Introduction
Radiotherapy has an increasingly notable role in the
treatment of the majority of malignant and a number of benign
neurological neoplasms, and the treatment of select nonmalignant
entities (1). However, the maximum
radiation dose that can be used is limited by the tolerance of
normal tissues surrounding the tumor (2). Thus far, in order to reduce
radiotherapy-induced central nervous system (CNS) damage, several
attempts are being made. One of these approaches is to apply the
total dose locally in fractions, in order to preserve healthy
neural tissue. Additionally, searches for novel treatment
opportunities to prevent radiation damage are continuing. Following
this, newer therapeutic methods may help diminish the risks of
radiotherapy by not limiting the volume treated.
Histone deacetylase (HDAC) inhibitors represent a
novel class of radiation protectors and mitigators against
total-body irradiation and can produce a significant reduction in
injury even when administered following the radiation exposure
(3). Various mechanisms have been
proposed for the radioprotective effects of HDAC inhibitors
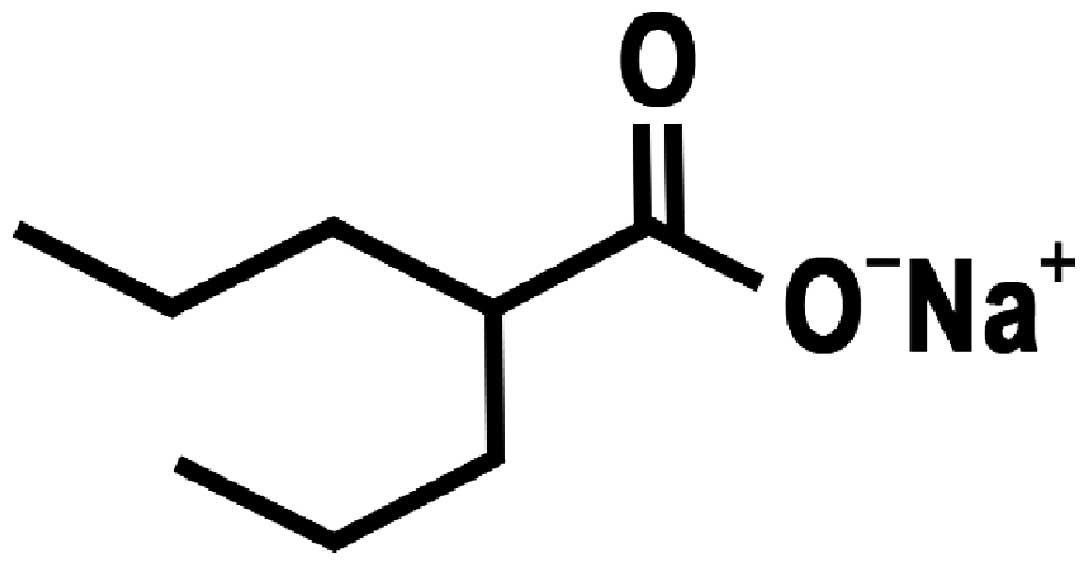
(4–6). Valproic acid (VPA) (Fig. 1), a HDAC inhibitor, is frequently
prescribed as an anti-epileptic drug in patients with brain tumor
due to its effectiveness, oral bioavailability and generally low
toxicity profile (7–10). Previous findings have shown that VPA
enhanced the radiation response of various brain tumor cell types
in vitro and in vivo (11–14).
Notably, VPA not only radiosensitizes tumor cells, but it may also
protect normal brain from radiation.
Results from the study by Lai et al (15) indicate that VPA may decrease human
neural cells vulnerability to cellular injury evoked by oxidative
stress, possibly arising from putative mitochondrial disturbances
involved in bipolar disorder. Similarly, VPA has neuroprotective
effects in cultured cortical neurons undergoing spontaneous cell
death. A previous study also indicated that the neuroprotective
properties of VPA involve modulation of neurotrophic factors and
receptors for melatonin, which is also believed to play a role in
neuroprotection (16). However, it
is also thought that VPA reduces DNA double-strand break repair
capacity and increases radiosensitivity in fibroblasts obtained
from healthy skin tissue (6).
The histological effects of radiation on normal
brain tissues have been studied by numerous investigators and the
morphological character described has generally been the same,
regardless of species or the type of radiation used to induce the
damage (17–20). The majority of these studies used
single X-ray (17–19) rather than fractionated radiotherapy
(FRT) (20,21) although fractionated radiation is
more commonly used clinically. However, radioprotection of VPA in
rat brain following moderate-dose FRT is not clearly
understood.
To maximize the therapeutic ratio of a
radiosensitizing agent, it is important that the drug does not
enhance the radiation-induced toxicity of normal cells or tissue.
The present study examined the whole-brain radioprotection of VPA
following FRT in vivo. Tissue samples from Wistar rats
treated with X-ray plus VPA were studied to gain a certain insight
as to how the compound modifies the radiation response of normal
brain. The primary goal was to perform evaluations of the cortex
cell response following VPA and fractionated X-ray irradiation. The
occurrence of radiation-induced apoptosis in normal brain was
investigated using immunohistochemistry (IHC). The development of
vascular changes in the rat brain was investigated by means of
transmission electron microscope (TEM). Animal weights of the TEM
group over 14 days starting from the injection of VPA were also
observed.
Materials and methods
Animals
At the beginning of the experiment, male Wistar rats
(Center for New Drugs Evaluation of Shandong University, Jinan,
China), 8 weeks old and weighing ~180–190 g, were allowed to
acclimatize for 1 week prior to the start of the study. The rats
were maintained in environment-controlled (temperature and
lighting) animal facilities, provided food and water ad
libitum and randomly assigned to the experimental groups. All
the studies were performed with prior approval and in accordance
with Institutional and National standards of Animal Care.
