Introduction
Cerebral infarction, a critical medical condition,
occurs when blood flow to a specific region of the brain is
blocked. This obstruction results in damage to the affected tissue
and the possibility of neurological impairment (1). Stroke is a medical condition
characterized by a sudden, localized neurological impairment caused
by damage to the central nervous system due to either a blockage or
bleeding in the blood vessels. Stroke is a critical public health
concern due to its significant impact on both mortality and
disability statistics. An obstruction of blood flow to the brain
characterizes ischemic stroke cases, which comprise ~87% of all
stroke cases. The obstruction of blood flow is often attributed to
the presence of atherosclerosis and thrombosis within the cerebral
arteries. Cerebral infarction is characterized by a complex
interaction of molecular processes. Inflammation and oxidation play
a central role in the state of cerebral infarction (2,3).
The increased production of reactive oxygen species
(ROS) and inflammation creates a detrimental feedback loop, as ROS
have the ability to initiate pro-inflammatory pathways and
stimulate the secretion of cytokines, thereby exacerbating tissue
injury and inflammation associated with brain injury. Oxidative
stress, along with malondialdehyde (MDA), is recognized as a major
contributor to the complex pathophysiology of brain injury
(2). The decrease in MDA levels
may be associated with the upregulated expression of nuclear factor
E2-related factor 2 (Nrf2), glutathione S-transferase (GST) and
glutathione peroxidase (GPX), which are antioxidant enzymes
activated by the Kelch like ECH-associated protein 1 (Keap1)/Nrf2
pathway (4). Researchers have
previously found that the blocking of Keap1 increases the
production of Nrf2. Nrf2 is a key factor in protecting neurons from
ischemic stroke and reducing brain damage caused by oxidative
stress. This process of activating the Keap1/Nrf2 pathway has also
been linked to higher levels of GPX, an enzyme that plays a
critical role in eliminating ROS and lipid hydroperoxides. The
Keap1/Nrf2 pathway has exhibited potential in mitigating oxidative
stress and providing neuroprotection in stroke and several
neurodegenerative diseases (5-7).
The Keap1/Nrf2 pathway controls the antioxidant
response and has been shown to influence inflammation in a number
of diseases, including ischemic stroke (8). Reactive nitrogen species bind to a
key regulatory cysteine thiol in the Keap1 protein during
oxidative-nitrosative stress (9).
When this chemical reaction occurs, Keap1 adducts are formed that
break the link between Keap1 and Nrf2. The disruption prevents
Keap1 from degrading Nrf2, thereby facilitating the accumulation of
Nrf2 in the nucleus. This localization allows Nrf2 to initiate the
activation of genes associated with the antioxidant response. It
has also been shown that a few small compounds can prevent Keap1
from functioning by S-alkylating the important thiol group. This
results in the suppression of the ability of Keap1 to degrade Nrf2
and instead promotes the activation of antioxidant defense pathways
(9,10).
For the prevention of oxidative damage in
cardiovascular tissue, an expanding array of medicinal herbs are
employed as innate antioxidants in clinical settings (11). Antioxidant activity can be
extracted from several medicinal plants, yielding active compounds
(12,13). Natural antioxidant medications have
the ability to reduce the generation of harmful free radicals by
boosting both particular and general immune responses, or by
directly limiting the cellular and tissue harm caused by free
radicals. Flavonoids, a class of organic chemicals found naturally
in plants, have exhibited great promise as agents with
anti-inflammatory properties. This is attributed to their ability
to modulate pro-inflammatory genes and inhibit the function of
enzymes involved in the inflammatory cascade (14). One particular flavonoid that has
been the subject of extensive research for its anti-inflammatory
properties is 7,8-dihydroxyflavone (7,8-DHF). Previous research has
documented the robust anti-inflammatory capabilities of 7,8-DHF
(15). Through the downregulation
of the nuclear factor-κB (NF-κB) and mitogen-activated protein
kinase (MAPK) pathways in lipopolysaccharide-treated RAW264.7
cells, the study by Park et al (15) demonstrated that 7,8-DHF exhibited
anti-inflammatory properties. 7,8-DHF directly suppressed the
inflammatory response in an obese adipose cell line by disrupting
the paracrine loop between adipocytes and macrophages. This is
evidenced by the reduction in pro-inflammatory cytokines and the
release of non-esterified fatty acids (16-18).
7,8-DHF may also decrease the activity of inflammatory mediators by
inhibiting the JNK and NF-κB signaling pathways. Furthermore,
7,8-DHF has been shown to reverse the decreased absorption of
glucose in the combined culture of adipocytes and macrophages,
along with an elevation in the phosphorylation of Akt (19). Thus, 7,8-DHF treatment is also
effective in reducing blood glucose levels.
The unusual properties of 7,8-DHF have attracted
considerable attention in the fields of neuroscience and neurology.
It has been found that the compound in question binds to and
activates the tyrosine kinase receptor B (TrkB), which is linked to
brain-derived neurotrophic factor (BDNF). A notable feature of
7,8-DHF is its ability to cross the blood-brain barrier (BBB);
thus, it holds promise for use in neurological applications
(16). Thus, it is worthy to
investigate the role of 7,8-DHF in stroke. Numerous studies have
been conducted to investigate the antioxidant properties of 7,8-DHF
(17,18,20);
however, further investigations are required to fully elucidate the
underlying mechanisms responsible for its antioxidant effects. It
is important to know the potential of 7,8 DHF for the treatment of
ischemic stroke.
The present study aimed to explore 7,8-DHF as an
antioxidant agent using in vivo and in silico
analyses. The present study demonstrates the potential of 7,8-DHF
as an antioxidant using molecular docking techniques and in
vivo experiments in a rat model of ischemia-reperfusion brain
injury.
Materials and methods
Experimental animals
A total of 25 male rats (Rattus norvegicus),
aged between 7 and 9 weeks and weighing at least 200 g from the
Animal House Faculty of Medicine Universitas Brawijaya (Malang,
Indonesia), were used in the present study. The duration of the
study was 4 months. These rats were divided into five different
groups, with each group consisting of 5 rats, as demonstrated in
Table I. The initial total number
of experimental animals used was 30 rats, with 6 rats per group. At
the final stage of the experiment, there were 2 rats that died
naturally and 3 rats that died following middle cerebral artery
occlusion (MCAO) due to complications associated with this
procedure. A total of 25 rats were used for parameter analysis. The
selection of these groups was made to evaluate the efficacy of
7,8-DHF treatment compared to the placebo and to normal rats. The
rats were placed in a suitable housing habitat and allowed to
acclimatize at room temperature (22-28˚C). The humidity in the room
was maintained at 45-50%, with a 12-h light-dark cycle. Food and
water were provided ad libitum. Each week, the experimental
animals were examined and monitored concerning overall health,
neurological functions, animal behavior and physical parameters,
such as body weight and body condition score. The indication for
euthanasia was determined based on various factors, including
self-mutilation, the amputation or crushing of limbs and tail,
generalized dermatitis, infection and severe dehydration. Death was
determined based on the absence of respiration, absence of
heartbeat, dilated pupils, the absence of reflexes, poor
pigmentation of the extremities and mucous membranes, and the
presence of dry and opaque corneas. The research was carried out
following internationally recognized ethical principles and the
guidelines outlined in the National Institutes of Health's Guide
for the Care and Use of Laboratory Animals of the Medical Faculty,
Brawijaya University (No. 160/EC/KEPK/09/2020).
 | Table IExperimental animal groups. |
Table I
Experimental animal groups.
| Group | Treatment
administered |
|---|
| Sham | Operation without
MCAO + normal saline for 14 days |
| MCAOP | MCAO induction +
normal saline for 14 days |
| MCAO10 | MCAO induction +
normal diet + 7,8-DHF administered orally at dose 10 mg/kg weight
body for 14 days |
| MCAO20 | MCAO induction +
normal diet + 7,8-DHF administered orally at dose 20 mg/kg weight
body for 14 days |
| MCAO50 | MCAO induction +
normal diet + 7,8-DHF administered orally at dose 50 mg/kg weight
body for 14 days |
MCAO
The experimental procedure involved the induction of
ischemia-reperfusion brain injury in the rats by performing MCAO.
In the present study, the rats were injected with ketamine (40-100
mg/kg) and xylazine (5-13 mg/kg) to induce and maintain anesthesia.
The experimental setup included the use of a heat lamp and a
heating pad to maintain body temperature at a constant level of
37˚C. A longitudinal incision of ~2 cm was made along the midline
of the neck. The right common carotid artery (CCA) was carefully
dissected from the vagus nerve and briefly occluded with cotton
thread. The left external carotid artery was ligated at its
bifurcation into the internal and external carotid arteries.
Following a delicate ligation procedure on the right internal
carotid artery (ICA), a 3-0 monofilament nylon was carefully
inserted into the ICA through the CCA incision. The nylon material
exhibited signs of detachment over a 90-min occlusion period. Upon
the completion of the experimental procedure, the rats were
transferred to the incubator for specialized and attentive medical
care (21,22). In the sham-operated (sham) group,
the rats underwent an identical surgical technique but without the
induction of MCAO. Following 14 days of treatment, the rats were
euthanized with intraperitoneal overdoses of ketamine (40-100 mg/kg
body weight) and xylazine (5-13 mg/kg body weight), followed by
decapitation. Blood and brain tissue samples were collected. Brain
tissue samples were fixed overnight by immersion in a 10% formalin
solution.
