Introduction
Microglial cells, the residential macrophages of the
brain, have a pivotal role in the central nervous system's (CNS)
innate immune response and act as the first line of defense against
microorganism invasion and injury (1).
Lipopolysaccharide (LPS) activates microglia to secrete various
pro-inflammatory and cytotoxic factors, including tumor necrosis
factor (TNF)-α, interleukin (IL)-1β and heat shock protein 60
(HSP60) (2,3). These factors are responsible for several
neurodegenerative diseases, including Parkinson's and Alzheimer's
disease as well as amyotrophic lateral sclerosis (4–6). Thus,
controlling microglia activation may represent an effective
therapeutic strategy against neurodegenerative diseases.
Oxymatrine (OMT) is a traditional Chinese medicine
derived from the root of Sophora flavescens. Previous
studies have revealed that OMT has numerous pharmacological
effects, including anti-inflammatory, antitumor and antioxidant
activities (7,8). Several studies have also indicated a
neuroprotective role of OMT. OMT has been reported to protect
neurons by downregulating 12/15-lipooxygenase, phospho-p38
mitogen-activated protein kinase and cytosolic phospholipase A2
(9). However, the neuroprotective
effects of OMT have remained to be fully elucidated.
HSP60 has pro-survival as well as pro-apoptotic
functions. Under stress conditions, microglial cells highly express
HSP60 and release it into the extracellular space, causing cell
death through binding Toll-like receptor (TLR)-4 (10). The present study aimed to assess the
neuroprotective effects of OMT in LPS-induced BV2 microglial cells
as well as to identify the possible mechanistic involvement of
HSP60. It was revealed that OMT likely suppressed microglial
activation via the HSP60/TLR-4/myeloid differentiation factor
(MYD)88/nuclear factor (NF)-κB signaling pathway.
Materials and methods
Chemicals
The BV2 murine microglial cell line was purchased
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China). LPS was purchased from Sigma-Aldrich (St. Louis, MO, USA).
OMT was from Guanyu Bio-tech (Xi'an, China). Antibodies against
caspase-3 (cat. no. 9664; 1:2,000 dilution), MYD88 (cat. no. 4283;
1:1,000 dilution) and TLR-4 (cat. no. 2219; 1:1,000 dilution) were
from Cell Signaling Technology, Inc. (Beverly, MA, USA). Anti-NF-κB
(cat. no. ab131485; 1:200 dilution) and anti-inducible nitric oxide
synthase (iNOS) antibodies (cat. no. ab3523; 1:200 dilution) were
from Abcam (Cambridge, MA, USA). Antibodies against HSP60
(ADI-SPA-806-D, 1:1,000) and heat shock factor (HSF)-1 (cat. no.
ADI-SPA-950-D; 1:1,000 dilution) were purchased from Stressgen (San
Diego, CA, USA). Anti-β-actin antibody (cat. no. ZM-0001; 1:1,000
dilution) was purchased from Zhongshan Goldenbridge Bio. IL-6,
IL-1β and TNF-α ELISA kits were acquired from Xinbosheng
Biotechnology Inc. (Shenzhen, China). Enhanced chemiluminescence
(ECL) and bicinchoninic acid (BCA) kits were from Pierce (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Dulbecco's modified
Eagle's medium (DMEM) and fetal bovine serum (FBS) were from Gibco
(Thermo Fisher Scientific, Inc). The Cell Counting Kit-8 (CCK-8)
and the Annexin V-fluorescein isothiocyanate (FITC)/propidium
iodide (PI) apoptosis detection kit were obtained from Beibo
(Shanghai, China).
Cell culture
BV2 cells were maintained in DMEM with 10% FBS and
grown at 37°C in a humidified atmosphere containing 5%
CO2. OMT was dissolved in DMEM. Cells were pre-treated
with LPS (1 µg/ml) for 30 min, followed by addition of different
concentrations of OMT. After 24 h, the supernatant of the cells was
obtained for ELISA, and cells were collected, a proportion of which
were lysed using a total protein extraction kit (KeyGen Biotech,
Jiangsu, China) for subsequent experiments.
Cell viability assay
Cell viability was evaluated using the CCK-8 assay.
Cells were seeded into 96-well microtiter plates at
5×104 cells/well and pre-treated with LPS (1 µg/ml) for
30 min, followed by treatment with various concentrations of OMT
(0, 1, 10, 20, 50 or 100 µg/ml). After 24 h of incubation, CCK-8
solution was added to each well, followed by incubation for 2 h.
The color intensity of the metabolized CCK-8 stain was determined
by measuring the absorbance at 450 nm with an immunoreader (PR4100;
Bio-Rad Laboratories, Inc., Hercules, CA, USA). Cell viability was
expressed as a percentage of the control group (untreated). Every
experiment was repeated three times.
