to the manufacturer's protocol. The fecal
specimens of each animal were analyzed in triplicate. Respective
positive and negative controls were used to confirm the validity of
the experiment. The absorbance was determined at a wavelength of
450, and 630 nm was used as a reference wavelength.
Assessment of the P-cresol amount
The concentration of p-cresol in the brain
homogenate was performed using an ELISA-based kit (EK711090; AFG
Bioscience) according to the manufacturer's protocol. The rat
brains were homogenized rapidly in a 5x volume PBS (pH 7.4) using a
hand Dounce homogenizer, then centrifuged for 20 min at 1,000 x g
at 4˚C. The supernatant was removed and assayed immediately.
Statistical analysis
One-way analysis of variance (ANOVA) was applied
when analyzing all the data obtained from the unique objective and
open field tests. ANY-maze Video Tracking Software 60000 (Stoelting
Co.) was used for this purpose. Tukey's post hoc multiple
comparison test was used to identify the statistically significant
differences between the groups. Statistics were used to express the
data as the mean ± SEM, and a value of P<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of vancomycin on novel object
recognition
The analysis of the effect of vancomycin on the
novel objective recognition test revealed that both groups spent
approximately the same amount of time investigating the two
familiar objects in the first trial (training phase). There was no
statistically significant difference between the groups. Object
recognition memory was tested by swapping out the object of the
first trial for copies of the original (familiar object) and
unfamiliar objects in the second trial (test phase). Performance
