Introduction
Recurrent spontaneous abortion (RSA) is defined by
the American Society for Reproductive Medicine (ASRM) as ≥2
consecutive spontaneous abortions, whereas the Royal College of
Obstetricians and Gynecologists and the European Society for Human
Reproduction and Embryology, specify it as the loss of ≥3
consecutive fetuses prior to 24 weeks of pregnancy with the same
partner. Statistically, 15–25% of pregnancies result in miscarriage
and <5% of women will experience two miscarriages, only 1% of
women will experience the loss of a fetus ≥3 times (1,2). The
causes of RSA are considered to include cytogenetic abnormalities,
anatomic irregularities, endocrine disorders, infection,
autoimmunity, atypical blood clotting, sperm quality and
environmental factors (2). However,
in ~50% of RSA cases no clearly defined etiology is identified
(3).
During pregnancy fetal trophoblast cells infiltrate
into the maternal uterine blood vessels and release blood into the
intervillous space (4). The outer
layer of the chorionic villi (syncytiotrophoblast) is bathed
directly in maternal blood, which enables the fetus to easily
acquire nutrients (5). However, the
exposure of trophoblast cells may lead to potential allogeneic
immune responses by the mother. Typically, uterine immune responses
allow the placenta to acquire maternal supplies and also prevent
excessive invasion (4). Disruption
of the normal balance between the itinerant trophoblast cells and
the uterine tissues they colonize during placentation, may result
in various clinical problems (4).
The hypothesis that maternal-fetal immune disorders lead to
miscarriage may be traced back to the 20th century when Medawar
postulated that the fetus may be considered as an allograft to the
mother, and that the absence of a maternal immune response is what
allows embryonic implantation (6).
Natural killer cells (NKs) are the third largest
granular cells and they account for 10–15% of the total lymphocytes
in the blood (7). NKs are activated
by external stimuli and they serve a vital role in immune
regulation and defense (5). A number
of uterine NKs (uNKs) gather in the endometrium prior to
placentation and they make up 75% of all uterine decidua
lymphocytes in early pregnancy (8).
At 20 weeks of pregnancy the uNKs begin to decline and they
disappear by the third trimester (9). Tissue NKs differ from circulating NKs
as their phenotype and function are modified by the local
microenvironment (10,11). A total of 90% of peripheral blood NK
cells (pbNKs) are CD56dim cells, which highly express
CD16 and possess clear cytotoxic activity (12). uNKs are primarily
CD56bright cells, which lack cytotoxicity and instead
secrete various cytokines, which serve an important role in
adjusting the invasion of trophoblastic cells (13,14).
NK activity depends on the integration of signals
from numerous germline-encoded activating and inhibitory receptors,
which are primarily from three families: C-type lectin-like
receptors, immunoglobulin (Ig)-like transcripts and killer cell
Ig-like receptors (KIRs) (15,16).
KIRs are encoded by an array of highly polymorphic genes located on
chromosome 19q13.4, which produce wide variations in KIR structure
and combination (17). KIRs are
classified according to their structure and function. Each KIR
molecule consists of two or three extracellular Ig domains (2D and
3D molecules, respectively), a transmembrane section and a short
(S) or long (L) intracellular tail (18). Each inhibitory KIR receptor contains
an immune receptor tyrosine inhibition motif in its cytoplasmic
domain, which may interact with SH2-containing protein-tyrosine
phosphatase-1 in the cytoplasm, and inhibit the activation of the
NK cell (19). Each activating KIR,
possesses a short charged cytoplasmic tail, which combines with the
immune receptor tyrosine activation sequence or DAP12 adapter
protein, thus enhancing activation signals and gene expression in
NKs (16,20). To the best of our knowledge, 15 KIR
gene loci and two pseudogenes have been identified to date, among
which a marked linkage disequilibrium exists (21). Two common haplotypes, designated A
and B, have been defined based on the number of KIR genes present.
The A haplotype is less variable and is recognized by the presence
of a minimum of 6 inhibitory receptor genes (KIR-2DL1, −2DL3,
−2DL4, −3DL1, −3DL2 and −3DL3) as well as the only activating
receptor gene (KIR2DS4) (22). The B
haplotype has a more variable and numerous gene content, which
differs due to the various possible combinations of activating
receptor genes (23). There are
three distinct groups of KIR genotype in humans, which are KIR-AA,
-AB and -BB. At present, >550 different KIR genotype IDs have
been described, which has greatly enriched the understanding of the
KIR gene family (allelefrequencies.net/kir6001a.asp; update in
2015).
KIRs combine with human leukocyte antigen (HLA)
ligands on the surface of target cells and are closely associated
with virus infection, malignancy, autoimmune diseases, organ
transplantation and the process of pregnancy (24–27). In
recent decades, an increasing number of studies have indicated that
KIRs may be associated with RSA. Certain previous studies consider
a maternal KIR-AA homozygous genotype as a protective factor, which
inhibits the activation of uNKs and prevents the fetus from being
rejected (28–30). However, others have reported that the
KIR-B haplotype promotes uNK activation and cytokine secretion,
which is conducive to trophoblastic cell implantation into the
endometrium and thus promotes a successful pregnancy (31,32). It
is possible that some specific combinations of KIR genes may serve
a role in the autoimmune mechanisms underlying recurrent
miscarriage. Individuals from different regions and ethnic groups
often have large differences in the frequencies of KIR genotypes.
The frequency of KIR-AA ranges from 1.5% in the Australian
aborigines to 67.9% in the China Yunnan Province Nu (33,34). As
numerous factors may lead to miscarriage, it is necessary to
consider these while analyzing the association between KIR and
spontaneous abortion. Considering the geographical and ethnic
differences that are observed in the KIR and HLA-C genes, the
present study investigated only those of a Han Chinese ethnicity.
The current study detected the KIR and HLA-C genes of 110
unexplained RSA and 105 healthy females. The aim of the present
study was to determine if certain KIR genotypes influence the
occurrence of spontaneous miscarriage.
Materials and methods
Study subjects and samples
In the present study the ASRM definition of RSA (as
defined above) was adhered to. Couples with ≥2 recurrent
spontaneous miscarriages prior to 20 weeks of pregnancy and no live
births were recruited from Henan Provincial People's Hospital
(Henan, China) between September 2015 and September 2016. The
parental chromosomes were karyotyped. At day 3, the
follicle-stimulating hormone, luteinizing hormone and testosterone
levels were measured in the females in a clinical laboratory at the
Henan Provincial People's Hospital (Henan, China) and the results
were reported to us, an ultrasound examination or hysteroscopy was
performed prior to the current study, semen quality was measured
and a thyroid function test was performed in order to rule out
abnormal thyroid function, which is related to miscarriage [thyroid
stimulating hormone (TSH), triiodothyronine (T3), thyroxine (T4),
free triiodothyronine (FT3), free thyroxine (FT4), anti-thyroid
peroxidase antibodies (anti-TPO) and antithyroglobulin antibody
(anti-TG)] (35). The blood glucose
level and level of immune antibodies, including antiphospholipid,
endometrial, sperm, anti human chorionic gonadotropin, anti zona
pellucida and anti ovarian antibodies were measured and a TORCH-IgM
(TOX-IgM, RV-IgM, CMV-IgM and HSV-IgM) screen was performed.
Studies have demonstrated that TORCH-IgM antibodies have a strong
association with the occurrence of RSA (36). Mycoplasma and chlamydia infection
were tested for and blood coagulation function (PT, ARTT, FIB, TT)
and D-dimer tests were performed identify if patients met inclusion
criteria. The hypercoagulation of blood is associated with the
occurrence of miscarriage (37).
Of the 188 patients tested who had normal
chromosomes, 78 were excluded because the suspected reasons for
their miscarriage were identified. Consequently, there were 110
couples of Han Chinese ethnicity that met the aforementioned
conditions and were recruited into the experimental (RSA) group
(age, 20–39 years). Each of the females enrolled in the study had
no history of poisonous or teratogenic chemical exposure, lived a
healthy lifestyle and had never fallen pregnant with a previous
partner. The control group consisted of 105 healthy females (age,
19–39 years) recruited from the Henan Red Cross Blood Center
between November 2015 and September 2016, who had given birth to a
minimum of 1 healthy child and had no history of miscarriage. The 2
groups had the same ethno-geographic origin and there were no
differences in ages. All participants were informed about the study
and written informed consent was obtained. Ethical approval for the
present study was obtained from the Board of Ethics Committee of
the Henan Provincial People's Hospital. Blood samples (5 ml) taken
from all female participants were preserved in tubes containing
EDTA at 4°C prior to genetic evaluation.