7,8-DHF treatment
7,8-DHF was obtained from the manufacturer, Tokyo
Chemical Industry Co., Ltd. (product no. D1916; lot no. OQ5VB). The
7,8-DHF isolate has a purity of 99.5%, as shown by high performance
liquid chromatography and 100% by neutralization titration (data
not shown). This isolate was dissolved using a concentration of 10%
dimethyl sulfoxide (Cas No. 67-68-5, MilliporeSigma) for the
dissolution (23). Isolate
preparations of 7,8-DHF were prepared every 5 days to prevent
oxidation and damage. The administration of 7,8-DHF was performed
by a professional assistant using a sonde/feeding tube (Thermo
Fisher Scientific, Inc.) as doses, as demonstrated in Table I.
Immunohistochemical staining
Immunohistochemistry (IHC) was used to measure the
amount of Nrf2 and GPX-1 protein in the brain tissues of rats in
the model of ischemic stroke. The kit used was from Bioss with the
following antibodies: [Nrf2, cat. no. bs-2013R; (1:75), and GPX-1,
cat.no. bs-3882R; (1:75)]. The calculation was based on the amount
of Nrf2, and GPX-1 protein found in the cytoplasm and nucleus of
brain tissue in the cortical lesion area. This included neurons,
and glia and Schwann cells observed under a microscope at x400
magnification. Briefly, the brain tissue samples were
deparaffinized and stored at room temperature for 24 h prior to
use. Following deparaffinization, the slides were immersed in a
sodium citrate buffer solution (pH 6.0) and then washed with
phosphate-buffered saline (PBS) (P-5119, MilliporeSigma) three
times for 5 min. Blocking was performed with a blocking buffer
(skimmed milk) (Blocker BLOTTO in Tris-buffered saline; cat. no
37530, Thermo Fisher Scientific, Inc.) followed by incubation for
60 min at room temperature. Nrf2 polyclonal antibody, unconjugated
(bs-2013R), GPX-1 polyclonal antibody, unconjugated (bs-3882R) at a
ratio of 1:75 were applied overnight at 4˚C. Secondary antibodies
(rabbit anti-rat IgM antibody (H+L), biotin-conjugated; 1:200;
bs-0346R-Biotin; Bioss) were used and incubated with the samples
for 60 min at room temperature, then washed three times for 5 min
with PBS. Streptavidin HRP was then incubated with the samples for
40 min at room temperature. The chromogen diaminobenzidine (DAB;
AMF080, ScyTek Laboratories, Inc.) ratio was 1:40 (DAB chromogen:
DAB buffer) and the samples were incubated for 5 min at room
temperature. Mayer's hematoxylin (cat. no. 109249, MilliporeSigma)
(Mayer's hematoxylin: distilled water, 1:3) was added, incubated
for 2 min at room temperature, then washed with distilled water and
mounted with Entella. The final result was observed under a
confocal microscope (BX51, Olympus Corporation) at x400
magnification. To minimize examination bias, IHC calculations were
conducted by two observers under the supervision of a pathologist.
The counting of positive cells was performed in a single-blind
manner, with codes known only to one individual to confirm the
results.
Enzyme-linked immunosorbent assay
(ELISA)
To assess the extent of oxidative damage in the
brain tissue, the concentration of MDA, a reliable indicator of
lipid peroxidation, was quantified in the brain samples. This was
performed using a commercially available ELISA kit (MDA
Elabscience® ELISA kit, cat. no. E-EL-0060, Elabscience
Biotechnology, Inc.) according to the manufacturer's
instructions.
Absorption, distribution, metabolism,
excretion and toxicity (ADMET) and drug-likeness
The ADMET of 7,8-DHF was evaluated using SwissADME
(http://www.swissadme.ch/) by inserting the SMILES
formula of 7,8-DHF. Lipinski's rule of 5 (LR5) was used to evaluate
the pharmacokinetic potential of the compounds. A chemical can be
classified as a drug if it meets the LR5 criteria. Finally, the
OSIRIS software tool (https://www.organic-chemistry.org/prog/peo/) was used
to predict the human safety of a compound based on its toxicity
analysis (24,25).
Biological activity and molecular
potential prediction
To derermine the potential activity of 7,8-DHF, the
biological activity was measured using the prediction of activity
spectra for substances (PASS) online web server (http://www.way2drug.com/index.php). The purpose
of the web server is to predict the biological activity spectrum of
organic compounds by analyzing their structural formulas. It covers
a wide range of >4,000 biological activities and achieves an
average accuracy rate of >95%. The predictions are obtained by
studying structure-activity relationships in a training set
consisting of structure and biological activity data for
>300,000 organic molecules (26,27).
Briefly, a canonical molecule from PubChem was entered into the
PASS online web server. The biological activity prediction of a
7,8-DHF was then calculated. Graph visualization was performed
using GraphPad Prism version 10 software (https://www.graphpad.com/features), while the
potential molecular electrostatics were measured using Gaussian 09W
and Gaussian View 6 software (https://gaussian.com/gaussview6/). The molecular
electrostatic potential (MESP) was also predicted using Chimera
(https://www.cgl.ucsf.edu/chimerax/).
Molecular docking
In the present study, molecular docking simulation
was performed using the Keap/Nrf2 complex structure (PDB ID: 2FLU).
7,8-DHF was downloaded from PubChem (https://pubchem.ncbi.nlm.nih.gov/compound/7_8-Dihydroxyflavone)
with code CID: 1880. Molecular docking was performed using PyRx
(https://pyrx.sourceforge.io/) on a
personal computer running Windows. Protein stabilization was
performed using PyMOL by removing water atoms and introducing
hydrogen atoms. To verify the accuracy and reliability of the
molecular docking process, a redocking procedure was performed. The
original ligand was repositioned into the target binding site using
the PyRx with accurate grid coordinates. After the redocking
process, an evaluation was performed to determine the position of
the ligand. Ligands were required to have a root mean square
deviation (RMSD) of <2.0. The lattice and docking parameters
were modified according to the docking validation results. The
interaction visualization was performed using the Protein ChimeraX
(https://www.cgl.ucsf.edu/chimera/)
and Discovery Studio (https://discover.3ds.com/discovery-studio-visualizer-download)
(28,29).
Molecular dynamics (MD)
simulations
MD simulations were used to determine how 7,8-DHF
and Keap1 interact. The OpenMM engine and the AMBER force field
were used for both the protein and ligand systems. The simulations
were performed within the Google Collaboratory platform (https://colab.research.google.com/?utm_source=scs-index)
as described in a previous study (30). In the present study, the model was
subjected to thorough solvation using the TIP3P water model with
108 water molecules contained in a cubic water box of 10x10x10
units. A solution of sodium chloride was introduced at a
concentration of 150 mM. The simulation was performed using the
ff19SB force field. The system underwent 1,000 steps of
minimization followed by 20 nsec of equilibration in the
isothermal-isobaric (NPT) ensemble using a 2-femtosecond time step.
Temperatures are set at 310 Kelvin, and pressures are in the range
of 1 atmosphere. The present study focuses on the investigation of
the RMSD.
Statistical analysis
Statistical analysis in the present study was
conducted using GraphPad Prism 9 software (Dotmatics). Data are
presented as mean ± SD. Normality testing was performed using the
Shapiro-Wilk test due to the sample size being <50 subjects. The
analysis of the Nrf2, GPX-1 and MDA parameters was carried out
using a one-way ANOVA parametric test to assess the overall
efficacy of administering the 7,8-DHF treatment, followed by
post-hoc Tukey analysis to examine the differences within each
group. P-values <0.05 were considered to indicate a
statistically significant difference.
Results
Antioxidant effects of 7,8-DHF in
brain tissue of rats with ischemia-reperfusion injury
To determine the antioxidant effects of 7,8-DHF,
Nrf2 expression was examined. 7,8-DHF was found to increase the
expression of Nrf2 in dose-dependent manner. The data demonstrated
that 7,8-DHF at 20 and 50 mg/kg body weight (MCAO20 and MCAO50,
respectively) significantly increased Nrf2 expression compared with
the control, as shown in Fig. 1A
and C.