ELISA
BV2 cells were pre-treated with LPS (1 µg/ml) for 30
min, followed by treatment with 0 or 20 µg/ml OMT for 24 h and
subsequent collection of the culture medium. In a control group,
the cells were left untreated. The levels of TNF-α, IL-6, IL-1β and
HSP60 in the culture medium were determined by ELISA kits according
to the manufacturer's instructions. The absorbance was measured at
450 nm on a microplate reader (Bio-Rad Laboratories, Inc.).
Western blot analysis
To determine the expression of certain proteins, BV2
cells were pre-treated with LPS (1 µg/ml) for 30 min, followed by
treatment with 0 or 20 µg/ml OMT for 24 h, and protein was
extracted using a lysis buffer containing 0.1% Triton X-100, 2%
sodium dodecyl sulfate (SDS), 5% glycerol, 5 mM
ethylenediaminetetraacetate, 150 mM NaCl and 20 mM Tris (pH 7.5)
(Bioscience, Shanghai, China). Following incubation at 4°C for 30
min, the lysates were centrifuged at 8,050 × g at 4°C for 30 min.
The BCA kit was then used to determine the protein concentration.
Protein denaturation was then performed in a sample buffer
containing 2-mercaptoethanol and bromophenol blue (KeyGen Biotech)
for 10 min at 95°C. Equal amounts of protein were resolved by 10%
SDS-polyacrylamide gel electrophoresis and transferred onto a
polyvinylidene difluoride membrane (EM Millipore, Billerica, MA,
USA). The membranes were blocked with 5% milk in Tris-buffered
saline containing Tween-20 and incubated at 4°C overnight with the
indicated antibodies, including anti-iNOS, -TLR-4, -heat shock
factor 1 (HSF-1), -NF-кB, -HSP60, -caspase-3, -MYD88 and -β-actin.
