Introduction
Ultraviolet B (UVB) irradiation induces acute and
chronic skin disorders, characterized by inflammation such as
erythema and edema, hyperplasia, dyspigmentation and deep wrinkle
formation (termed photoaging) (1–3). These
clinical symptoms have been reported to contribute to the
functional and structural abnormalities of the epidermis and
dermis, including an increase of epidermal thickness, a decrease of
epidermal polarity and the augmented degradation of extracellular
matrices such as collagen and elastin (4–6). On the
other hand, sebum secretion is considered to be involved in the
functional maintenance of the cutaneous surface as a biologic
barrier (7). Excessive sebum
production and secretion in sebaceous glands has been reported to
be associated with the development and aggravation of acne vulgaris
and seborrheic dermatitis (8). In
addition, UVB-irradiation has been reported to increase sebum
secretion and production in humans and hamsters in vivo
(9–11). Furthermore, UVB exposure has been
reported to facilitate free radical-mediated lipid peroxidation of
sebum in the skin (10,12). Therefore, sunscreen or topical and
systemic administration of antioxidants is likely to be an
efficient strategy for preventing the UV-mediated perturbation of
cutaneous barrier functions (13).
Extracts of the Chinese herb, Tripterygium
wilfordii Hook F (TWHF), have been reported to be effective in
the treatment of autoimmune diseases such as rheumatoid arthritis,
nephritis and lupus erythematosus (14–16).
Triptolide, a diterpenoid triepoxide, has been reported to be a
component of TWHF that possesses immunosuppressive,
anti-inflammatory and anti-oxidative actions in vivo and
in vitro (17–20). A previous study by the current
authors reported that triptolide inhibits the production of
prostaglandin E2 due to the transcriptional suppression of
prostaglandin H synthase (cyclooxygenase) 2 in human synovial
fibroblasts and mouse macrophages (18). In addition, prostaglandin E2 has been
reported to be a prominent indicator of UVB-mediated inflammation
in the skin, which contributes to the regulation of keratinocyte
proliferation and differentiation (21). Therefore, triptolide may be a
candidate for a novel therapeutic agent that improves skin barrier
functions under UVB-mediated acute and chronic inflammatory
conditions, including acne. However, it is not yet fully understood
whether triptolide can control the UVB-mediated dysregulation of
sebum production in sebaceous glands.
In the present study, the effects of triptolide on
UVB-augmented sebum production in sebaceous glands in hamsters were
examined both in vivo and in vitro. The results
indicated that triptolide inhibits UVB-enhanced sebum production in
sebaceous glands.
Materials and methods
Preparation of differentiated hamster
sebocytes
Hamster sebocytes were established from the
sebaceous glands of auricles of five-week-old male golden hamsters
(Japan SLC, Inc., Hamamatsu, Japan), as described previously
(22). Hamster sebocytes
(2.4×104 cells/cm2) were plated onto 12-well plates or
35-mm diameter culture dishes (BD Biosciences, Franklin Lakes, NJ,
USA), then cultured for 24 h at 37°C in Dulbecco's modified Eagle's
medium/F12 (Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 6% heat-denatured fetal bovine serum
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 2% human serum
(ICN Biomedicals, Irvine, CA, USA), 0.68 mM L-glutamine
(Invitrogen; Thermo Fisher Scientific, Inc.) and 10 nM recombinant
human epidermal growth factor (Progen Biotechnik GmbH, Heidelberg,
Germany) to achieve complete cell adhesion, as previously described
(11). The hamster sebocytes were
treated every two days for up to 10 days with 10 nM insulin
(Sigma-Aldrich; Merck KGaA), during which time intracellular lipid
droplets were abundantly formed. The resulting cells were termed
differentiated hamster sebocytes (DHS) (11). In this series of experiments, hamster
sebocytes were used up to the third passage level.
