Introduction
The dendritic cell (DC) tumor vaccine is currently
one of the most common active immunity strategies for tumor
treatment. The use of DCs in vitro and following antigen
stimulation, as a therapeutic cancer vaccine induced specific
immune responses and the corresponding tumor antigens have been
investigated in a large number of animal experiments and confirmed
in vitro, as well as in numerous clinical trials (1–3). Culture of
DCs in vitro is achieved with DC precursor cells, such as
cluster of differentiation (CD)14+ cells, that
differentiate into immature DCs via stimulation with
granulocyte-macrophage colony-stimulating factor (GM-CSF) and
interleukin-4 (IL-4). The preparation of DC as a specific tumor
vaccine is completed upon antigen stimulation and DC maturation.
The induction of DC maturation is particularly important and an
active tumor immune response may only be produced in mature DCs
(4), with immature DCs leading to
immune tolerance (5). Currently the
most common method for inducing DC maturation in the clinical
setting is incubation for 24–48 h with a variety of cytokines,
including IL-1, IL-6, tumor necrosis factor (TNF)-α and
prostaglandin E2 (PGE2) in immature DCs (6); however, this method is time-consuming and
costly. In addition, the gene transduction method is often adopted
to modify DCs to enhance their ability to stimulate the production
of tumor-specific cytotoxic T lymphocytes (CTLs). Furthermore, one
of the most commonly used methods is infecting DCs with adenovirus
vectors to express various genes (7).
The current study established a chimeric adenovirus (Ad) 5/F35
mosaic type adenoviral vector co-expressing CD40 ligand (CD40L) and
IL-2, which was able to efficiently infect human monocyte-derived
DCs (Mo-DC) and induced the maturation of Mo-DC and IL-12.
Materials and methods
Reagents and materials
EcoRI, BamHI, NcoI and
XhoI double-enzyme were purchased from Takara Bio, Inc.
(Otsu, Japan), husion® High-Fidelity DNA Polymerase and
T4 DNA ligase were purchased from New England BioLabs, Inc.
(Ipswich, MA, USA), and a ThermoScript™ kit, liposome Lipofectamine
2000™ and the eukaryotic expression vectors, pcDNA3.1/myc-His(−)B
were purchased from Invitrogen (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The antibodies for flow cytometry (CD80-PE (cat.
no. 560925; clonality, L307.4; mouse, mouse IgG1; dilution, 1:20),
CD86-PE (cat. no. 550616; clonality, MOPC-31C; mouse; mouse IgG1;
dilution, 1:20), CD83-PE (cat. no. 557720; clonality, R3-34; mouse;
mouse IgG1; dilution, 1:20), CD40-PE (cat. no. 560963; clonality,
5C3; mouse, mouse IgG1; dilution, 1:20), CD40L-PE (cat. no. 552559;
clonality, hCD40L-M90.1; mouse, mouse IgG1; dilution, 1:20), and
human leukocyte antigen-antigen D related (HLA-DR)-PE (cat. no.
562804; clonality, G46-6; mouse, mouse IgG1; dilution, 1:20)) were
purchased from BD Pharmingen (San Diego, CA, USA), and the RNA
extraction kit (RNeasy Mini kit) was purchased from Qiagen, Inc.,
(Valencia, CA, USA). Plasmid extraction, purification and gel
extraction kits were purchased from Omega Bio-Tek, Inc. (Norcross,
GA, USA) and The Cre-lox system recombinant adenovirus systems were
purchased from Microbix Biosystems Inc. (Mississauga, ON, Canada).
The IL-2 and IL-12 enzyme-linked immunosorbent assay (ELISA) kits
were purchased from Diaclone SAS (Besançon, France) and all the
culture mediums and fetal bovine serum (FBS) were purchased from
Gibco (Thermo Fisher Scientific, Inc.). Lymphocyte Separation
Medium was purchased from Haoyang Biological Technology Co., Ltd.
(Tianjin, China).
Cloning of human (h)CD40L and
IL-2
Total RNA was extracted using RNA extraction kits
following isolation of peripheral blood mononuclear cells (PMBCs)
with Lymphocyte Separation Medium, according to the manufacturer's
instructions. The first strand cDNA was synthesized according to
the instructions of the ThermoScript™ kit. According to the GenBank
registration accession numbers of CD40L and hIL-2 (NM_000074 and
NM_000586, respectively), two pairs of primers were designed during
the present study as follows: Upstream, 5′-gga att cgc cat ggt cga aac ata
caa cc-3′ (the EcoRI site was drawn from the 5′point, and
the restriction sites are underlined) and downstream,
5′-cgg gat cct cag
agt ttg agt aag cc-3′ (the BamHI site was drawn from the
5′point) for CD40L; upstream, 5′-cgg gat cca tgg aca gga tgc aac
tc-3′ (the BamHI site was drawn from the 5′point) and
downstream, 5′-ccc aag ctt caa gtc agt gtt gag atg atg-3′ (the
HindIII site was drawn from the 5′point) for IL-2. The
amplified fragments were inserted into the restriction enzyme sites
of the pcDNA3.1/myc-His(−)B eukaryotic expression carrier, and the
pcDNA3.1(−) CD40 and pcDNA3.1(−) IL-2 were constructed separately
to demonstrate the success of the cloned sequence.