for examining the novel object reflected memory for novel object
identification. The vancomycin-treated group exhibited a
substantial decrease in the new objective recognition rate compared
to the control group (P<0.05; Table
I).
 | Table IEffects of vancomycin on anxiety-like
behaviors in the open field and novel object recognition tests. |
Table I
Effects of vancomycin on anxiety-like
behaviors in the open field and novel object recognition tests.
| Test | Parameter | Control | Vancomycin | P-value |
|---|
| Open field | Line cross
(number) | 337 | 206 |
0.00888a |
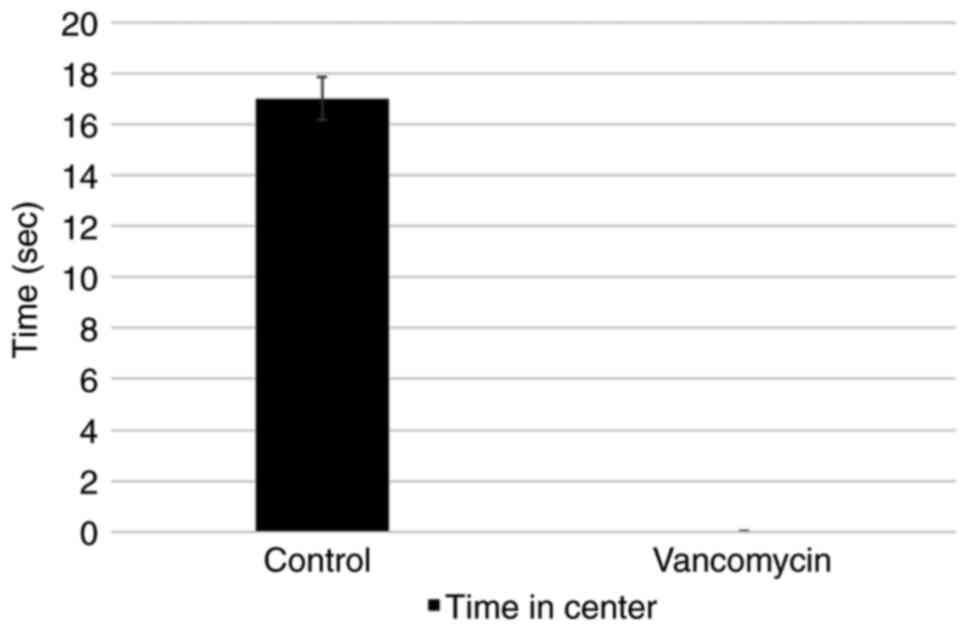
| Open field | Center time
(sec) | 17 | 0 |
0.02757b |
| Open field | Center cross
(number) | 8 | 2 |
0.05528c |
| Open field | Rearing
(number) | 196 | 104 |
0.00711a |
| Novel object | Preference of 2nd
object (day 2), % | 59 | 36 | 0.0024a |
Effects of vancomycin on grooming and
rearing
No significant differences were observed among the
groups in the grooming number (data not shown); however, the
rearing number was significantly lower in the vancomycin-treated
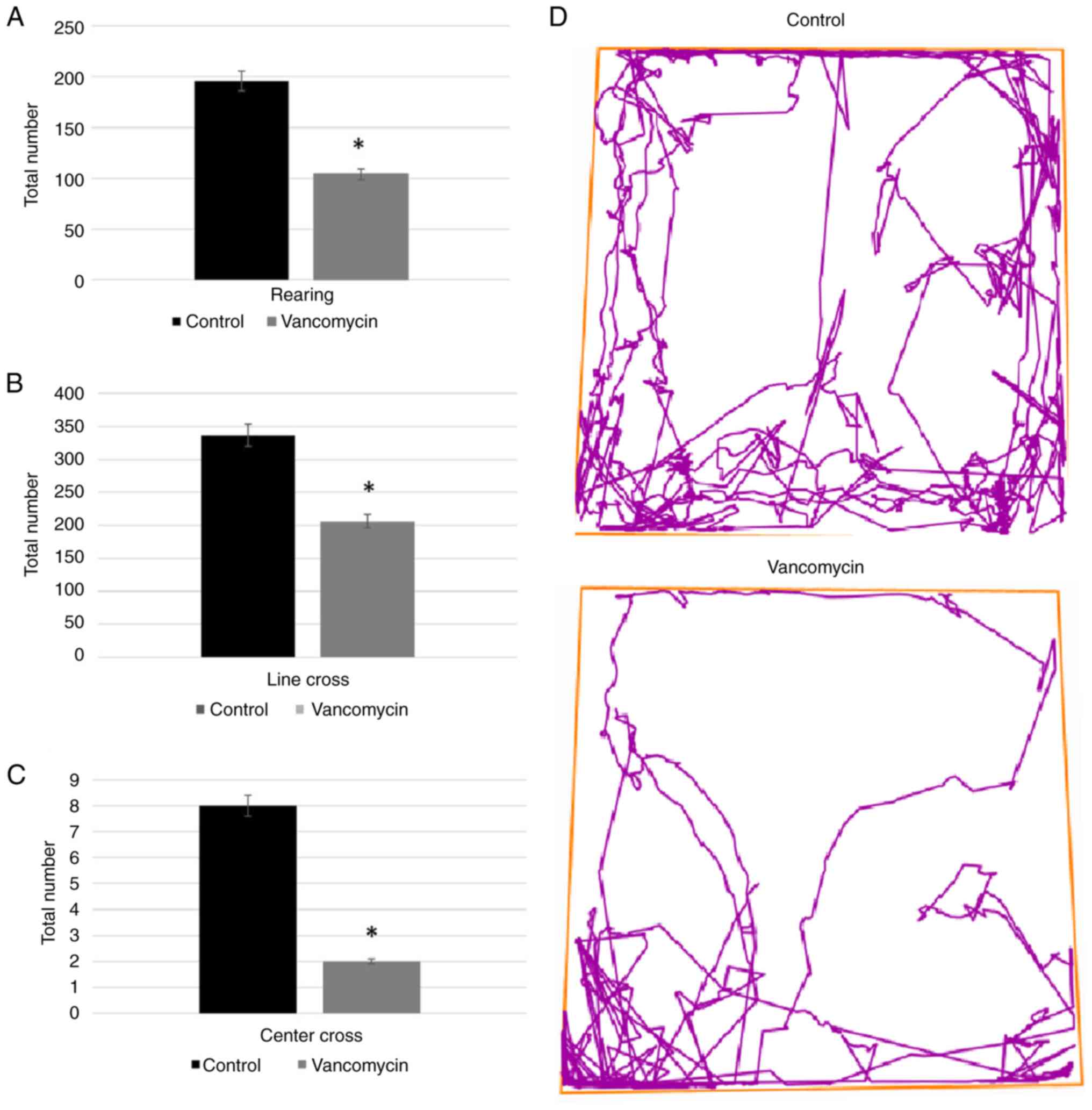
group (Fig. 1A). Rearing is a
vertical activity; however, unlike locomotion, it is carried out
while the animal stands on its rear legs and rests against the cage
walls. The animal performs the movement in an aim to become
familiar with the environment and, generally, to look for a food
source. The velocity of movement in the center and the periphery of
the box (Fig. 1D) was assessed by
the corresponding line crossing quantity and was significantly
lower in the vancomycin-treated group compared to the similar
parameters of the control group (Fig.
