Introduction
Diabetes mellitus (DM) is a chronic metabolic
disorder in which the pancreas produces insufficient or no insulin
to match the body's demands (type 1 DM), or in which the body is
unable to effectively utilize the insulin produced (type 2 DM)
(1). The prevalence of diabetes is
increasing to the extent that the number of adults with diabetes
has been estimated to be 592 million by 2035 (2–5). DM is
linked with oxidative stress occurring as a consequence of
increased formation of free radicals, including superoxide
(O2−) and hydroxyl (OH) radicals, and decreased activity
of antioxidant defense systems (6).
Hyperglycemia increases the formation of reactive oxygen species
(ROS) via several pathways including glucose autoxidation, the
polyol pathway and non-enzymatic protein glycation (7). Free radicals can adversely affect
important biomolecules including carbohydrates, proteins and DNA
(8). Oxidative damage to various
brain regions results in long-term complications, morphological
abnormalities and memory impairments (9). Emerging evidence suggests that oxidative
damage associated with DM may negatively impact the central nervous
system (CNS), causing cognitive impairment in addition to
complications in the peripheral nervous system (2). In the CNS, the hippocampus is considered
a particular target for diabetes-related changes (10). The hippocampus is a component of the
limbic system of the brain and considered as an integration center
for cognitive functions including learning and memory (11). Diabetes has been associated with
cognitive and memory impairments, indicating that the hippocampus
may be affected by the disease (12).
Apoptosis may be considered as a pathway for hyperglycemia-induced
hippocampal neuronal cell death (13). DNA fragmentation, cell shrinkage and
nuclei membrane blebbing are morphological characteristics of
apoptosis (14). The neuroprotective
effect of antioxidants has been indicated in the treatment of
experimental neurodegenerative animal models (5). Consumption of natural antioxidants
reportedly reduces the risk of cancer, cardiovascular disease and
diabetes, among other diseases (15).
Lycopene, which naturally occurs in tomato and other fruits
including papaya, pink guava and watermelon, has long been known to
have potential health-promoting properties (16). Among naturally-occurring carotenoids,
lycopene appears to best scavenge free radicals (17). Lycopene may also penetrate the
blood-brain barrier and prevail in the CNS (18). The purpose of the present study was to
investigate the effect of lycopene, insulin and their co-treatment
in preventing apoptosis, on the levels of total antioxidant
capacity (TAC) and malondialdehyde activity (MDA), within the
hippocampus of streptozotocin (STZ)-induced diabetic rats.
Materials and methods
Animals and study design
A total of 48 adult male Wistar rats (weighing
200–250 g, aged 8–10 weeks old) were purchased from the
Baqiyatallah University of Medical Sciences, Tehran, Iran. The
animals were housed under standard laboratory conditions (12-h
light/dark cycle, room temperature of 21–22°C and 45–55% humidity)
with ad libitum access to food and tap water. The
experimental protocols were reviewed and approved by the
Institutional Animal Ethics Committee of Zanjan University of
Medical Sciences (ZUMS), Zanjan, Iran (approval no.
ZUMS.REC.1394.29), and conducted in accordance with the ethical
guidelines approved by the Institutional Animal Ethics Committee of
ZUMS (IAEC no. 03/028/07). An overnight fast was followed by a
single dose of 60 mg/kg STZ (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) prepared in citrate buffer (pH 4.4; 0.1 M), injected
intraperitoneally (i.p.) to induce diabetes (19). Blood samples (0.2–0.5 ml) were
collected through the tail vein, and glucose levels measured using
a glucometer (Accu-Chek; Roche Diagnostics GmbH, Mannheim,
Germany); diabetes was confirmed 72 h after STZ injection. Animals
with fasting blood-glucose levels >250 mg/dl (20) were selected and used for the present
study. Body weights and blood-glucose levels were determined prior
to the experiment and at the end of the experiment to evaluate the
effects of lycopene and insulin. Rats were randomly divided into
six groups, with each group consisting of 8 animals. The first
group consisted of non-diabetic control animals. The second group
served as the control group treated with lycopene [4 mg/kg/day
per os (p.o.); lycopene was dissolved in double distilled
water following trituration with 5% Tween-80] (21). The third and fourth groups were the
diabetic control and diabetic animals treated with lycopene (4
mg/kg/day p.o.), respectively. The fifth group was the diabetic
group treated with insulin (1 to 2 units per day, i.p.), and the
sixth group comprised of diabetic animals administered lycopene (4
mg/kg p.o.) and insulin (1–2 units a day, i.p.) simultaneously. On
the third day of administration, the control and diabetic control
groups received 0.5 ml normal saline while the other groups
continued to receive insulin and/or lycopene. Drug administration
continued for 8 weeks and at the end of the eighth week, learning
and memory were evaluated for two days using the shuttle box test.
The animals under deep anesthesia were sacrificed; blood samples
(0.2–0.5 ml) were collected through the tail vein, and
blood-glucose levels were measured. The hippocampi were isolated
following rapid removal of the brains. To perform a terminal
deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay
and acridine orange (AO) staining, left hippocampi were fixed in
10% formalin, and right hippocampi were stored in cryotubes at
−70°C in order to assess TAC and MDA.
Passive avoidance task
The behavioral experiments were conducted using a
two-way shuttle box system (Borj Sanaat Co., Tehran, Iran). The
apparatus and procedure were as described previously (22). In brief, the step-through passive
avoidance apparatus consisted of a light chamber (27×14.5×14
cm3) made of transparent plastic and a dark chamber
(27×14.5×14 cm3) made of dark opaque plastic. The floors
of both chambers were made of stainless steel rods (3-mm diameter)
spaced 1 cm apart. The floor of the dark chamber could be
electrified using a shock generator. A rectangular opening (6×8
cm2) was located between the two chambers and could be
closed by an opaque guillotine door.
First, all experimental groups received two trials
to habituate and acclimatize the rats to the apparatus. For these
trials, the rats were placed in the light compartment of the
apparatus facing away from the door, and 10 sec later, the
guillotine door was raised. Upon the rat entering the dark
compartment, the door was closed, and after 30 sec, the rats were
taken from the dark compartment and placed in their home cage. The
habituation trial was repeated after 30 min and followed after the
same interval by the first acquisition trial. The entrance latency
to the dark compartment (step through latency, STLa) was recorded
when the animal had placed all four paws in the dark compartment.