After washing with phosphate-buffered saline (PBS), the membranes
were incubated with anti-mouse (cat. no. ZB-2305; 1:5,000) or
anti-rabbit (cat. no. ZB-2301; 1:5,000) secondary antibodies
(Zhongshan Goldenbridge Bio) at room temperature for 75 min. The
proteins were then visualized using the ECL kit and membranes were
exposed to X-ray film. The blotting results were semi-quantified
using Quantity One software, version 4.6.9 (Bio Rad Laboratories,
Inc.).
Flow cytometry
BV2 cells were divided into three groups: Control
group, LPS group [treated with LPS (1 µg/ml) for 24 h] and the
LPS+OMT group [treated with LPS (1 µg/ml) for 30 min and
subsequently with OMT (20 µg/ml)]. The cells were then collected
and washed twice with cold PBS. Following centrifugation at 126 × g
for 4 min at room temperature, cells were re-suspended in 400 µl
binding buffer. After incubation with 5 µl Annexin V-FITC for 15
min, 10 µl PI was added, followed by incubation for 5 min in the
dark. The apoptotic rate was measured by a FACSCalibur flow
cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. One-way analysis of variance, followed by the
Student-Newman-Keuls post hoc test were used for comparisons
between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
OMT inhibits LPS-induced decreases of
microglial cell viability
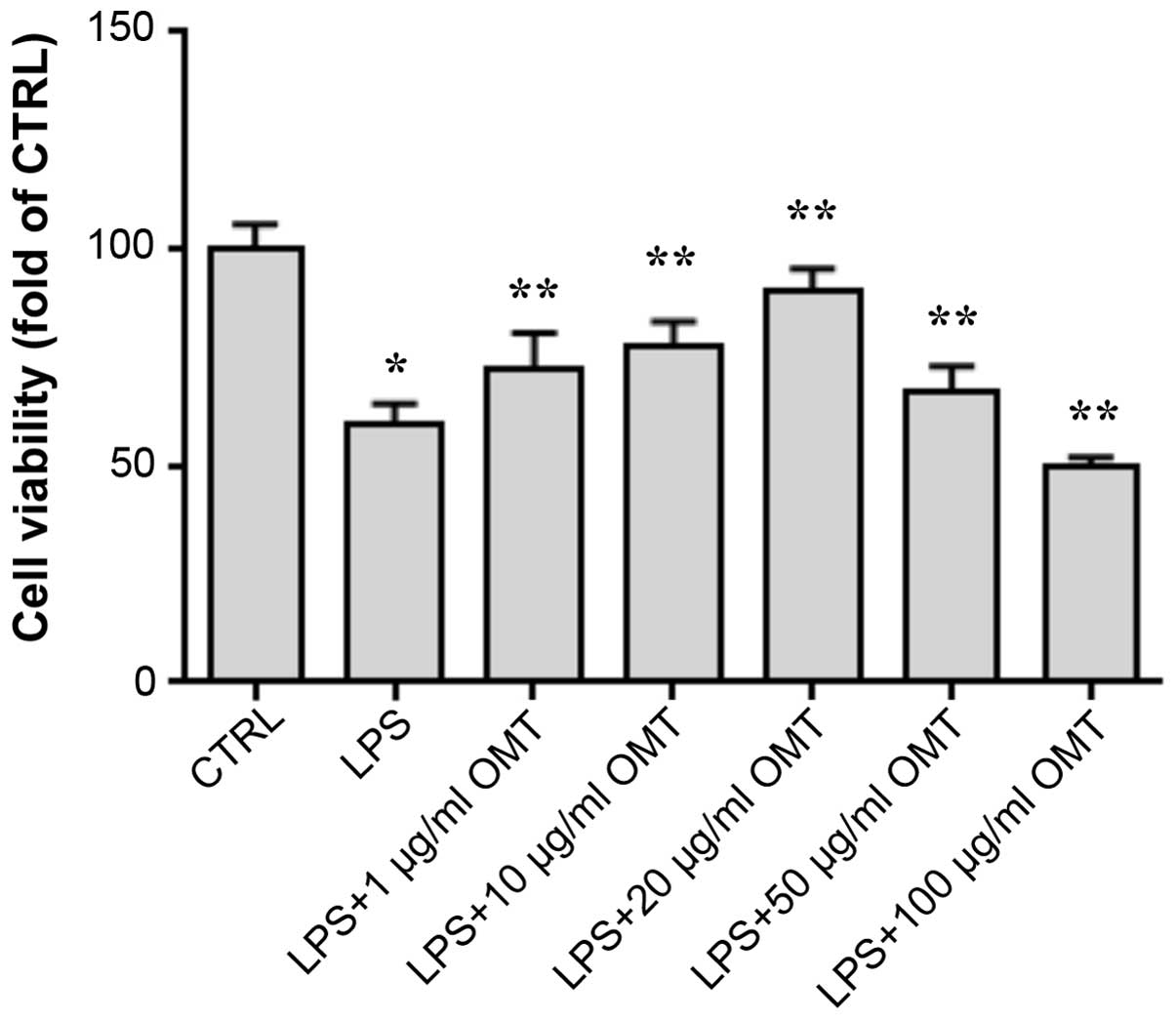
To evaluate the effects of OMT on LPS-induced
toxicity on BV2 cells, the CCK-8 assay was performed (Fig. 1). After LPS stimulation, the viability