In vivo and in vitro UVB irradiation
and triptolide treatment
The auricle skin of three-week-old male golden
hamsters (n=6; weight, 48.7±3.3 g; Japan SLC, Inc.) was topically
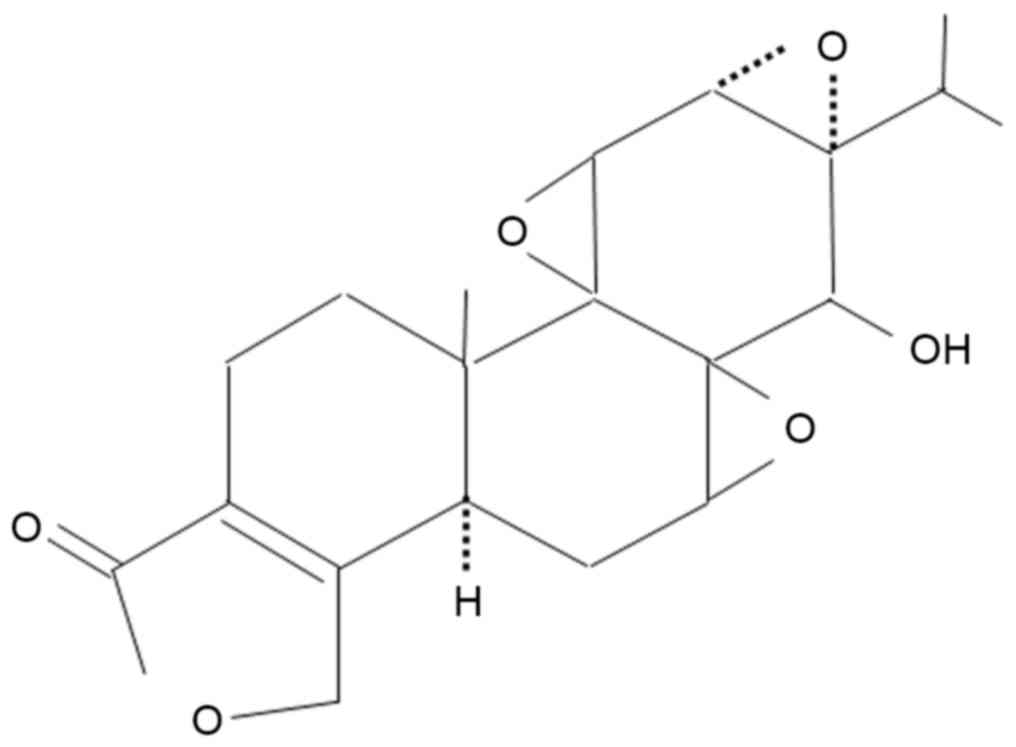
treated once a day with 50 µl of a 3.1 nmol solution of triptolide
(Enzo Life Sciences, Inc., Farmingdale, NY, USA; n=3; Fig. 1) in 95% ethanol and 5% glycerol, or
with the same volume of vehicle (n=3) before each UVB irradiation
(5.4 kJ/m2), once a day for seven days (11). For the in vitro UVB
irradiation experiments, DHS in 12-well plates were irradiated with
UVB (0.12–0.62 kJ/m2) (6) in a
serum-free culture medium supplemented with triptolide (28–112 nM),
then cultured for 24 h at 37°C. UVB irradiation was performed using
a Toshiba FL20S fluorescent sunlamp (Toshiba Corporation, Tokyo,
Japan), emitting rays between 275 and 375 nm with a peak emission
of 313 nm. The radiance was measured by a UV Indicator MI-340 (EKO
Instruments, Tokyo, Japan), as previously described (6). Hamsters were housed in a room
maintained at 23±1°C and 55±5% humidity with a 12-h light/dark
cycle (lights on 7:00 a.m. to 7:00 p.m.) during the experiments.
Food and water were available ad libitum. All experiments
presented in this study were performed according to the Guidelines
of Experimental Animal Care issued by the Prime Minister's Office
of Japan (Tokyo, Japan). The experimental protocol was approved by
the Committee of Animal Care and Use of Tokyo University of
Pharmacy and Life Sciences (Tokyo, Japan).
Oil red O staining
Animals were sacrificed by CO2 inhalation. Auricle
skin from hamsters irradiated with or without UVB as described
above was snap-frozen in liquid nitrogen. The frozen tissue
sections (8-µm) were incubated in 60% isopropanol after washing
with distilled H2O. Tissue sections were stained with 0.3% oil red
O (Sigma-Aldrich; Merck KGaA) in a solution of isopropanol and
distilled H2O (3:2, vol:vol) at 37°C for 15 min. Sections were then
viewed with a light microscope furnished with a digital camera
(Olympus Corporation, Tokyo, Japan). Sections were also
counterstained with Mayer's hematoxylin solution (Wako Pure
Chemical Industries, Ltd., Osaka, Japan), as previously described
(23).
Analysis of sebum components
Lipid components of sebum from the skin surface of
hamster auricles and in cultured hamster sebocytes were analyzed
using an automatic thin-layer chromatography (TLC) using a flame
ionization detector (Iatroscan; Iatron Laboratories, Inc., Tokyo,
Japan), as previously described (22). Briefly, the auricles were wiped with
acetone-impregnated cotton, then the sebum on the skin surface was
extracted twice for 30 sec with 50 µl of acetone in stainless steel
cups, 1 h after the wiping. The cells were harvested with 0.25%
trypsin and 0.02% EDTA in phosphate-buffered saline, then
sonicated. The cell lysate and harvested culture medium were mixed
with chloroform:methanol (2:1) for 5 min at room temperature. The
mixture was separated by centrifugation at 1,000 × g for 5 min at
room temperature following addition of 0.88% KCl. After removing
the methanol (the upper phase) with a pipette, the chloroform
fraction (the lower phase), including lipids, was collected. The
sebum extracts were subjected to Iatroscan and the levels of TG, a
major sebum component, and minor sebum components, free-fatty acids
(FFAs) and cholesterol (Ch), were calculated using an internal
control, authentic cholesterol acetate (2 g; Doosan Serdary
Research Laboratories, Toronto, Canada). Tripalmitin (for TG),
palmitic acid (for FFA) and Ch (Doosan Serdary Research
Laboratories) were used as lipid standards for chromatography.
