Introduction
The innate immune system recognizes
pathogen-associated molecular patterns through pattern-recognition
receptors (PRRs). Thus far, numerous PRRs have been identified,
including Toll-like receptors (TLRs), retinoic acid-inducible
gene-I (RIG-I)-like receptors (RLRs) and nucleotide-binding
oligomerization domain-like receptors (1–4). Among
these receptors, RLRs are cytosolic virus sensors and are
indispensable for antiviral immunity.
RLRs are DExD/H box-containing RNA helicases and
play a key role in sensing RNA virus invasion (5). They consist of RIG-I, melanoma
differentiation-associated gene 5 (MDA5) and laboratory of genetics
and physiology 2 (LGP2). RIG-I and MDA5 contain N-terminal domains,
consisting of the tandem caspase activation and recruitment domains
(CARDs), the central DExD/H box RNA helicase domain and the
C-terminal regulatory domain, whereas LGP2 lacks CARDs. RIG-I and
MDA5 share structural and functional similarities, but they
recognize distinct types of RNA viruses (6). RIG-I recognizes relatively short
double-stranded RNA (dsRNA) and 5′ triphosphate-single-stranded RNA
and are important in sensing influenza virus and hepatitis C virus.
By contrast, MDA5 recognizes long dsRNA (7) and they sense picornaviruses.
Subsequent to RIG-I and MDA5 sensing RNA virus invasion, they
interact with a CARD-containing adaptor protein and interferon
(IFN)-β promoter stimulator-1 through their CARDs, which results in
the induction of antiviral cytokine type I IFN, such as IFN-β.
Our recent study investigated the effects of
ionizing radiation on TLR2 and TLR4 using the human monocytic cell
line THP1 and THP1-derived macrophage-like cells and showed that
ionizing radiation affects these expression levels and the response
to their agonist depending on the cell differentiation state
(8). In THP1-derived macrophages,
the expression of TLR4 was decreased following X-irradiation and
the expression of its agonist lipopolysaccharide (LPS)-inducible
IFN-β was attenuated by X-irradiation. These results indicate that
the antiviral immune system of TLR4 in X-irradiated macrophages
cannot properly respond to viral infections subsequent to
LPS-containing gram-negative bacteria infections. Therefore, the
antiviral immune system by RLRs is important in this situation.
However, the effects of ionizing radiation on RLRs remain unknown.
Therefore, the present study investigated the effects of ionizing
radiation on the expression of RIG-I and MDA5 in
THP1-derived macrophages and the response to their agonist, a dsRNA
analogue polyinosinic-polycytidylic acid [poly(I:C)]/LyoVec™.
Materials and methods
Reagents
Phorbol 12-myristate 13-acetate (PMA) was purchased
from Sigma-Aldrich (St. Louis, MO, USA). The poly(I:C)-low
molecular weight (LMW)/LyoVec™ and poly(I:C)-high molecular weight
(HMW)/LyoVec™ were purchased from InvivoGen (San Diego, CA, USA).
Rabbit anti-human RIG-I (cat no. 4520), MDA5 (cat no. 5321)
monoclonal antibodies and anti-rabbit immunoglobulin G (IgG)
horseradish peroxidase (HRP)-linked (cat no. 7074) antibody were
purchased from Cell Signaling Technology Japan, K.K. (Tokyo,
Japan). Goat anti-human actin polyclonal antibody (sc-1615) and
HRP-conjugated donkey anti-goat IgG (sc-2056) were purchased from
Santa Cruz Biotechnology, Inc., (Santa Cruz, CA, USA).
Cell culture
THP1 human acute monocytic leukemia cells were
obtained from RIKEN BioResource Center (Tsukuba, Japan). Cells were
cultured in RPMI-1640 supplemented with 1% penicillin and
streptomycin (Gibco, Grand Island, NY, USA) and 10%
heat-inactivated fetal bovine serum (Japan Bioserum Co., Ltd.,
Fukuyama, Japan) at 37°C in a humidified atmosphere containing 5%
CO2. THP1-derived macrophages (macrophage-like cells)
were prepared as previously described (8). THP1 cells (2.0×105
cells/ml) were plated in 60-mm dishes (Iwaki, Tokyo, Japan) with 4
ml of medium containing 100 ng/ml PMA and cultured for 48 h. After
the 48-h culture, the medium containing PMA was replaced with fresh
medium not containing PMA and macrophage-like cells were used in
experiments.