Construction and amplification of the
Ad5/F35 CD40L-IL-2 adenovirus vector
In order to construct the co-expression adenovirus
vector of CD40L and IL-2, construction of pDC315 CD40L-IL-2, the
co-expressing adenovirus vector carrier of CD40L and IL-2-shuttle
plasmid, was required. The CD40L fragment was sliced from the
pcDNA3.1(−)CD40L, using the EcoRI and BamHI
double-enzyme, and inserted into the corresponding restriction
sites of the Adenovirus Shuttle Plasmid, pDC315 (Microbix
Biosystems Inc.) to constitute the pDC315 CD40L. A pair of IL-2
primers was designed for the construction of IL-2, which is
connected with the internal nucleic acid access site (internal
ribosome entry site; IRES). Upstream, 5¢-cat gcc atg gac agg atg
caa ctc c-3¢ (the NcoI site was drawn from the 5¢ point) and
downstream, 5¢-ccg ctc gag tca agt cag tgt tga gat-3¢ (the
XhoI site was drawn from the 5¢ point). The IL-2 fragment
was amplified by polymerase chain reaction (PCR) from the
pcDNA3.1(−) IL-2 plasmid using NcoI and XhoI
double-enzyme, and the NcoI and XhoI double-enzyme
was also used in the pmRNA IRES-luc vector (8). The fragment of IRES pmRNA was recovered
by gel electrophoresis, expression of CD40L and phenotype changes
of Mo-DCs infected with Ad5/F35 CD40L-IL-2 were analyzed using flow
cytometry, and pmRNA IRES-IL-2 was directly constructed from the
connection of IL-2 and pmRNA IRES. The IRES-IL-2 fragment was
recovered by gel electrophoresis (140 V for 20 min) using the
BamHI and XhoI double-enzyme and inserted into the
corresponding sites of pDC515 GM-CSF to form pDC315 CD40L-IRES-IL-2
(abbreviated as pDC315 CD40L- IL-2).
The packaging of the Ad5/F35 CD40L-IL-2 adenovirus
was performed by Beijing Benyuan Zhengyang Gene Technology Co.,
Ltd. (Beijing, China) and, subsequently, the virus seed was
obtained and identified by PCR (using pDC315 (Beijing Benyuan
Zhengyang Gene Technology Co., Ltd.) murine cytomegalovirus, Simian
virus 40 (1 µg/µl; 10 µl) and a LabCycler thermocycler under the
following conditions: 94°C for 30 sec, followed by annealing (64°C
for 30 sec) and extension (72°C for 30 sec) for 30 cycles). The 293
cells infected with the virus seed were then amplified by cell
culture (at 37°C for 72 h). Following freeze-thaw lysis of the 293
collected cells, the purified Ad GM-CSF-IL-2 was acquired through
CsCl gradient centrifugation, which was performed twice. The TCID50
method to quantify IL-2 and IL-12 in the supernatants of Mo-DCs
following infection of Ad5/F35 CD40L-IL-2 was used to identify the
virus titer.
Culture of human monocyte-derived
DC
Peripheral blood samples (20 ml) were obtained from
a healthy donors (one male, aged 36-years-old; written informed
consent was obtained and the study was ethically approved by the
review board of the Beijing Military General Hospital) and human
PBMCs were isolated using lymphocyte separation medium and density
gradient centrifugation. The adhesion method was adopted to culture
the DCs, according to the method described in a previous study
(9). Briefly, PBMCs were suspended
with RPMI-1640 complete medium (RPMI-1640 medium with 10% FBS) and
added to a 175-cm2 disposable plastic culture flask
(2×106/ml). The non-adherent cells and medium were
extracted after 1 h, and 20 ml complete medium containing RPMI-1640
supplemented with 800 U/ml GM-CSF and 250 U/ml IL-4 was added.
After three days of culture at 37°C, the above 20 ml GM-CSF and
IL-4 medium was supplemented. After a further five days of culture
at 37°C, phenotypic identification or detection for adenovirus
infection were conducted.
DC infection with adenovirus
vectors
DCs were cultured at 37°C for five days, and the
cells were suspended with RPMI-1640 complete medium containing
GM-CSF and IL-4, and collected by centrifugation (100 × g at 37°C
for 5 min). The cell number was adjusted to 5×105
cells/ml and added to a 6-well plate (3 ml/well). The Ad5/F35
CD40L-IL-2 or Ad5/F35 EGFP (10) was
added (viral titer, 100 plaque-forming unit (pfu)/cell), and 2 h
after infection, the cells were centrifuged (100 × g at 37°C for 5
min) and collected. The cells were placed into the 6-well plate
following suspension with RPMI-1640 complete medium containing
GM-CSF and IL-4. The supernatant was collected and frozen at −20°C
until subsequent ELISA testing, after 6, 12 and 24 h, and phenotype
testing was performed when the DCs were collected after 24 h. Eight
DCs were infected with Ad5/F35 enhanced green fluorescent protein
(EGFP) and fixed with 4% polyformaldehyde. The cells were analyzed
under a fluorescence microscope to observe GFP expression.