DNA isolation
Maternal genomic DNA was isolated from 5 ml of blood
using the TIANamp Genomic DNA kit (Tiangen Biotech Co., Ltd.,
Beijing, China). The quantity and quality of the DNA samples was
detected using a NanoDrop™ 2000 spectrophotometer at
wavelengths of 230, 260 and 280 nm (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The experiment procedure was conducted according
to the manufacturer's protocol.
KIR genotyping
KIR genotyping was performed on maternal genomic DNA
using a KIR Genotyping kit (cat. no. 54410D; Invitrogen; Thermo
Fisher Scientific, Inc.), which detected the presence or absence of
KIR genes using polymerase chain reaction (PCR)-sequence specific
primers. Formulations of locus specific primers from the kit were
used to amplify genomic DNA. The reaction buffer was mixed with the
genomic DNA sample and Taq DNA polymerase (cat. no. M8296; Promega
Corporation, Madison, WI, USA) in a 96-well thermal tray.
Thermocyling was performed as follows: Initial denaturation of 95°C
for 1 min; 30 cycles of denaturation at 94°C for 20 sec, annealing
at 63°C for 20 sec and extension at 72°C for 90 sec. 2% agarose gel
preparation was performed as follows: 2 g of agarose powder was
poured into a 250 ml conical flask and 100 ml of 0.5X
tetrabromoethane was dissolved in the flask. The microwave oven was
heated to boiling and ethidium bromide (cat. no. 15585011; Thermo
Fisher Scientific, Inc.) (6 µl) was added. The flask was shaken and
the mixture was poured into the template and cooled for 30 min.
When the thermocycling process was complete, 5 µl PCR products were
loaded onto a 2% agarose gel using a pipette. Electrophoresis was
carried out in the electrophoresis tank, which was filled with 0.5X
tetrabromoethane. The gel was run at a voltage of 150 V for 20 min.
Following electrophoresis, the gel was placed under UV light to
observe and record the results.
HLA-C genotyping
For HLA-C1 and -C2 genotyping, the same primers were
used as previously reported by Tajik et al (38). The PCR system in each reaction was 12
µl in total [60 ng genomic DNA (1 µl), 2X Taq PCR Master mix (Bio
Basic, Inc., Markham, ON, Canada) (5 µl), nuclease-free water (5
µl), forward (0.5 µl) and reverse (0.5 µl) primers]. PCR conditions
were: Initial denaturation for 2 min at 94°C; 10 cycles of 10 sec
denaturation at 94°C and 60 sec annealing and elongation at 65°C;
20 cycles of 10 sec denaturation at 94°C, 50 sec annealing at 61°C
and 30 sec elongation at 72°C; with a final extension step at 72°C
for 10 min. When the thermocycling process was complete, a total of
6 µl loading buffer (cat. no. B648314; Sangon Biotech Co., Ltd.,
Shanghai, China) was mixed with the amplification products and
electrophoresed on 2% agarose gels. 2% agarose gel preparation was
performed same as the above method and ethidium bromide was used as
a DNA stain. The gel was run at a voltage of 150 V for 25 min.
Following electrophoresis, the gel was placed under UV light
(WD-9403C; Beijing Liuyi Biotechnology Co., Ltd., Beijing, China)
to observe and record the results.
Statistical analysis
The percentage of KIR and HLA-C genes in the present
study was determined by direct counting. The frequency of the two
populations of KIR haplotypes (A and B) and HLA-C alleles (C1 and
C2) were obtained using the Hardy-Weinberg principle
(p2+2pq+q2=1) (39). All data were statistically analyzed
using Chi-square analysis, Pearson Chi-square continuity correction
and Fisher's exact test, on SPSS version 17 software (SPSS, Inc.,
Chicago, IL, USA). P≤0.05 was considered to indicate a
statistically significant difference. Data are presented as numbers
and percentages. Odds ratios (ORs) and 95% confidence intervals
(CIs) were calculated.
Results
Carrier frequency of KIR genes in the
RSA and control group
A total of 19 KIR genes were genotyped in all
participants: 2DL1, 2DL2, 2DL3, 2DL4, 2DL5A, 2DL5B, 2DS1, 2DS2,
2DS3, 2DS4FUL, 2DS4DEL (2DS4 allele with a 22 base pair deletion),
2DS5, 3DL1, 3DL2, 3DL3, 3DS1, 2DP1, 3DP1FUL and 3DP1DEL (absence of
exon 2 and its flanking intron sequences) (Table I and Fig.
1). The 4 framework genes (conservative gene positions fixed
and present in each person's KIR gene), KIR2DL4, 3DL2, 3DL3 and
3DP1FUL or 3DP1DEL were present in all of the samples. The KIR
pseudogene KIR2DP1 was present in all samples from the normal
group, but only 99.1% of the samples from the experimental RSA
group. In all participants, it was identified that compared with
the activating KIR genes, the inhibitory KIR genes had aT higher
frequency. Among the inhibitory KIR genes, KIR2DL1 demonstrated the
highest frequency, as it was present in 99.1 and 100% of the
experimental and control groups, respectively. The other most
frequent inhibitory KIR genes were KIR2DL3 and KIR3DL1, which were
present in ≥91.8% in each group. Apart from KIR2DS4 positive
genotypes (KIR2DS4FUL, KIR2DS4DEL or combined genotypes), the
frequency of the activating genes was <40%. KIR2DS2 demonstrated
the lowest observed carrier frequency of any activating gene, as it
was present in 18.2% of the experimental group and 16.2% of the
control group. No statistically significant differences were
identified between the frequency of each KIR gene in the
experimental group and the control group. Additionally, no
significant differences were identified in the frequency of the
inhibitory and activated KIR genes in each group.
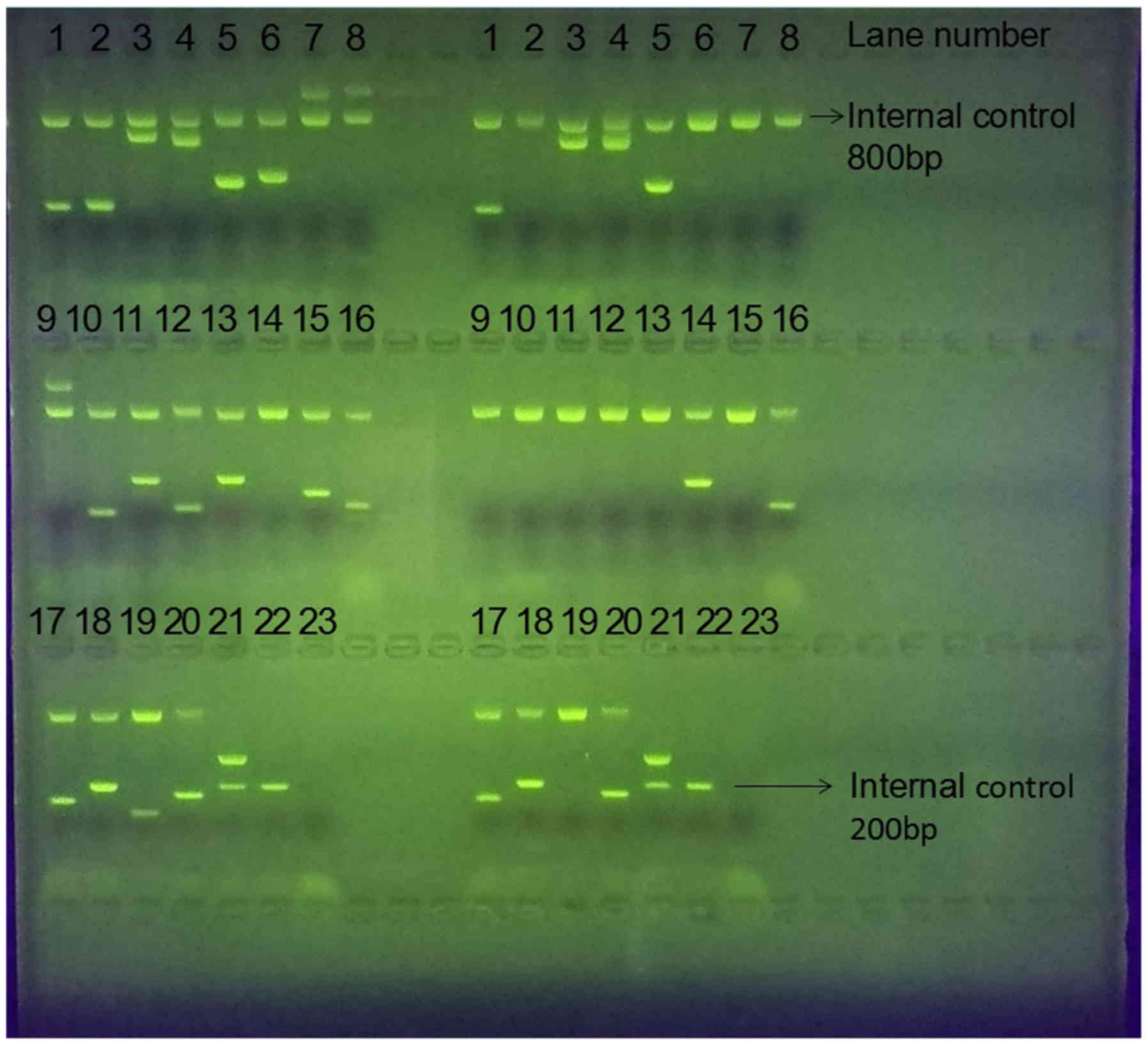 | Figure 1.Representative agarose gel
electrophoresis results. The DNA of two participants was separated
by agarose gel electrophoresis to reveal their KIR genes. Each
individual occupies 23 lanes consisting of 22 experiment lanes and
one blank control lane (some genes were assigned duplicate lanes).