Measurement of intracellular TG
After the treatment of DHS with UVB and/or
triptolide, the harvested cells were subjected to quantification of
TG using Liquitech TG-II (Roche Diagnostics K.K., Tokyo, Japan), as
previously described (11). The
quantity of intracellular TG was calculated using an authentic
trioleinate-standard solution (0.6 mg/ml).
Acyl coenzyme A (coA)/diacylglycerol
acyltransferase (DGAT) activity
Acyl CoA/DGAT activity in DHS treated with UVB
and/or triptolide was measured using 1,2-dioleoyl glycerol (Cayman
Chemical Co., Ann Arbor, MI, USA) and 14C-palmitoyl-CoA (GE
Healthcare Biosciences, Pittsburgh, PA, USA), as previously
described (24).
Statistical analysis
Data are presented as the mean ± standard deviation,
and were analyzed using one-way analysis of variance, and the
Fisher test for multiple comparisons. SPSS 17.0 software was used
to analyze the data (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of topical application of
triptolide on sebum levels on the skin surface
The effects of triptolide on sebum accumulation were
evaluated in sebaceous glands in three-week-old male golden
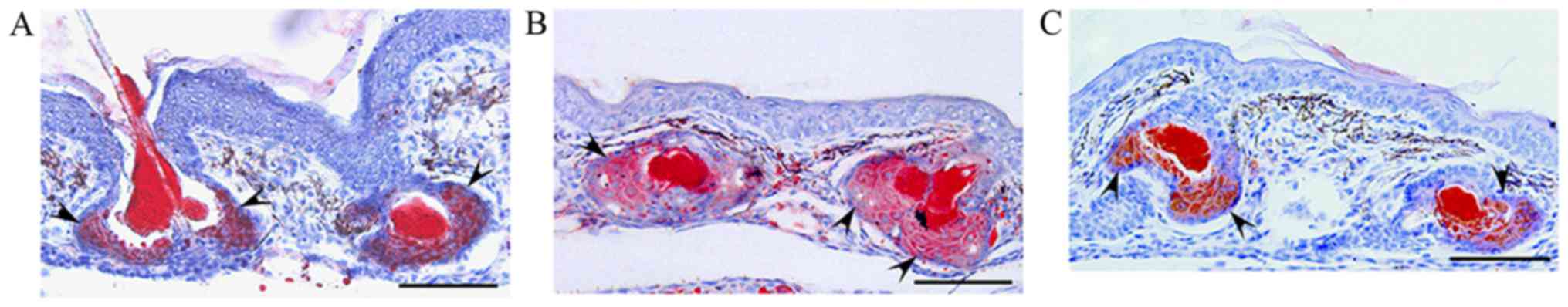
hamsters. Oil red O staining revealed that sebum accumulation in
the sebaceous glands and follicular ducts, as well as sebaceous
gland size, were increased in the UVB-irradiated hamsters compared
with the control (Fig. 2A and B). In
addition, when triptolide (3.1 nmol) was topically applied to the
skin of auricles before each UVB-irradiation, the UVB-enhanced
sebum accumulation and sebaceous size were decreased (Fig. 2B and C). Therefore, these results
suggested that topical application of triptolide reduced
UVB-induced aberrant sebum accumulation in sebaceous glands in
vivo.
Characterization of skin surface lipid
components in triptolide-treated hamsters
It was previously reported by the current authors
that sebum produced in hamster sebocytes consists of TG, the major
lipid component, and other minor components such as FFAs and Ch
(22). Therefore, in the current
study, alterations of sebum components on the skin surface were