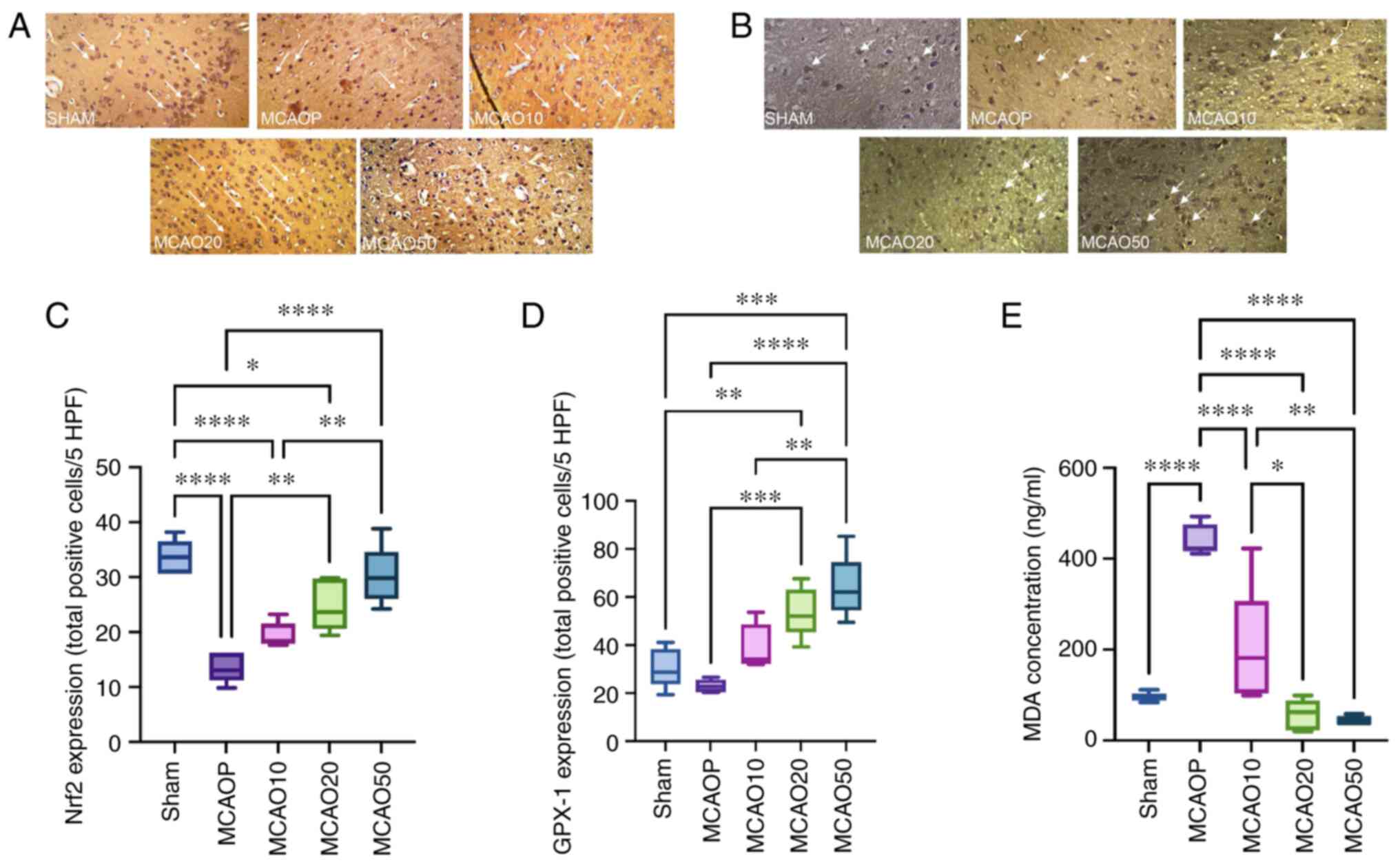 | Figure 1Antioxidant effects of 7,8-DHF in a
rat model of ischemia-reperfusion brain injury. (A) Nrf2 expression
in the brain tissue in each group. Arrows indicate Nrf2 expression.
(B) GPX-1 expression in the brain tissue in each group. Arrows
indicate GPX-1 expression (C) Statistical analysis of Nrf2
expression in each group. (D) Statistical analysis of GPX-1
expression in each group. (E) MDA level in each group.
*P≤0.05, **P≤0.01, ***P≤0.001 and
****P≤0.0001. The rat groups were as follows: Sham,
sham-operated group; MCAOP, control (MCAO induction and normal
saline); MCAO10, MCAO induction and 7,8-DHF at 10 mg/kg weight
body; MCAO20, MCAO induction and 7,8-DHF at 20 mg/kg weight body;
MCAO50, MCAO induction and 7,8-DHF at 50 mg/kg weight body.
7,8-DHF, 7,8-dihydroxyflavone; Nrf2, nuclear factor erythroid
2-related factor 2; GPX, glutathione peroxidase; MDA,
malondialdehyde; MCAO, middle cerebral artery occlusion. |
GPX-1 expression was also examined using IHC. The
results revealed a significant increase in GPX-1 expression
beginning at the dose of 20 mg/kg body weight compared with the
control. The most significant increase in GPX-1 expression was
observed in the MCAO50 group, as illustrated in Fig. 1B and D.
Furthermore, the level of MDA was lower in the
MCAO20 group than in the control (MCAOP group). The most notable
decrease level in the MDA level was found in the MCAO20 and MCAO50
groups compared with the other groups. There was no significant
difference in the MDA level between the MCAO20 and MCAO50 groups,
as shown in Fig. 1E.
ADMET and drug-likeness of
7.8-DHF
To examine the characteristics of 7,8-DHF as a drug,
ADMET, drug-likeness and toxicity analyses were performed. In drug
absorption, lipophilicity and solubility are critical molecular
characteristics. This property is critical to the drug feasibility
requirements, particularly for orally administered drugs (31). This drug feasibility test is also
known as the drug likeliness criteria. Numerous criteria have been
established, including the Lipinski criteria. A drug-like compound
that can be taken orally should have no more than one violation of
the criteria outlined by Lipinski's rule: No more than 5 hydrogen
bond donors, no more than 10 hydrogen bond acceptors, a molecular
weight of no more than 500 Da, and an octanol-water partition
coefficient (log P) of no more than 5(28). The results revealed that 7,8-DHF
fulfilled the LR5 and had a good ADMET, as presented in Table II. This indicates that the 7,8-DHF
compound has sufficient oral bioavailability to be administered
orally.
| Table IIPharmacokinetic prediction of
7,8-dihydroxyflavone. |
Table II
Pharmacokinetic prediction of
7,8-dihydroxyflavone.
| | Computationally
predicted values for the phytochemicals (ADMET profile) | |
|---|
| Pharmacokinetic
property | Model name | Reference
value | 7,8-DHF | Measurement
units |
|---|
| Lipinski rule of
5 | Molecular
weight | <500 | 254.24 | Numeric
(g/mol) |
| | Acceptor
hydrogen | <10 | 4 | Numeric (AH) |
| | Donor hydrogen | <5 | 2 | Numeric (DH) |
| | LogP | <5 | 2.87 | Numeric
(MLogP) |
| Absorption | Water
solubility | <0.4 mol/l
(low) | -3.605 | Numeric
(mol/l) |
| | | 0.4-41 mol/l
(medium) | | |
| | | >41mol/l
(high) | | |
| | Caco2
permeability | >0.90 | 1,048 | Numeric (Papp in
10-6 cm/sec) |
| | Intestinal
absorption | Low: 0-20% | 94,22 | Numeric (%
absorbed) |
| | (human) | Medium: 20-70% | | |
| | | High: 70-100% | | |
| | Skin
permeability | Likely skin
permeable: >-2.5 | -2.751 | Numeric (log
Kp) |
| | P-glycoprotein
substrate | Yes | Yes | Categorical
(yes/no) |
| | P-glycoprotein I
inhibitor | No | No | Categorical
(yes/no) |
| | P-glycoprotein II
inhibitor | No | No | Categorical
(yes/no) |
| Distribution | VDss (human) | Low: <-0.15 | 0,063 | Numeric (log
L/kg) |
| | | High >2.81 | | |
| | Fraction unbound
(human) | - | 0,146 | Numeric (Fu) |
| | BBB
permeability | Low: <0.1 | -0,057 | Numeric (log
BB) |
| | | Medium:
0.1-0.2 | | |
| | | High: >0.2 | | |
| | CNS
permeability | >-2 (penetrate
BBB) | -1,927 | Numeric (log
PS) |
| | | <-3 (Cannot
penetrate BBB) | | |
| Metabolism | CYP2D6
substrate | No | No | Categorical
(yes/no) |
| | CYP3A4
substrate | No | No | Categorical
(yes/no) |
| | CYP1A2
inhibitor | No | Yes | Categorical
(yes/no) |
| | CYP2C19
inhibitor | No | Yes | Categorical
(yes/no) |
| | CYP2C9
inhibitor | No | Yes | Categorical
(yes/no) |
| | CYP2D6
inhibitor | No | No | Categorical
(yes/no) |
| | CYP3A4
inhibitor | No | No | Categorical
(yes/no) |
| Excretion | Total
clearance | -0.002 | 0.229 | Numeric
(ml/min/kg) |
| | Renal OCT2
substrate | No | No | Categorical
(yes/no) |
| Toxicity | AMES toxicity | No | No | Categorical
(yes/no) |
| | hERG I
inhibitor | No | No | Categorical
(yes/no) |
| | hERG II
inhibitor | No | No | Categorical
(yes/no) |
| | Hepatotoxicity | No | No | Categorical
(yes/no) |
| | Skin
sensitization | No | No | Categorical
(yes/no) |
| | Tetrahymena
pyriformis | Non-toxic:
>0.5 | | |
| | toxicity | Toxic: <0.5 | 0,654 | Numeric (log
µg/l) |
| | Minnow
toxicity | Non-toxic:
>-0.3 | 1,178 | Numeric (log
mM) |
| | | Toxic:
<-0.3 | | |
| | Mutagenicity | No risk 100% | 60% | Numeric (%) |
| | | Medium risk
80% | | |
| | | High risk 60% | | |
| | Tumorigenicity | | 100% | Numeric (%) |
| | Irritating
effects | | 100% | Numeric (%) |
| | Reproductive
effects | | 100% | Numeric (%) |
7,8-DHF has a BBB score of -0.057, indicating low
permeability. BBB permeability is interpreted as logBBB >0.3,
indicating that molecules can easily pass through the BBB, whereas
molecules with logBB-1 are poorly distributed in the brain. The
difference between BBB permeability and central nervous system
(CNS) permeability lies in their respective properties. For CNS
permeability, logPS units accurately predict the compound
administered into the carotid artery. Compounds with logPS >-2
can penetrate the CNS, whereas compounds with logPS-3 cannot
penetrate the CNS. The 7,8-DHF score logPS was-1.927, indicating
that it can penetrate the CNS (Table
II). Finally, these data were confirmed with BOILED-Egg
analysis prediction using SwissADME (http://www.swissadme.ch/), demonstrating that 7,8-DHF
can penetrate the BBB, as shown in Fig. S1. As shown by the results, the
7,8-DHF compound has moderate potency based on its distribution
profile (32).
The toxicity of 7,8-DHF was also predicted using the
biocomputational approach, an important parameter for safety in the
human body. Several parameters are evaluated to determine toxicity.