1B and C). The
vancomycin-treated group did not spend time in the center (Fig. 2).
Qualitative detection of C.
difficile
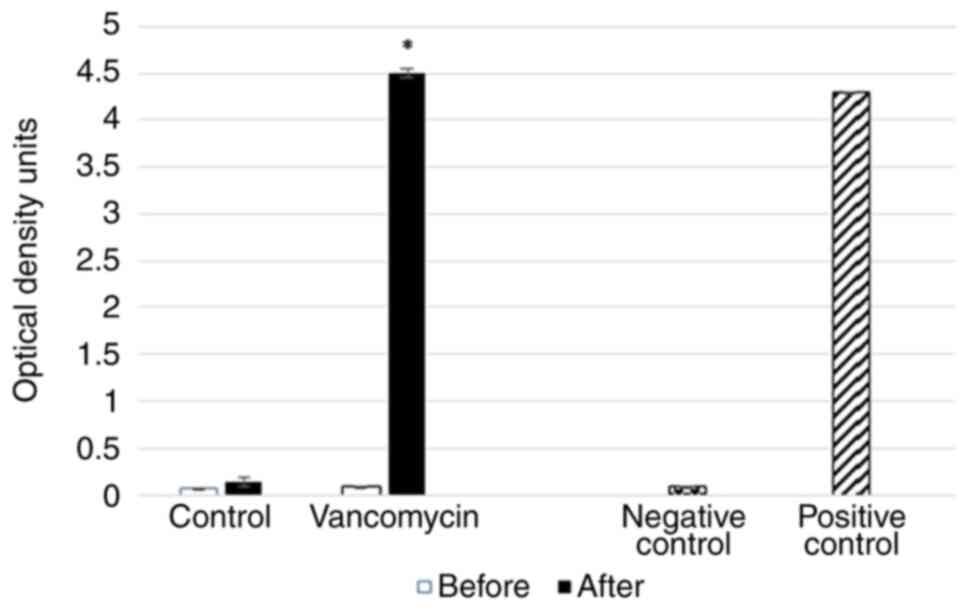
The qualitative detection of C. difficile GDH
in the animal fecal specimens was made using relevant ELISA tests
before and after placebo/vancomycin treatment. The analysis of the
obtained data with positive and negative controls (4.29 and 0.081,
respectively) revealed positive tests only in the fecal specimens
of the vancomycin-treated rats (Fig.
3). Since oral vancomycin is the first-line treatment against
the Clostridium species, and C. difficile is a major
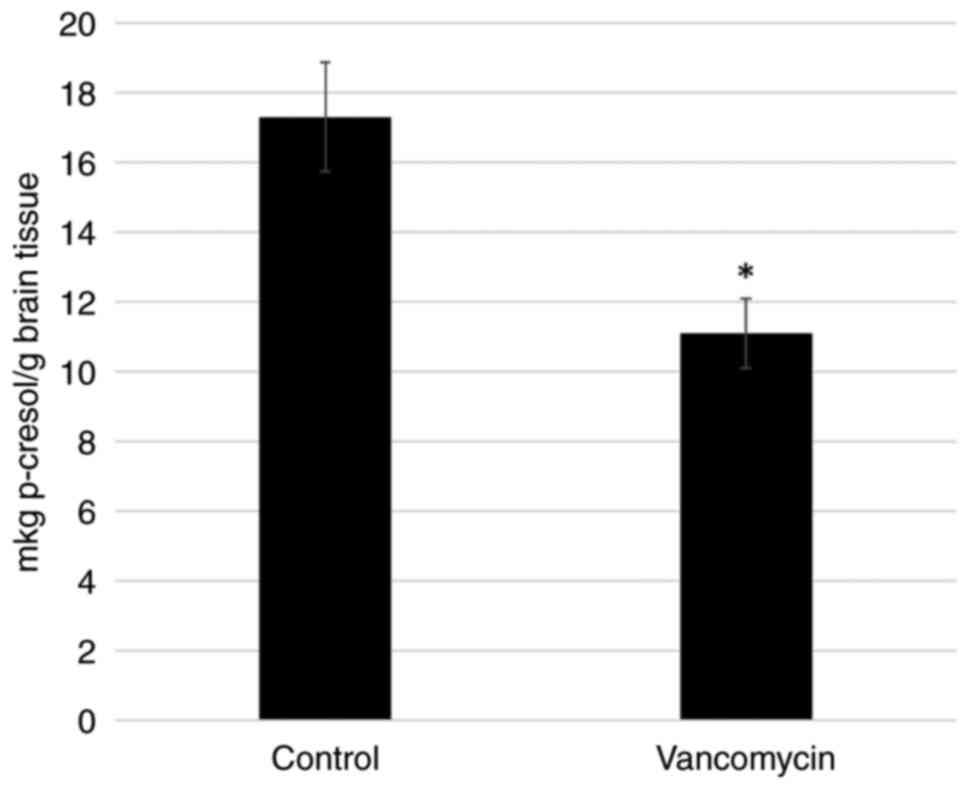
source of p-cresol, the p-cresol content was determined in the
brains of the control and vancomycin-treated rats. It was found
that the content of p-cresol in the vancomycin-treated animals was
significantly lower compared to that of the control group rats
(Fig. 4). These data indicate that
changes in the amount of p-cresol may occur due to the effects of
vancomycin on the intestinal microbiome, apparently through a
decrease in the Clostridium species.