In each experiment lane, there is an internal control polymerase
chain reaction fragment (800 bp in lanes 1–20 and 200 bp in lanes
21–22). The bright bands in the experiment lanes correspond to the
KIR genes. The two individuals' positive KIR genes are 2DL1, 2DL3,
2DL4, 2DS4FUL, 3DL1, 3DL2, 3DL3, 2DP1, 3DP1DEL (on the left) and
2DL1, 2DL3, 2DL4, 2DS4FUL, 2DS4DEL, 3DL1, 3DL2, 3DL3, 2DP1, 3DP1DEL
(on the right). KIR, killer cell immunoglobulin-like receptors; bp,
base pairs. |
 | Table I.Comparison of the frequency of KIR
genes in the RSA (n=110) and control (n=105) groups. |
Table I.
Comparison of the frequency of KIR
genes in the RSA (n=110) and control (n=105) groups.
|
|
| RSA group | Control group |
|
|
|
|---|
|
|
|
|
|
|
|
|
|---|
| KIR gene | n | % | n | % | P-value | Odds ratio | 95% confidence
interval |
|---|
| Inhibitory | 2DL1 | 109 | 99.1 | 105 | 100.0 | 1.000 | 0.991 | 0.973–1.010 |
| Inhibitory | 2DL2 | 21 | 19.1 | 17 | 16.2 | 0.591 | 1.221 | 0.589–2.532 |
| Inhibitory | 2DL3 | 108 | 98.2 | 105 | 100.0 | 0.497 | 0.982 | 0.956–1.008 |
| Inhibitory | 2DL4 | 110 | 100.0 | 105 | 100.0 | 1.000 | – | – |
| Inhibitory | 2DL5A | 41 | 37.3 | 37 | 35.2 | 0.757 | 1.095 | 0.615–1.950 |
| Inhibitory | 2DL5B | 10 |
9.1 | 10 |
9.5 | 0.922 | 0.954 | 0.367–2.477 |
| Activating | 2DS1 | 44 | 40.0 | 40 | 38.1 | 0.783 | 1.083 | 0.614–1.912 |
| Activating | 2DS2 | 20 | 18.2 | 17 | 16.2 | 0.708 | 1.151 | 0.552–2.401 |
| Activating | 2DS3 | 26 | 23.6 | 23 | 21.9 | 0.774 | 1.102 | 0.568–2.135 |
| Activating | 2DS5 | 31 | 28.2 | 23 | 21.9 | 0.304 | 1.401 | 0.736–2.666 |
| Inhibitory | 3DL1 | 101 | 91.8 | 101 | 96.2 | 0.190 | 0.442 | 0.127–1.543 |
| Inhibitory | 3DL2 | 110 | 100.0 | 105 | 100.0 | 1.000 | – | – |
| Inhibitory | 3DL3 | 110 | 100.0 | 105 | 100.0 | 1.000 | – | – |
| Activating | 3DS1 | 42 | 38.2 | 35 | 33.3 | 0.470 | 1.238 | 0.694–2.210 |
| Pseudogene | 2DP1 | 109 | 99.1 | 105 | 100.0 | 1.000 | 0.991 | 0.973–1.010 |
| Activating | 2DS4.FUL | 81 | 73.6 | 80 | 76.2 | 0.672 | 0.871 | 0.459–1.651 |
| Activating | 2DS4.DEL | 56 | 50.9 | 47 | 44.8 | 0.388 | 1.277 | 0.733–2.227 |
| Pseudogene | 3DP1.FUL | 15 | 13.6 | 8 |
7.6 | 0.168 | 1.914 | 0.751–4.874 |
| Pseudogene | 3DP1.DEL | 109 | 99.1 | 105 | 100.0 | 1.000 | 0.991 | 0.973–1.010 |
The experimental group was subsequently divided into
2 smaller groups depending on the number of miscarriages undergone
by each participant. The frequency of the KIR3DL1 gene was
significantly reduced in women who had aborted ≥3 times compared
with the control group (P=0.025; OR, 0.277; CI, 0.085–0.904;
Table II). However, there was still
no significant difference identified between the frequency of the
inhibitory and activating KIR genes in these 2 groups.
| Table II.Comparison of the frequency of KIR
alleles in women who had miscarriage ≥3 times (n=40) and the
control group (n=105). |
Table II.
Comparison of the frequency of KIR
alleles in women who had miscarriage ≥3 times (n=40) and the
control group (n=105).
|
|
| Patients | Control group |
|
|
|
|---|
|
|
|
|
|
|
|
|
|---|
| KIR gene | n | % | n | % | P-value | Odds ratio | 95% confidence
interval |
|---|
| Inhibitory | 2DL1 | 39 | 97.5 | 105 | 100.0 | 0.340 | 0.975 | 0.945–1.006 |
| Inhibitory | 2DL2 | 6 | 15.0 | 17 | 16.2 | 0.815 | 0.913 | 0.425–1.960 |
| Inhibitory | 2DL3 | 39 | 97.5 | 105 | 100.0 | 0.340 | 0.975 | 0.945–1.006 |
| Inhibitory | 2DL4 | 40 | 100.0 | 105 | 100.0 | 1.000 | – | – |
| Inhibitory | 2DL5A | 14 | 35.0 | 37 | 35.2 | 0.976 | 0.991 | 0.555–1.772 |
| Inhibitory | 2DL5B | 2 |
5.0 | 10 |
9.5 | 0.220 | 0.501 | 0.164–1.537 |
| Activating | 2DS1 | 15 | 37.5 | 40 | 38.1 | 0.930 | 0.975 | 0.550–1.727 |
| Activating | 2DS2 | 6 | 15.0 | 17 | 16.2 | 0.815 | 0.913 | 0.425–1.960 |
| Activating | 2DS3 | 8 | 20.0 | 23 | 21.9 | 0.741 | 0.892 | 0.451–1.763 |
| Activating | 2DS5 | 12 | 30.0 | 23 | 21.9 | 0.191 | 1.528 |
0.807–2.894 |
| Inhibitory | 3DL1 | 35 | 87.5 | 101 | 96.2 |
0.025a | 0.277 | 0.085–0.904 |
| Inhibitory | 3DL2 | 40 | 100.0 | 105 | 100.0 | 1.000 | – | – |
| Inhibitory | 3DL3 | 40 | 100.0 | 105 | 100.0 | 1.000 | – | – |
| Activating | 3DS1 | 14 | 35.0 | 35 | 33.3 | 0.800 | 1.079 | 0.601–1.935 |
| Pseudogene | 2DP1 | 39 | 97.5 | 105 | 100.0 | 0.340 | 0.975 | 0.945–1.006 |
| Activating | 2DS4.FUL | 28 | 70.0 | 80 | 76.2 | 0.323 | 0.729 | 0.389–1.366 |
| Activating | 2DS4.DEL | 17 | 42.5 | 47 | 44.8 | 0.743 | 0.911 | 0.521–1.593 |
| Pseudogene | 3DP1.FUL | 5 | 12.5 | 8 |
7.6 | 0.249 | 1.737 | 0.673–4.481 |
| Pseudogene | 3DP1.DEL | 39 | 97.5 | 105 | 100.0 | 0.340 | 0.975 | 0.945–1.006 |
KIR genotypes
The KIR genotypes were classified into three groups
(AA, AB and BB) in accordance with the following rules: i) The four
framework genes KIR2DL4, 3DL2, 3DL3 and 3DP1 were present in all
haplotypes; ii) the A haplotype contained at least six loci
encoding inhibitory receptors (KIR3DL3, 2DL3, 2DL1, 3DP1, 2DL4,
3DL1 and 3DL2) and only one activating receptor (KIR2DS4) (40); and iii) if activating receptors
characteristic of the B haplotypes were detected, the participants
were designated as having either an AB genotype or a BB
genotype.