Flow cytometric analysis
Flow cytometry was performed to identify DC
phenotypes and assess the infection efficiency of the adenovirus
vector in the DCs. The cells were collected, centrifuged (100 × g
for 5 min) and washed with phosphate-buffered saline (PBS) once.
Following an additional centrifugation (100 × g for 5 min), if
adenovirus infection efficiency was to be detected, this was
analyzed following resuspension with 400 µl PBS. However, for
phenotypic identification, a monoclonal antibody was added to the
cells and incubated for 30 min in the dark at room temperature, and
then washed two times with PBS. Subsequent to resuspension, the
cells were exposed to Lm400 PBS, and flow cytometry was performed
using a Beckman Coulter four-color flow cytometer (Coulter Epics
XL; Beckman Coulter, Inc., Brea, CA, USA).
ELISA detection
After thawing of the culture medium, which was
diluted five times, it was collected at each time-point (6, 12 and
24 h), and the level of IL-2 and IL-12 protein in the supernatant
was detected according to the instructions of the ELISA kit.
Statistical analysis
Continuous variables are presented as means ±
standard deviation and 95% confidence interval for the descriptive
statistics. Differences between the two groups were assessed by
Student's t-test for continuous variables. For comparison of
percentages (gender, multiorgan dysfunction syndrome, antibiotic
substance classes and mortality) the χ2 test was
employed using SPSS 13.0 statistical software (SPSS, Inc. Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Cloning of the CD40L and IL-2
genes
Total RNA that was obtained from healthy human PBMCs
was synthesized to the first strand cDNA by reverse transcription.
Two fragments of ~700 and 450 bp size, were obtained by PCR
amplification of the CD40L and IL-2 primers, and was inserted to
the corresponding locus of the eukaryotic expression vector,
pcDNA3.1/myc-His (−) B after a restriction enzyme fragment, then
identified by the corresponding primers. The positive clones were
sequenced by Shenggong (Beijing) Biological Technology Co., Ltd.
(Beijing, China), and it was confirmed that the sequence of the
CD40L and IL-2 clones were the same as CD40L (serial number,
NM_000074; 708 bp) and IL-2 (serial number, NM_000586 (462 bp)
according to GenBank.
Construction of Ad5/F35
CD40L-IL-2
CD40L was subcloned into an adenoviral shuttle
plasmid, pDC315, and then the IRES-IL-2 fragment was inserted into
the GM-CSF, so that the CD40L and IL-2 were connected in series
through the IRES sequence, and under the regulation and promoter of
the same cytomegalovirus (CMV). The constructed adenoviral shuttle
plasmid, pDC315 CD40L-IL-2 was packaged to produce the virus within
adenovirus packaging, and the virus DNA was extracted for
subsequent identification. PCR amplification was performed using
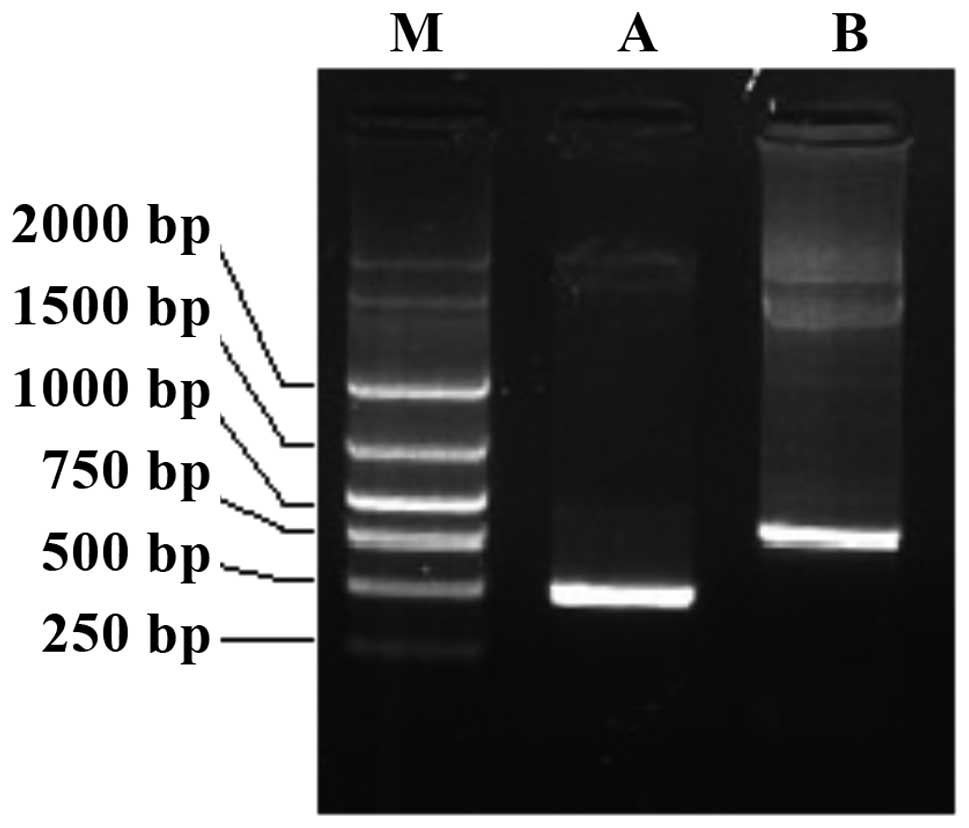
clone primers of CD40L and IL-2, and two fragments of the same size
as the CD40L and IL-2 gene fragments were obtained (Fig. 1). This demonstrated that CD40L and IL-2
were in the DNA adenovirus genome. Furthermore,
1010-1011 pfu viral titers were obtained by
repeated amplification of virus seeds in 293 cells and purification
by CsCl gradient centrifugation.