Irradiation
The whole-brain irradiation (WBI) was delivered by
X-ray (6 MeV-energy and 3 Gy/min). The rats were irradiated using a
6-MV linear accelerator (Elekta Synergy, Ltd., Stockholm, Sweden),
under mild anaesthesia by chloral hydrate [30 mg/kg,
intraperitoneally (i.p.)]. A total of four rats were placed in a
reproducible way; in a prone position on the linac couch with laser
alignment. The radiation field was 15×15 cm at a source-axis
distance of 100 cm. The isocenter was in the midline of the brain
and the posterior limit of the field corresponded to the line
passing by the posterior section of the two ears (22). The dose was delivered according to
the reading of the ‘Ionex’ ionization chamber, which was positioned
directly above the skull. Dosimetry was performed by ion chamber
and chemical Fricke dosimetry.
Experimental design
After 7 days of handling, 48 rats were randomly
divided into four groups, consisting of 12 rats each. The control
group (sham-exposed group) received sham irradiation plus
physiological saline. The VPA group received sham irradiation plus
150 mg/kg VPA (sodium salt; Sigma, St. Louis, MO, USA). The X-ray
group received X-ray WBI plus physiological saline. The combined
group received X-ray WBI plus 150 mg/kg VPA. The dose of VPA used
in the present study (150 mg/kg) was based on a previous study
(11). The rats were administered
10 injections (150 mg/kg) over 5 days, by i.p. every 12 h.
Following the third injection, FRT was delivered to the whole brain
of the rats at a daily dose of 3 Gy, 5 days/week for 1 week, for a
total of 15 Gy. Each group was randomly divided into two sections;
one section was obtained for histological study using histology
evaluation and the other was used for ultra-structural analysis
under the electron microscope (EM). The animal weights of the TEM
group over 14 days starting from the first injection of VPA were
also observed.
IHC
IHC staining was used to confirm the presence of
normal brain tissue damage in Wistar rats. One section of the rats
was sacrificed and the brains were harvested. Animals were
sacrificed 1 day after radiation using 0.4 mg/kg of chloral hydrate
(i.p.) and were subsequently perfused with 100 ml (10 min) of
saline before 500 ml (30 min) of 4% paraformaldehyde (Sigma) in
phosphate-buffered saline (PBS) for histological analysis. The
whole brain was collected and fixed in 4% paraformaldehyde at 4°C
overnight. The fixed brains were sliced coronally and embedded in
paraffin wax. Radiation brain injury blocks were processed and cut
into serial 4-µm sections for histology.
Cell apoptosis was analyzed in the cortex according
to the method of IHC using protein caspase-3 (Sigma). Caspase-3
immunostaining was performed on sections from each group. The
sections were washed in PBS and Tris-buffered saline and incubated
with 10% normal goat serum for 30 min. The sections were incubated
with a mixture of two monoclonal rat anti-mouse caspase-3
antibodies at a dilution of 20 mg/ml overnight at 4°C. Following
this, the sections were counterstained with hematoxylin (Sigma) and
mounted with aqueous mounting media. All the measurements were
performed in triplicate.
Histological images in the cerebral cortex overlying
the dentate gyrus of the hippocampus (20) were acquired by digital
photomicrography, using a light microscope under magnification,
x10-200. The tissue sections were coded. Histopathological
evaluation was carried out by two blinded pathologists at the
Second Hospital of Shandong University.
TEM
TEM analysis was used to assess the effects of VPA
on the radioprotection of rat brain microvascular. At an interval
of 6 months after irradiation, groups of rats were sacrificed under
deep chloral hydrate anaesthesia (400 mg/kg) for the other
sections. The brains were removed quickly and the regions of
interest on the cortex (overlying the dentate gyrus of the
hippocampus) (20) were removed
quickly and sliced into 1.0-mm sections prior to fixing in cold EM
grade 2.5% glutaraldehyde in 0.1 mol/l PBS (pH 7.3). Subsequently,
the sections were rinsed in PBS, postfixed in 1% OsO4
with 0.1% K3Fe(CN)6, dehydrated through a
graded series of EtOH and embedded in Epon (dodecenyl succinic
anhydride, nadic methyl anthydride, Scipoxy 812 Resin and
dimethylaminomethyl; Energy Beam Sciences, East Granby, CT, USA).
Ultrathin sections (65-nm) were cut and stained with 2% uranyl
acetate and Reynold's lead citrate, and examined on a JEM-1011 TEM
(Jeol, Tokyo, Japan). The sections were qualitatively analyzed for
the degree of radiation injury by blinded investigators, unaware of
the experimental condition.
Statistical analysis
For statistical analyses, Statistical Product and
Service Solutions software package (SPSS, Inc., Chicago, IL, USA)
was used. The statistical significance of the experimental data
among the groups was assessed with the Student's t-test. All the
values were expressed as mean ± standard deviation for n≥3.
P<0.05 in each experiment was considered to indicate a
statistically significant difference and statistical power was
calculated accordingly.