For the training of the animals, as soon as they had spontaneously
entered into the dark compartment, the guillotine door was lowered,
and a mild electrical shock (0.5 mA) was applied for 3 sec; after
30 sec, the rat was returned to its home cage. Then, after 2 min,
the procedure was repeated. The rat received a foot-shock each time
it reentered the dark compartment with all four paws placed in the
compartment; training was terminated when the rat remained in the
light compartment for 120 sec. The number of trials (entries into
the dark chamber) were recorded. Long-term memory was tested within
24 h after the passive avoidance learning (PAL) acquisition trial.
The rats were placed in the lighted chamber as in PAL training
session, and 10 sec later, the guillotine door was raised, and the
step-through latency (STLr) and the time spent in the dark
compartment (TDC) were recorded for up to 600 sec. If the rat did
not enter the dark compartment within 600 sec, the retention test
was terminated, and a ceiling score of 600 sec was assigned. During
this session, the electric shocks were not applied to the grid
floor.
Tissue preparation
The hippocampus samples obtained from rats were
immersed in 10% formalin for 72 h to allow fixation at room
temperature. A slow step-wise dehydration was performed for tissue
processing by adding a series of ethanol solutions (50–100%) of
increasing concentration. Samples were washed with xylene prior to
the addition of each new alcohol solution. Following dehydration,
the hippocampal samples were embedded in melt paraffin. Sectioning
was performed using a rotary microtome, from which slices of 5 µm
thickness were obtained. The slices were mounted on microscope
slides and any paraffin adhering to the mounted sections was
dissolved by passive clearance with chloroform.
TUNEL assay
In-situ DNA fragmentation was visualized by the
TUNEL method according to a previous study (23) using an in situ cell death detection
POD kit (Roche Diagnostics GmbH) (24). Briefly, dewaxed tissue sections were
predigested with proteinase K (20 mg/ml in 10 mmol/l Tris-HCl, pH
7.6) for 30 min at room temperature, then rinsed three times with
phosphate-buffered saline solution (PBS). Subsequently, treatment
with 3% H2O2 for 10 min at room temperature
was used to block endogenous peroxidase activity, and the sections
were incubated with the TUNEL reaction mixture for 60 min at 37°C.
The slides were then rinsed three times with PBS and incubated with
convertor peroxidase for 30 min in a humidified chamber at room
temperature. Following three washes with PBS, diaminobenzidine
chromogenic substrate (Roche Diagnostics GmbH) was added to
sections for 10 min at room temperature. The sections were
counterstained with hematoxylin for 30 sec at room temperature. The
number of TUNEL-positive cells in the hippocampal region were
counted under a light microscope (Olympus BX51; Olympus
Corporation, Tokyo, Japan) by double-blinded observation and
analyzed with Digital Imaging Solution Cell Software version 1.1
(Olympus Soft Imaging Solutions GmbH, Münster, Germany).
AO staining
AO is a metachromatic fluorescence probe which is
used to demonstrate the degree of nuclear DNA susceptibility to
diabetes-induced denaturation by distinguishing between native
double-stranded DNA (green fluorescence) and denatured
single-stranded DNA (red fluorescence) (5). Samples were washed with distilled water
(5 min) and PBS (5 min) and then stained with freshly prepared AO
(0.19 mg/ml in McIlvain phosphate-citrate buffer, pH 4.0) for 20
min, and finally counterstained with hematoxylin for 5 min, all at
room temperature. To measure the percentage of dead cells, samples
were assessed on the same day using a fluorescent microscope
(Olympus BX51). A total of 8 samples from each group were selected
randomly. For each sample, 5 sections, and in each section, 5
regions [CA1, CA2, CA3, CA4 and dentate gyrus (DG)], were evaluated
at random. The total number of cells and the number of dead cells
were determined in each region (23).
TAC
The hippocampus samples obtained from rats were
lysed by adding 1 ml lysis buffer (154 mM NH4Cl, 10 mM
KHCO3, 0.1 mM EDTA, pH 7.4). Samples were then
homogenized with a sterile syringe and homogenizer. The samples
were centrifuged at 7,000 × g for 20 min at room temperature, and
the supernatants were collected and aliquoted. Samples were stored
at −20°C prior to assessment of TAC and MDA. The TAC assay was
performed using a TAC kit (Abcam, Cambridge, MA, USA) and the
method described by Prieto et al (25). Different concentrations of extracts
(0.50, 0.75, 1.00 and 1.50 mg/ml) and ascorbic acid as standard
were added to 3 ml reagent solution (0.6 M
H2SO4, 28 mM Na2HPO4
and 4 mM ammonium molybdate). The samples were incubated for 90 min
at 25°C following shaking, and centrifuged at 700 × g for 10 min at
room temperature. The absorbance of the supernatant was then
determined at 490 nm with an ELISA plate reader. Distilled water (1
ml) was used as blank, processed in the same way. TAC was expressed
as ascorbic acid equivalent per dry weight of extract.
MDA assay
MDA content, as a measure of lipid peroxidation, was
assayed in the form of thiobarbituric acid-reactive substances
(TBARS) by the method of Wills (26)
using a lipid peroxidation MDA assay kit (Abcam). Briefly, 0.5 ml
post mitochondrial supernatant and 0.5 ml Tris HCl were incubated
at 37°C for 2 h. Following incubation, 1 ml 10% trichloroacetic
acid was added and centrifuged at 1,000 × g for 10 min at room
temperature. To 1 ml of supernatant, 1 ml 0.67% thiobarbituric acid
was added and the tubes were retained in boiling water for 10 min.
Following cooling, 1 ml double distilled water was added and
absorbance was measured at 535 nm with an ELISA plate reader. TBARS
were quantified using an extinction coefficient of
1.56×105 M−1cm−1. Tissue protein
was estimated using the Biuret method and the brain malondialdehyde
content expressed as nmol malondialdehyde/mg protein (27).
Statistical analysis
For data analysis, SPSS 15.0 software (SPSS, Inc.,
Chicago, IL, USA) was used. All data are presented as the mean ±
standard error of the mean. To compare differences between the
means of multiple groups, one-way analysis of variance followed by
Tukey's post hoc test was used. P<0.05 was considered to
indicate statistical significance.
Results
Step-through latency in the
acquisition trial
On assessing delay in the acquisition phase
(step-through to the dark compartment), there were no significant
differences between the groups (Fig.
1A).