Discussion
The enteropathogen, C. difficile, an
opportunistic anaerobic bacterium, can be carried by animals and
humans and distributed globally (26). C. difficile has also been
found in rodents, including rats and mice (27). C. difficile produces high
concentrations of p-cresol through the fermentation of tyrosine,
which, apart from perturbations in the beneficial microbiome
through gut-brain axes, changes the dopamine metabolism of the
brain (4).
A recent study by the authors using rat
pheochromocytoma cells (PC-12 cells) demonstrated the effects of
p-cresol on the secretion of BDNF and neurofilament subunit
expression (17). Low doses of
p-cresol enhanced BDNF secretion and nerve growth factor induced
the differentiation in PC-12 cells in culture. It was hypothesized
that low doses of p-cresol may cause mild oxidative stress that
stimulates the release of BDNF by turning on redox-sensitive genes
(17). Given that the intestinal
microbiome is the main source of p-cresol, the equilibrium between
gut microbiome strains, particularly Clostridium species,
and various endogenous neuroactive substances, such as opioids, may
directly affect neuroplasticity.
Vancomycin is increasingly used as the first-line
drug in the treatment of Clostridium infections (28). Notably, the selected dose of
vancomycin and the route of administration (oral) used in the
present study significantly decrease its bactericidal effects on
other target bacteria (e.g., Staphylococcus aureus). It was
recently shown that oral vancomycin administration selectively
affected indole and p-cresol-producing gut bacterial taxa from
Clostridium species (29).
The specificity of vancomycin against Clostridium lies in
its mechanism of action. Vancomycin is a glycopeptide antibiotic
that inhibits cell wall synthesis in susceptible bacteria. This
antibiotic binds to D-alanyl D-alanine and inhibits
glucosyltransferase and the P-phospholipid carrier, thereby
preventing the synthesis and polymerization of N-acetylmuramic acid
and N-acetylglucosamine within the peptidoglycan layer of bacteria
(30). In the case of C.
difficile, the binding of vancomycin disrupts the cell wall
synthesis process, weakening or killing the bacteria. One of the
reasons vancomycin is effective against C. difficile
infections is that this bacterium is susceptible to the antibiotic
due to the unique composition of its cell wall. Unlike certain
other Gram-positive bacteria (e.g., Bacillus, Campylobacter,
Lactobacillus), the cell walls of C. difficile contain a
unique peptidoglycan structure that allows vancomycin to bind
effectively and disrupt cell wall synthesis (31,32).
This inhibition weakens bacterial cell walls and ultimately causes
the leakage of intracellular components, resulting in cell death.
Thus, lysed cell fragments, including GDH, can be liberated into
the stool. The present study revealed an increase in the release of
such fragments under the effects of vancomycin.
The present study is the continuation of previous
research (7). The results of the
present study regarding the novel object recognition test revealed
that vancomycin administration in the rats significantly decreased
the new object recognition processes compared to the control rats.
Additionally, the results from the open field test revealed that
the vancomycin-treated rats traveled a substantially shorter
distance in the periphery of the box than the control group.
Furthermore, compared to the control group, this one's movement in
the box's middle was noticeably farther. Moreover, the number of
rearings in the vancomycin-treated group was significantly lower
than that in the control group rats. These data suggest that
vancomycin decreases rats' behavioral projection of cognitive
ability. This may indicate a potential link between C.
difficile colonies in the gut and the cognitive functions of
the brain. Furthermore, the administration of vancomycin in rats,
in parallel with disruptions in behavioral responses, decreases the
content of p-cresol in the brain. It is thus suggested that this
interaction is carried out through the dopaminergic activity of the
brain.