Results
IHC slide of the rat brain at the 15
Gy dose level
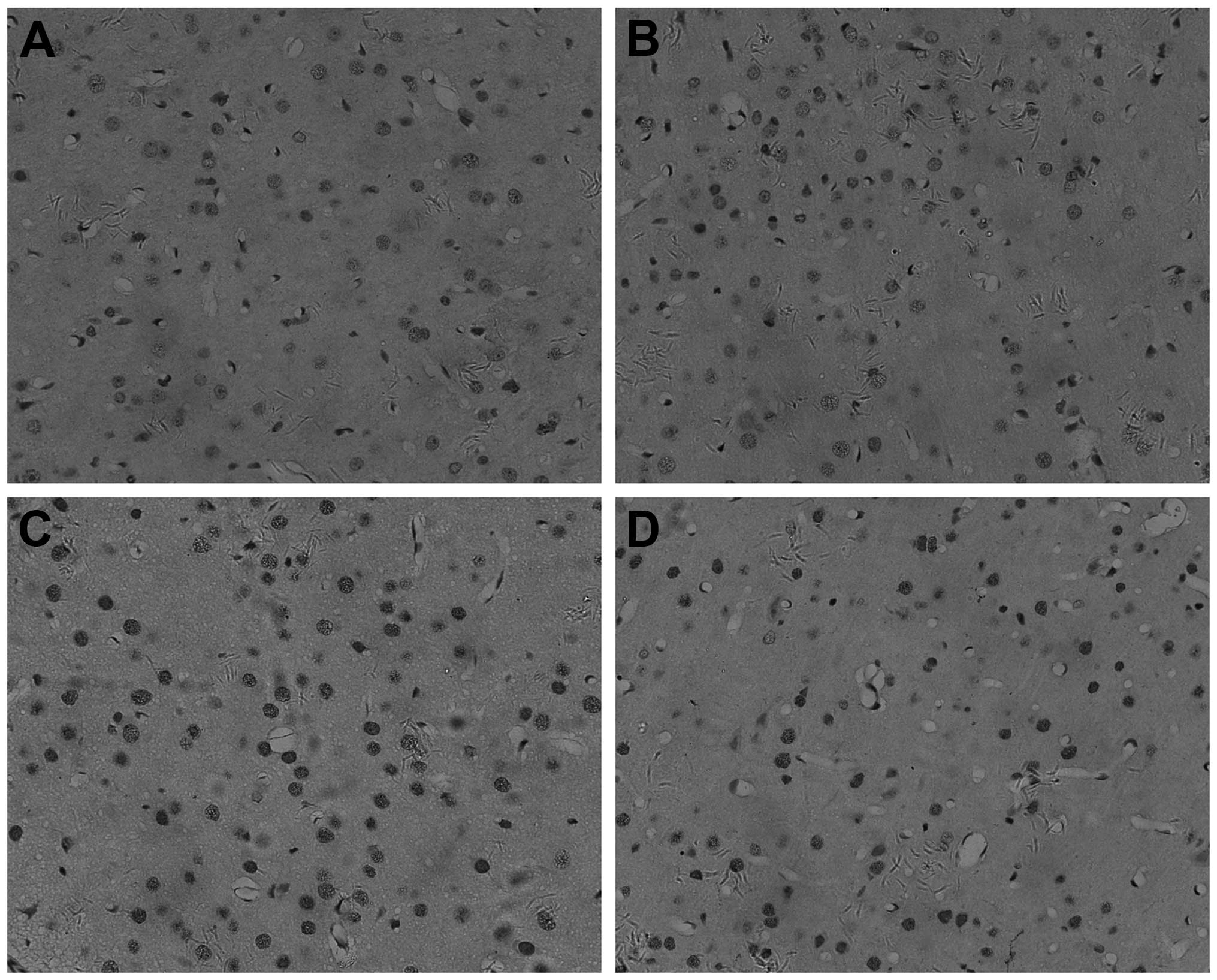
Cell apoptosis was quantified using a protein
caspase-3 and a morphological assessment of cell staining (original
magnification, x200). These sections were from the cerebral cortex
overlying the dentate gyrus of the hippocampus. Oligodendrocytes
and their processes stain dark brown (Fig. 2). By IHC, caspase-3 was found to be
present mainly in the nucleus of cells in the unirradiated cortex
of oligodendrocyte (Fig. 2A and B).
Following irradiation, the number of positive nuclei and the
intensity of the reaction product were greatly increased (Fig. 2C and D).
Apoptosis occurred in the cortex 1 day after
treatment, peaking in the X-ray group (Fig. 2C). The cells of the combined group
(Fig. 2D) showed moderate caspase-3
staining in the cytoplasm and nucleus. Caspase-3 was significantly
decreased in the combined group (Fig.
2D) compared to the X-ray group (Fig. 2C) in the cortex of the brain.
Caspase-3 had the lowest expression in the control (Fig. 2A) and VPA groups (Fig. 2B).