of the cells was obviously decreased. Of note, at concentrations of
1–20 µg/ml, OMT increased the cell viability in a dose-dependent
manner, indicating that it had a protective effect on BV2 cells
against LPS-induced toxicity. However, compared to that in the LPS
+ 20 µg/ml OMT group, higher concentrations of OMT (50 and 100
µg/ml) decreased the cell viability; therefore, the OMT
concentration of 20 µg/ml with the maximum protective effect was
used in all subsequent experiments.
OMT has anti-apoptotic effects on
microglia
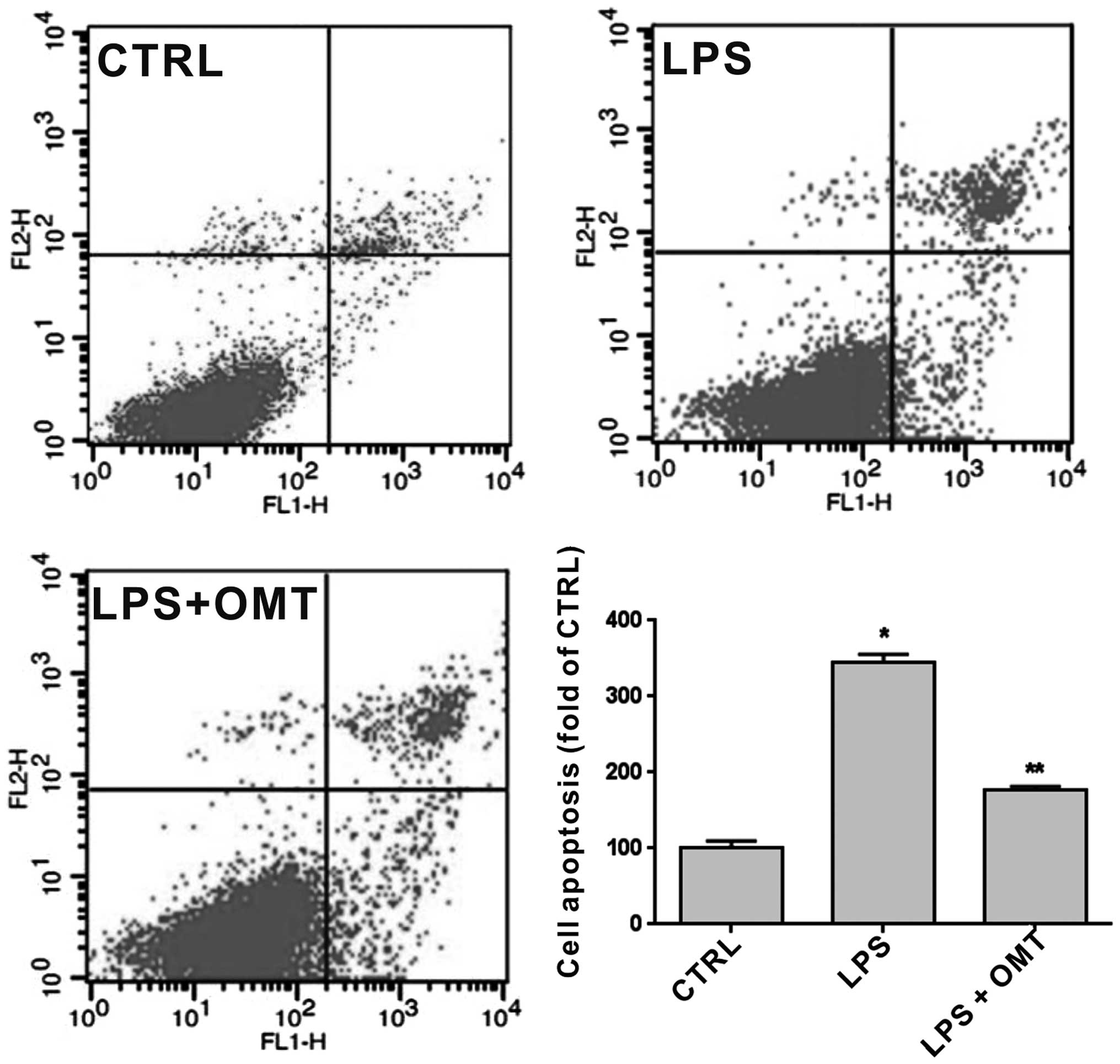
To further confirm the protective effect of OMT on
BV2 cells, Annexin V and PI double staining followed by flow
cytometric analysis were performed on cells pre-treated with LPS (1
µg/ml) for 30 min and incubated with 20 µg/ml OMT for 24 h
(Fig. 2). The results showed that OMT
significantly reduced LPS-induced apoptosis of BV2 cells.
Therefore, OMT exerted a significant anti-apoptotic effect on
microglia (P<0.05).
OMT inhibits LPS-induced expression
and release of HSP60 by BV2 cells
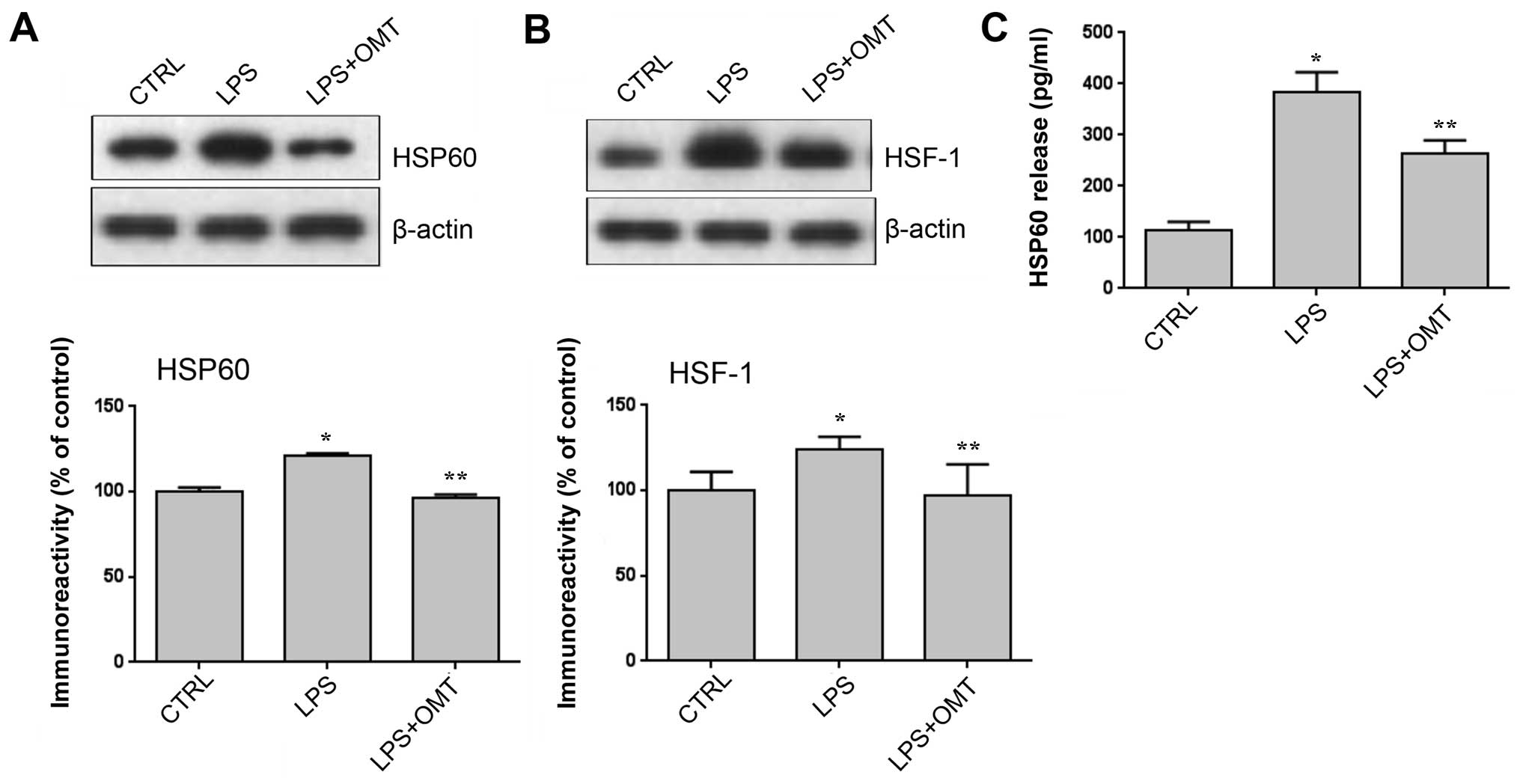
To further explore the mechanisms of the protective
effects of OMT, HSP60 expression and release by BV2 cells were
assessed. Numerous studies have indicated that HSP60 has a role in
microglia-induced inflammation, which was confirmed by a study by
our group (3). Therefore, it was
hypothesized that OMT exerts its anti-inflammatory effects via
HSP60. Western blot analysis showed that the expression of HSP60
was significantly increased after LPS stimulation, which was
completely inhibited by OMT (Fig. 3A).