evaluated in UVB and/or triptolide-treated hamsters. A TLC analysis
of sebum on the skin surface indicated that UVB irradiation
significantly augmented the levels of TG (2.3±1.2-fold, P<0.05),
FFAs (1.5±0.3-fold, P<0.05) and Ch (2.0±0.5-fold, P<0.05)
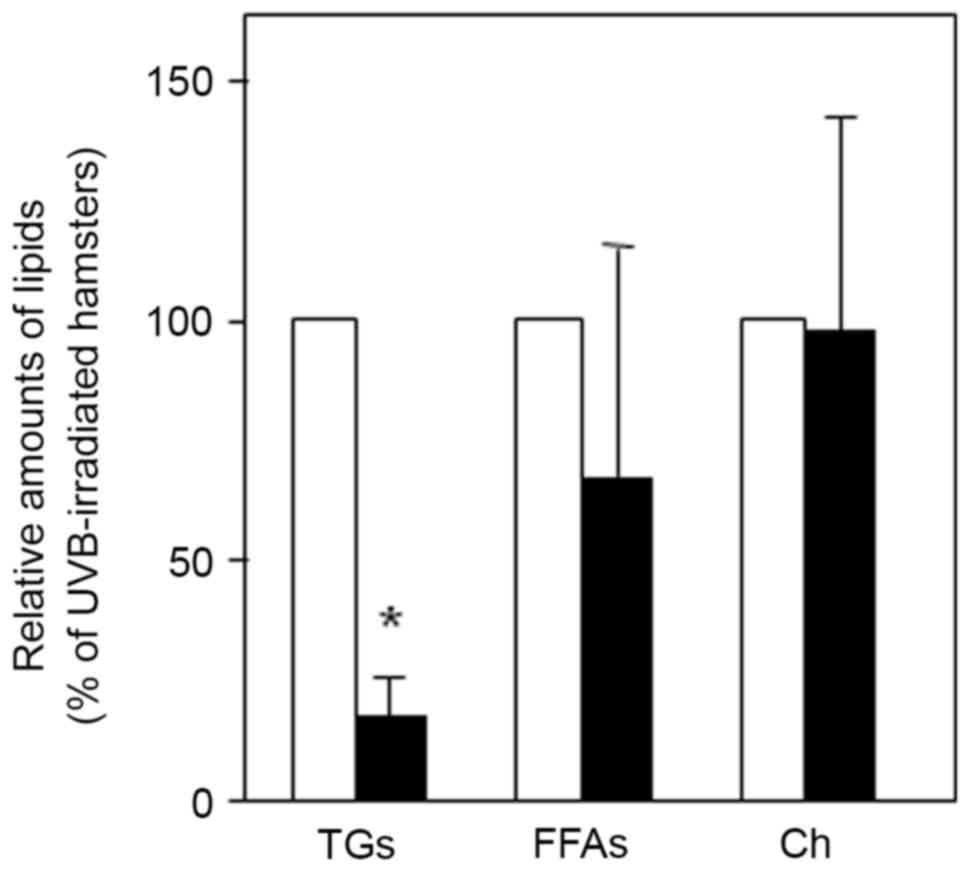
(Table I). In addition, topical
application of triptolide was found to significantly decrease the
level of TG (83% inhibition, P<0.05) compared with control
hamsters treated with a vehicle (Fig.
3). However, there was little change in the level of FFAs and
Ch on the skin surface of UVB-irradiated hamsters compared with
controls. These results suggested that triptolide selectively
modulated the level of TG rather than other sebum components in the
skin of UVB-irradiated hamsters.
 | Table I.Lipid composition on the skin surface
of auricles in UVB-irradiated hamsters. |
Table I.
Lipid composition on the skin surface
of auricles in UVB-irradiated hamsters.
|
| Relative amounts of
lipids (fold vs. control) |
|---|
|
|
|
|---|
| Treatment | TG | FFAs | Ch |
|---|
| Control | 1.0 | 1.0 | 1.0 |
| UVB (5.4
kJ/m2) | 2.3±1.2a | 1.5±0.3a | 2.0±0.5a |
Effect of UVB irradiation on sebaceous
lipogenesis and DGAT activity in DHS
In order to clarify the molecular mechanisms by
which triptolide selectively decreased the TG level on the skin
surface of UVB-irradiated hamsters, the regulation of TG production
and DGAT expression was evaluated in UVB-irradiated DHS. When the
DHS were irradiated with UVB (0.62 kJ/m2) and cultured for 24 h,
TLC analysis indicated that the extracellular levels of TG, FFAs
and Ch were significantly increased compared with non-irradiated
controls (Fig. 4A). In addition, UVB
irradiation of DHS was indicated to significantly increase the
intracellular levels of TG and FFAs compared with non-irradiated
controls, but had no significant effect on the levels of Ch
(Fig. 4B). This indicated that UVB
irradiation increased the levels of TG in DHS. Furthermore, the
enzymatic activity of DGAT, a rate-limiting enzyme of TG synthesis,
was dose-dependently increased by UVB irradiation in DHS (Fig. 4C).