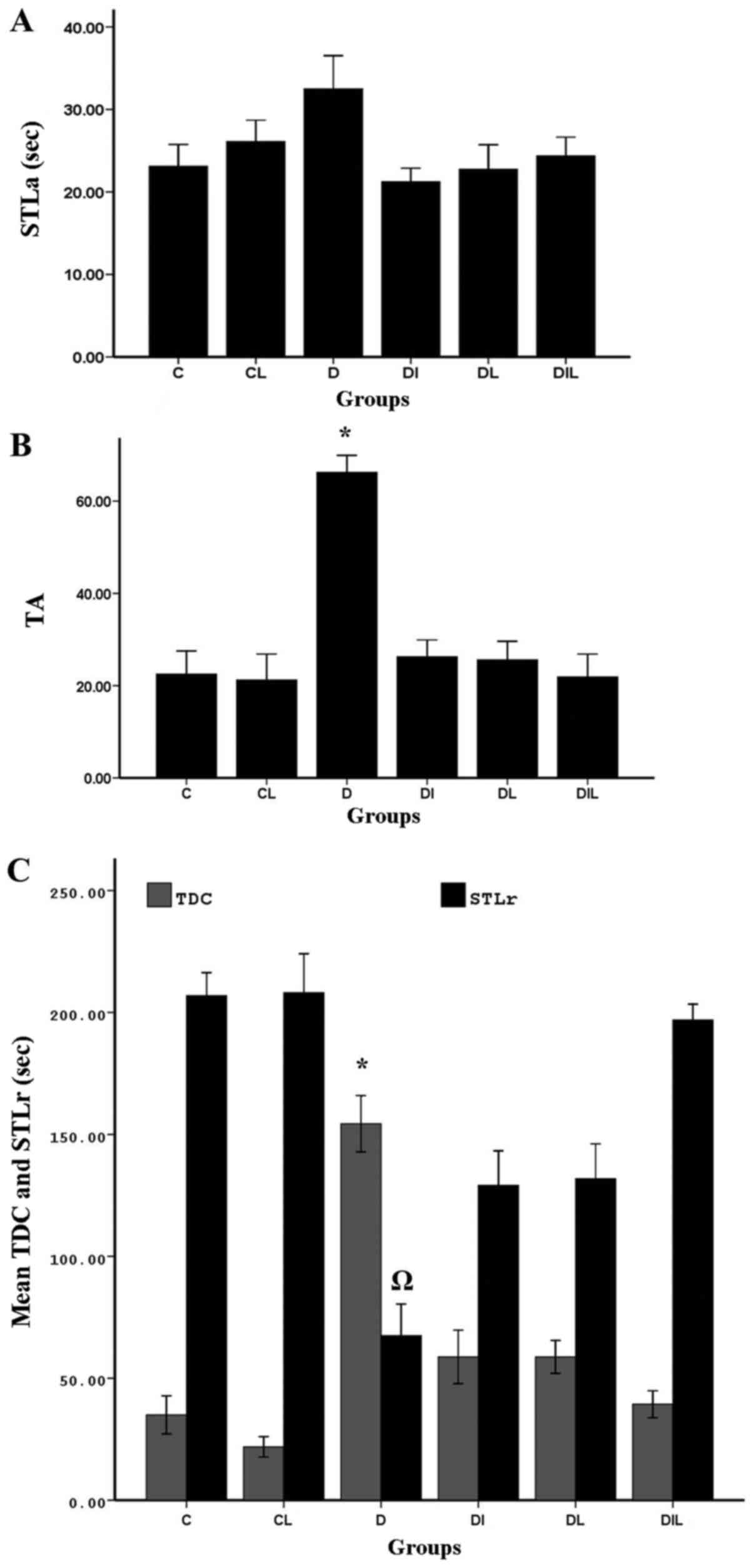 | Figure 1.Effect of lycopene and insulin on (A)
STLa, (B) trial to acquisition (TA) and (C) STLr and TDC in
diabetic rats subjected to the passive avoidance learning test.
*P<0.05, group D vs. other experimental groups;
ΩP<0.05, group D vs. other experimental groups. The
bars indicate the mean ± standard error of the mean. C, control;
CL, control group treated with lycopene; D, untreated diabetic
group; DI, diabetic group treated with insulin; DL, diabetic group
treated with lycopene; DIL, diabetic group treated with insulin and
lycopene; STLa, step-through latency in the acquisition trial;
STLr, step-through latency; TDC, time spent in dark
compartment. |
Responding at the end of trial to
acquisition (TA)
At 24 h after training, memory assessment showed
that TA for step-through between the two chambers was significantly
increased in the diabetic group without treatment compared with in
the control group (P<0.05), indicating that the induction of
diabetes could increase TA. Comparing the diabetic group without
treatment with the groups treated with insulin and lycopene
individually and simultaneously, the increase in TA observed was
statistically significant (P<0.05; Fig. 1B).
Step-through latency (STLr)
At 24 h after training, STLr of the diabetic group
without treatment was significantly decreased compared with that of
the control group (P<0.05), Furthermore, in the diabetic group
without treatment, STLr was significantly reduced compared with in
the diabetic groups treated with insulin, lycopene or both
compounds simultaneously (P<0.05; Fig.
1C).
Time spent in dark compartment
(TDC)
Comparisons of total time spent in the dark
compartment of the shuttle box between diabetic and control rats
revealed significant differences among groups. In the diabetic
group without treatment, probably owing to increased TA between the
two chambers, and thereby reduced STLr, TDC was significantly
increased compared with in the control group (P<0.05). In the
diabetic groups treated with insulin, lycopene or both compounds
simultaneously, TA between the two chambers was reduced compared
with in the diabetic group without treatment, resulting in a
significant reduction in TDC in these groups, compared with in the
diabetic group without treatment (P<0.05; Fig. 1C).
TAC in the hippocampus
The TAC of blood in the diabetic group was
significantly lower than that in the control group (P<0.05). By
contrast, the diabetic rats that underwent treatment for 8 weeks
with insulin, lycopene and a combination thereof exhibited
significant increases in TAC compared with the diabetic rats
without treatment (P<0.05; Fig.
2).