Acknowledgements
Not applicable.
Funding
Funding: The Basic Science Research Program of Ilia State
University supported the present study by providing the annual
budget for the purchasing of reagents.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GT proposed the main hypothesis for the study,
designed the main experiments and participated in the writing of
the manuscript. NK carried out the main experiments with animal
behavior. EZ designed the main stages of the research protocol, and
organized the study and participated in data analysis. NN carried
out the animal euthanasia, decapitation and brain homogenization.
TB performed the biochemical analysis of p-cresol, and participated
in behavioral recording and analysis. LS performed the statistical
analysis of the obtained data and prepared the figures and graphs
for the study. DM performed the interpretation of the obtained
data, prepared the introduction of obtained data, and prepared the
introduction and discussion sections of the manuscript. EZ and TB
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
Animal care during experimental procedures was
carried out by the recommendation of the Ilia State University
Research Projects Ethics Commission (Decision no. R/266-23) and by
Council of Europe Directive 2010/63/EU for animal experiments.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Russell WR, Duncan SH, Scobbie L, Duncan
G, Cantlay L, Calder AG, Anderson SE and Flint HJ: Major
phenylpropanoid-derived metabolites in the human gut can arise from
microbial fermentation of protein. Mol Nutr Food Res. 57:523–535.
2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lin L and Zhang J: Role of intestinal
microbiota and metabolites on gut homeostasis and human diseases.
BMC Immunol. 18(2)2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Persico AM and Napolioni V: Urinary
p-cresol in autism spectrum disorder. Neurotoxicol Teratol.
36:82–90. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Saito Y, Sato T, Nomoto K and Tsuji H:
Identification of phenol- and p-cresol-producing intestinal
bacteria by using media supplemented with tyrosine and its
metabolites. FEMS Microbiol Ecol. 94(fiy125)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Morinaga Y, Fuke C, Arao T and Miyazaki T:
Quantitative analysis of cresol and its metabolites in biological
materials and distribution in rats after oral administration. Leg
Med (Tokyo). 6:32–40. 2004.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Gabriele S, Sacco R, Cerullo S, Neri C,
Urbani A, Tripi G, Malvy J, Barthelemy C, Bonnet-Brihault F and
Persico AM: Urinary p-cresol is elevated in young French children
with autism spectrum disorder: A replication study. Biomarkers.
19:463–470. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tevzadze G, Shanshiashvilli L and
Mikeladze D: Children with epilepsy and autistic spectrum disorders
show similarly high levels of urinary p-cresol. J Biol Phys Chem.
17:77–80. 2017.
|
|
8
|
DeWolf WE Jr, Carr SA, Varrichio A,
Goodhart PJ, Mentzer MA, Roberts GD, Southan C, Dolle RE and Kruse
LI: Inactivation of dopamine .beta.-hydroxylase by p-cresol:
Isolation and characterization of covalently modified active site
peptides. Biochemistry. 27:9093–9101. 1988.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Goodhart PJ, DeWolf WE and Kruse LI:
Mechanism-based inactivation of dopamine beta.-hydroxylase by
p-cresol and related alkylphenols. Biochemistry. 26:2576–2583.
1987.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sun Z, Bo Q, Mao Z, Li F, He F, Pao C, Li
W, He Y, Ma X and Wang C: Reduced plasma dopamine-β-hydroxylase
activity is associated with the severity of bipolar disorder: A
pilot study. Front Psychiatry. 12(566091)2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Robinson PD, Schutz CK, Macciardi F, White
BN and Holden JJA: Genetically determined low maternal serum
dopamine b-hydroxylase levels and the etiology of autism spectrum
disorders. Am J Med Genet. 100:30–36. 2001.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Perkovic MN, Erjavec GN, Stefulj J,
Muck-Seler D, Pivac N, Hercigonja DK, Hranilovic D, Curkovic M and
Dodig-Curkovic K: Association between the polymorphisms of the
selected genes encoding dopaminergic system with ADHD and autism.
Psychiatry Res. 215:260–261. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Peláez T, Alcalá L, Alonso R,
Rodríguez-Créixems M, García-Lechuz JM and Bouza E: Reassessment
of Clostridium difficile susceptibility to metronidazole and
vancomycin. Antimicrob Agents Chemother. 46:1647–1650.