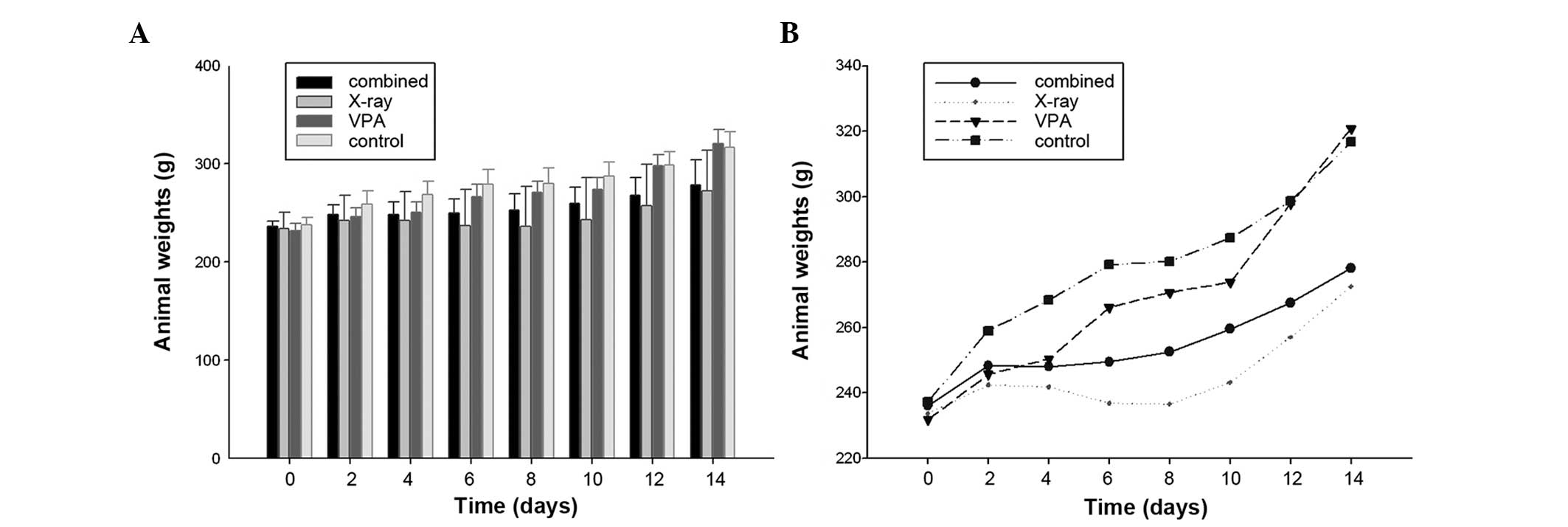
Animal weight throughout the course of
the experiment
Following the third injection of VPA, 15 Gy (3
Gy/day) was delivered locally to the brain over five days. The FRT
treatments were well tolerated by all the animals, as no apparent
adverse effects were observed in any group throughout the course of
the study. The animal weights of the TEM group over two weeks
starting from the injection of VPA were also observed. Following
VPA and irradiation, there were significant differences in the
average weight of the animals between the combined and control
groups, on day 6, 12 and 14 day (P<0.05) (Table I). By contrast, two days after
irradiation the average weight of the X-ray group was significantly
less than that of the controls (P<0.05) (Table I). There was a trend towards a lower
body weight in X-ray group following irradiation compared to either
no-irradiation or rats of the combined group (Fig. 3), although there was no significant
difference in the average weight between the combined group and
irradiated rats (Table I). However,
two days after VPA, the weight change of the average weight in the
combined group was characterized by a plateau from days 2–8 and a
steep increase following irradiation, compared to the X-ray group
(Fig. 3B). A tendency of an
increase for the average body weight of combined group at 15 Gy was
evident, but this difference was not statistically significant
(Fig. 3B)
 | Table IAnimal weights of the TEM group over
14 days starting from the first injection of valproic acid
(VPA). |
Table I
Animal weights of the TEM group over
14 days starting from the first injection of valproic acid
(VPA).
| Day |
|---|
|
|
|---|
| Group | 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 |
|---|
|
Combineda | 236.00±5.329 | 248.33±9.832 | 248.00±13.115 |
249.50±14.653b | 252.50±16.991 | 259.50±16.742 |
267.50±18.097b |
278.17±25.795b |
| X-raya | 233.67±17.072 | 242.33±25.586 |
241.83±29.404b |
236.83±37.156b |
236.50±40.134b |
243.17±42.466b |
257.00±42.506b |
272.50±41.215b |
| VPAa | 231.83±7.250 | 245.67±9.647 | 250.33±10.930 | 266.17±13.258 | 270.67±11.308 | 273.83±12.007 | 297.83±11.125 | 320.83±13.834 |
|
Controla | 237.33±7.866 | 259.00±13.446 | 268.33±14.109 | 279.17±15.342 | 280.17±15.715 | 287.33±14.404 | 298.67±13.909 | 316.67±16.182 |
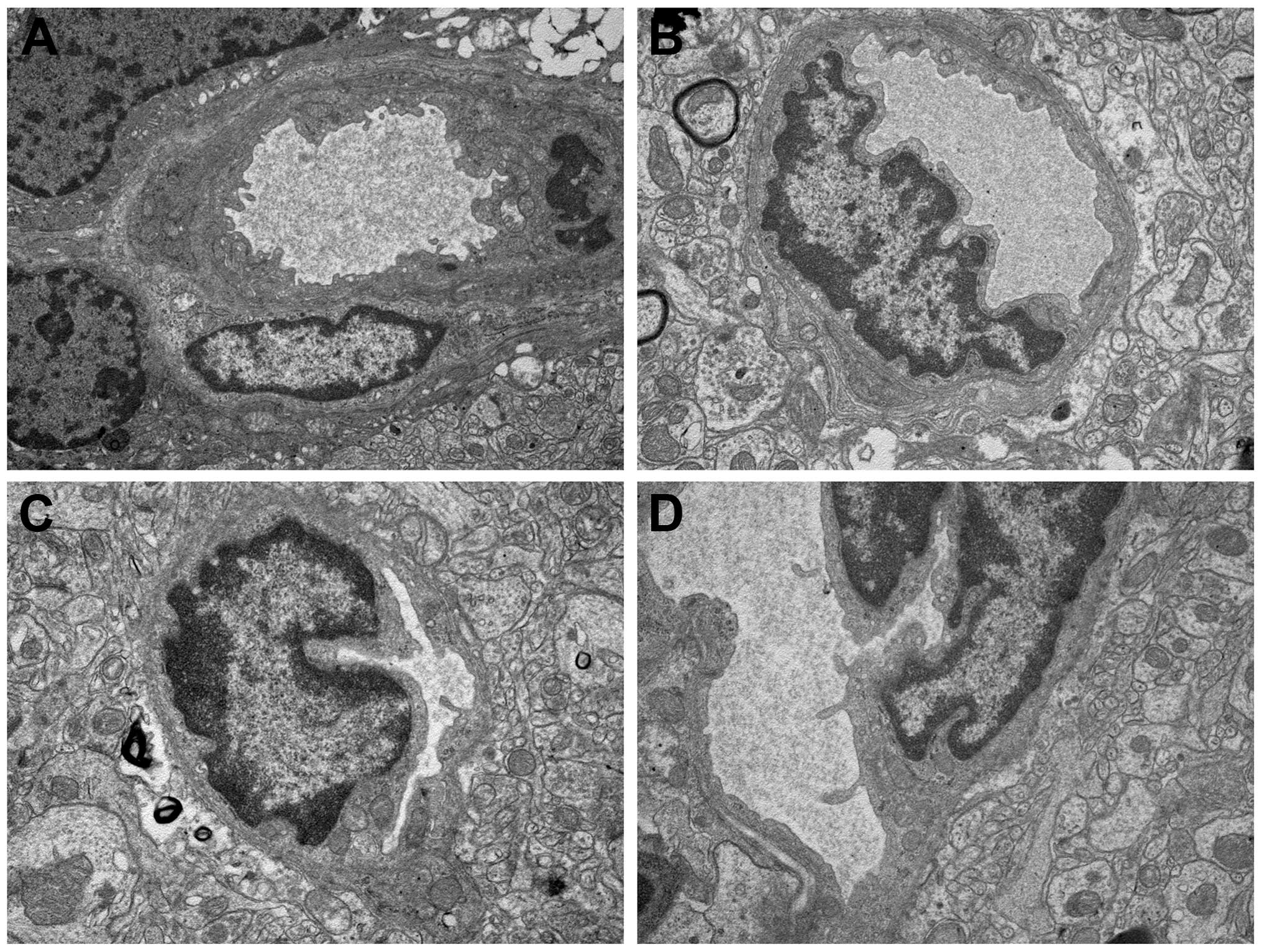
Observation of ultrastructure
TEM analysis was also used to assess the
radioprotection of VPA on the cortex cells of rat normal brain. The
development of vascular changes in the rat brain following a
moderate-dose FRT X-ray (15 Gy) was investigated. The results
indicate that, even with this dose, vascular abnormalities occur 6
months after irradiation. The X-ray group (Fig. 4C) showed a severe structural
disorder, loss and thickening of microvascular wall, hyaline
degeneration and fibrosis generally changed. Focal swelling of the