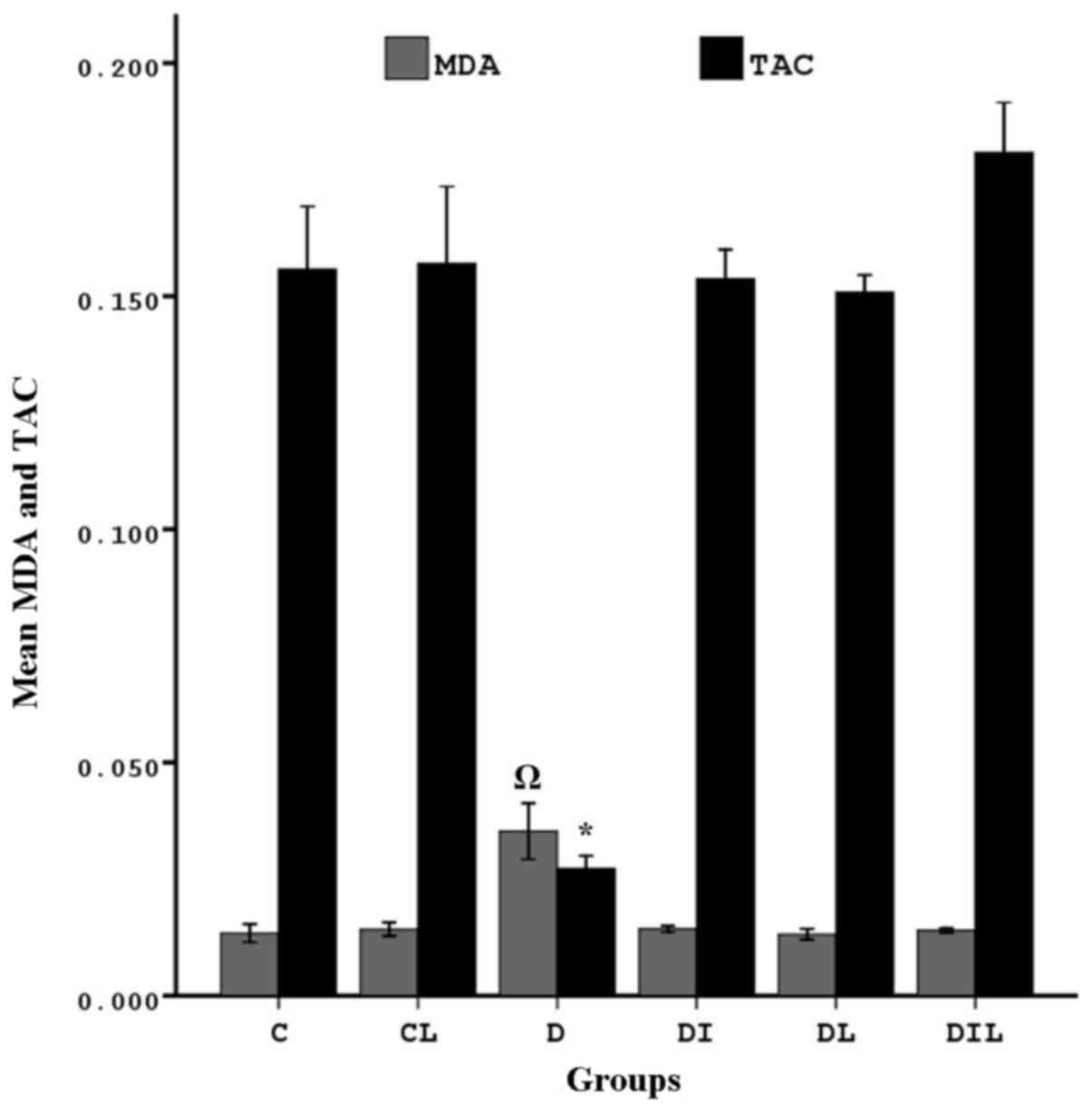 | Figure 2.Effect of lycopene and insulin on TAC
and MDA in the hippocampus of diabetic rats. MDA:
ΩP<0.05, group D vs. other experimental groups. TAC:
*P<0.05, group D vs. other experimental groups. The bars
indicate the mean ± standard error of the mean. C, control; CL,
control group treated with lycopene; D, untreated diabetic group;
DI, diabetic group treated with insulin; DL, diabetic group treated
with lycopene; DIL, diabetic group treated with insulin and
lycopene; TAC, total antioxidant capacity; MDA, malondialdehyde
activity. |
MDA in the hippocampus
MDA was significantly increased in the diabetic
group compared with in the control group (P<0.05). In the
diabetic rats treated for 8 weeks with insulin, lycopene or a
combination thereof, MDA was significantly lower than that in the
diabetic rats without treatment (P<0.05; Fig. 2).
AO staining results
AO is a nucleic acid-selective metachromatic stain
useful for cell cycle determination. In this staining assay, the
dead and intact cells were red and green, respectively. Cell counts
in the different regions of the hippocampus (CA1, CA2, CA3, CA4 and
DG; Fig. 3) revealed that the mean
percentage of intact cells in each of the treated groups was
significantly higher than that for diabetic rats without treatment
(P<0.05). The results of all experimental groups are shown in
Table I.
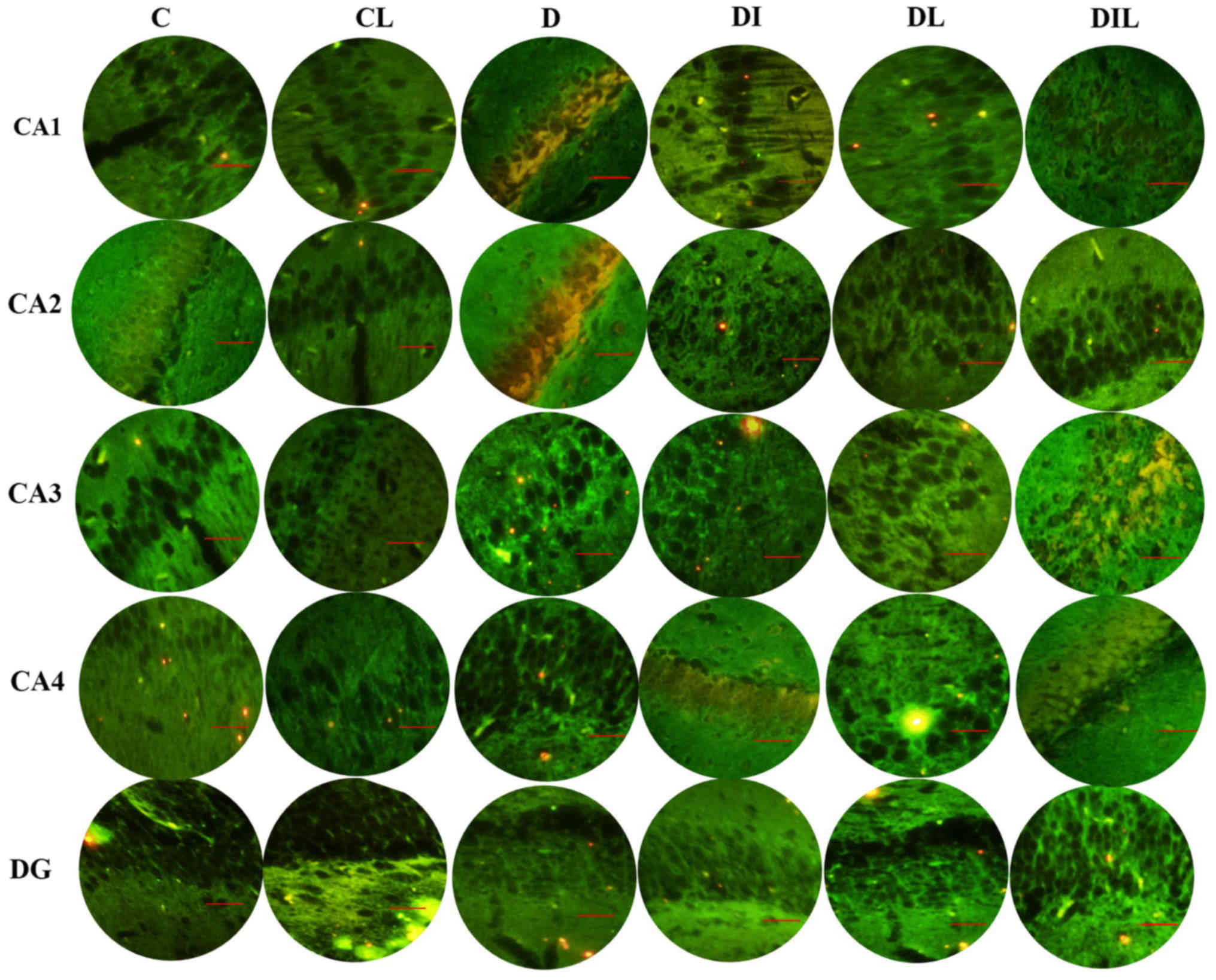 | Figure 3.Effect of lycopene and insulin on
cell viability in the different regions (CA1, CA2, CA3, CA4 and DG)
of the hippocampus according to acridine orange staining.