In vitro X-irradiation
X-irradiation (150 kVp, 20 mA, 0.5 mm Al and 0.3 mm
Cu filters) was performed using an X-ray generator (MBR-1520R-3;
Hitachi Medical Corporation, Tokyo, Japan) at a distance of 45 cm
from the focus and a dose rate of 1.00 Gy/min.
Stimulation with
poly(I:C)/LyoVec™
To stimulate RLRs, two types of poly(I:C)/LyoVec™
(InvivoGen), which is a complex of poly(I:C) and the transfection
reagent LyoVec™, were used. In brief, macrophage-like cells were
exposed to X-rays and 500 ng/ml poly(I:C)-LMW/LyoVec™ or
poly(I:C)-HMW/LyoVec™ was added to the culture 24 h after
X-irradiation. After an additional 24 h, cells were harvested for
reverse transcription-polymerase chain reaction (RT-PCR) and
western blot analysis.
RT-PCR
Total RNA was extracted using the RNeasy Mini kit
(Qiagen, Valencia, CA, USA) and quantified using a NanoDrop
(Thermo, Wilmington, DE, USA). cDNA templates were synthesized from
1 µg RNA using the iScript cDNA Synthesis kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA), according to the
manufacturer's instructions. PCR was performed using the AccuPrime™
Taq DNA Polymerase system (Invitrogen Life Technologies, Carlsbad,
CA, USA). The primer sequences used are shown in Table I. The reaction conditions for
RIG-I were 94°C for 1 min followed by 30 cycles of 94°C for
1 min, 55°C for 1 min and 72°C for 1 min, and subsequently 72°C for
10 min. The reaction conditions for MDA5 were 94°C for 1 min
followed by 30 cycles of 94°C for 1 min, 64°C for 1 min and 72°C
for 1 min, and subsequently 72°C for 10 min. The reaction
conditions for IFN-β and β-actin were as reported
elsewhere (8). The PCR products
were confirmed using electrophoresis on ethidium bromide-stained
1.5% agarose gels.
 | Table IPrimer sequences for reverse
transcription-polymerase chain reaction. |
Table I
Primer sequences for reverse
transcription-polymerase chain reaction.
| Primer | Sequence (5′→3′) | PCR products
(bp) |
|---|
| RIG-I | F:
GCATATTGACTGGACGTGGCA | 644 |
| R:
CAGTCATGGCTGCAGTTCTGTC | |
| MDA5 | F:
GCAAGAGCATCCCCGGAGCC | 601 |
| R:
TCGTGGCCCCTCCAACACCA | |
| IFN−β | F:
CCTGTGGCAATTGAATGGGAGGC | 370 |
| R:
CCAGGCACAGTGACTGTACTCCTT | |
| β-actin | F:
GGCACCCAGCACATTGAAGA | 632 |
| R:
GGCACGAAGGCTCATCATTC | |
SDS-PAGE and western blotting
Cells were harvested and suspended in CelLytic™ M
Cell Lysis reagent (Sigma-Aldrich) containing 1% Protease Inhibitor
cocktail (Sigma-Aldrich) on ice for 30 min. After centrifugation at
20,600 x g for 20 min at 4°C, supernatants were collected. The
protein concentration was determined using the Bio-Rad Protein
Assay kit and a SmartSpec™ plus spectrophotometer (Bio-Rad
Laboratories, Inc.). Each lysate was mixed with 2X sample buffer
(Bio-Rad Laboratories, Inc.) containing 5% 2-mercaptoethanol. After
boiling for 5 min, proteins were separated using 4–20%
Mini-PROTEAN® TGX™ Precast gels (Bio-Rad Laboratories,
Inc.) and transferred onto polyvinylidene difluoride membranes of
Trans-Blot® Turbo™ Mini PVDF Transfer pack (Bio-Rad
Laboratories, Inc.) using the Trans-Blot® Turbo™
Transfer system (Bio-Rad Laboratories, Inc.). The membranes were
blocked in TBST buffer [10 mmol/l HCl (pH 7.5), 100 mmol/l NaCl and
0.1% Tween-20] containing 4% ECL Prime Blocking agent (GE
Healthcare UK Ltd., Little Chalfont, England). The membranes were
probed with each primary antibody in Can Get Signal®
Immunoreaction Enhancer solution 1 (Toyobo, Co., Ltd, Osaka, Japan)
overnight at 4°C. Following the reaction with the primary
antibodies, the membranes were labeled with HRP-conjugated
secondary antibodies in Can Get Signal® Immunoreaction
Enhancer solution 2 (Toyobo, Co., Ltd) for 1 h. The antigens were
visualized by the ECL Prime western blotting detection system (GE
Healthcare UK Ltd.).