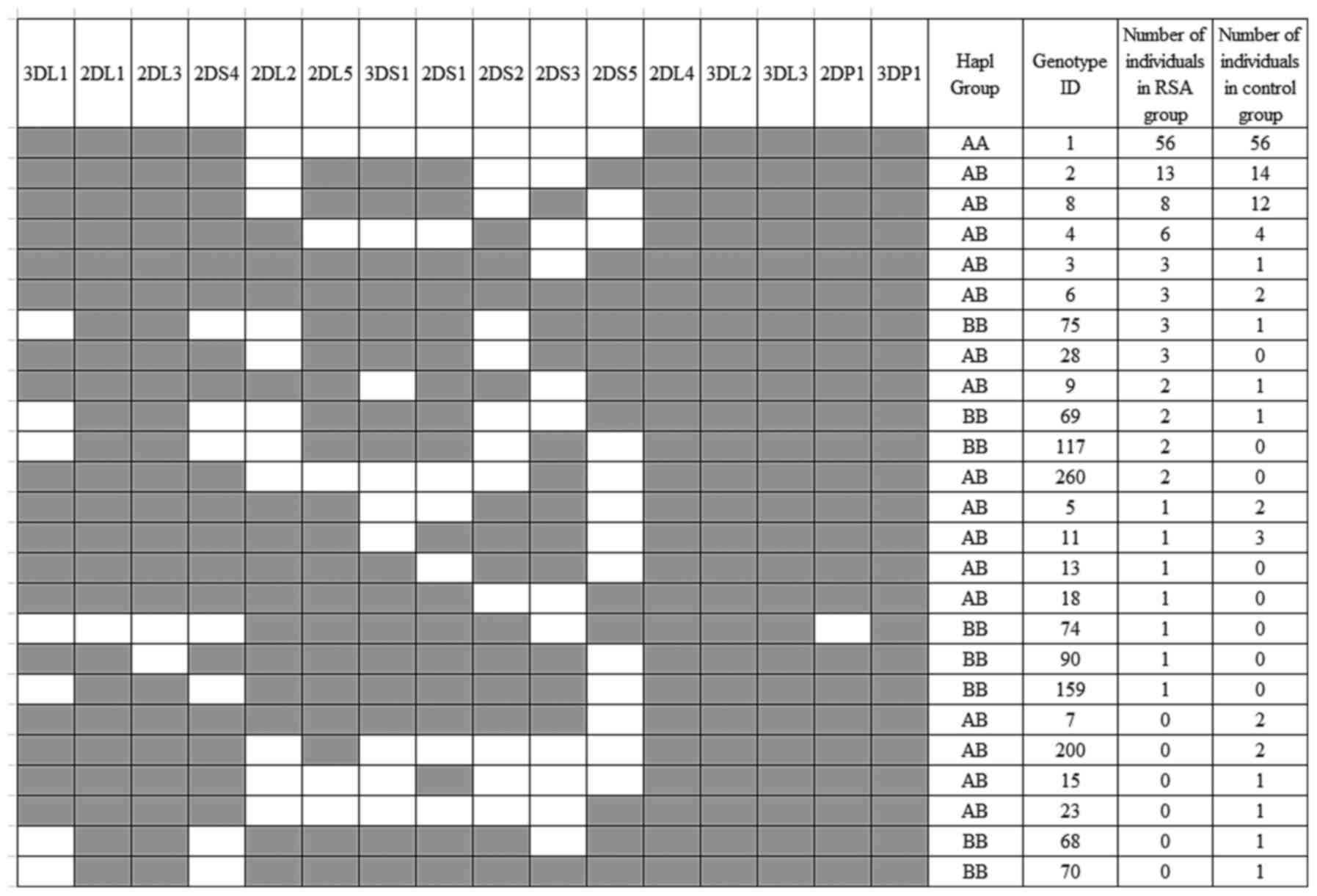
In the present study, the RSA group was revealed as
having 19 different genotypes, whereas the control group only had
17 (Fig. 2). The frequency of KIR
genotypes was similar in the RSA and control group. The most
frequent genotype identified in all participants (50.9% of the RSA
group and 53.3% of the control group) consisted of the 3DL1, 2DL1,
2DL3, 2DS4, 2DL4, 3DL2, 3DL3, 2DP1 and 3DP1 genes, which
corresponds with an AA genotype (genotype ID 1; Table III). The KIR AB genotypes accounted
for 40.0 and 42.9% of patients in the RSA and control groups,
respectively. The frequency of KIR BB genotypes was revealed as
9.1% of patients in the RSA group, compared with 3.8% of patients
in the control group (P=0.127; OR, 2.534; CI, 0.740–8.679). The
haplotypes may also be subdivided into centromeric and telomeric
contents: Cen-A (2DL3 and 2DL1), Tel-A (3DL1 and 2DS4), Cen-B
(2DS2, 2DL2, 2DL5B and 2DS3) and Tel-B (3DS1, 2DL5A, 2DS5 and
2DS1). Comparing the control group with the RSA group, no
significant differences were identified between the centromeric and
telomeric classifications exhibited.
| Table III.Frequency of the KIR genotypes
carriers in the RSA (n=110) and the control (n=105) groups. |
Table III.
Frequency of the KIR genotypes
carriers in the RSA (n=110) and the control (n=105) groups.
|
| RSA group | Control group |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| KIR genotype | n | % | n | % | P-value | Odds ratio | 95% confidence
interval |
|---|
| KIR |
| AA | 56 | 50.9 | 56 | 53.3 | 0.734 | 0.908 | 0.521–1.582 |
| AB | 44 | 40.0 | 45 | 42.9 | 0.677 | 0.887 | 0.505–1.558 |
| BB | 10 |
9.1 | 4 |
3.8 | 0.127 | 2.534 | 0.740–8.679 |
| Cen |
| AA | 71 | 64.5 | 74 | 70.5 | 0.365 | 0.760 | 0.420–1.377 |
| AB | 37 | 33.6 | 31 | 29.5 | 0.533 | 1.209 | 0.665–2.198 |
| BB | 2 |
1.8 | 0 |
0.0 | 0.497 | 0.982 | 0.956–1.008 |
| Tel |
| AA | 65 | 59.1 | 62 | 59.0 | 0.989 | 1.004 | 0.571–1.764 |
| AB | 36 | 32.7 | 39 | 37.1 | 0.514 | 0.824 | 0.460–1.475 |
| BB | 9 |
8.2 | 4 |
3.8 | 0.190 | 2.261 | 0.648–7.889 |
When the women who had miscarried ≥3 times were
compared with the control group, the rate of the BB haplotype was
significantly higher (P=0.025; OR, 3.617; CI, 1.107–11.818;
Table IV). Additionally, the
frequency of the Tel-BB haplotype was also significantly higher in
the women who have aborted ≥3 times compared with the control group
(P=0.025; OR, 3.617; CI, 1.107–11.818).
| Table IV.Frequency of KIR genotypes carriers
in women who had miscarriage ≥3 times (n=40) and the control group
(n=105). |
Table IV.
Frequency of KIR genotypes carriers
in women who had miscarriage ≥3 times (n=40) and the control group
(n=105).
|
| RSA group with ≥3
miscarriages | Controls |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| KIR genotype | n | % | n | % | P-value | Odds ratio | 95% confidence
interval |
|---|
| KIR |
| AA | 22 | 55.0 | 56 | 53.3 | 0.809 | 1.071 | 0.614–1.868 |
| AB | 13 | 32.5 | 45 | 42.9 | 0.129 | 0.641 | 0.360–1.140 |
| BB | 5 | 12.5 | 4 |
3.8 | 0.025a | 3.617 | 1.107–11.818 |
| Cen |
| AA | 27 | 67.5 | 74 | 70.5 | 0.646 | 0.869 | 0.477–1.583 |
| AB | 12 | 30.0 | 31 | 29.5 | 0.938 | 1.024 | 0.559–1.878 |
| BB | 1 |
2.5 | 0 |
0.0 | 0.246 | 0.975 | 0.945–1.006 |
| Tel |
| AA | 25 | 62.5 | 62 | 59.0 | 0.612 | 1.158 | 0.656–2.044 |
| AB | 10 | 25.0 | 39 | 37.1 | 0.064 | 0.565 | 0.308–1.038 |
| BB | 5 | 12.5 | 4 |
3.8 | 0.025a | 3.617 | 1.107–11.818 |
Frequency of HLA-C alleles in RSA
patients and controls
The frequencies of the HLA-C alleles C1 and C2, in
the RSA and control groups were analyzed. No significant
differences were identified between the two groups (Tables V and VI). The present study also analyzed the
distribution of the HLA-C genotypes of KIR AA, AB and BB in the 2
groups (Tables V and VI). The frequency of HLA-C2C2 in KIR AB
and KIR BB was significantly higher in the RSA group compared with
the control group (P=0.024 for KIR AB; P<0.001 for KIR BB;
Table V and P=0.014 for KIR AB;
P<0.001 for KIR BB; Table VI).