Phenotypic characteristics of
Mo-DCs
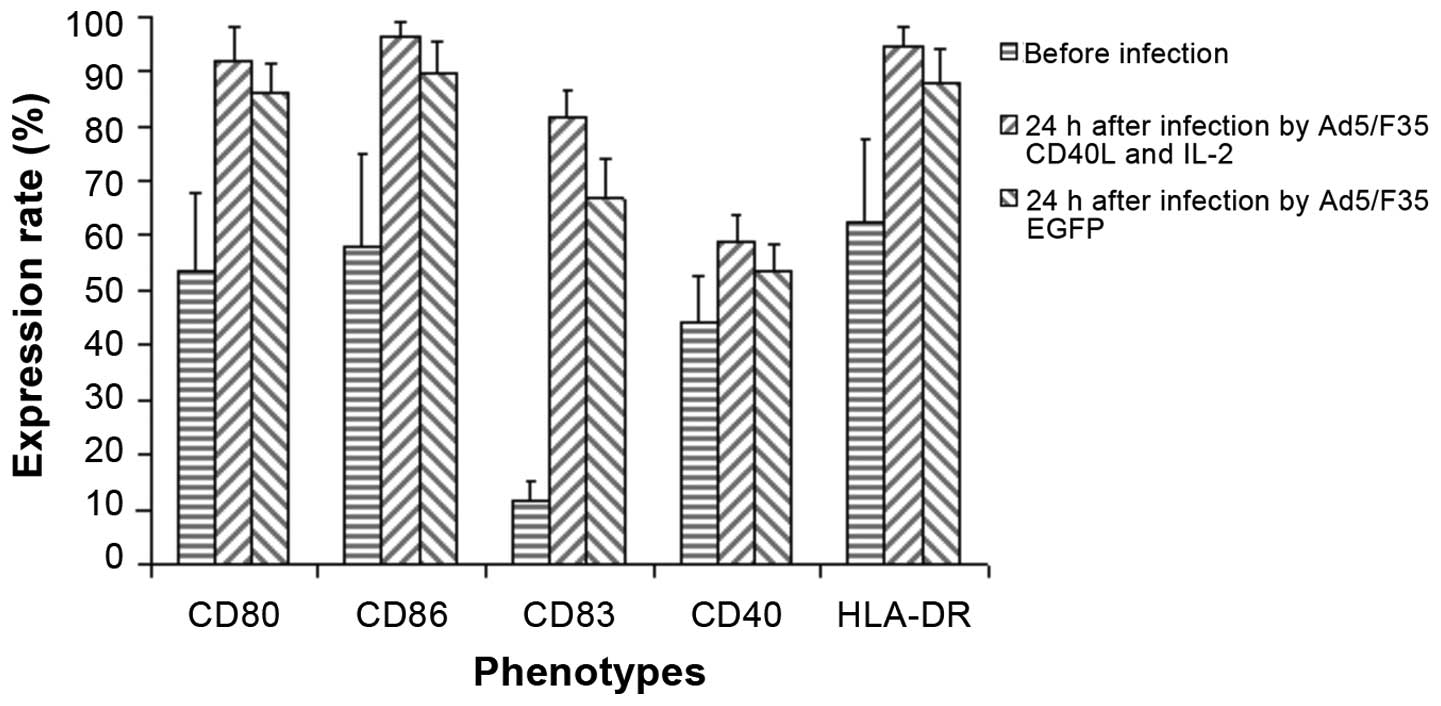
DCs were cultured for five days and stained with
CD86, CD80, CD83, CD40 and HLA-DR antibodies. The results indicated
that following five days of culturing, DCs demonstrated the
immature phenotype, with moderate expression (>50%) of
co-stimulatory molecules, CD80, CD86 and CD40 and major
histocompatibility complex (MHC) II molecule, HLA-DR and low
expression (<20%) of CD83 (Fig.
2).