HSF-1 is a transcription factor regulating HSP60 expression and was
therefore examined as well. The results showed that OMT completely
abrogated the LPS-induced increases in HSF-1 expression (Fig. 3B). Furthermore, an ELISA revealed that
OMT significantly decreased the LPS-induced release of
extracellular HSP60 (Fig. 3C). These
results indicated that under the stress conditions, HSP60 is highly
expressed and secreted into the extracellular space to induce cell
apoptosis, which is consistent with the hypothesis of the present
study.
OMT inhibits LPS-induced upregulation
of iNOS, TLR-4, NF-κB p65, caspase-3 and MYD88 expression
TLR-4 is a receptor of LPS and HSP60 on the surface
of microglia, which can activate downstream signaling molecules,
including NF-κB, caspase-3, MYD88 and iNOS. To determine whether
extracellular HSP60 released by BV2 cells can bind TLR-4 and
activate the TLR-4 signaling pathway and whether OMT inhibits this
activation, the effects of LPS and OMT on the protein levels of
TLR-4, Myd88, NF-κB, caspase-3 and iNOS were assessed by western
blotting. The results showed that OMT abrogated the LPS-induced
expression of TLR-4, NF-κB, caspase-3, MYD88 and iNOS in BV2 cells
(Fig. 4). These experiments indicated
that OMT may exert its anti-inflammatory effects via inhibiting the