Magnification, ×400; scale bar, 200 µm. C, control; CL, control
group treated with lycopene; D, untreated diabetic group; DI,
diabetic group treated with insulin; DL, diabetic group treated
with lycopene; DIL, diabetic group treated with insulin and
lycopene; DG, dentate gyrus. |
 | Table I.Effects of insulin and lycopene on
hippocampal neuronal cell viability (acridine orange staining). |
Table I.
Effects of insulin and lycopene on
hippocampal neuronal cell viability (acridine orange staining).
|
| Group cell death
rate, % |
|---|
|
|
|
|---|
| Region | C | CL | D | DI | DL | DIL |
|---|
| CA1 |
0.36±0.31a | 0.42±0.19 | 18.90±1.75 |
2.91±0.65a |
2.73±0.45a |
2.36±0.52a |
| CA2 |
0.30±0.34a | 0.34±0.26 | 21.50±2.16 |
2.74±0.65a |
2.58±0.63a |
2.51±0.43a |
| CA3 |
0.33±0.27a | 0.38±0.25 | 19.00±1.70 |
2.65±0.65a |
2.84±0.68a |
2.10±0.58a |
| CA3 |
0.46±0.32a | 0.56±0.31 | 20.57±1.84 |
2.60±0.92a |
2.48±0.41a |
2.36±0.73a |
| Dentate gyrus |
0.45±0.46a | 0.61±0.23 | 18.43±1.50 |
2.30±0.95a |
2.80±0.70a |
2.23±0.65a |
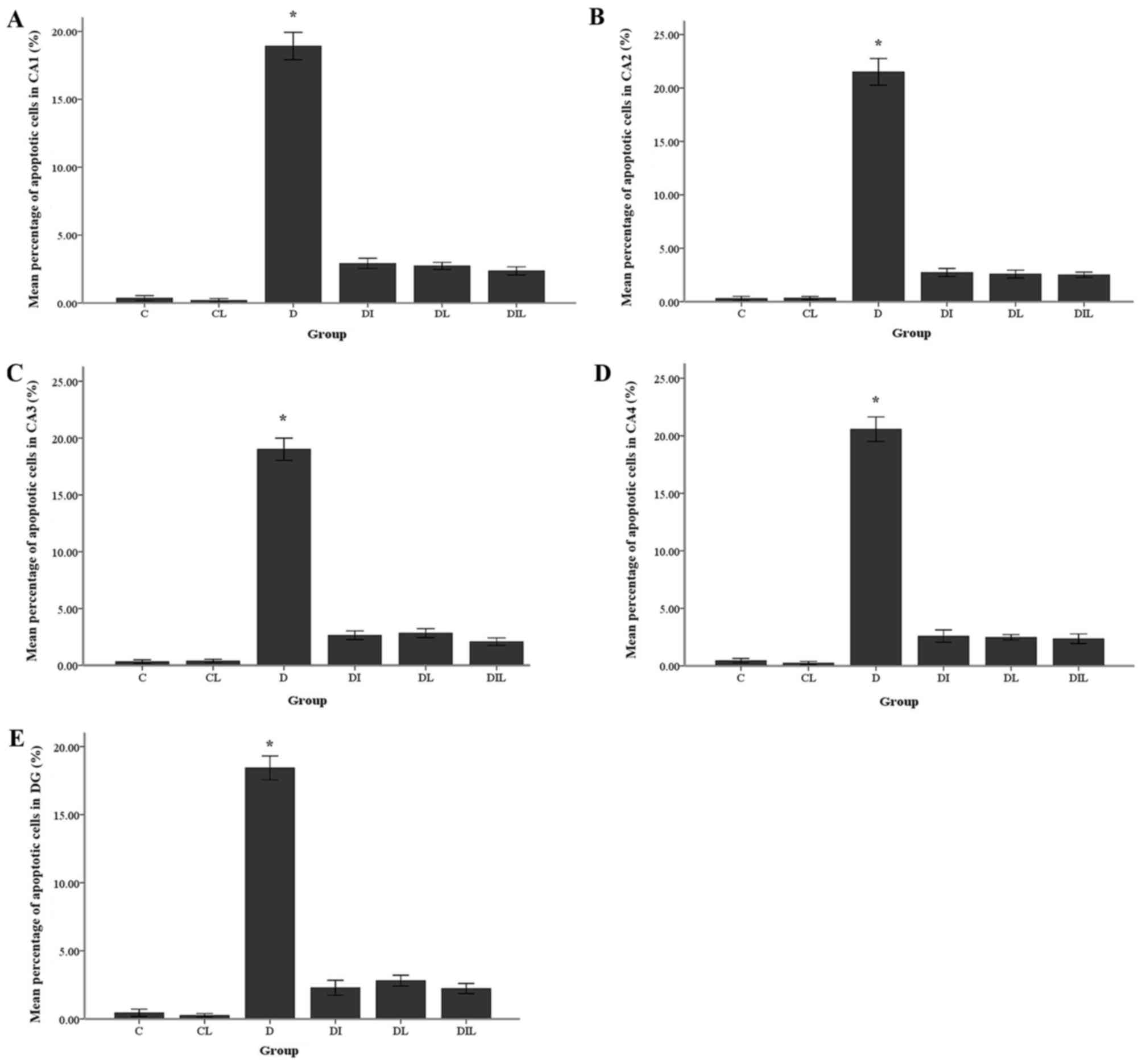
TUNEL staining results
The mean percentage of TUNEL-positive cells in the
diabetic group was significantly increased compared with that in
the control group (P<0.05). By contrast, in each of the insulin,
lycopene and combination-treated groups, the mean percentage of
apoptotic cells was significantly lower than that in diabetic rats
without treatment (P<0.05; Figs. 4
and 5).