The Ames test is a widely used technique to determine the mutagenic
potential of substances using bacteria (33). The Ames test used in the present
study was a prediction test using OSIRIS (https://www.organic-chemistry.org/prog/peo/). The
OSIRIS toxicity prediction model uses structural features and
substructures associated with toxic effects to determine the
potential toxicity of a chemical molecule. OSIRIS evaluates the
submitted chemical structure by comparing it to a database of
recognized toxicophores (substructures associated with toxic
effects) and structural alerts. Toxicophores refer to distinct
chemical properties that have been associated with various forms of
toxicity. OSIRIS provides toxicity predictions for several domains,
including mutagenicity, tumorigenicity, irritation, reproductive
effects and others. Each category can have different warnings
associated with it (28). The data
demonstrated that 7,8-DHF was mostly safe, although there was a
moderate risk of mutagenicity, as shown in Table II.
7,8-DHF activity and molecular
electrostatic potential prediction
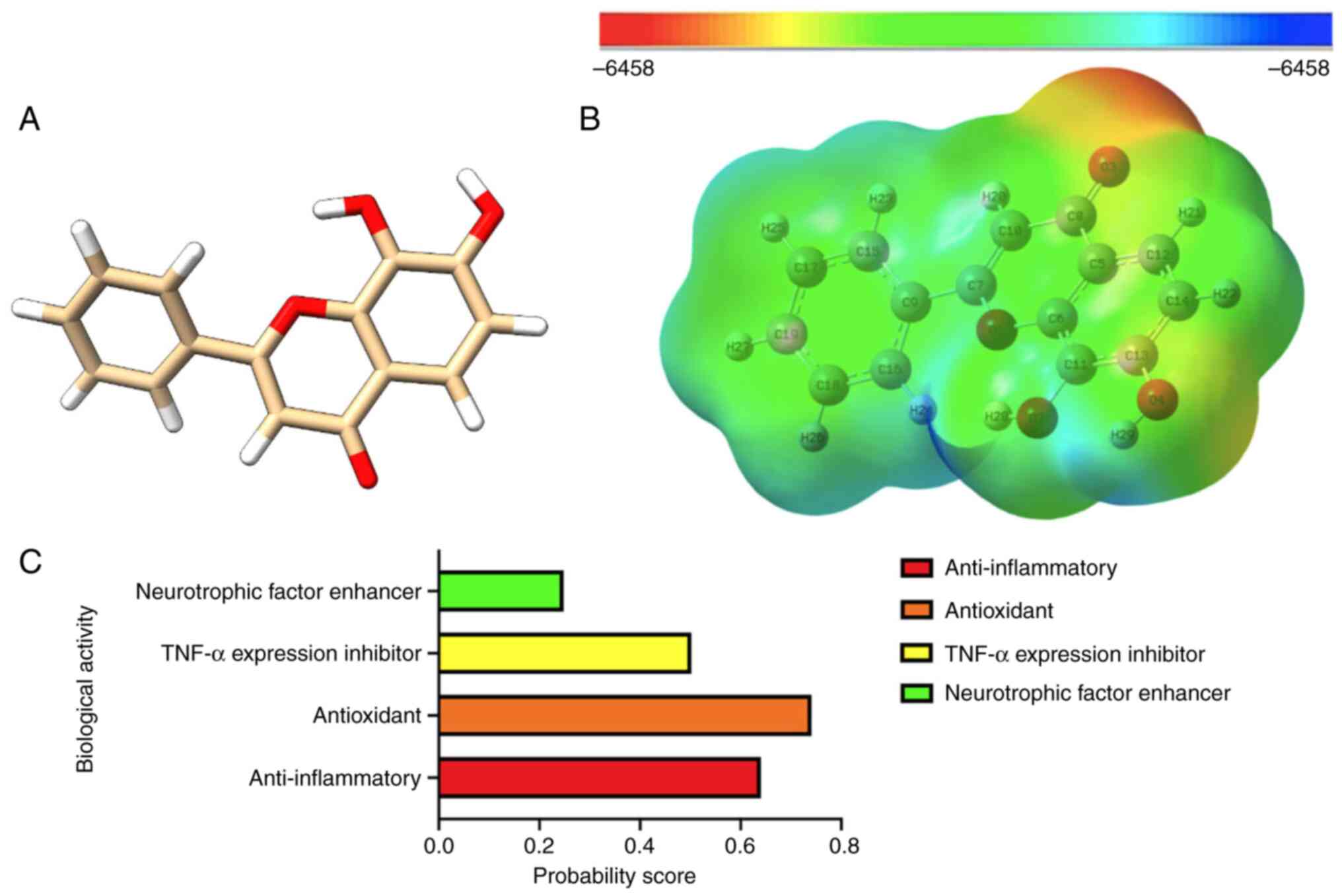
7,8-DHF has a molecular structure consisting of two
aromatic rings connected by a single rotatable bond. It has four
acceptor sites and two donor hydrogen bonds. The flavone skeleton
contains two hydroxyl groups at positions 7 and 8 with a
trans configuration. The biological activity of the molecule
is influenced by several functional groups, with the hydroxyl
groups being critical for protein-receptor interaction. The
planarity and stability of the flavone backbone are due to the
presence of a carbonyl group at position 4 and a double bond
between positions 2 and 3. A representative 3D structure of 7,8-DHF
is presented in Fig. 2A.
To determine the characteristics of the structure,
MESP was performed. The MESP surface is a tool used to identify
electrophilic and nucleophilic regions of a molecule. Different
shades on the surface represent different levels of electrostatic
potential. The ability of a molecule to bind to biomolecules can be
determined by calculating its electrophilicity index (34). Different colors represent the
electrostatic potential levels at the surface in the following
order: red-orange-yellow-green-blue. The color coding for these
maps in the identified compound varies from -6.457 (deepest red) to
+6.457 (deepest blue) (blue indicates the most electronegative and
red is the most electropositive). The most electronegative region
of 7,8-DHF in a hydrogen atom is shown in Fig. 2B.
To determine the biological activity of 7,8-DHF
based on chemical structure, the biological activity was predicted
using a PASS online web server. PASS is a computer program designed
to predict the biological activities of chemical substances. The
PASS online tool allows users to predict the pharmacological,
biological and toxicological properties of organic substances by
analyzing their chemical structures. This can be beneficial in the
drug discovery and development process (35). The biological activity prediction
of 7,8-DHF shows that it has the probability of anti-inflammatory
and antioxidant activity using PASS online. The highest predicted
score is antioxidant activity with a score of 0.741, followed by
anti-inflammatory activity with a score of 0.640, indicating that
7,8-DHF has promising anti-inflammatory and antioxidant activity.
Notably, the prediction also specified 7,8-DHF as an inhibitor of
tumor necrosis factor (TNF)-α expression with a score of 0.502, as
shown in Fig. 2C. This suggests a
possible mechanism of anti-inflammatory activity through the
inhibition of TNF-α expression. Finally, this prediction also
consists of a neurotrophic factor enhancer with a score of 0.248.
Neurotrophic factor enhancer activity is important in the
pathogenesis of stroke, which is reduced in stroke conditions. It
is important to recognize that while PASS and other similar
predictive methods are valuable for screening large databases of
chemical compounds, they have limitations. Predictions are derived
from the current information contained in the training data set,
and the accuracy of these predictions depends on the quality and
variety of the data used to develop the model. Experimental
validation is critical to verifying and improving the expected
biological activity of compounds.
Molecular docking of 7,8-DHF against
Keap1
To determine the antioxidant effect related to the
molecular prediction mechanism, molecular docking was performed by
targeting Keap1. The interaction of the Keap1/Nrf2 complex is
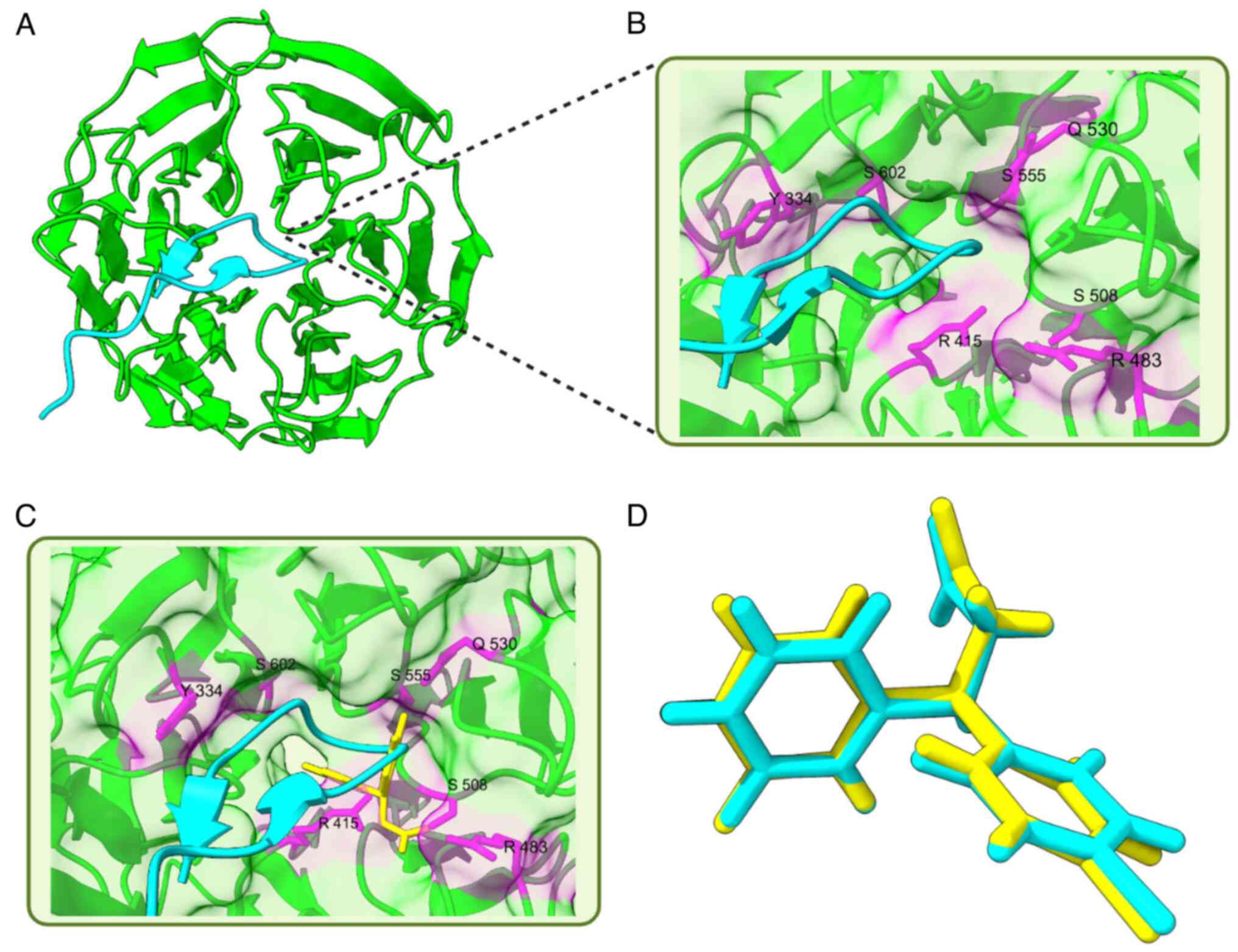
illustrated in Fig. 3A. Nrf2
appears to interact with the binding pocket of Keap1 in the middle
of the protein. There are several key amino acid residues that play
critical roles in the complex interaction between Keap1 and Nrf2.