Results and Discussion
In the present study, the effects of ionizing
radiation on RIG-I and MDA5 in THP1-derived
macrophage-like cells were investigated.
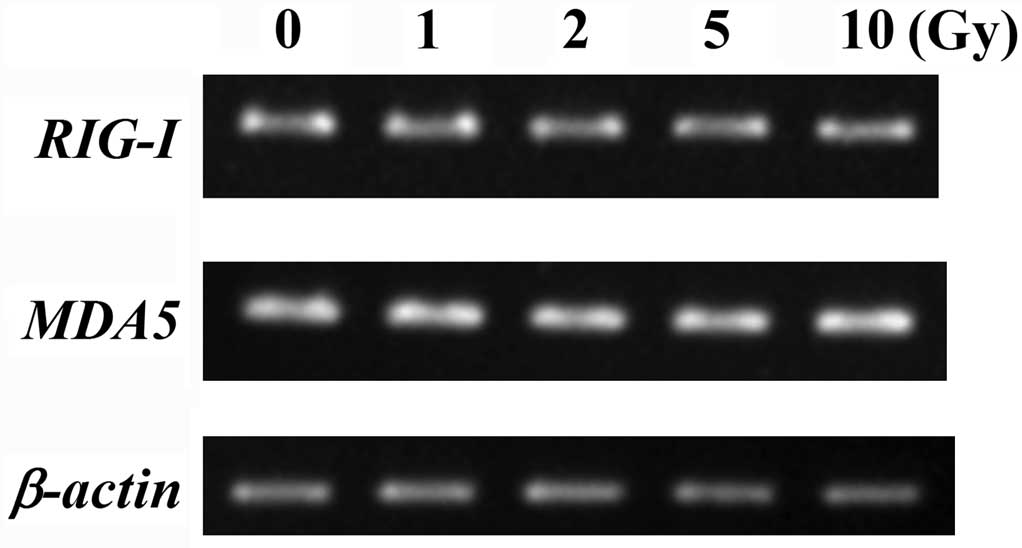
As shown in Fig. 1,
macrophage-like cells expressed RIG-I and MDA5 at the
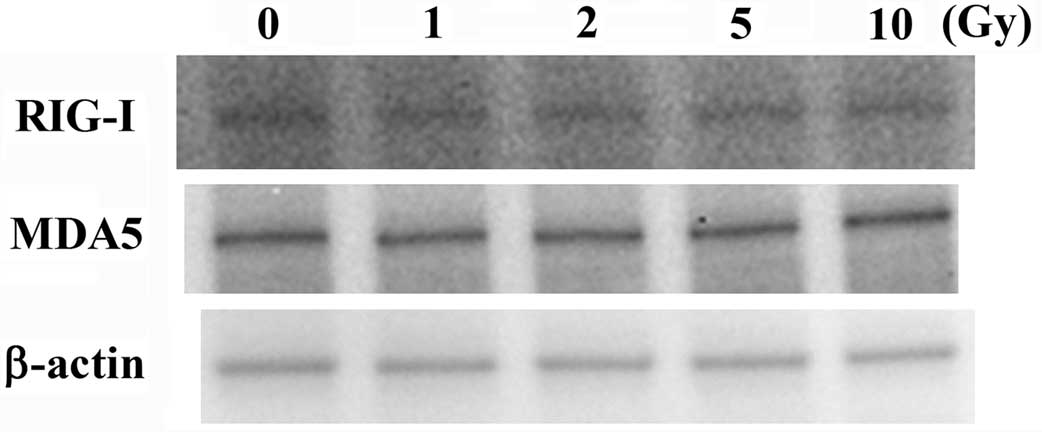
mRNA level. Furthermore, they also expressed these receptors at
protein level (Fig. 2). In
addition, X-irradiated macrophage-like cells expressed RIG-I
and MDA5 at mRNA and protein levels and these expression
levels were identical to those of non-irradiated cells (Figs. 1 and 2). These results indicate that ionizing
radiation neither affects transcriptional nor post-transcriptional
regulation of RIG-I and MDA5 expression. Our recent
study reported that ionizing radiation decreased the expression of
TLR2 and TLR4 on macrophage-like cells; thus, showing that the
effect of ionizing radiation on PRRs depends on the types of PRRs,
particularly affecting the expression of TLR2 and TLR4. Certain
stimuli, such as IFN-γ and LPS, have been reported to upregulate
the expression of RIG-I, as well as TLR2 and
TLR4 (9–11). Therefore, it is likely that ionizing
radiation does not affect the signaling pathway required for the
induction of TLRs expression by IFN-γ or LPS.