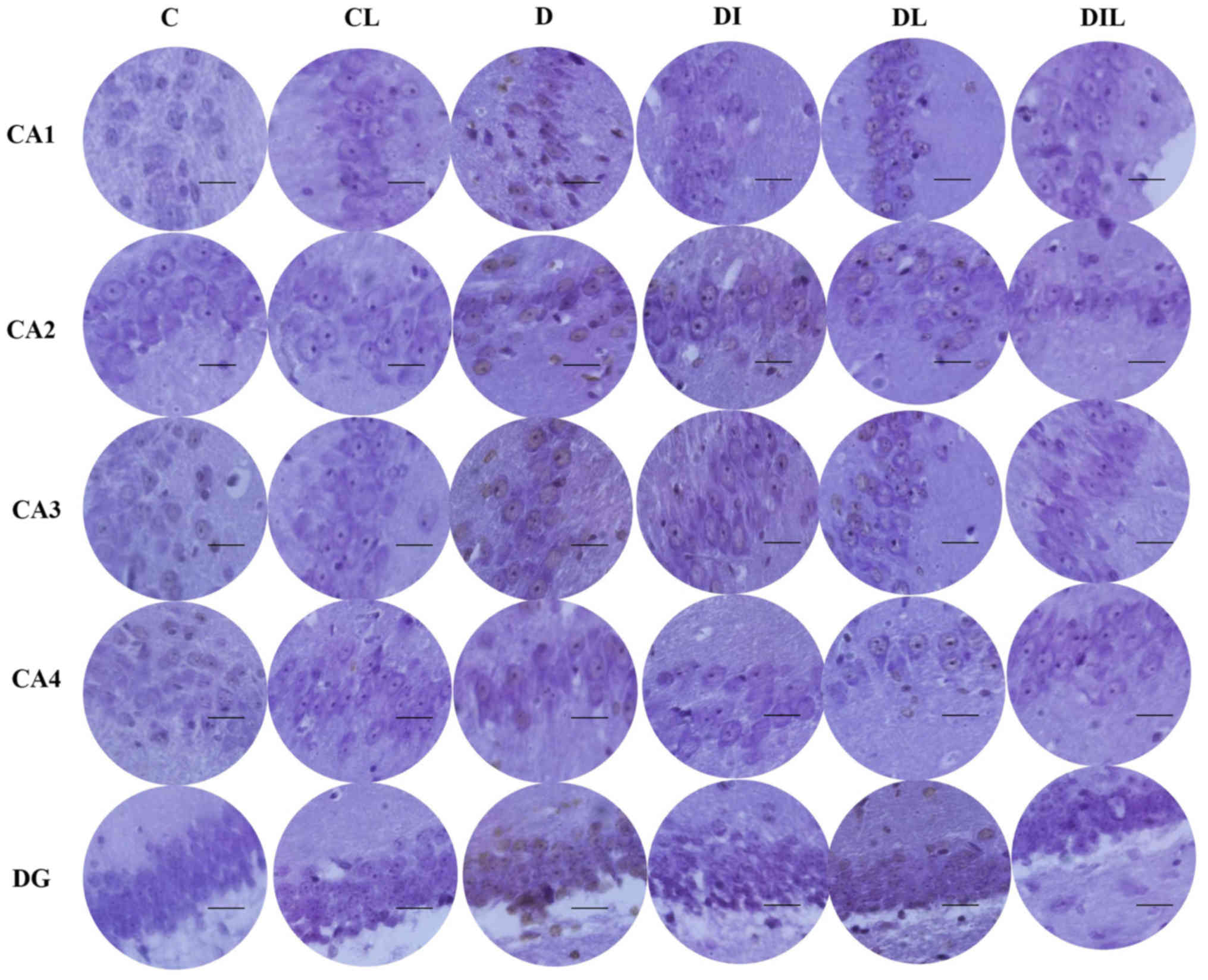 | Figure 4.Effect of lycopene and insulin on
cell apoptosis in the different regions (CA1, CA2, CA3 and CA4 and
DG) of the hippocampus. Apoptosis was evaluated by terminal
deoxynucleotidyl transferase dUTP nick-end labeling assay.
Magnification, ×400; scale bar, 200 µm. C, control; CL, control
group treated with lycopene; D, untreated diabetic group; DI,
diabetic group treated with insulin; DL, diabetic group treated
with lycopene; DIL, diabetic group treated with insulin and
lycopene; DG, dentate gyrus. |
Discussion
The findings of the present study firstly verified
that STZ induced neuronal cell death in the hippocampus region.
Furthermore, diabetic animals treated with lycopene and insulin for
8 weeks exhibited attenuated STZ-induced learning and memory
impairments, associated with increased TAC and decreased MDA.
STZ-induced diabetic hyperglycemia serves an
important role in the degeneration of neurons in various regions of
the brain, including the cingulate cortex, thalamic nuclei and
hippocampus, by increasing the production of ROS (28). Oxidative stress is now recognized as
an important etiological factor associated with the development of
several chronic diseases, including cancer, cardiovascular
diseases, osteoporosis and diabetes (5). Nishikawa et al (29) reported that direct glucose toxicity to
neurons may be due in particular to enhanced intracellular glucose
oxidation, which leads to an increase in ROS production. According
to Arvanitakis et al (30),
oxidative stress is a major factor influencing the extent of
neuronal damage in both experimental diabetic rats and humans.