capillary endothelial cells can be detected with the TEM method in
the combined group (Fig. 4D). Mild
swelling of the capillary endothelial cells in the irregular lumen
was observed. However, the control (Fig. 4A) and VPA groups (Fig. 4B) exhibited normal vessel walls with
well-preserved tight junctions.
Discussion
In the present study, the dose to the brain was
purposely limited to 15 Gy. This dose level was selected on the
basis that the induced effects should be similar to the ones
observed in the human brain following a full course of FRT.
Moderate-dose fractionated X-ray irradiation did not appear to
affect the rat well-being. Whereas there is no doubt that a higher
dose will result in the earlier appearance of gross damage, the
underlying pathology may be quite different. This schedule was
designed to allow us to assess the acute and late effects of brain
irradiation. As there was no precedence to follow, 1 day and 6
months were chosen as an adequate interval to investigate the
changes induced by FRT. The experiments were terminated at 6 months
after the start of FRT, which were considered to be sufficient to
observe the late changes induced by radiation treatment.
Previous studies have reported that exposure to HDAC
inhibitors does not increase the radiosensitivity of various normal
tissue cell lines (12,23,24),
whereas certain HDAC inhibitors inhibit the repair of
radiation-induced DNA double-strand break and increase
radiosensitivity of normal cells, specifically lymphocytes
(25) and primary skin fibroblasts
in vitro (6). Additionally,
in vitro and in vivo data indicate that HDAC
inhibitors may protect normal tissue from radiation-induced side
effects (4,26).
Irradiation has been shown to induce apoptosis in
oligodendrocytes in vivo (27–30)
and in vitro (31).
Stress-induced oligodendrocyte apoptosis was mediated by caspase
activation (32). Caspases are
cysteine proteases that play a key role in cascade activation
during apoptosis induced by a number of stimuli (33–36).
In addition to cell death, the active caspase-3 plays a role in
normal cellular processes, including neuronal differentiation and
migration. The results of the study by Soane et al (37) indicate that upregulation of B-cell
lymphoma 2 protein and inhibition of caspase-3 activation are
potential mechanisms by which C5b-9 increases survival of
oligodendrocyte in vitro and possibly in vivo during
inflammation and immune-mediated demyelination affecting the CNS.
Although caspases, including caspase-3, are activated following
ionizing radiation, it has also been indicated that the
radiation-induced apoptosis is not dependent on them in the
developing CNS (38). However,
using primary cultures of neural precursor cells of the cortex from
developing rat brain, the study by Michelin et al (39) demonstrated the protection from
radiation-induced apoptosis by caspase-3 inhibitor is involved in
the apoptotic mechanism.