TLR-4 pathway.
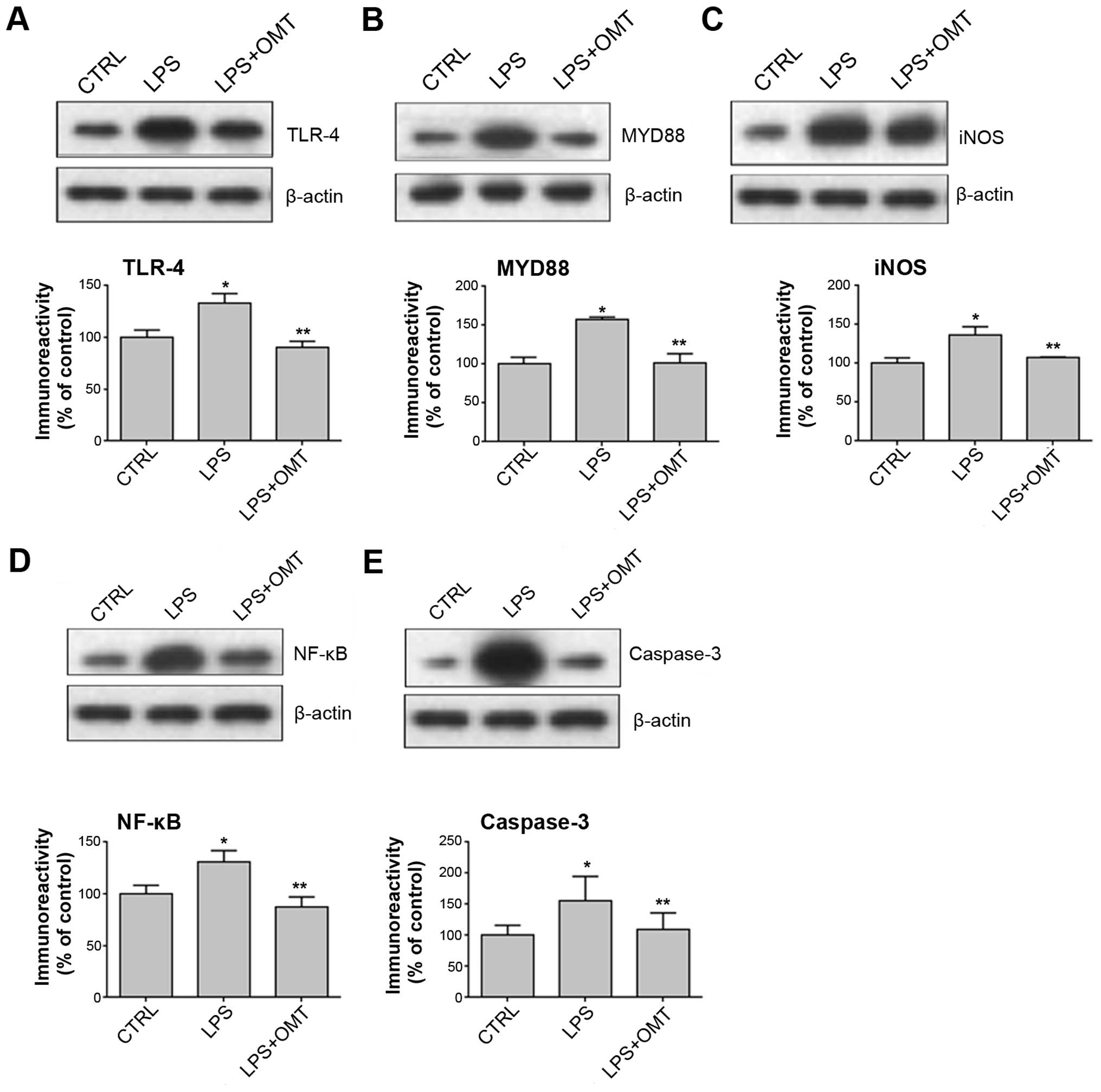 | Figure 4.OMT reduces the expression of (A)
TLR-4, (B) MYD88, (C) iNOS, (D) NF-κB and (E) caspase-3 in
LPS-stimulated BV2 microglia. BV2 cells were pre-treated with LPS
for 30 min, followed by incubation with 20 µg/ml OMT for 24 h.
β-actin was used as a reference. Values are expressed as the mean ±
standard error of three independent experiments. *P<0.05 vs.
CTRL; **P<0.05 vs. LPS group. CTRL, control; LPS,
lipopolysaccharide; OMT, oxymatrine; TLR, Toll-like receptor;
MYD88, myeloid differentiation factor 88; iNOS, inducible nitric
oxide synthase; NF-κB, nuclear factor-κB. |
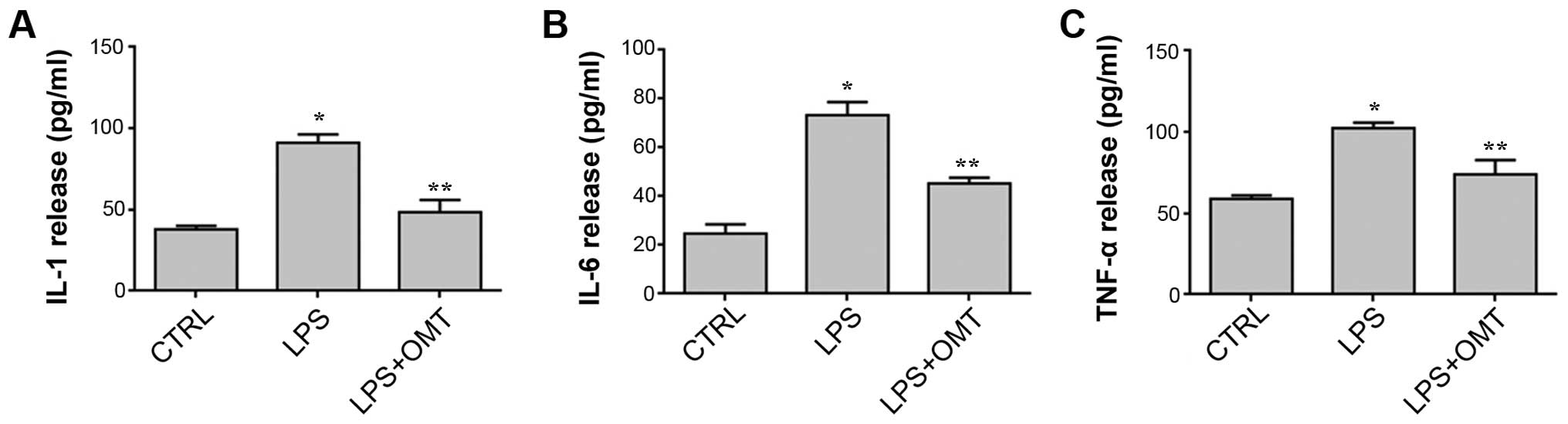
OMT inhibits LPS-induced inflammatory
cytokine release by BV2 microglial cells
Activation of the TLR-4 signaling pathway is
accompanied by the production and release of inflammatory
cytokines. To further investigate the effects of OMT on
LPS-stimulated cytokines, the levels of TNF-α, IL-1β and IL-6 in
the culture medium were measured by ELISA. As shown in Fig. 5, OMT markedly attenuated LPS-induced
release of TNF-α, IL-1β and IL-6 by BV2 microglial cells. These
results indicated that OMT exerts its anti-inflammatory effects by
suppressing the release of inflammatory factors.