Lycopene treatment may reduce the enzyme activities of superoxide
dismutase, glutathione peroxidase and catalase to control levels of
ROS in cadmium-treated rats, and may attenuate oxidative stress by
reducing xanthine oxidase activity (31). Akbaraly et al (32) observed that lycopene enhanced
cognitive performance in the elderly. Furthermore, a study by Kuhad
et al (21) demonstrated that
lycopene attenuated diabetes-associated cognitive decline in rats,
at the levels of oxidative-nitrosative stress and peripheral
inflammation associated with the development of cognitive
impairment, which suggested the therapeutic potential of lycopene
in diabetes-induced learning and memory impairment. Thus, lycopene
may protect against diabetes-associated memory decline through
reducing oxidative stress, as evident in rat models.
The TUNEL assay results indicated that lycopene and
insulin individually and simultaneously prevented apoptosis. In
support of this result, Lim et al (33) reported that lycopene inhibited
apoptosis in neural cells via reducing oxidative stress. They also
concluded that the consumption of lycopene-rich foods may prevent
neuronal damage induced by oxidative stress in certain
neuropathological conditions including Alzheimer's disease. Sadek
et al (34) demonstrated that
lycopene could protect brain tissue from apoptotic cell death
through inhibiting monosodium glutamate formation. Feng et
al (35) also revealed that
lycopene was a potent neuroprotectant against apoptosis during
oxidative stress and mitochondrial dysfunction in the hippocampal
neuroglia. Previous study by our group demonstrated that lycopene
and insulin, alone or in combination, could prevent apoptosis in
the hippocampus region of STZ-induced diabetic rats, with increased
expression of anti-apoptotic genes (Bcl-2 and Bcl-xL) and decreased
expression of pro-apoptotic gene Bax (36). Data obtained presently indicated
decreased TAC and increased MDA in the hippocampi of diabetic rats.
Plasma TAC represents an appropriate biochemical parameter for
comparing the overall antioxidant status. Lipid peroxidation is a
well-established mechanism associated with cellular injury and is
frequently used as an indicator of oxidative stress (37). It has previously been revealed that in
STZ-induced diabetic rats, TCA, MDA and antioxidative enzymes
including catalase are altered not only in plasma, but also in
other organs (including the kidney, liver, heart and brain)
(1). The current results indicated
that lipid peroxidation levels were significantly increased, while
glutathione, superoxide dismutase and catalase activities were
markedly decreased in the hippocampus of diabetic rats. It has been
shown that treatment with lycopene may recover the levels of lipid
peroxides, glutathione, superoxide dismutase and catalase to
control values (38). Witztum
(39) reported that lipid
peroxidation products increased oxidative stress conditions and
ROS. Consistent with the present study, Zhang et al
(40) and Prakash and Kumar (41) reported that dietary antioxidants,
including lycopene, could protect important biomolecules and DNA
from oxidation and serve an important role in the prevention of
chronic diseases such as Alzheimer's and Parkinson's disease.
In conclusion, the current data suggested that
insulin and lycopene are effective in protecting hippocampal
neuroglia against STZ-induced damage leading to learning and memory
impairment. Therefore, insulin and lycopene, alone or in
combination, may be considered as drug candidates for neurological
disorders in which oxidative stress serves an important role in
pathogenesis.
Acknowledgements
The authors would like to thank Dr Alireza Shoghli,
Deputy of Research and Technology of Zanjan University of Medical
Sciences, Zanjan, Iran, for assisting with the research.
Funding
The present study was part of an anatomy graduate
student research thesis, which was funded by the Deputy of Research
and Technology at the Zanjan University of Medical Sciences,
Zanjan, Iran with code (grant no. A-12-202-3).
Availability of data and materials
All primary data generated in the present study are
archived at the Zanjan University of Medical Sciences, Zanjan,
Iran, and available from the corresponding author on request.
Authors' contributions
RM performed the experiments. AA acted as study
advisor and helped to revise the manuscript. DS acted as the study
advisor. IJA is the corresponding author, analyzed data, supervised
the project, and wrote the manuscript. All authors discussed the
results and contributed to the final manuscript.
Ethics approval and consent to
participate
The animal experimental protocols were reviewed and
approved by the Institutional Animal Ethics Committee of Zanjan
University of Medical Sciences, Zanjan, Iran (approval no.
ZUMS.REC.1394.29).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tian X, Liu Y, Ren G, Yin L, Liang X, Geng
T, Dang H and An R: Resveratrol limits diabetes-associated
cognitive decline in rats by preventing oxidative stress and
inflammation and modulating hippocampal structural synaptic
plasticity. Brain Res. 1650:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rahmeier FL, Zavalhia LS, Tortorelli LS,
Huf F, Géa LP, Meurer RT, Machado AC, Gomez R and Fernandes MDC:
The effect of taurine and enriched environment on behaviour, memory
and hippocampus of diabetic rats. Neurosci Lett. 630:84–92. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Esteghamati A, Larijani B, Aghajani MH,
Ghaemi F, Kermanchi J, Shahrami A, Saadat M, Esfahani EN, Ganji M,
Noshad S, et al: Diabetes in Iran: Prospective Analysis from First
Nationwide Diabetes Report of National Program for Prevention and
Control of Diabetes (NPPCD-2016). Sci Rep. 7:134612017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Diaz-Valencia PA, Bougnères P and Valleron
AJ: Global epidemiology of type 1 diabetes in young adults and
adults: A systematic review. BMC Public Health. 15:2552015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koppula S, Kumar H, More SV, Kim BW, Kim
IS and Choi DK: Recent advances on the neuroprotective potential of
antioxidants in experimental models of Parkinson's disease. Int J
Mol Sci. 13:10608–10629. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martín-Gallán P, Carrascosa A, Gussinyé M
and Domínguez C: Biomarkers of diabetes-associated oxidative stress
and antioxidant status in young diabetic patients with or without
subclinical complications. Free Radic Biol Med. 34:1563–1574. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wolff SP, Jiang ZY and Hunt JV: Protein
glycation and oxidative stress in diabetes mellitus and ageing.
Free Radic Biol Med. 10:339–352. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baynes JW: Role of oxidative stress in
development of complications in diabetes. Diabetes. 40:405–412.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fukui K, Onodera K, Shinkai T, Suzuki S
and Urano S: Impairment of learning and memory in rats caused by
oxidative stress and aging, and changes in antioxidative defense
systems. Ann N Y Acad Sci. 928:168–175. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reagan LP: Glucose, Stress, and
Hippocampal Neuronal Vulnerability. Int Rev Neurobiol. 51:289–324.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Reagan LP: Insulin signaling effects on
memory and mood. Curr Opin Pharmacol. 7:633–637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zilliox LA, Chadrasekaran K, Kwan JY and
Russell JW: Diabetes and Cognitive Impairment. Curr Diab Rep.