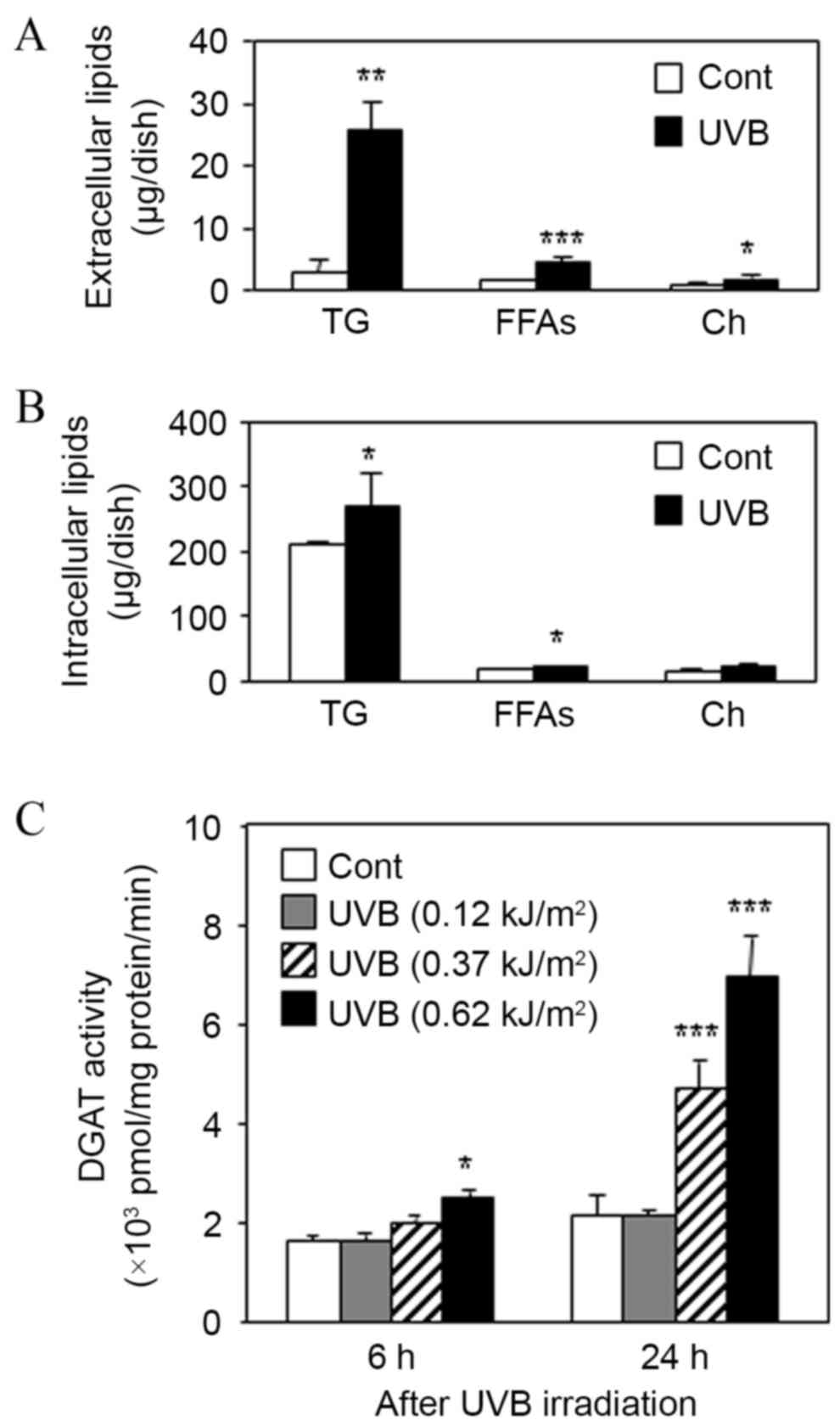 | Figure 4.Effect of UVB irradiation on sebaceous
lipogenesis and DGAT activity in DHS. DHS at the third passage were
irradiated with UVB (0.62 kJ/m2) in serum-free
conditions and then maintained for 24 h. (A) Harvested culture
media and (B) cells were subjected to TLC analysis. (C) DHS at the
third passage were irradiated with UVB at 0.12, 0.37 and 0.62
kJ/m2. After the irradiation, the cells were maintained
for 6 or 24 h, then DGAT activity was measured in the harvested
cells. Data are presented as the mean ± standard deviation of three
dishes. *P<0.05, **P<0.01 and ***P<0.01 vs. untreated
controls. DHS, differentiated hamster sebocytes; TLC, thin-layer
chromatography; DGAT, diglyceride acyltransferase; UVB, ultraviolet
B; TG, triacylglycerol; FFAs, free fatty acids; Ch,
cholesterol. |
Effect of triptolide on UVB-enhanced
TG production and DGAT activity in DHS
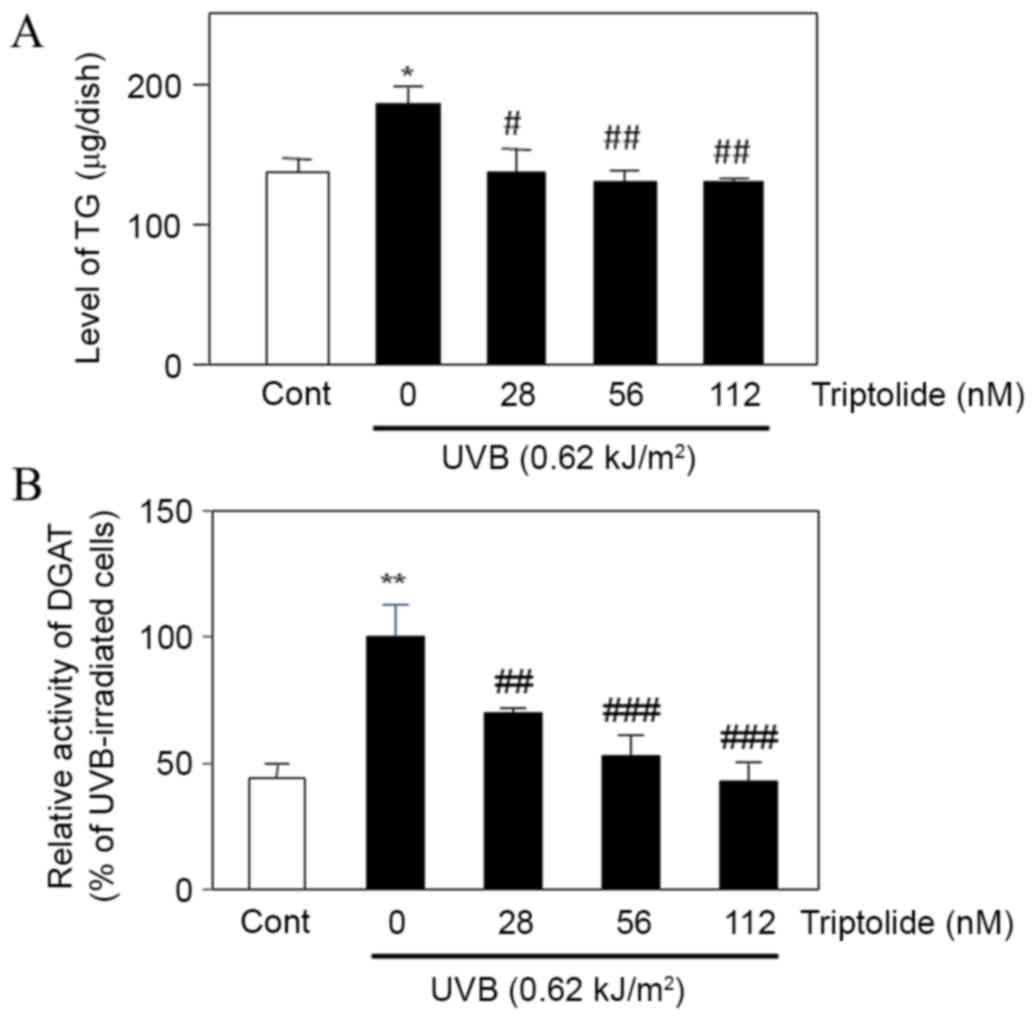
Treatment of UVB-irradiated DHS with triptolide was
found to significantly decrease the intracellular level of TG
(P<0.05 at 28 nM, P<0.01 at 56 and 112 nM triptolide;
Fig. 5A) compared with the
non-treated cells. Triptolide also significantly decreased DGAT
activity in UVB-irradiated triptolide-treated cells compared with
UVB-irradiated non-treated cells (P<0.01 at 28 nM, P<0.001 at
56 and 112 nM triptolide; Fig. 5B).
These results suggested that triptolide suppressed the UVB-enhanced
DGAT activity and TG production in DHS.
Discussion
UVB-irradiation has been reported to facilitate
sebum secretion and production in vivo (9,10), yet
it remains unclear how UVB stimulates sebum production in sebaceous
glands. To the best of our knowledge, the present study is the
first to demonstrate that UVB irradiation increases DGAT activity,
leading to an increase in TG production in DHS. Taken together with
a previous report using DGAT knockout mice, in which DGAT was shown
to be involved in TG biosynthesis and lipid-droplet formation in
adipocytes (25). UVB is likely to
facilitate sebum production due to DGAT-mediated de novo
synthesis of TG in DHS. Furthermore, as most people are exposed to
UV in sunlight on an everyday basis (3), it is not only endogenous factors, such