Infection efficiency of Ad5/F35
adenovirus vector in Mo-DCs
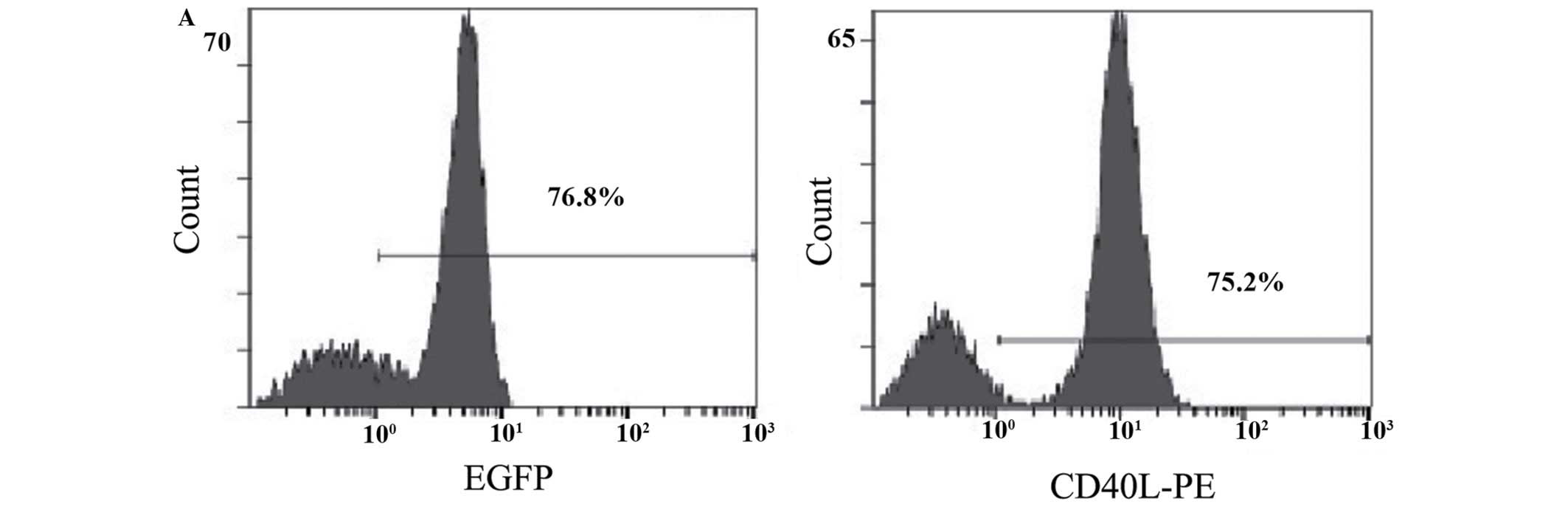
The expression efficiency of EGFP was detected by
flow cytometry after 24 h of infection of Mo-DC by Ad5/F35 EGFP or
Ad5/F35 CD40L-IL-2 (virus titer, 100 pfu/cells), and the results
indicate that the infection efficiency of Ad5/F35 EGFP to Mo-DC was
76.8%. The expression of CD40L was detected by flow cytometry and
the expression rate was identified to be 75.2%, which was similar
to EGFP (Fig. 3A). The green
fluorescence was observed under a fluorescence microscope (Fig. 3B).
Maturation of Mo-DC by induction of
Ad5/F35 CD40L-IL-2 and production of IL-12
The phenotypic changes of Mo-DC were detected
through flow cytometry 24 h after infection by Ad5/F35 EGFP or
Ad5/F35 CD40L-IL-2 (virus titer, 100 pfu/cells), and the results
indicated that the expression of CD86, CD80, CD83, CD40 and HLA-DR
was increased; however, after 24 h, when DCs were infected
CD40L-IL-2, the expression levels of CD86, CD80, CD83, CD40 and
HLA-DR were increased more markedly (Fig.
3). This indicated that activation and maturation of DCs may be
induced by autoinfection of Mo-DC, although phenotypic changes were
more obvious when CD40L was expressed by DCs and DCs were trending
towards a mature phenotype. ELISA was used to detect the contents
of IL-2 and IL-12 in the cell supernatant at different time-points
(6, 12 and 24 h) subsequent to Mo-DC infection with Ad5/F35
CD40L-IL-2 or Ad5/F35 EGFP (Fig. 4A).
The expression of IL-12 was detected 6 h after Mo-DCs were infected
with CD40L-IL-2 and the expression continued to rise after 24 h.
Although production of IL-12 was induced by infection with pure
Ad5/F35 EGFP and exhibited a similar mode of secretion, the IL-12
secretion was significantly lower (P<0.05) when compared with
the secretion from Mo-DCs after infection with Ad5/F35 CD40L-IL-2;
the difference between the two was 2–8 times. In addition,
following infection of Mo-DCs with Ad5/F35 CD40L-IL-2, high levels
of IL-2 were detected in the culture supernatant; however, Mo-DCs
infected with Ad5/F35 EGFP only exhibited low levels of expression
(Fig. 4B).
Discussion
Tumor immunotherapy based on DCs is currently a
popular tumor immunotherapeutic strategy. Subsequent to antigen
stimulation by the tumor, and after being stimulated by tumor
associated antigens, DCs process the antigens and deliver them to
the T cells, and the initial (naïve) T lymphocytes differentiate
into CTLs, which recognize and kill the corresponding tumor cells.