These are tyrosine (TYR)334, arginine (ARG)415, ARG483, serine
(SER)508, glutamic acid (GLU)30, SER555 and SER602, as shown in
Fig. 3B. To clarify whether the
binding of 7,8-DHF can inhibit Keap1, a small-molecule inhibitor
[(3~{S})-3-(4-chlorophenyl)-3-(2-oxidanylidene-1~{H}-pyridin-4-yl)propanoic
acid] of Keap1 was used as a control in the present study (36). It was found that this
small-molecule inhibitor can bind to the binding pocket site of the
Keap1-Nrf2 interaction, thereby directly disrupting the Keap1/Nrf2
interaction, as shown in Fig. 3C.
Before beginning molecular docking, redocking was performed to
clarify the molecular procedure. The redocking result was good as
it had the same configuration, which has an RMSD value <2 Å, as
shown in Fig. 3D.
Following successful validation, molecular docking
simulations were performed between 7,8-DHF and Keap1 proteins,
whose value data, or ΔG (kcal/mol), are presented in Table III. The results of molecular
docking revealed that 7,8-DHF can bind to Keap1 with a binding
affinity of -7.4 Kcal/mol. This binding affinity is lower than that
of the control (-7.8 Kcal/mol). To confirm whether the binding is
similar to the control at important residues, visualization was
performed using ChimeraX and Discovery Studio. It was found that
7,8-DHF has a bond at the same site as the control ligand, as shown
in Fig. 4A. This interaction
appears to have an interaction with an important amino acid in
Keap1, as shown in Fig. 4B. The
control was found to be able bind to SER508, ARG483 and glutamine
(GLN)530 in a hydrogen bond manner; isoleucine (ILE)461,
phenylalanine (PHE)478, GLY462, glycine (GLY)509, GLY364, GLY603,
SER555 and TYR572 with van der Waals; and TYR525, alanine (ALA)556,
and ARG415 in a hydrophobic bond manner. While 7,8-DHF can bind to
the Keap1 protein via GLY364, ILE461, GLY462, ARG483, GLY509,
TYR525, SER555, TYR572, SER602 and GLY603 in a hydrogen bond
manner, while ARG415, GLN530 and ALA556 have hydrophobic bond
modes. Almost all bonds formed between 7,8-DHF and Keap1 are
similar to the control, except for SER602, as shown in Fig. 4C.
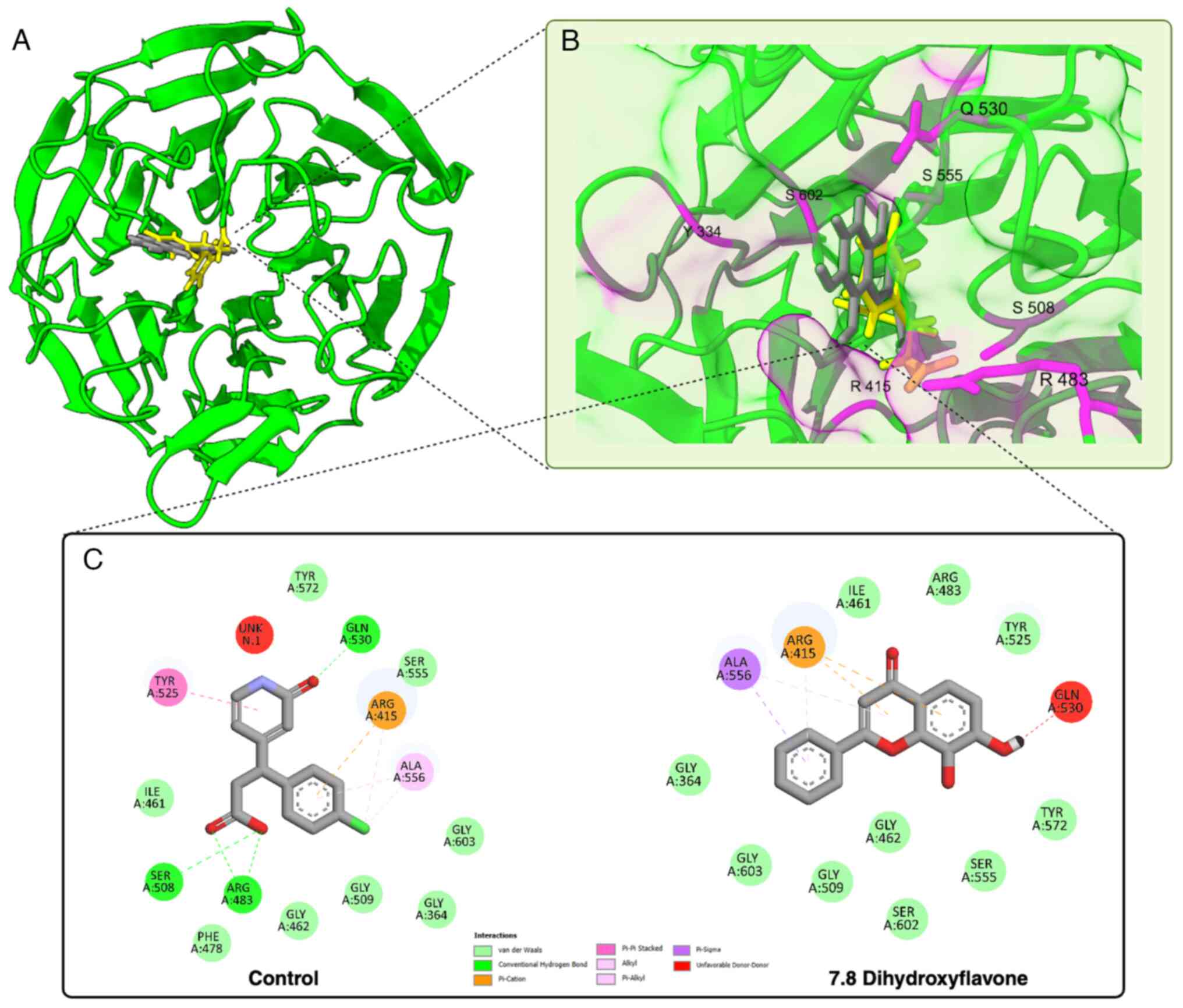 | Figure 4The binding interaction of 7,8-DHF
with Keap1. (A) Superimposed image of 7,8-DHF (grey) and control
(yellow) in Keap1. 7,8-DHF has a similar binding with the control.
(B) Detailed visualization interaction 7,8-DHF with important
residue in Keap1 protein. 7,8-DHF has near interaction with a key
residue in Keap1. (C) Comparison of the interaction of 7,8-DHF and
the control. 7,8-DHF has several similar residues in interaction
with the control. 7,8-DHF, 7,8-dihydroxyflavone; Keap1, Kelch like
ECH-associated protein 1; Nrf2, nuclear factor erythroid 2-related
factor 2. |
| Table IIIBinding affinity interaction of
7,8-DHF and Keap1. |
Table III
Binding affinity interaction of
7,8-DHF and Keap1.