To investigate the signaling pathways implicated in
the radioprotection-induced apoptosis of the rat cortex cells in
the four groups, an IHC analysis was used to examine the caspase-3
protein levels. Subsequently, the present study demonstrated that
irradiation induces an evident increase of caspase-3 activity in
oligodendrocytes irradiated in vivo. In addition, caspase-3
was significantly decreased in the combined (Fig. 2D) compared to the X-ray group
(Fig. 2C) on the cortex of the
brain. The results of the study demonstrated that exposure to VPA
leads to a decrease in radiation-induce apoptosis in the rat
cortex. Caspase-3 had the lowest expression in the control
(Fig. 2A) and VPA groups (Fig. 2B). These results indicate an
important role of VPA in irradiation-induced apoptosis. As a
result, although quantitative studies to determine the proportions
of the different cellular types affected by X-irradiation and VPA
injection were not carried out, the present data indicate that VPA,
a HDAC inhibitor, developed a radioprotective effect from
radiation-induced apoptosis of in the rat cortex.
Oligodendrocytes have been shown to undergo
radiation-induced apoptosis in the CNS in vivo. An
understanding of the signalling pathways in mediating
radiation-induced apoptosis of oligodendrocytes may lead to
targeting strategies that protect the CNS from radiation injury
(30). Following radiation, cells
undergo rapid sphingomyelin hydrolysis and ceramide generation
followed by apoptosis (40).
Results of the study by Ferrer (41) suggest that X-irradiation enhances
naturally occurring cell death in the cerebral cortex and
subcortical white matter and that this process depends on the
activation of protein synthesis. On the basis of previous studies
regarding cell death (Fig. 4) in
vivo, one could expect that oligodendrocytes death caused by
X-ray would be inhibited after an i.p. injection of VPA.
Furthermore, the detection of caspase-3 induction in the cortex,
indicates that the cortex has an acute response to radiation.
Whether the expression of this stress protein contributes or
protects against the generation of the late radiation injury
remains to be determined. The cardinal features of radiation
exposure of the CNS are demyelination, gliosis and vascular damage
(42).
Following irradiation with a single dose of 8, 18 or
22 Gy, a peak response was observed at ~8 h after irradiation in
the rat spinal cord, and the apoptosis index returned to the levels
in non-irradiated oligodendrocytes at 24 h (28,43,44).
The present result (Fig. 2) showed
the activation of caspase-3 in irradiated oligodendrocytes 24 h
after IR. The fractionated irradiation dose of 15 Gy-induced
apoptosis was mediated by caspase-3 in oligodendrocytes.
Morphologically, apoptosis in 8-week-old rat cortex appeared to be
confined to oligodendrocytes cells and little evidence of apoptosis
was found in neurons and vascular endothelial cells. Vascular and
glial changes are significant in the development of late radiation
damage to the nervous system. Vascular effects occur at lower dose
levels, but following a longer latent period with effects mediated
through damage to neuroglia (45).
WBI of rats with 15 Gy X-ray has been reported to
suppress cerebral glucose utilization in 1 month (46) and decreased cerebrovascular volume
in ~1 year (47). Electron
microscopic examinations in the study by Kubota et al
(48) showed that injury in CNS has
initiated within 1 week after the application of radiation. The
present results indicated that even with the dose of 15 Gy FRT, the
X-ray group showed a severe structural disorder in 6 months. To a
certain extent, these findings are in accordance with a previous
study in which cerebral endothelial cells are more sensitive to
radiation damage than astrocytes (49). Mild swelling of capillary
endothelial cells in the irregular lumen could be observed in the
combined group. The findings indicate the protective effects of VPA
pretreatment from radiation injury of the CNS partially at this
dose. However, only a small local was observed in EM and further
studies are required to improve the effect of VPA on the treatment
of radiation injury. Subsequently, further study for ultrastructure
changes of rat nerve caused by X-ray is required.
These data indicate a possible association, not
necessarily causal, between the damage of the
microvascular/oligodendrocytes unit of tissue injury and
development of radiation-induced brain injury following the
exposure of rats to conventional fractionation (21). The average weight of the irradiated
animals was significantly lower than that of the sham-treated
animals when assessed 30 days from the start of FRT, with a daily
dose of 2 Gy delivered 5 days/week (total, 40 Gy) (20). In the present study, there was a
trend towards the lowest body weight in the X-ray group following
irradiation, when a 3 Gy daily dose was delivered 5 days/week
(total, 15 Gy). Compared to the X-ray group, a tendency of an
increase in body weight for the combined group was evident.
However, this difference was not statistically significant
(P>0.05). The observation time may be too short for assessment
of body weight change.
Following WBI, there is a sequence of cellular
events involving cell apoptosis and cerebrovascular change.
Although it is unclear how experimental results obtained from
animal models may be applied clinically, it is possible that such
results can be used together with theoretical models of
radioprotection. Animal models may also be used to test drug
therapies that may ameliorate radiation injury to normal brain.
In conclusion, the present results demonstrate that
VPA evidently provides radioprotection to the cortex of rats
against the X-ray injury. Thus, treatment with VPA in combination
with radiotherapy could be a noteworthy approach for protection of
the normal brain. Considering the surge of interest in
radioprotection, such results may be useful for improving future
clinical strategies of VPA. Although it is promising as a protector
of radiotherapy, further studies with VPA are warranted.
Acknowledgements
The present study was supported by the Shandong
Provincial Natural Science Foundation, China (grant no.
ZR2010HM080). We are grateful to the Central Research Laboratory
(The Second Hospital of Shandong University) for their technical
assistance and generous support.