CTL production by DCs require at least three signals (11): Tumor antigens, in the form of
MHC-antigen peptide complex (the first signal), processed and
delivered by DCs, which stimulate T cells, including
CD4+ and CD8+ T lymphocytes. Co-stimulatory
molecules of DC surface expression, such as CD80, CD86 and CD40 are
the second signal and interact with co-stimulatory molecule ligand
ions of T cell surface expression. The activated T cells, DCs and
CD4+ T cells then secrete various cytokines (the third
signal), including Th1 cytokines, such as IL-2, IL-12 and TNF-PGE2
that promote significant cell proliferation of responsive CD8+
T-lymphocytes, and thereby CTLs with tumor antigen specificity are
produced. DC culturing in vitro is induced and
differentiated mostly from the peripheral blood monocytes, CD14+ by
GM-CSF and IL-4; however, DCs induced by these two types of
cytokine are of the immature phenotype, which express low levels of
co-stimulatory and MHC II molecules. Furthermore, these immature
DCs promote organisms to produce specific antitumor immunity
reactions as a tumor vaccine to stimulate the body to produce
specific antitumor immune response. Bacterial lipopolysaccharides
(LPSs) have been used to induce maturation of DCs in in
vitro studies and animal models (12). In a previous study, DCs were incubated
with a variety of cytokines, including IL-1, IL-6, TNF-PGE2 for
24–48 h (3); however, this method is
time-consuming and costly. The interaction of CD40 and CD40L is
known to be particularly important; the functions of DCs in
vivo and the interaction of CD40L with the expression of
CD4+ and CD40 on the surface of DCs is the condition
required to produce tumor specific CTL with DC stimulation
(13). Previous studies found that an
effective specific antitumor immune response is produced by DCs
independent of CD4+ with the stimulation of anti-CD40
monoclonal antibody or CD40L (14,15). DCs
infected by adenovirus carriers (16,17) and
expressing CD40L effectively induced an antitumor immune response
in animal experiments, and the induction of the antitumor immune
response was associated with DC maturation induced by CD40L and the
production of various cytokines (18,19),
particularly IL-12 (20). Certain DCs,
which are of the mature phenotype, but cannot produce IL-12, are
occasionally referred to as half-mature DCs, while only DCs of the
mature phenotype and producing IL-12 are termed mature DCs. Only
the mature DCs effectively induce an antitumor immune response
(21).
The present study hypothesized that infecting DCs
derived from PBMCs with a chimeric adenovirus 5/F35 carrier
co-expressing CD40L and IL-2 induces mature phenotype DCs,
producing large quantities of IL-12. The present study has the
following characteristics: First, the co-expression of CD40L and
IL-2 was demonstrated to enhance the antitumor effect. Consistent
with this, CD40L expressed alone with an adenovirus vector has been
demonstrated to effectively induce an antitumor immune response
(22); however, marked proliferation
of CTLs requires the stimulation of a variety of lymphocyte
proliferation factors following production of tumor specific CTLs
by DC stimulation, particularly IL-2. However, DCs and
CD4+ T lymphocytes also produce certain quantities of
IL-2, although exogenous expression of IL-2 exerts more stable and
durable functions. A previous study showed that CD40L and IL-2
exert a congenerous antitumor effect (23), and Rousseau et al applied these
results from the in vitro and animal experiments into a
clinical setting in which a mixture of primary cultured skin
fibroblasts infected with Ad IL-2 and Ad CD40L, and autologous
leukemia cells were administered by intradermal injection. The
pertinent leukemia cell responses of the patients were then
monitored (24). CD40L and IL-2 are
co-expressed in the same carrier, and in future applications it may
be possible to reduce the number of steps (by only producing a
carrier) and reduce the cost. In addition, the maturation time of
DCs in vitro may be significantly shortened following the
infection of DCs with the adenovirus vector, as the process of DC
maturation induced by CD40L was constant and did not change when
DCs were infected by adenovirus in vivo or in vitro.
Therefore, DCs may be inoculated following adenovirus infection,
and the maturation process of DCs may be completed in vivo,
rather than culturing for 24–48 h in vitro, as is necessary
when inducing DC maturation with cytokines.
In addition, hematopoietic system cells, including
DCs, may be used as carriers of chimeric adenovirus 5/F35, as it is
possible to efficiently infect these cells. It was known that
adenovirus type 5 from the hematopoietic system, including
dendritic cells, have a low infection efficiency, as the cells of
hematopoietic origin exhibit low Coxsackie adenovirus receptor
(CAR) expression or a complete lack of CAR (25), and the infection of adenovirus type 5
into cells is dependent on the receptor. The mechanism of the
chimeric adenovirus 5/F35 vector entering into cells involves
passage through CD46, and the molecule is commonly expressed in all
types of cells, including in the hematopoietic system (26). Chimeric adenovirus 5/F35 vector was
effectively infected into human Mo-DCs (27), which was consistent with the findings
of the present study.