| Target protein (PDB
ID) | Substance | Binding affinity
(kcal/mol) | Number of
clusters | Residue
interaction | Grid box | Dimension | RMSD | Validation (<2
Å) |
|---|
| Keap1 | 7,8-DHF | -7.4 | 6 | GLY364* | X: 16.212 | X: 21 | 1.8 | Valid |
| | | | | ARG415* | Y: 63.988 | Y: 21 | | |
| | | | | ILE461* | Z: 27.113 | Z:21 | | |
| | | | | GLY462* | | | | |
| | | | | ARG483* | | | | |
| | | | | GLY509* | | | | |
| | | | | TYR525* | | | | |
| | | | | GLN530* | | | | |
| | | | | SER555* | | | | |
| | | | | ALA556* | | | | |
| | | | | TYR572* | | | | |
| | | | | SER602 | | | | |
| | | | | GLY603* | | | | |
| | Control | -7.8 | 1 | GLY462* | | | 1.2 | Valid |
| | | | | GLY364* | | | | |
| | | | | ARG415* | | | | |
| | | | | ILE461* | | | | |
| | | | | PHE478 | | | | |
| | | | | ARG483* | | | | |
| | | | | SER508 | | | | |
| | | | | GLY509* | | | | |
| | | | | TYR525* | | | | |
| | | | | GLN530* | | | | |
| | | | | SER555* | | | | |
| | | | | ALA556* | | | | |
| | | | | TYR572* | | | | |
| | | | | GLY603* | | | | |
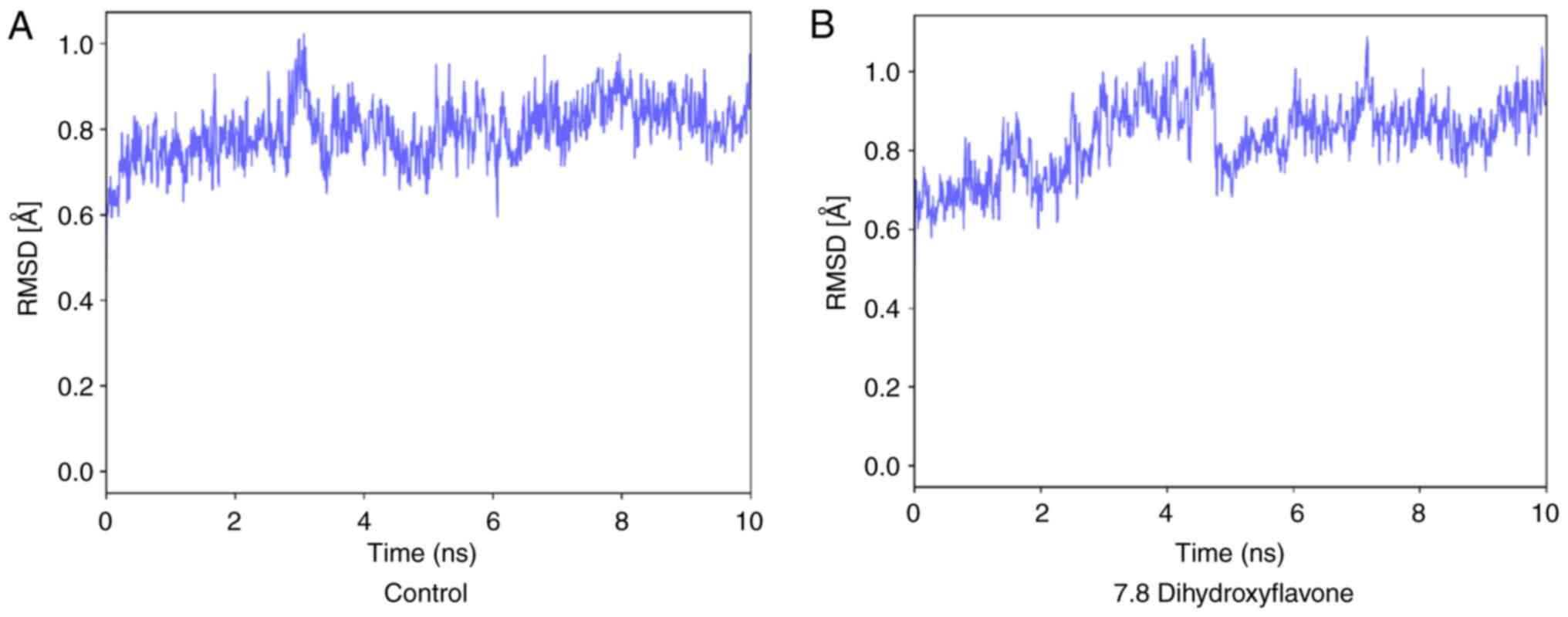
MD simulations 7,8-DHF against
Keap1
Based on the molecular dynamic analysis of 7,8-DHF
and control against Keap1. It was found that 7,8-DHF was slightly
more unstable than the control. The data revealed that the Keap1
protein complex against the control had an average RMSD fluctuation
of 0.8 angstrom (Å) with an equilibrium of 5 nsec, while 7.8 DHF
has an RMSD fluctuation from 2 to 6 ns and an equilibrium from 6 to
9, with an average value of 0.81 Å, as shown in Fig. 5. This indicates that there is no
notable difference in the stability interaction between 7,8-DHF and
the control.
Discussion
As aforementioned, the Keap1/Nrf2 complex plays a
critical role in stroke-ischemic situations by regulating cellular
defense against oxidative stress and promoting the activation of
genes involved in antioxidant and detoxification pathways, such as
superoxide dismutase (SOD), catalase (CAT) and glutathione or GST
(37,38). Under conditions of oxidative
stress, BDNF is also modulated by activated Nrf2(39). BDNF has been found to play a role
in regulating neural function, plasticity and neurogenesis in
individuals who have survived ischemic stroke (40). The data of the present study
demonstrated that the administration of 7,8-DHF increased the
expression of Nrf2. Nrf2 translocates to the nucleus and induces
the production of antioxidants. Antioxidant response elements
(AREs) are specific enhancer sequences found in gene regulatory
regions that Nrf2 targets. These AREs play a critical role in
facilitating the recruitment of key transcription factors (41). The Nrf2 protein has a significant
impact on the regulation of ~500 genes. These genes are responsible
for encoding various proteins that play key roles in redox
homeostasis, detoxification processes, stress response mechanisms
and metabolic enzyme activities (42,43).
The result of increasing Nrf2 expression will
increase the antioxidant regulatory genes such as SOD, CAT and
GPX-1, which may improve oxidative stress conditions in ischemic
stroke (44). GPX-1 plays a
crucial role as an antioxidant enzyme in reducing intracellular
hydrogen peroxide accumulation. This enzyme is ubiquitous within
cellular structures, including the cytosol, mitochondria and
certain cells. Previous studies have demonstrated that it is
superior to CAT in removing intracellular peroxides in a number of
physiological conditions (45,46).
The effects of GPx-1 on various processes involving oxidants have
been extensively studied. GPX-1 has been studied in the context of
disease and tissue injury processes, such as atherogenesis, drug
toxicity and ischemia-reperfusion injury (44).
Based on the measured MDA levels in the present
study, the administration of 7,8-DHF also improved the oxidative
stress condition, as evidenced the significant decrease in MDA
levels in the MCAO20 and MCAO50 groups. According to the study by
Wei et al (47), MDA levels
as a marker of oxidative stress are inversely proportional to BDNF
and SOD levels in patients with chronic schizophrenia, and the
expected compensation does not occur. If there is a condition of
abnormal BDNF levels accompanied by a disturbance in redox
homeostasis (oxidative stress), this condition will also cause
disruptions in neurotrophic processes and neuron development
(47). From the aforementioned
result, the dose of 50 mg/kg body weight 7,8-DHF was the optimal
dose among all parameters. Thus, 7,8-DHF has antioxidant activity
by increasing Nrf2 and GPX-1 expression, while decreasing MDA
levels.
The prediction of biological activity was in
agreement with the experimental result. This prediction was also in
agreement with the findings of previous studies. The study by Park
et al (15) demonstrated
that 7,8-DHF exhibited anti-inflammatory properties via the
downregulation of the NF-κB and MAPK pathways in
lipopolysaccharide-treated RAW264.7 cells. In addition, 7,8-DHF
functions as a potent TrkB agonist and a potent antioxidant through
direct free radical scavenging and increased SOD activity. It
significantly inhibits 6-hydroxydopamine-induced oxidative damage,
preventing cell death, apoptosis and mitochondrial dysfunction
(48).
Molecular docking revealed that 7,8-DHF could bind
to the Keap1 protein. 7,8-DHF can bind to key amino acid residues
in Keap 1. 7,8-DHF had amino acid residue similarity with the
control (>80%), indicating it has a biological function
(49). The molecular dynamic
result indicates that there is no notable difference in stability
interaction between 7,8-DHF and control, suggesting that it has an
equal stability interaction with Keap 1. It was expected that
7,8-DHF may function as a Keap1 inhibitor similar to the
control.
7,8-DHF has been shown to function as a TrkB agonist
(16). It activates TrkB receptors
and induces downstream signaling pathways that promote cell
survival, growth, differentiation and plasticity. Additionally,
7,8-DHF has demonstrated neuroprotective properties and has been
found to restore motor function deficits in a mouse model of
Parkinson's disease (11,19,48).
7,8-DHF has also been reported to upregulate heme oxygenase 1
(HO-1) to protect PC12 cells against
1-methyl-4-phenylpyridinium-induced cytotoxicity (48). The data of the present study
demonstrated that 7,8-DHF administration increased GPX-1
expression, thus contributing to decreased oxidation processes in
pathological conditions. This supported the data demonstrating that
it also upregulated GPX-1. Thus, it is possible that 7,8-DHF
regulates the antioxidant mechanism at the upper site of GPX-1 and
HO-1. Thus, it is suggested that a mechanism responsible for the
antioxidant effects of 7,8-DHF may be the inhibition of Keap1,
which allows the translocation of Nrf2 to induce antioxidant
production, as shown in Fig.
6.