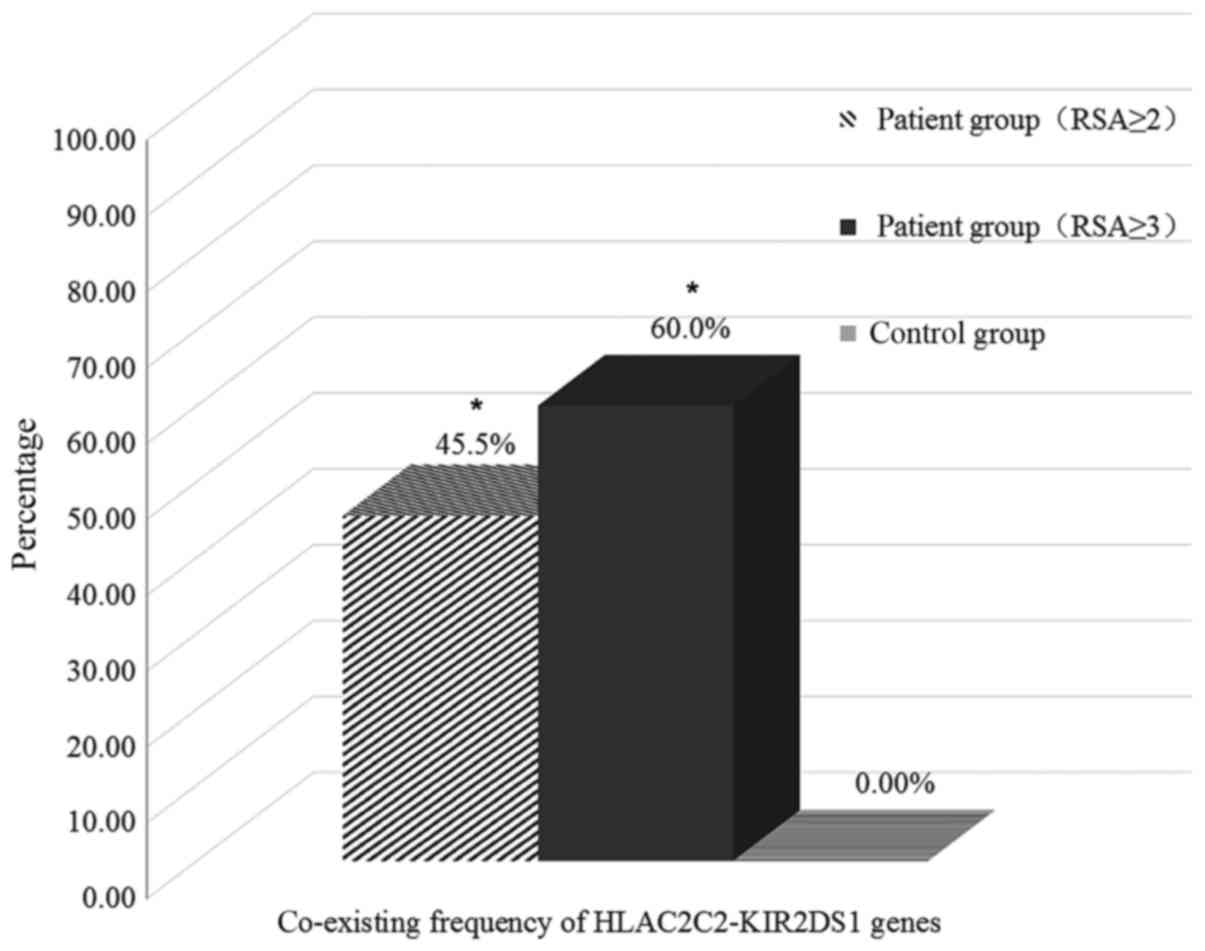
The co-existing frequency of KIR2DS1-C2 genes in the RSA group was
slightly higher than in the control group, however it was not
statistically different (20.9 vs. 18.1%, P=0.617). The patients in
the RSA group who had a homozygous HLA-C2C2 had a significantly
higher frequency of the 2DS1 gene compared with the control group
(45.5 vs. 0.0%, respectively; P<0.001; Fig. 3).
| Table V.Comparison of the frequency of HLA-C
genotypes in the RSA (n=110) and control (n=105) groups. |
Table V.
Comparison of the frequency of HLA-C
genotypes in the RSA (n=110) and control (n=105) groups.
|
|
| RSA group | Control group |
|
|---|
|
|
|
|
|
|
|---|
| KIR genotype | HLA-C genotype | n | % | n | % | P-value |
|---|
| Total no. of
genotype carriers in each group | C1C1 | 53 | 48.2 | 48 | 45.7 | 0.723 |
|
| C1C2 | 46 | 41.8 | 50 | 47.6 | 0.409 |
|
| C2C2 | 11 | 10.0 | 7 |
6.7 | 0.399 |
| KIRAA | C1C1 | 25 | 44.6 | 21 | 37.5 | 0.307 |
|
| C1C2 | 25 | 44.6 | 28 | 50.0 | 0.444 |
|
| C2C2 | 6 | 10.7 | 7 | 12.5 | 0.691 |
| KIRAB | C1C1 | 24 | 54.5 | 25 | 55.6 | 0.876 |
|
| C1C2 | 17 | 38.6 | 20 | 44.4 | 0.405 |
|
| C2C2 | 3 |
6.8 | 0 |
0.0 | 0.024a |
| KIRBB | C1C1 | 4 | 40.0 | 2 | 50.0 | 0.155 |
|
| C1C2 | 4 | 40.0 | 2 | 50.0 | 0.155 |
|
| C2C2 | 2 | 20.0 | 0 |
0.0 |
<0.001a |
| Table VI.Comparison of the frequency of HLA-C
genotypes in women who miscarried ≥3 times (n=40) and the control
group (n=105). |
Table VI.
Comparison of the frequency of HLA-C
genotypes in women who miscarried ≥3 times (n=40) and the control
group (n=105).
|
|
| RSA group with ≥3
miscarriages | Controls |
|
|---|
|
|
|
|
|
|
|---|
| KIR genotype | HLA-C genotype | n | % | N | % | P-value |
|---|
| Total no. of
carriers in each group genotype | C1C1 | 19 | 47.5 | 48 | 45.7 | 0.799 |
|
| C1C2 | 16 | 40.0 | 50 | 47.6 | 0.279 |
|
| C2C2 | 5 | 12.5 | 7 |
6.7 | 0.164 |
| KIRAA | C1C1 | 9 | 40.9 | 21 | 37.5 | 0.622 |
|
| C1C2 | 11 | 50.0 | 28 | 50.0 | 1.000 |
|
| C2C2 | 2 |
9.1 | 7 | 12.5 | 0.439 |
| KIRAB | C1C1 | 8 | 61.5 | 25 | 55.6 | 0.397 |
|
| C1C2 | 4 | 30.8 | 20 | 44.4 | 0.047a |
|
| C2C2 | 1 |
7.7 | 0 |
0.0 | 0.014a |
| KIRBB | C1C1 | 2 | 40.0 | 2 | 50.0 | 0.155 |
|
| C1C2 | 1 | 20.0 | 2 | 50.0 |
<0.001a |
|
| C2C2 | 2 | 40.0 | 0 |
0.0 |
<0.001a |
Discussion
During a normal pregnancy, a fetus grows in the
maternal womb for nearly 40 weeks as an allogeneic entity without
causing immune rejection. Immune dysfunction at the maternal-fetal
interface may be associated with RSA (41). Previous studies have revealed that
uNKs serve an important role in regulating the invasion of
trophoblastic cells (13,14). During pregnancy a large number of
uNKs converge in the maternal decidua and express KIRs (8). Meanwhile, the placental villous
trophoblastic cells express HLA-C, -E, and -G molecules, which
predominantly combine with uNK receptors (42–44). As
fetal trophoblast cells make direct contact with the uterine
tissues, appropriate recognition and immune responses in the
maternal-fetal interface are essential for fetal growth and to
prevent excessive placenta invasion or allogeneic rejection
(4). Human uNKs secrete abundant
cytokines and angiogenic growth factors, which serve a prominent
role in trophoblast cell invasion and placentation (13,14).
Changes to the number or function of uNKs may lead to the
development of an abnormal uterine local environment, which makes
it difficult for the fetus to survive (45). Several previous studies have
demonstrated that certain deleterious HLA-KIR pairings affect the
secretion of cytokines and the expression of receptors, which may
result in a range of pregnancy complications, including
pre-eclampsia, fetal growth restriction and RSA (27,31,46).