as androgens and insulin, that serve key roles in the regulation of
sebum production and secretion, but potentially also environmental
ones. Moreover, excess and/or long-term UVB irradiation is likely
to perturb epidermal barrier functions through the abnormal
enhancement of sebum production and secretion, as well as
alterations in the expression pattern of tight junction-related
molecules (26).
Akitomo et al (10) reported that UV exposure facilitates
the peroxidation of skin surface lipids such as TG and Ch, which in
turn affects the cutaneous barrier functions by increasing
transepidermal water loss. Regarding the anti-oxidative actions of
triptolide, previous studies (19,20) have
reported that triptolide exhibits anti-oxidative actions due to the
inhibition of reactive oxygen species levels in the mouse liver,
and superoxide anions in murine peritoneal macrophages. In
addition, the current authors previously demonstrated that a
natural anti-oxidative polymethoxy flavonoid, nobiletin,
predominantly decreases the level of TG, which is the target lipid
for UVB-peroxidation (11).
Therefore, it is suggested that the topical application of
triptolide is effective for the prevention of sebum peroxidation on
the skin, not only due to its own anti-oxidative activity but also
due its ability to inhibit sebaceous TG synthesis.
In the present study, it was indicated that the
extracellular level of TG was increased in UVB-irradiated DHS,
suggesting that sebum excretion in DHS is facilitated by UVB. Since
sebum secretion is generally considered to be regulated by a
holocrine mechanism, which may involve the apoptosis of sebocytes
(27), it was confirmed that there
were no apoptotic cells in this experimental condition (data not
shown). In addition, the current authors have previously reported
on the apoptosis-independent and ATP-binding cassette (ABC)
transporter (ABCB1/P-glycoprotein) -mediated TG secretion in DHS
(28). Regarding the relationship
between ABCB1 activity and UV irradiation, Trindade et al
(29) have reported that
ABCB1-overexpressing leukemia cells with a multidrug-resistance
phenotype are resistant to UVA but sensitive to UVB and UVC.