Discussion
OMT is the major alkaloid of Sophora
flavescens and has anti-inflammatory, anti-allergic,
anti-viral, anti-fibrotic and cardiovascular protective properties
(11–13). A previous study has reported that in
rats with cerebral ischemia, OMT reduced the production of NF-κB
and suppressed the inflammatory reaction of the CNS to decrease the
area of cerebral infarction (14).
However, the neuroprotective effects of OMT and the underlying
molecular mechanisms have yet to be fully elucidated. The
inhibition of microglia activation has been proposed to be an
effective therapeutic option for neurodegenerative diseases.
Therefore, the present study investigated whether OMT treatment can
inhibit microglia activation and examined the underlying
mechanisms. It was revealed that OMT attenuated microglia
activation by suppressing the HSP60/TLR-4/MYD88/NF-κB signaling
pathway.
The mitochondrial matrix protein HSP60 can be
induced by stress and forms a chaperon complex within the
mitochondria, which is important for mitochondrial protein folding
and function (15). Activated HSP60
mainly locates in the plasma membrane or the extracellular space
and can mediate cell apoptosis. Furthermore, HSP60 has also been
reported to enhance caspase activation to exert a pro-apoptotic
function (16). In the present study,
the effects of OMT on microglia apoptosis were assessed by Annexin
V and PI double staining. The results indicated that LPS induced
apoptosis in BV2 microglial cells, which was inhibited by OMT.
Further mechanistic study revealed that OMT decreased the
expression and release of HSP60 triggered by LPS. It has been
reported that HSP60 is a ligand of TLR-4, indicating that HSP60
evokes an immune response (17).
TLR-4 signaling may proceed via two possible
pathways-the MYD88-dependent and the MYD88-independent pathway.
MYD88 is the universal intracellular adaptor recruited by all known
TLRs except TLR-3 (18). The results
of the present study showed that the expression of MYD88 was
increased in LPS-induced BV2 cells, which was abrogated by OMT,
indicating that HSP60 activates microglia via the MYD88-dependent
pathway. NF-κB is a vital factor in the TLR-4 pathway and
translocates to the nucleus upon its activation, where it induces
the production of several pro-inflammatory cytokines, including
TNF-α, IL-1β and IL-6 (19). NF-κB is
also the principal regulator of the transactivation of
pro-inflammatory genes such as iNOS (20). Furthermore, NF-κB also initiates the
transcription of the HSP60 gene, which leads to a potent
inflammatory response in innate immune cells (21). The results of the present study showed
that OMT suppressed the expression of NF-κB and iNOS in LPS-induced
BV2 cells and decreased the release of inflammatory factors. The
activation of NF-κB by caspase-3 is critical in inflammation and
cell apoptosis (22). LPS-stimulated
microglia have been shown not to be toxic to neighboring neurons
when caspase-3 is inhibited (23). The
results of the present study showed that OMT reduced the levels of
cleaved caspase-3, which is consistent with the fact that OMT
inhibited apoptosis, as indicated by flow cytometry.
In conclusion, the results of the present study
demonstrated that OMT exerts neuroprotective and anti-inflammatory
effects on LPS-induced microglial cells, which may be attributed to
the inhibition of the HSP60-TLR-4-MYD88-NF-κB pathway. Therefore,
these findings provide a potential therapeutic application of OMT
for neurodegenerative diseases.
Acknowledgements
The present study was supported by grants from
National Natural Science Foundation of China (nos. 81460182,
31460257, 81571098, 31560273, 81260051 and 31260243) and the
Ningxia Natural Science Foundation (no. NZ14057). Additional
funding was provided for Dr Yin Wang by the Program for New Century
Excellent Talents in University.
References
|
1
|
Kreutzberg GW: Microglia: A sensor for
pathological events in the CNS. Trends Neurosci. 19:312–318. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jung WK, Lee DY, Park C, Choi YH, Choi I,
Park SG, Seo SK, Lee SW, Yea SS, Ahn SC, et al: Cilostazol is
anti-inflammatory in BV2 microglial cells by inactivating nuclear
factor-kappaB and inhibiting mitogen-activated protein kinases. Br
J Pharmacol. 159:1274–1285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li YH, Teng P, Wang Y, Zhang YM, Ma CJ and
Pu J: Expression and regulation of HSP60 in activated microglia
cells. J Ningxia Med Col. 8:712–714. 2011.
|
|
4
|
Mhatre M, Floyd RA and Hensley K:
Oxidative stress and neuroinflammation in Alzheimer's disease and
amyotrophic lateral sclerosis: Common links and potential
therapeutic targets. J Alzheimers Dis. 6:147–157. 2004.PubMed/NCBI
|
|
5
|
Tansey MG, McCoy MK and Frank-Cannon TC:
Neuroinflammatory mechanisms in Parkinson's disease: Potential
environmental triggers, pathways, and targets for early therapeutic
intervention. Exp Neurol. 208:1–25. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McGeer PL and McGeer EG: Inflammatory
processes in amyotrophic lateral sclerosis. Muscle Nerve.