Furthermore, the adenovirus vector encoded two genes
simultaneously, and the two genes, CD40L and IL-2, were connected
by the internal nucleic acid access site (IRES) sequence, and
controlled by CMV promoter regulation, resulting in the efficient
expression of the two genes. IRES is a commonly used method for
producing double gene expression vectors (28,29);
however, the expression levels of the two genes connected via the
IRES is not completely consistent, and often the gene expression
level close to the promoter is high, while the gene expression
level downstream of the IRES is low (30). In conclusion, the present study
indicates that CD40L and IL-2 connected by IRES may be highly
expressed, and IRES and IL-2 may be connected by NcoI locus
(ccatgg), with the Kozak sequence (accatgg) being formed in favor
of the expression of IL-2. The present study demonstrates the
connection between IRES CD40L and IL-2 may lead to efficient
expression. The IRES and IL-2 are connected by the NcoI
sites and form the Kozak Sequence (accatgg), which is conducive to
increased IL-2 expression. Ad5/F35 CD40L-IL-2 is efficiently
infected into human Mo-DCs and expresses CD86, CD80, CD83, CD40 and
HLA-DR genes. In addition, Ad5/F35 CD40L-IL-2 stimulates the
maturation of Mo-DCs and results in high levels of IL-12
production.
References
|
1
|
Cun Y, Zhang Q, Xiong C, Li M, Dai N,
Zhang S and Wang D: Combined use of adenoviral vector
Ad5/F35-mediated APE1 siRNA enhances the therapeutic efficacy of
adenoviral-mediated p53 gene transfer in hepatoma cells in vitro
and in vivo. Oncol Rep. 29:2197–2204. 2013.PubMed/NCBI
|
|
2
|
Kim SY, Kang S, Song JJ and Kim JH: The
effectiveness of the oncolytic activity induced by Ad5/F35
adenoviral vector is dependent on the cumulative cellular
conditions of survival and autophagy. Int J Oncol. 42:1337–1348.
2013.PubMed/NCBI
|
|
3
|
Hu X, Cao Y, Meng Y and Hou M: A novel
modulation of structural and functional changes of mouse bone
marrow derived dendritic cells (BMDCs) by interleukin-2(IL-2). Hum
Vaccin Immunother. 11:516–521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
de Vries IJ, Lesterhuis WJ, Scharenborg
NM, Engelen LP, Ruiter DJ, Gerritsen MJ, Croockewit S, Britten CM,
Torensma R, Adema GJ, et al: Maturation of dendritic cells is a
prerequisite for inducing immune responses in advanced melanoma
patients. Clin Cancer Res. 9:5091–5100. 2003.PubMed/NCBI
|
|
5
|
Kim R, Emi M and Tanabe K: Functional
roles of immature dendritic cells in impaired immunity of solid
tumour and their targeted strategies for provoking tumour immunity.
Clin Exp Immunol. 146:189–196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Castiello L, Sabatino M, Jin P, Clayberger
C, Marincola FM, Krensky AM and Stroncek DF: Monocyte-derived DC
maturation strategies and related pathways: A transcriptional view.
Cancer Immunol Immunother. 60:457–466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mossoba ME and Medin JA: Cancer
immunotherapy using virally transduced dendritic cells: Animal
studies and human clinical trials. Expert Rev Vaccines. 5:717–732.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cavallini C, Lovato O, Bertolaso A,
Pacelli L, Zoratti E, Zanolin E, Krampera M, Zamò A, Tecchio C,
Cassatella MA, et al: The TNF-family cytokine TL1A inhibits
proliferation of human activated B cells. PLoS One. 8:e601362013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Erdmann M and Schuler-Thurner B: Towards a
standardized protocol for the generation of monocyte-derived
dendritic cell vaccines. Methods Mol Biol. 595:149–163. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo Z, Liu H, He XP, Tan XH, Zhou Y, Chen
X, Shi YJ, Liu XD and Chen HR: A clinical study of cytokine-induced
killer cells for the treatment of refractory lymphoma. Oncol Lett.
2:531–536. 2011.PubMed/NCBI
|
|
11
|
Boudreau JE, Bonehill A, Thielemans K and
Wan Y: Engineering dendritic cells to enhance cancer immunotherapy.
Mol Ther. 19:841–853. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Morelli AE, Zahorchak AF, Larregina AT,
Colvin BL, Logar AJ, Takayama T, Falo LD and Thomson AW: Cytokine
production by mouse myeloid dendritic cells in relation to
differentiation and terminal maturation induced by
lipopolysaccharide or CD40 ligation. Blood. 98:1512–1523. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kelleher M and Beverley PC:
Lipopolysaccharide modulation of dendritic cells is insufficient to
mature dendritic cells to generate CTLs from naive polyclonal
CD8+ T cells in vitro, whereas CD40 ligation is
essential. J Immunol. 167:6247–6255. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tcherepanova IY, Adams MD, Feng X,
Hinohara A, Horvatinovich J, Calderhead D, Healey D and Nicolette
CA: Ectopic expression of a truncated CD40L protein from synthetic
post-transcriptionally capped RNA in dendritic cells induces high
levels of IL-12 secretion. BMC Mol Biol. 9:902008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hernandez MG, Shen L and Rock KL:
CD40-CD40 ligand interaction between dendritic cells and
CD8+ T cells is needed to stimulate maximal T cell
responses in the absence of CD4+ T cell help. J Immunol.