16:872016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gaspar JM, Baptista FI, Galvão J, Castilho
AF, Cunha RA and Ambrósio AF: Diabetes differentially affects the
content of exocytotic proteins in hippocampal and retinal nerve
terminals. Neuroscience. 169:1589–1600. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Anjum R and Khar A: Apoptosis: An
overview. Indian J Biotechnol. 1:58–72. 2002.
|
|
15
|
Sun J, Chu Y-F, Wu X and Liu RH:
Antioxidant and antiproliferative activities of common fruits. J
Agric Food Chem. 50:7449–7454. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Klipstein-Grobusch K, Launer LJ, Geleijnse
JM, Boeing H, Hofman A and Witteman JC: Serum carotenoids and
atherosclerosis. The Rotterdam Study. Atherosclerosis. 148:49–56.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miller NJ, Sampson J, Candeias LP, Bramley
PM and Rice-Evans CA: Antioxidant activities of carotenes and
xanthophylls. FEBS Lett. 384:240–242. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rao AV and Rao LG: Carotenoids and human
health. Pharmacol Res. 55:207–216. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Anarkooli Jafari I, Ganji Barzegar H and
Pourheidar M: The protective effects of insulin and natural honey
against hippocampal cell death in streptozotocin-induced diabetic
rats. J Diabetes Res. 2014:4915712014.PubMed/NCBI
|
|
20
|
Özkan Y, Yilmaz O, Oztürk Aİ and Erşan Y:
Effects of triple antioxidant combination (vitamin E, vitamin C and
α-lipoic acid) with insulin on lipid and cholesterol levels and
fatty acid composition of brain tissue in experimental diabetic and
non-diabetic rats. Cell Biol Int. 29:754–760. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kuhad A, Sethi R and Chopra K: Lycopene
attenuates diabetes-associated cognitive decline in rats. Life Sci.
83:128–134. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Firouzjaei MA, Jafari MR, Eskandari M,
Anarkoli IJ and Alipour M: Aminoguanidine changes hippocampal
expression of apoptosis-related genes, improves passive avoidance
learning and memory in streptozotocin-induced diabetic rats. Cell
Mol Neurobiol. 34:343–350. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khaki A, Novin MG, Khaki AA, Nouri M,
Sanati E and Nikmanesh M: Comparative study of the effects of
gentamicin, neomycin, streptomycin and ofloxacin antibiotics on
sperm parameters and testis apoptosis in rats. Pak J Biol Sci.
11:1683–1689. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alipour M, Gholami MR, Anarkooli Jafari I,
Sohrabi D, Tajki J and Pourheidar M: Intraperitoneal aminoguanidine
improves sciatic nerve ischemia-reperfusion injury in male
sprague-dawley rats. Cell Mol Neurobiol. 31:765–773. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Prieto P, Pineda M and Aguilar M:
Spectrophotometric quantitation of antioxidant capacity through the
formation of a phosphomolybdenum complex: Specific application to
the determination of vitamin E. Anal Biochem. 269:337–341. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wills ED: Mechanisms of lipid peroxide
formation in animal tissues. Biochem J. 99:667–676. 1966.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wokes F and Still BM: The estimation of
protein by the biuret and Greenberg methods. Biochem J. 36:797–806.
1942. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li ZG and Sima AA: C-peptide and central
nervous system complications in diabetes. Exp Diabesity Res.
5:79–90. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nishikawa T, Edelstein D, Du XL, Yamagishi
S, Matsumura T, Kaneda Y, Yorek MA, Beebe D, Oates PJ, Hammes HP,
et al: Normalizing mitochondrial superoxide production blocks three
pathways of hyperglycaemic damage. Nature. 404:787–790. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Arvanitakis Z, Wilson RS, Bienias JL,
Evans DA and Bennett DA: Diabetes mellitus and risk of Alzheimer
disease and decline in cognitive function. Arch Neurol. 61:661–666.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rencuzogullari N and Erdogan S: Oral
administration of lycopene reverses cadmium-suppressed body weight
loss and lipid peroxidation in rats. Biol Trace Elem Res.
118:175–183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Akbaraly NT, Faure H, Gourlet V, Favier A
and Berr C: Plasma carotenoid levels and cognitive performance in
an elderly population: Results of the EVA Study. J Gerontol A Biol
Sci Med Sci. 62:308–316. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lim S, Hwang S, Yu JH, Lim JW and Kim H:
Lycopene inhibits regulator of calcineurin 1-mediated apoptosis by
reducing oxidative stress and down-regulating Nucling in neuronal
cells. Mol Nutr Food Res. 61:16005302017. View Article : Google Scholar
|
|
34
|
Sadek K, Abouzed T and Nasr S: Lycopene
modulates cholinergic dysfunction, Bcl-2/Bax balance, and
antioxidant enzymes gene transcripts in monosodium glutamate (E621)
induced neurotoxicity in a rat model. Can J Physiol Pharmacol.
94:394–401. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng C, Luo T, Zhang S, Liu K, Zhang Y,
Luo Y and Ge P: Lycopene protects human SH SY5Y neuroblastoma cells
against hydrogen peroxide induced death via inhibition of oxidative
stress and mitochondria associated apoptotic pathways. Mol Med Rep.
13:4205–4214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Soleymaninejad M, Joursaraei SG, Feizi F
and Anarkooli Jafari I: The Effects of lycopene and insulin on
histological changes and the expression level of Bcl-2 family genes
in the hippocampus of streptozotocin-induced diabetic rats. J
Diabetes Res. 2017:46509392017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Moore K and Roberts LJ II: Measurement of
lipid peroxidation. Free Radic Res. 28:659–671. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sharma S, Anjaneyulu M, Kulkarni SK and
Chopra K: Resveratrol, a polyphenolic phytoalexin, attenuates
diabetic nephropathy in rats. Pharmacology. 76:69–75. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Witztum JL: The oxidation hypothesis of
atherosclerosis. Lancet. 344:793–795. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang YJ, Gan RY, Li S, Zhou Y, Li AN, Xu
DP and Li HB: Antioxidant phytochemicals for the prevention and
treatment of chronic diseases. Molecules. 20:21138–21156. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Prakash A and Kumar A: Lycopene protects
against memory impairment and mito-oxidative damage induced by
colchicine in rats: An evidence of nitric oxide signaling. Eur J
Pharmacol. 721:373–381. 2013. View Article : Google Scholar : PubMed/NCBI
|