26:459–470. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan X, Wang Y, Du D, Hu Z, Xu M, Xu M and
Liu Z: The effects of the combination of sodium ferulate and
oxymatrine on lipopolysaccharide-induced acute lung injury in mice.
Inflammation. 35:1161–1168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Y, Xu Y, Ji W, Li X, Sun B, Gao Q and
Su C: Anti-tumor activities of matrine and oxymatrine: Literature
review. Tumour Biol. 35:5111–5119. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cui L, Zhang X, Yang R, Wang L, Liu L, Li
M and Du W: Neuroprotection and underlying mechanisms of oxymatrine
in cerebral ischemia of rats. Neurol Res. 33:319–324. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chandra D, Choy G and Tang DG: Cytosolic
accumulation of HSP60 during apoptosis with or without apparent
mitochondrial release: evidence that its pro-apoptotic or
pro-survival functions involve differential interactions with
caspase-3. J Biol Chem. 282:31289–31301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang M, Hu YY, Dong XQ, Xu QP, Yu WH and
Zhang ZY: The protective role of oxymatrine on neuronal cell
apoptosis in the hemorrhagic rat brain. J Ethnopharmacol.
143:228–235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chai NL, Fu Q, Shi H, Cai CH, Wan J, Xu SP
and Wu BY: Oxymatrine liposome attenuates hepatic fibrosis via
targeting hepatic stellate cells. World J Gastroenterol.
18:4199–4206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li S Hong, Lei L, Lei S, Dan Z, De Li D,
Guo Fen Q, Yan L, Wen Feng C and Bao Feng Y: Cardioprotective
effects and underlying mechanisms of oxymatrine against ischemic
myocardial injuries of rats. Phytother Res. 22:985–989. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Y, Zhang XJ, Yang CH and Fan HG:
Oxymatrine protects rat brains against permanent focal ischemia and
downregulates NF-kappaB expression. Brain Res. 1268:174–180. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Voos W and Röttgers K: Molecular
chaperones as essential mediators of mitochondrial biogenesis.
Biochim Biophys Acta. 1592:51–62. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Samali A, Cai J, Zhivotovsky B, Jones DP
and Orrenius S: Presence of a pre-apoptotic complex of
pro-caspase-3, Hsp60 and Hsp10 in the mitochondrial fraction of
jurkat cells. EMBO J. 18:2040–2048. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lehnardt S, Schott E, Trimbuch T, Laubisch
D, Krueger C, Wulczyn G, Nitsch R and Weber JR: A vicious cycle
involving release of heat shock protein 60 from injured cells and
activation of toll-like receptor 4 mediates neurodegeneration in
the CNS. J Neurosci. 28:2320–2331. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rosenberger K, Dembny P, Derkow K, Engel
O, Krüger C, Wolf SA, Kettenmann H, Schott E, Meisel A and Lehnardt
S: Intrathecal heat shock protein 60 mediates neurodegeneration and
demyelination in the CNS through a TLR4-and MyD88-dependent
pathway. Mol Neurodegener. 10:52015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsung A, McCoy SL, Klune JR, Geller DA,
Billiar TR and Hefeneider SH: A novel inhibitory peptide of
Toll-like receptor signaling limits lipopolysaccharide-induced
production of inflammatory mediators and enhances survival in mice.
Shock. 27:364–369. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dasgupta S, Jana M, Zhou Y, Fung YK, Ghosh
S and Pahan K: Antineuroinflammatory effect of NF-kappaB essential
modifier-binding domain peptides in the adoptive transfer model of
experimental allergic encephalomyelitis. J Immunol. 173:1344–1354.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Chen L, Hagiwara N and Knowlton
AA: Regulation of heat shock protein 60 and 72 expression in the
failing heart. J Mol Cell Cardiol. 48:360–366. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Soria JA, Arroyo DS, Gaviglio EA,
Rodriguez-Galan MC, Wang JM and Iribarren P: Interleukin 4 induces
the apoptosis of mouse microglial cells by a caspase dependent
mechanism. Neurobiol Dis. 43:616–624. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Burguillos MA, Deierborg T, Kavanagh E,
Persson A, Hajji N, Garcia-Quintanilla A, Cano J, Brundin P,
Englund E, Venero JL and Joseph B: Caspase signalling controls
microglia activation and neurotoxicity. Nature. 472:319–324. 2011.
View Article : Google Scholar : PubMed/NCBI
|