2002.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Johnson S: Recurrent Clostridium
difficile infection: A review of risk factors, treatments, and
outcomes. J Infect. 58:403–410. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Settanni CR, Bibbò S, Ianiro G, Rinninella
E, Cintoni M, Mele MC, Cammarota G and Gasbarrini A:
Gastrointestinal involvement of autism spectrum disorder: Focus on
gut microbiota. Expert Rev Gastroenterol Hepatol. 15:599–622.
2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Alharthi A, Alhazmi S, Alburae N and
Bahieldin A: The human gut microbiome as a potential factor in
autism spectrum disorder. Int J Mol Sci. 23(1363)2022.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tevzadze G, Barbakadze T, Kvergelidze E,
Zhuravliova E, Shanshiashvili L and Mikeladze D: Gut neurotoxin
p-cresol induces brain-derived neurotrophic factor secretion and
increases the expression of neurofilament subunits in PC-12 cells.
AIMS Neurosci. 9:12–23. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Saribas Z, Ergun H, Mamuk S, Köseoglu-Eser
O and Melli M: Critical appraisal of air pouch infection model in
rats. Ann Clin Lab Sci. 42:50–56. 2012.PubMed/NCBI
|
|
19
|
Gonzales M, Pepin J, Frost EH, Carrier JC,
Sirard S, Fortier LC and Valiquette L: Faecal pharmacokinetics of
orally administered vancomycin in patients with suspected
Clostridium difficile infection. BMC Infect Dis.
10(363)2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Rajizadeh MA, Sheibani V, Bejeshk MA,
Borzadaran FM, Saghari H and Esmaeilpour K: The effects of high
intensity exercise on learning and memory impairments followed by
combination of sleep deprivation and demyelination induced by
etidium bromide. Int J Neurosci. 129:1166–1178. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rajizadeh MA, Esmaeilpour K,
Masoumi-Ardakani Y, Bejeshk MA, Shabani M, Nakhaee N, Ranjbar MP,
Borzadaran FM and Sheibani V: Voluntary exercise impact on
cognitive impairments in sleep-deprived intact female rats. Physiol
Behav. 188:58–66. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Esmaeilpour K, Sheibani V, Shabani M and
Mirnajafi-Zadeh J: Low frequency electrical stimulation has time
dependent improving effect on kindling-induced impairment in
long-term potentiation in rats. Brain Res. 1668:20–27.
2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Iulita MF, Allard S, Richter L, Munter LM,
Ducatenzeiler A, Weise C, Do Carmo S, Klein WL, Multhaup G and
Cuello AC: Intracellular Aβ pathology and early cognitive
impairments in a transgenic rat overexpressing human amyloid
precursor protein: A multidimensional study. Acta Neuropathol
Commun. 2(61)2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lueptow LM: Novel object recognition test
for the investigation of learning and memory in mice. J Vis Exp.
126(55718)2017.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Ennaceur A: One-trial object recognition
in rats and mice: Methodological and theoretical issues. Behav
Brain Res. 215:244–254. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Public Health England. Clostridioides
difficile: guidance, data and analysis. Available from:
https://www.gov.uk/government/collections/clostridium-difficile-guidance-data-and-analysis.
|
|
27
|
Krijger IM, Meerburg BG, Harmanus C and
Burt SA: Clostridium difficile in wild rodents and
insectivores in the Netherlands. Lett Appl Microbiol. 69:35–40.
2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chiu CY, Sarwal A, Feinstein A and
Hennessey K: Effective dosage of oral vancomycin in treatment for
initial episode of clostridioides difficile infection: A
systematic review and meta-analysis. Antibiotics.
8(173)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Nazzal L, Soiefer L, Chang M, Tamizuddin
F, Schatoff D, Cofer L, Aguero-Rosenfeld ME, Matalon A, Meijers B,
Holzman R and Lowenstein J: Effect of vancomycin on the gut
microbiome and plasma concentrations of gut-derived uremic solutes.
Kidney Int Rep. 6:2122–2133. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Koyama N, Inokoshi J and Tomoda H:
Anti-infectious agents against MRSA. Molecules. 18:204–224.
2012.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kirk JA, Banerji O and Fagan RP:
Characteristics of the Clostridium difficile cell envelope
and its importance in therapeutics. Microb Biotechnol. 10:76–90.
2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Eubank TA, Gonzales-Luna AJ, Hurdle JG and
Garey KW: Genetic mechanisms of vancomycin resistance in
clostridioides difficile: A systematic review. Antibiotics
(Basel). 11(258)2022.PubMed/NCBI View Article : Google Scholar
|