Further experiments are required to clarify whether ABCB1 is
associated with the UVB-increased TG excretion in DHS.
Acne vulgaris is characterized by: i) Excess sebum
production in sebaceous glands; ii) microcomedone formation; and
iii) the hyperproliferation of Propionibacterium acnes,
which results in inflammatory conditions in acne lesions (8). The aggravation of acne is likely to
result in a disfiguring and permanent scar formation that carries a
psychological and social impact on the patient's quality of life
(8,30,31).
Allen and LoPresti (32) reported
the involvement of sunlight in the aggravation of acne vulgaris,
and the current results indicated that triptolide inhibits
UVB-enhanced sebaceous TG production in vivo and in
vitro. These findings suggest that triptolide could be used in
the future as a therapeutic agent or in cosmetics, for the
effective prevention of sunlight-associated acne aggravation.
In conclusion, the present study demonstrated that
triptolide inhibits UVB-induced sebaceous lipogenesis in hamsters
both in vivo and in vitro. Furthermore, the study has
supplied novel evidence indicating that triptolide decreases
UVB-augmented TG production by suppressing DGAT activity in DHS.
Thus, triptolide is a candidate to be used as a therapeutic agent
or in cosmetics for acne treatment.
Acknowledgements
This study was supported by a Grant-in-Aid for
Scientific Research (C) (grant no. 26460633) awarded to Professor
Takashi Sato.
References
|
1
|
Rabe JH, Mamelak AJ, McElgunn PJ, Morison
WL and Sauder DN: Photoaging: Mechanisms and repair. J Am Acad
Dermatol. 55:1–19. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Quan T, Qin Z, Xia W, Shao Y, Voorhees JJ
and Fisher GJ: Matrix-degrading metalloproteinases in photoaging. J
Invest Dermatol Symp Proc. 14:pp. 20–24. 2009; View Article : Google Scholar
|
|
3
|
Sambandan DR and Ratner D: Sunscreens: An
overview and update. J Am Acad Dermatol. 64:748–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jenkins G: Molecular mechanisms of skin
aging. Mech Ageing Dev. 123:801–810. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Inomata S, Matsunaga Y, Amano S, Takada K,
Kobayashi K, Tsunenaga M, Nishiyama T, Kohno Y and Fukuda M:
Possible involvement of gelatinases in basement membrane damage and
wrinkle formation in chronically ultraviolet B-exposed hairless
mouse. J Invest Dermatol. 120:128–134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tanaka S, Sato T, Akimoto N, Yano M and
Ito A: Prevention of UVB-induced photoinflammation and photoaging
by a polymethoxy flavonoid, nobiletin, in human keratinocytes in
vivo and in vitro. Biochem Pharmacol. 68:433–439. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thody AJ and Shuster S: Control and
function of sebaceous glands. Physiol Rev. 69:383–416.
1989.PubMed/NCBI
|
|
8
|
Kurokawa I, Danby FW, Ju Q, Wang X, Xiang
LF, Xia L, Chen W, Nagy I, Picardo M, Suh DH, et al: New
developments in our understanding of acne pathogenesis and
treatment. Exp Dermatol. 18:821–832. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Suh DH, Kwon TE and Youn IJ: Changes of
comedonal cytokines and sebum secretion after UV irradiation in
acne patients. Eur J Dermatol. 12:139–144. 2002.PubMed/NCBI
|
|
10
|
Akitomo Y, Akamatsu H, Okano Y, Masaki H
and Horio T: Effects of UV irradiation on the sebaceous gland and
sebum secretion in hamsters. J Dermatol Sci. 31:151–159. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sato T, Takahashi A, Kojima M, Akimoto N,
Yano M and Ito A: A citrus polymethoxy flavonoid, nobiletin
inhibits sebum production and sebocyte proliferation, and augments
sebum excretion in hamsters. J Invest Dermatol. 127:2740–2748.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Merle C, Laugel C and Baillet-Guffroy A:
Effect of UVA or UVB irradiation on cutaneous lipids in films or in
solution. Photochem Photobiol. 86:553–562. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kohl E, Steinbauer J, Landthaler M and
Szeimies RM: Skin ageing. J Eur Acad Dermatol Venereol. 25:873–884.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qin WZ, Zhu GD, Yang SM, Han KY and Wang
J: Clinical observations on Tripterygium wilfordii in treatment of
26 cases of discoid lupus erythematosus. J Tradit Chin Med.
3:131–132. 1983.PubMed/NCBI
|
|
15
|
Tao XL, Sun Y, Dong Y, Xiao YL, Hu DW, Shi
YP, Zhu QL, Dai H and Zhang NZ: A prospective, controlled,
double-blind, crossover study of Tripterygium wilfordii Hook F. in
treatment of rheumatoid arthritis. Chin Med J (Engl). 102:327–332.