References
|
1
|
Melian E: Radiation therapy in neurologic
disease. Handb Clin Neurol. 121:1181–1198. 2014. View Article : Google Scholar
|
|
2
|
Wong CS and Van der Kogel AJ: Mechanisms
of radiation injury to the central nervous system: implications for
neuroprotection. Mol Interv. 4:273–284. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brown SL, Kolozsvary A, Liu J, Ryu S and
Kim JH: Histone deacetylase inhibitors protect against and mitigate
the lethality of total-body irradiation in mice. Radiat Res.
169:474–478. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chung YL, Lee MY and Pui NN: Epigenetic
therapy using the histone deacetylase inhibitor for increasing
therapeutic gain in oral cancer: prevention of radiation-induced
oral mucositis and inhibition of chemical-induced oral
carcinogenesis. Carcinogenesis. 30:1387–1397. 2009. View Article : Google Scholar
|
|
5
|
Wu X, Chen PS, Dallas S, et al: Histone
deacetylase inhibitors up-regulate astrocyte GDNF and BDNF gene
transcription and protect dopaminergic neurons. Int J
Neuropsychopharmacol. 11:1123–1134. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Purrucker JC, Fricke A, Ong MF, Rube C,
Rube CE and Mahlknecht U: HDAC inhibition radiosensitizes human
normal tissue cells and reduces DNA double-strand break repair
capacity. Oncol Rep. 23:263–269. 2010.PubMed/NCBI
|
|
7
|
Peterson GM and Naunton M: Valproate: a
simple chemical with so much to offer. J Clin Pharm Ther.
30:417–421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Phiel CJ, Zhang F, Huang EY, Guenther MG,
Lazar MA and Klein PS: Histone deacetylase is a direct target of
valproic acid, a potent anticonvulsant, mood stabilizer, and
teratogen. J Biol Chem. 276:36734–36741. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gӧttlicher M, Minucci S, Zhu P, et al:
Valproic acid defines a novel class of HDAC inhibitors inducing
differentiation of transformed cells. EMBO J. 20:6969–6978.
2001.
|
|
10
|
Marks PA, Miller T and Richon VM: Histone
deacetylases. Curr Opin Pharmacol. 3:344–351. 2003. View Article : Google Scholar
|
|
11
|
Camphausen K, Cerna D, Scott T, et al:
Enhancement of in vitro and in vivo tumor cell radiosensitivity by
valproic acid. Int J Cancer. 114:380–386. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chinnaiyan P, Cerna D, Burgan WE, Beam K,
Williams ES, Camphausen K and Tofilon PJ: Postradiation
sensitization of the histone deacetylase inhibitor valproic acid.
Clin Cancer Res. 14:5410–5415. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Van Nifterik KA, Van den Berg J, Slotman
BJ, Lafleur MV, Sminia P and Stalpers LJ: Valproic acid sensitizes
human glioma cells for temozolomide and γ-radiation. J Neurooncol.
107:61–67. 2012.
|
|
14
|
Zhou Y, Xu Y, Wang H, Niu J, Hou H and
Jiang Y: Histone deacetylase inhibitor, valproic acid,
radiosensitizes the C6 glioma cell line in vitro. Oncol
Lett. 7:203–208. 2014.PubMed/NCBI
|
|
15
|
Lai JS, Zhao C, Warsh JJ and Li PP:
Cytoprotection by lithium and valproate varies between cell types
and cellular stresses. Eur J Pharmacol. 539:18–26. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Castro LM, Gallant M and Niles LP: Novel
targets for valproic acid: up-regulation of melatonin receptors and
neurotrophic factors in C6 glioma cells. J Neurochem. 95:1227–1236.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mizumatsu S, Monje ML, Morhardt DR, Rola
R, Palmer TD and Fike JR: Extreme sensitivity of adult neurogenesis
to low doses of X-irradiation. Cancer Res. 63:4021–4027.
2003.PubMed/NCBI
|
|
18
|
Zabolotskii NN, Onishchenko LS and Galeev
IS: Mitochondrial megaconia and pleioconia in the rat brain as
possible adaptive reactions in conditions of lethal radiation and
radiomodified lesions. Neurosci Behav Physiol. 30:497–501. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yildirim O, Comoğlu S, Yardimci S, Akmansu
M, Bozkurt G and Sürücü S: Preserving effect of melatonin on the
levels of glutathione and malondialdehyde in rats exposed to
irradiation. Gen Physiol Biophys. 27:32–37. 2008.PubMed/NCBI
|
|
20
|
Yuan H, Gaber MW, Boyd K, Wilson CM, Kiani
MF and Merchant TE: Effects of fractionated radiation on the brain
vasculature in murine model: blood-brain barrier permeability,
astrocyte proliferation, and ultrastructural changes. Int J Radiat
Oncol Biol Phys. 66:860–866. 2006. View Article : Google Scholar
|
|
21
|
Cicciarello R, d'Avella D, Gagliardi ME,
et al: Time-related ultrastructural changes in an experimental
model of whole brain irradiation. Neurosurgery. 38:772–780. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vinchon-Petit S, Janet D, Jadaud E,
Feuvret L, Garcion E and Menei P: External irradiation models for
intracranial 9L glioma studies. J Exp Clin Cancer Res. 29:1422010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Munshi A, Kurland JF, Nishikawa T, et al:
Histone deacetylase inhibitors radiosensitize human melanoma cells
by suppressing DNA repair activity. Clin Cancer Res. 11:4912–4922.