A recent study demonstrated that significant
differences exist in the frequency of KIR3DL1, 2DS4, 2DS1 and 2DS5
genes in 10 Han Chinese populations form different regions in China
(47). This suggests that
investigators who have considered the differences between ethnic
groups, but have not considered the differences in geographical
regions, may form mistaken conclusions when analyzing the
correlations between KIR and disease. In the present study, the
variation of KIRs in different ethnic and geographical groups was
fully considered and all participants recruited to the study were
from the same area and of the same ethnicity with no other regional
migration known for three generations. The KIR and HLA-C genes
present in the Han Chinese normal women and patients with RSA were
detected, and it was investigated whether certain genotypes were
more susceptible to RSA. To the best of our knowledge, the present
study is the first time an association has been demonstrated
between the uNK KIR and HLA-C genes in women of Han Chinese
ethnicity with RSA. As it was not possible to collect all of the
aborted fetuses from patients with RSA, there is a possibility that
chromosomal aneuploidy in the fetus may have led to miscarriage,
although this is unlikely to have been the cause in all cases. A
previous study evaluated the karyotypes of the products of
conception in women with RSA and revealed the frequency of abnormal
chromosomes did not differ from the whole population (48).
The mechanism by which NK cells develop a tolerance
to normal self-tissues depends mainly on the ‘missing self’
mechanism (49). In a normal
situation, inhibitory KIRs interact with specific cognate HLA
ligands that generate inhibitory signals and thereby prevent NK
cells from being activated (50).
The interactions of activation receptors with their self-specific
ligands may result in NK cells hyporesponsiveness or reduced
expression of cognate receptors (51). A previous study revealed that many NK
cells with anti-HLA-C2 reactivity were present in HLA-C1 homozygous
and heterozygous healthy individuals with 2DS1, but not in HLA-C2
homozygous donors (52). This
suggests that the functions of NK cells in vivo are
critically regulated. A self-stabilization mechanism may exist in
NK cells, allowing them to maintain homeostasis by self-regulating,
however, when interference forces are above the self-regulation
ability, referred to as the ‘threshold effect’, an immune attack
will be activated and external factors are required to restore
homeostasis (53).
In the present study, it was demonstrated that the
KIR AA haplotype frequency of the two groups was similar, which was
in line with the results reported by other studies in Han Chinese
populations (54,55). No statistical significances were
identified in the frequency of KIR genes in the RSA group compared
with the control group. No significant difference was identified in
the frequency of HLA-C alleles between the patients with RSA and
the controls. A similar conclusion was reached by Christiansen
et al (56). In the present
study, the frequency of the KIR3DL1 gene was significantly reduced
in women who had aborted ≥3 times (n=40) compared with the control
(n=105). The inhibitory KIR3DL1 gene is one of the most polymorphic
KIR genes (57). The ligand for the
KIR3DL1 receptor is the Bw4 epitope present on several HLA-B
molecules, which is not expressed by fetal trophoblast cells
(58). In patients with pulmonary
artery hypertension, the expression of 3DL1 is lower and its
function is disrupted, suggesting a central role for KIRs in the
occurrence and development of immune-associated vascular diseases
(59). The decreased frequency of
the KIR3DL1 gene in patients who have miscarried ≥3 times may lead
to the weakened inhibition of NK cells, which contributes to the
imbalance of steady state and works against fetal survival. The
different significance results that were identified between women
who had miscarried ≥3 times and those who had not, suggests that
this should be considered as a selection criteria when miscarriage
diseases are being researched. A previous study also demonstrated
that there was a decreased expression frequency of the uNK KIR2DL1
receptor in women with RSA who carried a C2 epitope (10). In the present study, the high rate of
HLA-C2C2 in KIR AB and KIR BB individuals may decrease the
expression of the KIR2DL1 receptor and simultaneously generate
strong activation effects, which breaks the self-adjustment ability
of NKs and activates NK-mediated immunological rejection of the
fetus.
Komlos et al (60) first demonstrated that couples sharing
common HLA antigens had a significantly higher percentage of
repeated miscarriages compared with the control group. This led to
further research on the subject and certain studies identified that
KIRs were associated with RSA, however, there are disagreements
over the underlying mechanisms. Varla-Leftherioti et al
(61) revealed there was a lower
frequency of inhibitory KIRs (inhibitory 2DL1, 2 and 3) in the
peripheral blood and decidual tissue of women suffering multiple
miscarriages, compared with fertile females. Other studies
identified inhibitory genotypes as more common in the control
subjects and activating genotypes as more common in patients with
RSA (29,30,46,62). The
frequency of the KIR2DS1 gene has been demonstrated to be
significantly higher in patients with RSA compared with healthy
controls (63). However, the
frequency of activating KIR genes demonstrated no significant
differences between the two groups in the present study. The
KIR2DS1 gene combined with a specific ligand may generate a strong
activation effect in NKs (64). The
activation of KIR2DS1+uNKs, stimulated by fetal HLA-C2, may form
soluble products, including granulocyte-macrophage
colony-stimulating factor, which enhances the migration of primary
trophoblast cells (65). A recent
study also indicated that the co-expression of KIR2DS1 and HLA-C2
may be associated with RSA (66). In
the present study, the co-existing frequency of KIR2DS1-C2 genes in
patients with RSA was slightly higher than in the control groups
(20.9 vs. 18.1%). However, the women in the RSA group who had a
homozygous HLA-C2C2 had a significantly higher frequency of the
2DS1 gene compared with the control group (45.5 vs 0.0%;
P<0.001). Furthermore, in women who had aborted ≥3 times this
frequency was at 60.0%. This supports the hypothesis that strong
activation effects surpass the self-regulation of NKs and alter
their secretory activity into cell toxicity, which may ultimately
lead to spontaneous miscarriage.
Previous research reported that the frequency of the
KIR BB genotype was significantly lower and the KIR AB genotype was
significantly higher in the RSA group compared with the control
cohort (29). Additionally,
infertile patients who carry the KIR A haplotype experienced fewer
pregnancy losses following euploid single-embryo transfer compared
with KIR B haplotype carriers (28).
In the Han Chinese population, the KIR AA genotype has a high
prevalence, which may reach up to half of the total number of the
population in some areas (47),
however, no difference was identified in the frequency of the KIR
AA genotype between the two groups in the present study. The rate
of the BB genotype was significantly higher in women who had
aborted ≥3 times compared with the control group. Alternatively,
certain previous studies have suggested that the maternal AA
genotype is associated with an increased risk of pregnancy
complications and the activation of uNK cells is a generic
mechanism promoting trophoblast invasion into the decidua (27,31,32,46,67). The
2DL1 gene in all AA individuals represents the highest inhibitory
influence when interacting with HLA-C2, which is characterized by a
lack of appropriate cytokines to enforce uterine arterial
remodeling, thus leading to poor placentation and ultimately fetal
loss (32,46,58,68). In
the present study, these tendencies were not identified.
In summary, many viewpoints have been published on
how RSA is associated with individual genetics, but the underlying
molecular basis of the disease has not yet been determined. To
date, a variety of therapeutic regimens have been used in the
clinical treatment of RSA, with varying results (6,69,70).
Meta-analyses have concluded that paternal cell immunization,
third-party donor leukocytes, trophoblast membranes and intravenous
immunoglobulin trials did not improve the live birth rate or have a
beneficial effect in women with previous unexplained recurrent
miscarriage (69,71–73). A
full understanding of the underlying pathogenesis of RSA is
required prior to the development of an effective treatment plan,
which may assist patients in completing successful pregnancies. The
results of the present study revealed that in patients with RSA
(miscarried ≥3 times), the frequency of inhibitory genes was
reduced while activated gene combinations increased when compared
with the control group. The self-activation of the maternal NK
cells or the reduction of the activation threshold may result in NK
cells becoming susceptible to irritability, which may be associated
with the occurrence of RSA. The experimental results provide an
insight for further study on miscarriage induced by maternal-fetal
interface immune disorders. In addition, the results may have
clinical therapeutic significance, helping select suitable
fertilized eggs for women who are at high risk of recurrent
spontaneous abortion and reduce the activation of uNKs and the
incidence of pregnancy complications. Comprehensive analysis of
large groups is required to obtain useful results. This is
dependent on concerted efforts from multiple regions due to the
varied nature of KIR genes throughout different ethnicities and
locations.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81501336 and
81450018).
References
|
1
|
Stirrat GM: Recurrent miscarriage. Lancet.