178:2844–2852. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thacker EE, Nakayama M, Smith BF, Bird RC,
Muminova Z, Strong TV, Timares L, Korokhov N, O'Neill AM, de Gruijl
TD, et al: A genetically engineered adenovirus vector targeted to
CD40 mediates transduction of canine dendritic cells and promotes
antigen-specific immune responses in vivo. Vaccine. 27:7116–7124.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kikuchi T, Moore MA and Crystal RG:
Dendritic cells modified to express CD40 ligand elicit therapeutic
immunity against preexisting murine tumors. Blood. 96:91–99.
2000.PubMed/NCBI
|
|
18
|
Tada Y, O-Wang J, Yu L, Shimozato O, Wang
YQ, Takiguchi Y, Tatsumi K, Kuriyama T, Takenaga K, Sakiyama S, et
al: T-cell-dependent antitumor effects produced by CD40 ligand
expressed on mouse lung carcinoma cells are linked with the
maturation of dendritic cells and secretion of a variety of
cytokines. Cancer Gene Ther. 10:451–456. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wolf B, Schwarzer A, Côté AL, Hampton TH,
Schwaab T, Huarte E, Tomlinson CR, Gui J, Fisher JL, Fadul CE, et
al: Gene expression profile of peripheral blood lymphocytes from
renal cell carcinoma patients treated with IL-2, interferon-α and
dendritic cell vaccine. PLoS One. 7:e502212012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stax AM, Crul C, Kamerling SW, Schlagwein
N, van der Geest RN, Woltman AM and van Kooten C: CD40L stimulation
of rat dendritic cells specifically favors the IL-12/IL-10 ratio
resulting in a strong T cell stimulatory capacity. Mol Immunol.
45:2641–2650. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagy ZS, Ross JA, Rodriguez G, Balint BL,
Szeles L, Nagy L and Kirken RA: Genome wide mapping reveals PDE4B
as an IL-2 induced STAT5 target gene in activated human PBMCs and
lymphoid cancer cells. PLoS One. 8:e573262013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gonzalez-Carmona MA, Lukacs-Kornek V,
Timmerman A, Shabani S, Kornek M, Vogt A, Yildiz Y, Sievers E,
Schmidt-Wolf IG, Caselmann WH, et al: CD40ligand-expressing
dendritic cells induce regression of hepatocellular carcinoma by
activating innate and acquired immunity in vivo. Hepatology.
48:157–168. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
DeBenedette MA, Calderhead DM,
Tcherepanova IY, Nicolette CA and Healey DG: Potency of mature
CD40L RNA electroporated dendritic cells correlates with IL-12
secretion by tracking multifunctional CD8(+)/CD28(+) cytotoxic
T-cell responses in vitro. J Immunother. 34:45–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rousseau RF, Biagi E, Dutour A, Yvon ES,
Brown MP, Lin T, Mei Z, Grilley B, Popek E, Heslop HE, et al:
Immunotherapy of high-risk acute leukemia with a recipient
(autologous) vaccine expressing transgenic human CD40L and IL-2
after chemotherapy and allogeneic stem cell transplantation. Blood.
107:1332–1341. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Balamotis MA, Huang K and Mitani K:
Efficient delivery and stable gene expression in a hematopoietic
cell line using a chimeric serotype 35 fiber pseudotyped
helper-dependent adenoviral vector. Virology. 324:229–237. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gaggar A, Shayakhmetov DM and Lieber A:
CD46 is a cellular receptor for group B adenoviruses. Nat Med.
9:1408–1412. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ophorst OJ, Kostense S, Goudsmit J, De
Swart RL, Verhaagh S, Zakhartchouk A, Van Meijer M, Sprangers M,
Van Amerongen G, Yüksel S, et al: An adenoviral type 5 vector
carrying a type 35 fiber as a vaccine vehicle: DC targeting, cross
neutralization, and immunogenicity. Vaccine. 22:3035–3044. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vitoriano-Souza J, Moreira N,
Teixeira-Carvalho A, Carneiro CM, Siqueira FA, Vieira PM,
Giunchetti RC, Moura SA, Fujiwara RT, Melo MN, et al: Cell
recruitment and cytokines in skin mice sensitized with the vaccine
adjuvants: Saponin, incomplete Freund's adjuvant, and
monophosphoryl lipid A. PLoS One. 7:e407452012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang S, Cohen CJ, Peng PD, Zhao Y, Cassard
L, Yu Z, Zheng Z, Jones S, Restifo NP, Rosenberg SA, et al:
Development of optimal bicistronic lentiviral vectors facilitates
high-level TCR gene expression and robust tumor cell recognition.
Gene Ther. 15:1411–1423. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ngoi SM, Chien AC and Lee CG: Exploiting
internal ribosome entry sites in gene therapy vector design. Curr
Gene Ther. 4:15–31. 2004. View Article : Google Scholar : PubMed/NCBI
|