As the recognition of dsRNA, such as poly(I:C)
through RIG-I or MDA5 leads to the induction of the antiviral
cytokine IFN-β, IFN-β mRNA expression of macrophage-like
cells following poly(I:C)-LMW/LyoVec™ or poly(I:C)-HMW/LyoVec™
stimulation was investigated. According to the manufacture's data
sheet, the average size of poly(I:C)-LMW and poly(I:C)-HMW is 0.2–1
and 1.5–8 kb, respectively. Poly(I:C)-LMW and poly(I:C)-HMW/LyoVec™
are believed to be recognized by RIG-I and MDA5, respectively, as
RIG-I and MDA5 recognize short (<0.3 kb) and long (>4 kb)
poly(I:C), respectively (7).
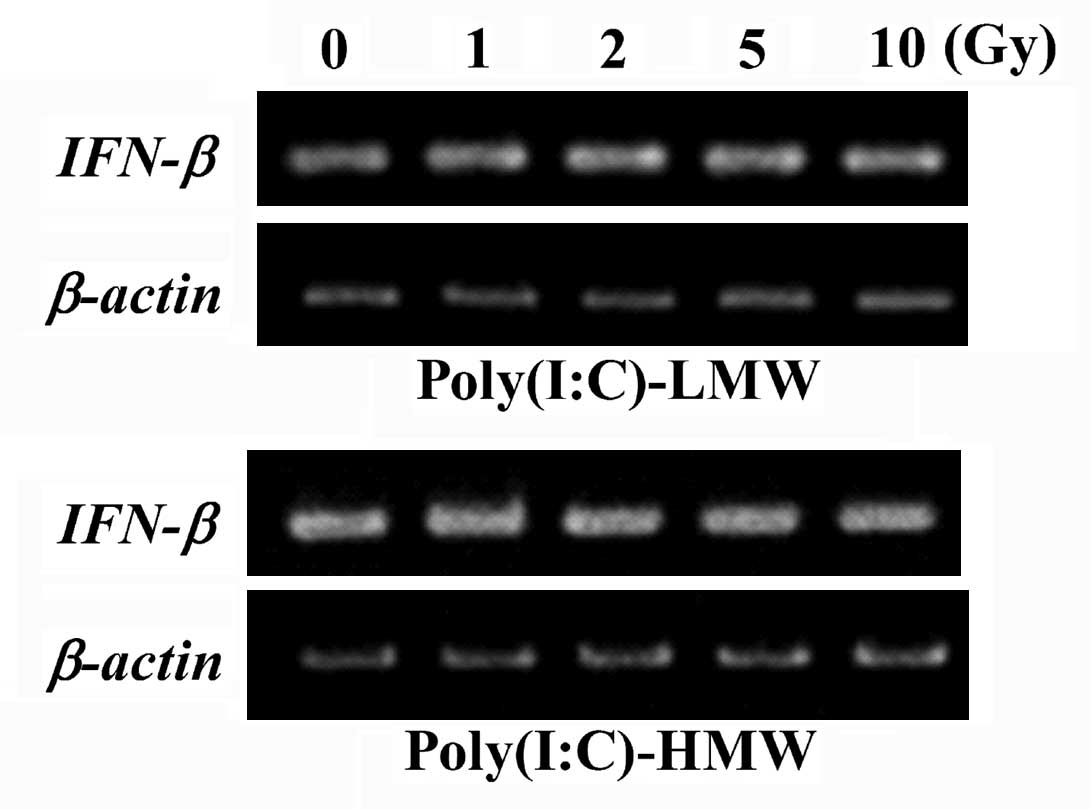
Although non-stimulated macrophage-like cells did not express
detectable IFN-β mRNA (data not shown), the expression of
IFN-β mRNA was observed after treatment with poly(I:C)-LMW
or poly(I:C)-HMW/LyoVec™ for 24 h (Fig.