336:673–675. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Evaluation and treatment of recurrent
pregnancy loss: A committee opinion. Fertil Steril. 98:1103–1111.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
van den Boogaard E, Kaandorp SP, Franssen
MT, Mol BW, Leschot NJ, Wouters CH, van der Veen F, Korevaar JC and
Goddijn M: Consecutive or non-consecutive recurrent miscarriage: Is
there any difference in carrier status? Hum Reprod. 25:1411–1414.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moffett A and Loke C: Immunology of
placentation in eutherian mammals. Nat Rev Immunol. 6:584–594.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moffett-King A: Natural killer cells and
pregnancy. Nat Rev Immunol. 2:656–663. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cavalcante MB, Sarno M, Junior E Araujo,
Da Silva Costa F and Barini R: Lymphocyte immunotherapy in the
treatment of recurrent miscarriage: Systematic review and
meta-analysis. Arch Gynecol Obstet. 295:511–518. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ravet S, Scott-Algara D, Bonnet E, Tran
HK, Tran T, Nguyen N, Truong LX, Theodorou I, Barré-Sinoussi F,
Pancino G and Paul P: Distinctive NK-cell receptor repertoires
sustain high-level constitutive NK-cell activation in HIV-exposed
uninfected individuals. Blood. 109:4296–4305. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Koopman LA, Kopcow HD, Rybalov B, Boyson
JE, Orange JS, Schatz F, Masch R, Lockwood CJ, Schachter AD, Park
PJ and Strominger JL: Human decidual natural killer cells are a
unique NK cell subset with immunomodulatory potential. J Exp Med.
198:1201–1212. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
King A, Burrows T, Verma S, Hiby S and
Loke YW: Human uterine lymphocytes. Hum Reprod Update. 4:480–485.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sharkey AM, Xiong S, Kennedy PR, Gardner
L, Farrell LE, Chazara O, Ivarsson MA, Hiby SE, Colucci F and
Moffett A: Tissue-specific education of decidual NK cells. J
Immunol. 195:3026–3032. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi FD, Ljunggren HG, La Cava A and Van
Kaer L: Organ-specific features of natural killer cells. Nat Rev
Immunol. 11:658–671. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ferlazzo G, Tsang ML, Moretta L, Melioli
G, Steinman RM and Münz C: Human dendritic cells activate resting
natural killer (NK) cells and are recognized via the NKp30 receptor
by activated NK cells. J Exp Med. 195:343–351. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lash GE, Naruse K, Robson A, Innes BA,
Searle RF, Robson SC and Bulmer JN: Interaction between uterine
natural killer cells and extravillous trophoblast cells: Effect on
cytokine and angiogenic growth factor production. Hum Reprod.
26:2289–2295. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van der Meer A, Lukassen HG, van Lierop
MJ, Wijnands F, Mosselman S, Braat DD and Joosten I: Membrane-bound
HLA-G activates proliferation and interferon-gamma production by
uterine natural killer cells. Mol Hum Reprod. 10:189–195. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Raulet DH and Vance RE: Self-tolerance of
natural killer cells. Nat Rev Immunol. 6:520–531. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lanier LL: NK cell recognition. Annu Rev
Immunol. 23:225–274. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Uhrberg M, Valiante NM, Shum BP, Shilling
HG, Lienert-Weidenbach K, Corliss B, Tyan D, Lanier LL and Parham
P: Human diversity in killer cell inhibitory receptor genes.
Immunity. 7:753–763. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Varla-Leftherioti M, Spyropoulou-Vlachou
M, Niokou D, Keramitsoglou T, Darlamitsou A, Tsekoura C,
Papadimitropoulos M, Lepage V, Balafoutas C and Stavropoulos-Giokas
C: Natural killer (NK) cell receptors' repertoire in couples with
recurrent spontaneous abortions. Am J Reprod Immunol. 49:183–191.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vivier E, Nunès JA and Vély F: Natural
killer cell signaling pathways. Science. 306:1517–1519. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yokoyama WM and Plougastel BF: Immune
functions encoded by the natural killer gene complex. Nat Rev
Immunol. 3:304–316. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vilches C and Parham P: KIR: Diverse,
rapidly evolving receptors of innate and adaptive immunity. Annu
Rev Immunol. 20:217–251. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Uhrberg M, Parham P and Wernet P:
Definition of gene content for nine common group B haplotypes of
the Caucasoid population: KIR haplotypes contain between seven and
eleven KIR genes. Immunogenetics. 54:221–229. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pyo CW, Guethlein LA, Vu Q, Wang R,
Abi-Rached L, Norman PJ, Marsh SG, Miller JS, Parham P and Geraghty
DE: Different patterns of evolution in the centromeric and
telomeric regions of group A and B haplotypes of the human killer
cell Ig-like receptor locus. PloS one. 5:e151152010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alter G, Heckerman D, Schneidewind A,
Fadda L, Kadie CM, Carlson JM, Oniangue-Ndza C, Martin M, Li B,
Khakoo SI, et al: HIV-1 adaptation to NK-cell-mediated immune
pressure. Nature. 476:96–100. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
de Smith AJ, Walsh KM, Ladner MB, Zhang S,
Xiao C, Cohen F, Moore TB, Chokkalingam AP, Metayer C, Buffler PA,
et al: The role of KIR genes and their cognate HLA class I ligands
in childhood acute lymphoblastic leukemia. Blood. 123:2497–2503.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cooley S, Weisdorf DJ, Guethlein LA, Klein
JP, Wang T, Marsh SG, Spellman S, Haagenson MD, Saeturn K, Ladner
M, et al: Donor killer cell Ig-like receptor B haplotypes,
recipient HLA-C1 and HLA-C mismatch enhance the clinical benefit of
unrelated transplantation for acute myelogenous leukemia. J
Immunol. 192:4592–4600. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakimuli A, Chazara O, Hiby SE, Farrell L,
Tukwasibwe S, Jayaraman J, Traherne JA, Trowsdale J, Colucci F,
Lougee E, et al: A KIR B centromeric region present in Africans but
not Europeans protects pregnant women from pre-eclampsia. Proc Natl
Acad Sci USA. 112:pp. 845–850. 2015; View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Morin SJ, Treff NR, Tao X, Scott RT III,
Franasiak JM, Juneau CR, Maguire M and Scott RT: Combination of
uterine natural killer cell immunoglobulin receptor haplotype and
trophoblastic HLA-C ligand influences the risk of pregnancy loss: A
retrospective cohort analysis of direct embryo genotyping data from
euploid transfers. Fertil Steril. 107:677–683. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ozturk OG, Sahin G, Karacor ED and
Kucukgoz U: Evaluation of KIR genes in recurrent miscarriage. J
Assist Reprod Genet. 29:933–938. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vargas RG, Bompeixe EP, Franca PP, de
Moraes M Marques and da Graca Bicalho M: Activating killer cell
immunoglobulin-like receptor genes' association with recurrent
miscarriage. Am J Reprod Immunol. 62:34–43. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Alecsandru D, Garrido N, Vicario JL,
Barrio A, Aparicio P, Requena A and García-Velasco JA: Maternal KIR
haplotype influences live birth rate after double embryo transfer
in IVF cycles in patients with recurrent miscarriages and
implantation failure. Hum Reprod. 29:2637–2643. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hiby SE, Apps R, Sharkey AM, Farrell LE,
Gardner L, Mulder A, Claas FH, Walker JJ, Redman CW, Morgan L, et
al: Maternal activating KIRs protect against human reproductive
failure mediated by fetal HLA-C2. J Clin Invest. 120:4102–4110.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Toneva M, Lepage V, Lafay G, Dulphy N,
Busson M, Lester S, Vu-Trien A, Michaylova A, Naumova E, McCluskey
J and Charron D: Genomic diversity of natural killer cell receptor
genes in three populations. Tissue Antigens. 57:358–362. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yao Y, Shi L, Tao Y, Lin K, Liu S, Yu L,
Yang Z, Yi W, Huang X, Sun H, et al: Diversity of killer cell
immunoglobulin-like receptor genes in four ethnic groups in China.
Immunogenetics. 63:475–483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu H, Shan Z, Li C, Mao J, Xie X, Wang W,
Fan C, Wang H, Zhang H, Han C, et al: Maternal subclinical
hypothyroidism, thyroid autoimmunity, and the risk of miscarriage:
A prospective cohort study. Thyroid. 24:1642–1649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kishore J, Agarwal J, Agrawal S and
Ayyagari A: Seroanalysis of Chlamydia trachomatis and S-TORCH
agents in women with recurrent spontaneous abortions. Indian J
Pathol Microbiol. 46:684–687. 2003.PubMed/NCBI
|
|
37
|
Simcox LE, Ormesher L, Tower C and Greer
IA: Thrombophilia and pregnancy complications. Int J Mol Sci.