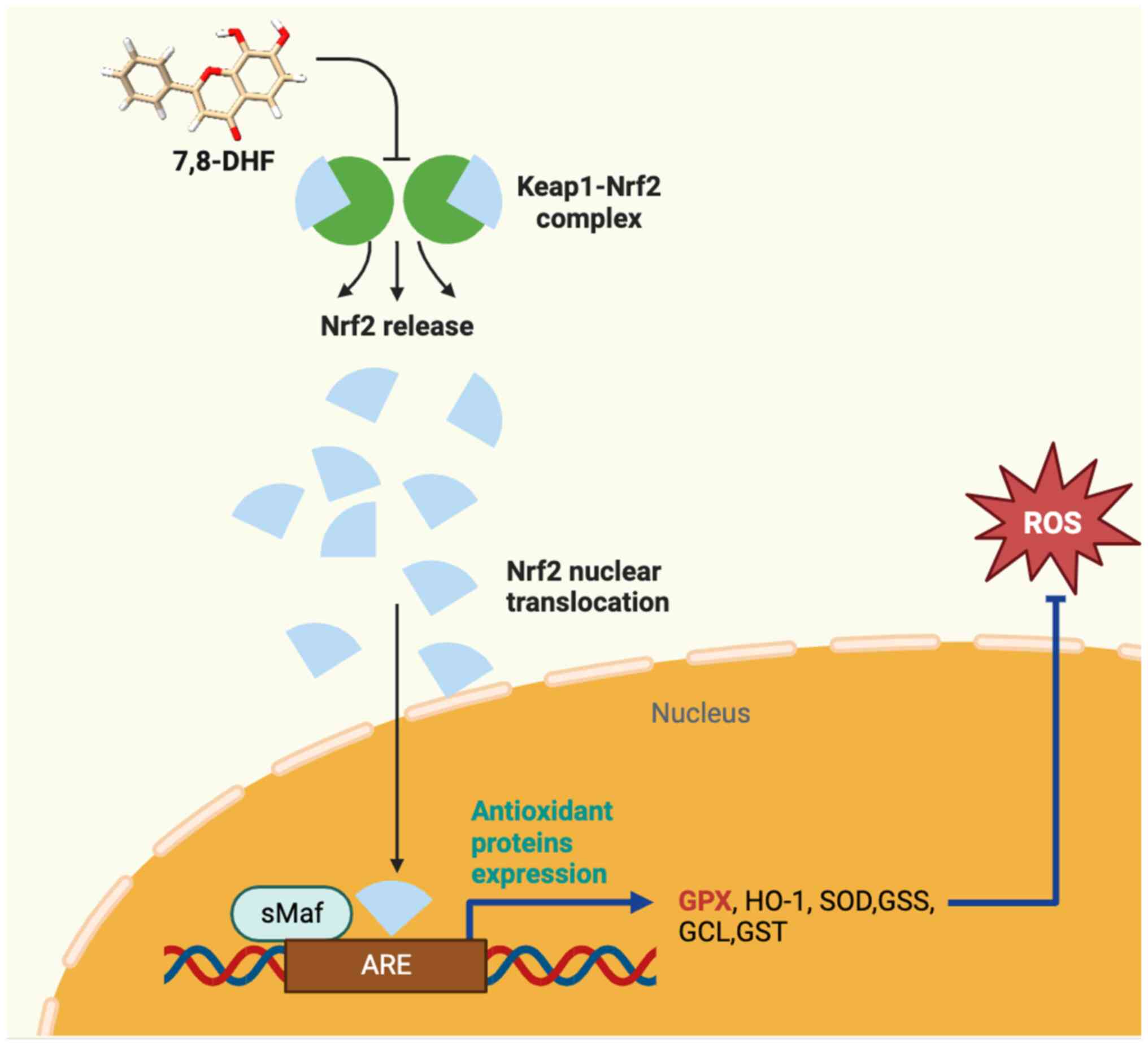 | Figure 6Proposed mechanism of 7,8-DHF as an
antioxidant in ischemic reperfusion brain injury rat model. 7,8-DHF
inhibits the Keap1 interaction, thereby stabilizing Nrf2, which
accumulates in the cytoplasm and then translocates to the nucleus.
Nrf2 interacts with the ARE, which then activates the transcription
of genes involved in antioxidant defense, detoxification, and cell
protection, such as HO-1, GPX, SOD and various phase II
detoxification enzymes. 7,8-DHF, 7,8-dihydroxyflavone; Keap1, Kelch
like ECH-associated protein 1; Nrf2, nuclear factor erythroid
2-related factor 2; ARE, antioxidant response element; HO-1, heme
oxygenase-1; GPX, glutathione peroxidase; SOD, superoxide
dismutase; GST, glutathione S-transferase; GCL, glutamate-cysteine
ligase; GSS, glutathione synthetase. |
A limitation of the present study is that the
analyses performed were not sufficient to judge that 7,8-DHF can
directly inhibit the interaction of the Keap1/Nrf2 complex; thus,
further studies are warranted to clarify the underlying mechanisms
in a more in-depth manner. However, the present study demonstrated
that 7,8-DHF increases Nrf2 and GPX-1 expression, while decreasing
the MDA levels. Molecular docking elucidated the possible
mechanisms and interactions that occurred in the in vivo
results. Further follow-up studies, such as the experimental
identification of interactions and the visualization of the model
using X-ray or cryo-EM, are required to clarify these results and
confirm the potential of 7,8-DHF as a novel therapy for ischemic
stroke through the inhibition of Keap1. In addition, experimental
toxicity studies are required to further investigate the safety of
7,8-DHF.
In conclusion, from the findings presented herein,
it can be concluded that 7,8-DHF has antioxidant activity through
the Keap1 inhibition, thereby increasing Nrf2 expression. 7,8-DHF
can decrease the levels of MDA. It can also increase the expression
of GPX-1 in brain tissue. Molecular docking also revealed a
promising result; 7,8-DHF can inhibit the interaction of the
Keap1/Nrf2 complex. However, further studies are warranted to
clarify the mechanisms of 7,8-DHF in ameliorating other biomarkers
of stroke. Therefore, further more advanced studies on 7,8-DHF are
required to clarify the findings of the present study.
Supplementary Material
Result of BOILED-Egg analysis of
7,8-DHF. 7,8-DHF, 7,8-dihydroxyflavone; BBB, blood-brain barrier;
HIA, human intestinal absorption; PGP+, active efflux by
P-glycoprotein (P-gp); PGP., non-substrate of P-gp.
Acknowledgements
The authors are grateful to the Medical Faculty of
Brawijaya University Animal Laboratory staff for providing
technical assistance.
Funding
Funding: The present study was supported by the Faculty of
Medicine at Brawijaya University (grant no. 3392.1/Uni 0.F08/PN
l2O2O).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YY and MFRS were involved in the conception and
design of the study, in data collection and analysis, and in the
writing, revising and reviewing of the manuscript. DBK, AAA, DMA,
YNS, RFM and BA were involved in data collection and analysis, and
in the writing of the manuscript. AHV, SDA, AH, EN, YF, AM, KAS,
URZ, DYP and SU were involved in data analysis, and in the writing,
revising and reviewing of the manuscript. YY and MFRS confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was conducted by following the
University of Brawijaya animal ethics guidelines and was approved
by the Ethics Committee (No. 160/EC/KEPK/09/2020), Faculty of
Medicine, Brawijaya University, Malang, Indonesia.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, AI tools
(QuillBot Premium; https://quillbot.com/) were used to improve the
readability and language of the manuscript or to generate images,
and subsequently, the authors revised and edited the content
produced by the AI tools as necessary, taking full responsibility
for the ultimate content of the present manuscript.
References
|
1
|
Lin SY, Wang YY, Chang CY, Wu CC, Chen WY,
Liao SL and Chen CJ: TNF-α receptor inhibitor alleviates metabolic
and inflammatory changes in a rat model of ischemic stroke.
Antioxidants (Basel). 10(851)2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Li Z, Bi R, Sun S, Chen S, Chen J, Hu B
and Jin H: The role of oxidative stress in acute ischemic
stroke-related thrombosis. Oxid Med Cell Longev.
2022(8418820)2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kurniawan DB, Rizki Syaban MF, Mufidah A,
Rafsi Zulfikri MU and Riawan W: Protective effect of Saccharomyces
cerevisiae in Rattus norvegicus Ischemic Stroke Model. Res J
Pharmacy Technol. 5785–5789. 2021.
|
|
4
|
Qian CL and Fan R: Effect of Pingchuan
Guben decoction on patients with chronic obstructive pulmonary
disease: Results from a randomized comparative effectiveness
research trial. Exp Ther Med. 14:3915–3925. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tran KT, Pallesen JS, Solbak SMØ,
Narayanan D, Baig A, Zang J, Aguayo-Orozco A, Carmona RMC, Garcia
AD and Bach A: A Comparative assessment study of known
small-molecule Keap1-Nrf2 Protein-Protein interaction inhibitors:
Chemical synthesis, binding properties, and cellular activity. J
Med Chem. 62:8028–8052. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
B Gowda SG, Fuda H, Tsukui T, Chiba H and
Hui SP: Discovery of eicosapentaenoic acid esters of hydroxy fatty
acids as potent Nrf2 activators. Antioxidants (Basel).
9(397)2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lee KH, Cha M and Lee BH: Neuroprotective
Effect of Antioxidants in the Brain. Int J Mol Sci.
21(7152)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
González-Bosch C, Boorman E, Zunszain PA
and Mann GE: Short-chain fatty acids as modulators of redox
signaling in health and disease. Redox Biol.
47(102165)2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hardingham GE and Lipton SA: Regulation of
neuronal oxidative and nitrosative stress by endogenous protective
pathways and disease processes. Antioxid Redox Signal.
14:1421–1424. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sokolov AV, Dubrovskaya NM, Kostevich VA,
Vasilev DS, Voynova IV, Zakharova ET, Runova OL, Semak IV, Budevich
AI, Nalivaeva NN and Vasilyev VB: Lactoferrin induces
erythropoietin synthesis and rescues cognitive functions in the
offspring of rats subjected to prenatal hypoxia. Nutrients.
14(1399)2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lotfi MS and Kalalinia F: Flavonoids in
Combination with stem cells for the treatment of neurological
disorders. Neurochem Res. 48:3270–3282. 2023.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Xu DP, Li Y, Meng X, Zhou T, Zhou Y, Zheng
J, Zhang JJ and Li HB: Natural antioxidants in foods and medicinal
plants: Extraction, assessment and resources. Int J Mol Sci.