1989.PubMed/NCBI
|
|
16
|
Jiang X: Clinical observations on the use
of the Chinese herb Tripterygium wilfordii Hook for the treatment
of nephrotic syndrome. Pediatr Nephrol. 8:343–344. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qiu D, Zhao G, Aoki Y, Shi L, Uyei A,
Nazarian S, Ng JC and Kao PN: Immunosuppressant PG490 (triptolide)
inhibits T-cell interleukin-2 expression at the level of
purine-box/nuclear factor of activated T-cells and NF-kappaB
transcriptional activation. J Biol Chem. 274:13443–13450. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin N, Sato T and Ito A: Triptolide, a
novel diterpenoid triepoxide from Tripterygium wilfordii Hook. f.,
suppresses the production and gene expression of pro-matrix
metalloproteinases 1 and 3 and augments those of tissue inhibitors
of metalloproteinases 1 and 2 in human synovial fibroblasts.
Arthritis Rheum. 44:2193–2200. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu Y, Cui J, Bao X, Chan S, Young DO, Liu
D and Shen P: Triptolide attenuates oxidative stress, NF-κB
activation and multiple cytokine gene expression in murine
peritoneal macrophage. Int J Mol Med. 17:141–150. 2006.PubMed/NCBI
|
|
20
|
Lu Y, Bao X, Sun T, Xu J, Zheng W and Shen
P: Triptolide attenuates the oxidative stress induced by LPS/D-GalN
in mice. J Cell Biochem. 113:1022–1033. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kabashima K, Nagamachi M, Honda T,
Nishigori C, Miyachi Y, Tokura Y and Narumiya S: Prostaglandin E2
is required for ultraviolet B-induced skin inflammation via EP2 and
EP4 receptors. Lab Invest. 87:49–55. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sato T, Imai N, Akimoto N, Sakiguchi T,
Kitamura K and Ito A: Epidermal growth factor and
1alpha,25-dihydroxyvitamin D3 suppress lipogenesis in hamster
sebaceous gland cells in vitro. J Invest Dermatol. 117:965–970.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Akimoto N, Sato T, Iwata C, Koshizuka M,
Shibata F, Nagai A, Sumida M and Ito A: Expression of perilipin A
on the surface of lipid droplets increases along with the
differentiation of hamster sebocytes in vivo and in vitro. J Invest
Dermatol. 124:1127–1133. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Iwata C, Akimoto N, Sato T, Morokuma Y and
Ito A: Augmentation of lipogenesis by
15-deoxy-Delta12,14-prostaglandin J2 in hamster sebaceous glands:
Identification of cytochrome P-450-mediated
15-deoxy-Delta12,14-prostaglandin J2 production. J Invest Dermatol.
125:865–872. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harris CA, Haas JT, Streeper RS, Stone SJ,
Kumari M, Yang K, Han X, Brownell N, Gross RW, Zechner R and Farese
RV Jr.: DGAT enzymes are required for triacylglycerol synthesis and
lipid droplets in adipocytes. J Lipid Res. 52:657–667. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamamoto T, Kurasawa M, Hattori T, Maeda
T, Nakano H and Sasaki H: Relationship between expression of tight
junction-related molecules and perturbed epidermal barrier function
in UVB-irradiated hairless mice. Arch Dermatol Res. 300:61–68.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wróbel A, Seltmann H, Fimmel S,
Müller-Decker K, Tsukada M, Bogdanoff B, Mandt N, Blume-Peytavi U,
Orfanos CE and Zouboulis CC: Differentiation and apoptosis in human
immortalized sebocytes. J Invest Dermatol. 120:175–181. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kurihara H, Sato T, Akimoto N, Ogura T and
Ito A: Identification and characterization of ABCB1-mediated and
non-apoptotic sebum secretion in differentiated hamster sebocytes.
Biochim Biophys Acta. 1811:1090–1096. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Trindade GS, Capella MA, Capella LS,
Affonso-Mitidieri OR and Rumjanek VM: Differences in sensitivity to
UVC UVB and UVA radiation of a multidrug-resistant cell line
overexpressing P-glycoprotein. Photochem Photobiol. 69:694–699.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang S, Cho S, Chung JH, Hammerberg C,
Fisher GJ and Voorhees JJ: Inflammation and extracellular matrix
degradation mediated by activated transcription factors nuclear
factor-kappaB and activator protein-1 in inflammatory acne lesions
in vivo. Am J Pathol. 166:1691–1699. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sato T, Kurihara H, Akimoto N, Noguchi N,
Sasatsu M and Ito A: Augmentation of gene expression and production
of promatrix metalloproteinase 2 by Propionibacterium acnes-derived
factors in hamster sebocytes and dermal fibroblasts: A possible
mechanism for acne scarring. Biol Pharm Bull. 34:295–299. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Allen HB and LoPresti PJ: Acne vulgaris
aggravated by sunlight. Cutis. 26:254–256. 1980.PubMed/NCBI
|