3). The X-irradiated macrophage-like cells also expressed
IFN-β mRNA following each poly(I:C)/LyoVec™ stimulation and
their expression levels were identical to those of non-irradiated
cells (Fig. 3); thus, indicating
that X-irradiated macrophage-like cells retain the ability to
induce IFN-β following poly(I:C) stimulation. Our recent
study showed that the induction of IFN-β mRNA subsequent to
LPS stimulation was lower in X-irradiated macrophage-like cells
compared to non-irradiated cells (8), indicating that the antiviral immune
system of TLR4 in X-irradiated macrophages cannot properly respond
to viral infections following the gram-negative bacteria
infections. However, this theory could be rejected by the present
study as antiviral immune systems of RLRs normally function
following exposure to ionizing radiation. By contrast, Besch et
al (12) reported that
RIG-I/MDA5 agonists induce proapoptotic signaling in human melanoma
cells; thus, showing that the stimulation of RIG-I/MDA5 is a
potentially useful therapeutic application in cancer treatment.
Therefore, the present results also indicate a possibility that
RLRs are an effective target for the induction of antitumor
immunity during the radiation therapy. To confirm this possibility,
further investigations regarding the antitumor effects of
cotreatment with poly(I:C)/LyoVec™ and ionizing radiation are
required.
Acknowledgements
The authors would like to thank Enago (www.enago.jp) for the English language review. The
present study was supported by a JSPS KAKENHI Grant-in-Aid for
Young Scientists (B; nos. 23791383 and 25861053). The study was
also partially supported by a Hirosaki University Grant for
Exploratory Research by Young Scientists and a Priority Research
Grant for Young Scientists Designated by the President of Hirosaki
University.
References
|
1
|
Creagh EM and O'Neill LA: TLRs, NLRs and
RLRs: a trinity of pathogen sensors that co-operate in innate
immunity. Trends Immunol. 27:352–357. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yan H, Ohno N and Tsuji NM: The role of
C-type lectin receptors in immune homeostasis. Int Immunopharmacol.
16:353–357. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vabret N and Blander JM: Sensing microbial
RNA in the cytosol. Front Immunol. 4:4682013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu D, Rhebergen AM and Eisenbarth SC:
Licensing adaptive immunity by NOD-like receptors. Front Immunol.
4:4862013.PubMed/NCBI
|
|
5
|
Matsumiya T and Stafforini DM: Function
and regulation of retinoic acid-inducible gene-I. Crit Rev Immunol.
30:489–513. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kato H, Takeuchi O, Mikamo-Satoh E, Hirai
R, Kawai T, Matsushita K, Hiiragi A, Dermody TS, Fujita T and Akira
S: Length-dependent recognition of double-stranded ribonucleic
acids by retinoic acid-inducible gene-I and melanoma
differentiation-associated gene 5. J Exp Med. 205:1601–1610. 2008.
View Article : Google Scholar
|
|
7
|
Kato H, Takeuchi O, Sato S, et al:
Differential roles of MDA5 and RIG-I helicases in the recognition
of RNA viruses. Nature. 441:101–105. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yoshino H, Chiba K, Saitoh T and
Kashiwakura I: Ionizing radiation affects the expression of
Toll-like receptors 2 and 4 in human monocytic cells through c-Jun
N-terminal kinase activation. J Radiat Res. 55:876–884. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Faure E, Thomas L, Xu H, Medvedev A,
Equils O and Arditi M: Bacterial lipopolysaccharide and IFN-gamma
induce Toll-like receptor 2 and Toll-like receptor 4 expression in
human endothelial cells: role of NF-kappa B activation. J Immunol.
166:2018–2024. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Imaizumi T, Aratani S, Nakajima T, et al:
Retinoic acid-inducible gene-I is induced in endothelial cells by
LPS and regulates expression of COX-2. Biochem Biophys Res Commun.
292:274–279. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Imaizumi T, Yagihashi N, Kubota K, Yoshida
H, Sakaki H, Yagihashi S, Kimura H and Satoh K: Expression of
retinoic acid-inducible gene-I (RIG-I) in macrophages: possible
involvement of RIG-I in atherosclerosis. J Atheroscler Thromb.
14:51–55. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Besch R, Poeck H, Hohenauer T, et al:
Proapoptotic signaling induced by RIG-I and MDA-5 results in type I
interferon-independent apoptosis in human melanoma cells. J Clin
Invest. 119:2399–2411. 2009.PubMed/NCBI
|