18(96)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Aldughaylibi FS, Raza MA, Naeem S, Naeem
S, Rafi H, Alam MW, Souayeh B, Farhan M, Aamir M, Zaidi N and Mir
TA: Extraction of bioactive compounds for antioxidant,
antimicrobial, and antidiabetic applications. Molecules.
27(5935)2022.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Laksmitawati DR, Prasanti AP, Larasinta N,
Agitha Syauta G, Hilda R, Ramadaniati HU, Widyastuti A, Karami N,
Afni M, Rihibiha DD, et al: Anti-inflammatory potential of
Gandarusa (Gendarussa vulgaris Nees) and Soursoup (Annona
muricata L) extracts in LPS stimulated-macrophage cell
(RAW264.7). J Natural Remedies. 16:73–81. 2016.
|
|
15
|
Park HY, Kim GY, Hyun JW, Hwang HJ, Kim
ND, Kim BW and Choi YH: 7,8-Dihydroxyflavone exhibits
anti-inflammatory properties by downregulating the NF-κB and MAPK
signaling pathways in lipopolysaccharide-treated RAW264.7 cells.
Int J Mol Med. 29:1146–1152. 2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Liu C, Chan CB and Ye K:
7,8-dihydroxyflavone, a small molecular TrkB agonist, is useful for
treating various BDNF-implicated human disorders. Transl
Neurodegener. 5(2)2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Chen J, Chua KW, Chua CC, Yu H, Pei A,
Chua BH, Hamdy RC, Xu X and Liu CF: Antioxidant activity of
7,8-dihydroxyflavone provides neuroprotection against
glutamate-induced toxicity. Neurosci Lett. 499:181–185.
2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kozics K, Valovicova Z and Slamenova D:
Structure of flavonoids influences the degree inhibition of
Benzo(a)pyrene-induced DNA damage and micronuclei in HepG2 cells.
Neoplasma. 58:516–524. 2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jin W: Regulation of BDNF-TrkB signaling
and potential therapeutic strategies for Parkinson's disease. J
Clin Med. 9(257)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang R, Kang KA, Piao MJ, Ko DO, Wang ZH,
Chang WY, You HJ, Lee IK, Kim BJ, Kang SS and Hyun JW: Preventive
effect of 7,8-dihydroxyflavone against oxidative stress induced
genotoxicity. Biol Pharm Bull. 32:166–171. 2009.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rupadevi M, Parasuraman S and Raveendran
R: Protocol for middle cerebral artery occlusion by an intraluminal
suture method. J Pharmacol Pharmacother. 2:36–39. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Xu W, Zhang Y, Su J, Liu AF, Wang K, Li C,
Liu YE, Zhang YQ, Lv J and Jiang W: Ischemia reperfusion injury
after gradual versus rapid flow restoration for middle cerebral
artery occlusion rats. Sci Rep. 8(1638)2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
García-Díaz Barriga G, Giralt A,
Anglada-Huguet M, Gaja-Capdevila N, Orlandi JG, Soriano J, Canals
JM and Alberch J: 7,8-dihydroxyflavone ameliorates cognitive and
motor deficits in a Huntington's disease mouse model through
specific activation of the PLCγ1 pathway. Hum Mol Genet.
26:3144–3160. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Syaban MFR, Muhammad RF, Adnani B, Ami
Putra GF, Arvina SD, Budi D, Erwan NE and Krisnayana AD: Molecular
docking studies of interaction curcumin against Beta-secretase 1,
Amyloid A4 Protein, Gamma-secretase and Glycogen Synthase Kinase-3β
as Target Therapy for Alzheimer Disease. Res J Pharmacy Technol.
15(3074)2022.
|
|
25
|
Daina A, Michielin O and Zoete V:
SwissADME: A free web tool to evaluate pharmacokinetics,
drug-likeness and medicinal chemistry friendliness of small
molecules. Sci Rep. 7(42717)2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Filimonov DA, Lagunin AA, Gloriozova TA,
Rudik AV, Druzhilovskii DS, Pogodin PV and Poroikov VV: Prediction
of the biological activity spectra of organic compounds using the
pass online web resource. Chem Heterocycl Comp. 50:444–457.
2014.
|
|
27
|
Syaban MFR, Faratisha IFD, Yunita KC,
Erwan NE, Kurniawan DB and Putra GFA: Molecular docking and
interaction analysis of propolis compounds against SARS-CoV-2
receptor: Propolis compound against SARS-CoV-2 receptor. J Tropical
Life Sci. 12:219–230. 2022.
|
|
28
|
Arviana SD, Yueniwati Y, Rahayu M and
Syaban MFR: 7,8-dihydroxyflavone as a neuroprotective agent in
ischemic stroke through the regulation of HIF-1α protein. Res J
Pharmacy Technol. 15:3980–3986. 2022.
|
|
29
|
Meng EC, Goddard TD, Pettersen EF, Couch
GS, Pearson ZJ, Morris JH and Ferrin TE: UCSF ChimeraX: Tools for
structure building and analysis. Protein Sci.
32(e4792)2023.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Santoso W, Putra G, Syaban M, Vadhana R,
Khamid J, Maududi I and Sujuti H: Integrated molecular docking,
dynamic simulations and in vivo analysis of ethanol extract
Citrus sinensis peel as an antioxidant and neurotrophic
agent for ameliorating motor and cognitive functions in traumatic
brain injury. World Acad Sci J. 5(29)2023.
|
|
31
|
Ugale VG and Bari SB: Structural
Exploration of Quinazolin-4(3H)-ones as Anticonvulsants: Rational
Design, Synthesis, Pharmacological Evaluation, and Molecular
Docking Studies. Arch Pharm (Weinheim). 349:864–880.
2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Daina A and Zoete V: A BOILED-Egg to
predict gastrointestinal absorption and brain penetration of small
molecules. ChemMedChem. 11:1117–1121. 2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zeiger E: The test that changed the world:
The Ames test and the regulation of chemicals. Mutat Res Genet
Toxicol Environ Mutagen. 841:43–48. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Vetrivel R, Deka RC, Chatterjee A, Kubo M,
Broclawik E and Miyamoto A: Studies on the molecular electrostatic
potential inside the microporous material and its relevance to
their catalytic activity. In: Theoretical and Computational
Chemistry. Vol 3. Murray JS and Sen K (eds). Elsevier, Amsterdam.
pp509-541, 1996.
|
|
35
|
Parasuraman S: Prediction of activity
spectra for substances. J Pharmacol Pharmacother.
2(5253)2011.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Heightman TD, Callahan JF, Chiarparin E,
Coyle JE, Griffiths-Jones C, Lakdawala AS, McMenamin R, Mortenson
PN, Norton D, Peakman TM, et al: Structure-activity and
structure-conformation relationships of Aryl propionic acid
inhibitors of the Kelch-like ECH-associated protein 1/nuclear
factor erythroid 2-related factor 2 (KEAP1/NRF2) protein-protein
interaction. J Med Chem. 62:4683–4702. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Li N, Pang Q, Zhang Y, Lin J, Li H, Li Z,
Liu Y, Fang X, An Y, Bai H, et al: Ginsenoside compound K reduces
neuronal damage and improves neuronal synaptic dysfunction by
targeting Aβ. Front Pharmacol. 14(1103012)2023.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Frandsen JR and Narayanasamy P:
Neuroprotection through flavonoid: Enhancement of the glyoxalase
pathway. Redox Biol. 14:465–473. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Sivandzade F, Prasad S, Bhalerao A and
Cucullo L: NRF2 and NF-қB interplay in cerebrovascular and
neurodegenerative disorders: Molecular mechanisms and possible
therapeutic approaches. Redox Biol. 21(101059)2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Handayani F, Setyowati S, Pudjonarko D and
Sawitri DR: The Effect of ‘SELF-HELP Packages’ on post stroke
depression among ischemic stroke survivors. Nurse Media J Nursing.
10:361–375. 2020.
|
|
41
|
Suzuki T and Yamamoto M: Molecular basis
of the Keap1-Nrf2 system. Free Radic Biol Med. 88:93–100.
2015.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hahn ME, Timme-Laragy AR, Karchner SI and
Stegeman JJ: Nrf2 and Nrf2-related proteins in development and
developmental toxicity: Insights from studies in zebrafish (Danio
rerio). Free Radic Biol Med. 88:275–289. 2015.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Bellezza I, Giambanco I, Minelli A and
Donato R: Nrf2-Keap1 signaling in oxidative and reductive stress.
Biochim Biophys Acta Mol Cell Res. 1865:721–733. 2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Lubos E, Loscalzo J and Handy DE:
Glutathione peroxidase-1 in Health and Disease: From molecular
mechanisms to therapeutic opportunities. Antioxid Redox Signal.
15:1957–1997. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Antunes F, Han D and Cadenas E: Relative
contributions of heart mitochondria glutathione peroxidase and
catalase to H(2)O(2) detoxification in in vivo conditions. Free
Radic Biol Med. 33:1260–1267. 2002.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Cohen G and Hochstein P: Glutathione
peroxidase: The primary agent for the elimination of hydrogen
peroxide in erythrocytes. Biochemistry. 2:1420–1428.
1963.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wei C, Sun Y, Chen N, Chen S, Xiu M and
Zhang X: Interaction of oxidative stress and BDNF on executive
dysfunction in patients with chronic schizophrenia.
Psychoneuroendocrinology. 111(104473)2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Song Y and Han X: 7,8-Dihydroxyflavone
Protects PC12 Cells against MPP+-Induced Cytotoxicity by Heme
Oxygenase-1. ijSciences. 8:55–59. 2019.
|
|
49
|
Martin YC, Kofron JL and Traphagen LM: Do
structurally similar molecules have similar biological activity? J
Med Chem. 45:4350–4358. 2002.PubMed/NCBI View Article : Google Scholar
|