2005. View Article : Google Scholar
|
|
24
|
Blattmann C, Oertel S, Ehemann V, et al:
Enhancement of radiation response in osteosarcoma and
rhabdomyosarcoma cell lines by histone deacetylase inhibition. Int
J Radiat Oncol Biol Phys. 78:237–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stoilov L, Darroudi F, Meschini R, van der
Schans G, Mullenders LH and Nararajan AT: Inhibition of repair of
X-ray-induced DNA double-strand breaks in human lymphocytes exposed
to sodium butyrate. Int J Radiat Biol. 76:1485–1491. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chung YL, Wang AJ and Yao LF: Antitumor
histone deacetylase inhibitors suppress cutaneous radiation
syndrome: Implications for increasing therapeutic gain in cancer
radiotherapy. Mol Cancer Ther. 3:317–325. 2004. View Article : Google Scholar
|
|
27
|
Li YQ, Jay V and Wong CS: Oligodendrocytes
in the adult rat spinal cord undergo radiation-induced apoptosis.
Cancer Res. 56:5417–5422. 1996.PubMed/NCBI
|
|
28
|
Chow BM, Li YQ and Wong CS:
Radiation-induced apoptosis in the adult central nervous system is
p53-dependent. Cell Death Differ. 7:712–720. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Atkinson SL, Li YQ and Wong CS: Apoptosis
and proliferation of oligodendrocyte progenitor cells in the
irradiated rodent spinal cord. Int J Radiat Oncol Biol Phys.
62:535–544. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu FG and Wong CS: Radiation-induced
apoptosis of oligodendrocytes and its association with increased
ceramide and down-regulated protein kinase B/Akt activity. Int J
Radiat Biol. 80:39–51. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vrdoljak E, Bill CA, Stephens LC, van der
Kogel AJ, Ang KK and Tofilon PJ: Radiation-induce apoptosis of
oligodendrocytes in vitro. Int J Radiat Biol. 62:475–480. 1992.
View Article : Google Scholar
|
|
32
|
Gu C, Casaccia-Bonnefil P, Srinivasan A
and Chao MV: Oligodendrocyte apoptosis mediated by caspase
activation. J Neurosci. 19:3043–3049. 1999.PubMed/NCBI
|
|
33
|
Lin HI, Lee YJ, Chen BF, et al:
Involvement of Bcl-2 family, cytochrome c and caspase 3 in
induction of apoptosis by beauvericin in human non-small cell lung
cancer cells. Cancer Lett. 230:248–259. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim DS, Jeon SE, Jeong YM, Kim SY, Kwon SB
and Park KC: Hydrogen peroxide is a mediator of indole-3-acetic
acid/horseradish peroxidise-induced apoptosis. FEBS Lett.
580:1439–1446. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zuliani T, Obriot H, Tual M, et al:
Variable Bax antigenicity is linked to keratinocyte position within
epidermal strata and UV-induced apoptosis. Exp Dermatol.
17:125–132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Park SK, Kang H and Kwon CH:
Caspase-dependent cell death mediates potent cytotoxicity of
sulfide derivatives of 9-anilinoacridine. Anticancer Drugs.
19:381–389. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Soane L, Rus H, Niculescu F and Shin ML:
Inhibition of oligodendrocyte apoptosis by sublytic C5b-9 is
associated with enhanced synthesis of bcl-2 and mediated by
inhibition of caspase-3 activation. J Immunol. 163:6132–6138.
1999.PubMed/NCBI
|
|
38
|
Ferrer I: Role of caspases in ionizing
radiation-induced apoptosis in the developing cerebellum. J
Neurobiol. 41:549–558. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Michelin S, del Rosario Perez M, Dubner D
and Gisone P: Increased activity and involvement of caspase-3 in
radiation-induced apoptosis in neural cells precursors from
developing rat brain. Neurotoxicology. 25:387–398. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mathias S, Pena LA and Kolesnick RN:
Signal transduction of stress via ceramide. Biochem J. 335:465–480.
1998.PubMed/NCBI
|
|
41
|
Ferrer I: The effect of cycloheximide on
natural and X-ray-induced cell death in the developing cerebral
cortex. Brain Res. 588:351–357. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Brush J, Lipnick SL, Phillips T, Sitko J,
McDonald JT and McBride WH: Molecular mechanisms of late normal
tissue injury. Semin Radiat Oncol. 17:121–130. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li YQ, Guo YP, Jay V, Stewart PA and Wong
CS: Time course of radiation-induced apoptosis in the adult rat
spinal cord. Radiother Oncol. 39:35–42. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li YQ and Wong CS: Radiation-induced
apoptosis in the neonatal and adult rat spinal cord. Radiat Res.
154:268–276. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hopewell JW: Late radiation damage to the
central nervous system: a radiobiological interpretation.
Neuropathol Appl Neurobiol. 5:329–343. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ito M, Patronas NJ, Di Chiro G, Mansi L
and Kennedy C: Effect of moderate level X-radiation to brain on
cerebral glucose utilization. J Comput Assist Tomogr. 10:584–588.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Keyeux A: Late modifications of cephalic
circulation in head x-irradiated rats. Radiat Environ Biophys.
13:125–135. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kubota Y, Takahashi S, Sun XZ, Sato H,
Aizawa S and Yoshida K: Radiation-induced tissue abnormalities in
fetal brain are related to apoptosis immediately after irradiation.
Int J Radiat Biol. 76:649–659. 2000. View Article : Google Scholar
|
|
49
|
Fike JR, Gobbel GT, Chou D, Wijnhoven BP,
Bellinzona M, Nakagawa M and Seilhan TM: Cellular proliferation and
infiltration following interstitial irradiation of normal dog brain
is altered by an inhibitor of polyamine synthesis. Int J Radiat
Oncol Biol Phys. 32:1035–1045. 1995. View Article : Google Scholar : PubMed/NCBI
|