16:28418–28428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tajik N, Shahsavar F, Nasiri M and
Radjabzadeh MF: Compound KIR-HLA genotype analyses in the Iranian
population by a novel PCR-SSP assay. Int J Immunogenet. 37:159–168.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schaap T: The applicability of the
Hardy-Weinberg principle in the study of populations. Ann Hum
Genet. 44:211–215. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Martin AM, Freitas EM, Witt CS and
Christiansen FT: The genomic organization and evolution of the
natural killer immunoglobulin-like receptor (KIR) gene cluster.
Immunogenetics. 51:268–280. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Meuleman T, Cohen D, Swings GM, Veraar K,
Claas FH and Bloemenkamp KW: Increased complement C4d deposition at
the maternal-fetal interface in unexplained recurrent miscarriage.
J Reprod Immunol. 113:54–60. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dahl M and Hviid TV: Human leucocyte
antigen class Ib molecules in pregnancy success and early pregnancy
loss. Hum Reprod Update. 18:92–109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
King A, Allan DS, Bowen M, Powis SJ,
Joseph S, Verma S, Hiby SE, McMichael AJ, Loke YW and Braud VM:
HLA-E is expressed on trophoblast and interacts with CD94/NKG2
receptors on decidual NK cells. Eur J Immunol. 30:1623–1631. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Apps R, Gardner L, Sharkey AM, Holmes N
and Moffett A: A homodimeric complex of HLA-G on normal trophoblast
cells modulates antigen-presenting cells via LILRB1. Eur J Immunol.
37:1924–1937. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Brosens I, Pijnenborg R, Vercruysse L and
Romero R: The ‘Great Obstetrical Syndromes’ are associated with
disorders of deep placentation. Am J Obstet Gynecol. 204:193–201.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hiby SE, Walker JJ, O'Shaughnessy KM,
Redman CW, Carrington M, Trowsdale J and Moffett A: Combinations of
maternal KIR and fetal HLA-C genes influence the risk of
preeclampsia and reproductive success. J Exp Med. 200:957–965.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yin C, Hu L, Huang H, Yu Y, Li Z, Ji Q,
Kong X, Wang Z, Yan J, Yan J, et al: Genetic polymorphism and
evolutionary differentiation of Eastern Chinese Han: A
comprehensive and comparative analysis on KIRs. Sci Rep.
7:424862017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Orlando J and Coulam C: Is superfertility
associated with recurrent pregnancy loss? Am J Reprod Immunol.
72:549–554. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Raulet DH, Vance RE and McMahon CW:
Regulation of the natural killer cell receptor repertoire. Annu Rev
Immunol. 19:291–330. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Emmer PM, Steegers EA, Kerstens HM, Bulten
J, Nelen WL, Boer K and Joosten I: Altered phenotype of HLA-G
expressing trophoblast and decidual natural killer cells in
pathological pregnancies. Hum Reprod. 17:1072–1080. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tripathy SK, Keyel PA, Yang L, Pingel JT,
Cheng TP, Schneeberger A and Yokoyama WM: Continuous engagement of
a self-specific activation receptor induces NK cell tolerance. J
Exp Med. 205:1829–1841. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pittari G, Liu XR, Selvakumar A, Zhao Z,
Merino E, Huse M, Chewning JH, Hsu KC and Dupont B: NK cell
tolerance of self-specific activating receptor KIR2DS1 in
individuals with cognate HLA-C2 ligand. J Immunol. 190:4650–4660.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shifrin N, Raulet DH and Ardolino M: NK
cell self tolerance, responsiveness and missing self recognition.
Semin Immunol. 26:138–144. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jiang K, Zhu FM, Lv QF and Yan LX:
Distribution of killer cell immunoglobulin-like receptor genes in
the Chinese Han population. Tissue Antigens. 65:556–563. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu GQ, Zhao YM, Lai XY, Yang KL, Zhu FM,
Zhang W, Wang W, Luo Y, Tan YM, Wang YJ, et al: Distribution of
killer-cell immunoglobulin-like receptor genes in eastern mainland
Chinese han and taiwanese han populations. Tissue antigens.
74:499–507. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Christiansen OB, Mohapeloa HP, Steffensen
R and Jersild C: HLA-C and -Bw typing of couples with unexplained
recurrent miscarriages. J Reprod Immunol. 37:63–77. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ahn RS, Moslehi H, Martin MP, Abad-Santos
M, Bowcock AM, Carrington M and Liao W: Inhibitory KIR3DL1 alleles
are associated with psoriasis. Br J Dermatol. 174:449–451. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kennedy PR, Chazara O, Gardner L, Ivarsson
MA, Farrell LE, Xiong S, Hiby SE, Colucci F, Sharkey AM and Moffett
A: Activating KIR2DS4 is expressed by uterine NK cells and
contributes to successful pregnancy. J Immunol. 197:4292–4300.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ormiston ML, Chang C, Long LL, Soon E,
Jones D, Machado R, Treacy C, Toshner MR, Campbell K, Riding A, et
al: Impaired natural killer cell phenotype and function in
idiopathic and heritable pulmonary arterial hypertension.
Circulation. 126:1099–1109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Komlos L, Zamir R, Joshua H and Halbrecht
I: Common HLA antigens in couples with repeated abortions. Clin
Immunol Immunopathol. 7:330–335. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Varla-Leftherioti M, Spyropoulou-Vlachou
M, Keramitsoglou T, Papadimitropoulos M, Tsekoura C, Graphou O,
Papadopoulou C, Gerondi M and Stavropoulos-Giokas C: Lack of the
appropriate natural killer cell inhibitory receptors in women with
spontaneous abortion. Hum Immunol. 66:65–71. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Faridi RM, Das V, Tripthi G, Talwar S,
Parveen F and Agrawal S: Influence of activating and inhibitory
killer immunoglobulin-like receptors on predisposition to recurrent
miscarriages. Hum Reprod. 24:1758–1764. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang S, Zhao YR, Jiao YL, Wang LC, Li JF,
Cui B, Xu CY, Shi YH and Chen ZJ: Increased activating killer
immunoglobulin-like receptor genes and decreased specific HLA-C
alleles in couples with recurrent spontaneous abortion. Biochem
Biophys Res Commun. 360:696–701. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Sivori S, Carlomagno S, Falco M, Romeo E,
Moretta L and Moretta A: Natural killer cells expressing the
KIR2DS1-activating receptor efficiently kill T-cell blasts and
dendritic cells: Implications in haploidentical HSCT. Blood.
117:4284–4292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Xiong S, Sharkey AM, Kennedy PR, Gardner
L, Farrell LE, Chazara O, Bauer J, Hiby SE, Colucci F and Moffett
A: Maternal uterine NK cell-activating receptor KIR2DS1 enhances
placentation. J Clin Invest. 123:4264–4272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Dambaeva SV, Lee DH, Sung N, Chen CY, Bao
S, Gilman-Sachs A, Kwak-Kim J and Beaman KD: Recurrent pregnancy
loss in women with killer cell immunoglobulin-like receptor KIR2DS1
is associated with an increased HLA-C2 allelic frequency. Am J
Reprod Immunol. 75:94–103. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hiby SE, Regan L, Lo W, Farrell L,
Carrington M and Moffett A: Association of maternal killer-cell
immunoglobulin-like receptors and parental HLA-C genotypes with
recurrent miscarriage. Hum Reprod. 23:972–976. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Flores AC, Marcos CY, Paladino N, Arruvito
L, Williams F, Middleton D and Fainboim L: KIR receptors and HLA-C
in the maintenance of pregnancy. Tissue Antigens. 1(69 Suppl):
S112–S113. 2007. View Article : Google Scholar
|
|
69
|
Porter TF, Wong LF and Scott JR:
Immunotherapy for recurrent miscarriage. Cochrane Database Syst
Rev. 21:CD0001122006.
|
|
70
|
Chen JL, Yang JM, Huang YZ and Li Y:
Clinical observation of lymphocyte active immunotherapy in 380
patients with unexplained recurrent spontaneous abortion. Int
immunopharmacol. 40:347–350. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hutton B, Sharma R, Fergusson D, Tinmouth
A, Hebert P, Jamieson J and Walker M: Use of intravenous
immunoglobulin for treatment of recurrent miscarriage: A systematic
review. BJOG. 114:134–142. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ata B, Tan SL, Shehata F, Holzer H and
Buckett W: A systematic review of intravenous immunoglobulin for
treatment of unexplained recurrent miscarriage. Fertil Steril.
95:1080–1085. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu Z, Xu H, Kang X, Wang T, He L and Zhao
A: Allogenic lymphocyte immunotherapy for unexplained recurrent
spontaneous abortion: A meta-analysis. Am J Reprod Immunol.
76:443–453. 2016. View Article : Google Scholar : PubMed/NCBI
|