Introduction
Ulcerative colitis (UC), a chronic intestinal
inflammation condition, is characterized by repeated flare-ups of
inflammation that can result in oncogenic insults to the colonic
epithelial. UC is a known risk factor for intestinal cancer,
especially for colorectal cancer. The etiology of UC is unknown but
it is generally considered to be the result of a combination of
genetic susceptibility, changes in the commensal enteric flora, and
an immune imbalance, in which the inflammatory response is
considered to be one of the important pathogenic factors (1–3). Markers
of inflammation, TNF-α (tumor necrosis factor-α) and IL-6
(interleukin-6), are useful in assessing disease activity in
patients with UC, as they have been demonstrated to contribute to
the development of colon mucosal damage and lead to a chronic
inflammatory process (4).
LPS (lipopolysaccharide), the specialized structural
component of bacteria cell walls, represents a major virulence
factor of gram-negative bacteria, as it initiates the occurrence
and development of a strong inflammatory response (5). Oral administration of dextran sodium
sulphate (DSS) in mice triggers intestinal inflammation that is
closely related to human ulcerative colitis (6).
Dandelion is a perennial herb which is used to treat
acute conjunctivitis, malignant boils, skin ulcers and other
inflammatory conditions (7).
Taraxasterol is an active compound extracted from dandelion that
has a molecular structure resembling a steroid hormone, and has
been demonstrated to display anti-inflammatory effects in
vivo and in vitro (8).
However, there are no reports on the treatment of UC using
taraxasterol. In the present study, taraxasterol was demonstrated
to significantly decrease expression of inflammatory factors IL-6
and TNF-α, in vivo and vitro. In addition,
taraxasterol significantly decreased the apoptosis rate, as well as
the protein expression levels of caspase-3 and p53, which suggested
that the inhibitory effects of taraxasterol on inflammatory factors
may have an effect on apoptosis. Finally, taraxasterol decreased
the pathological damage of colonic epithelial cells in a
dose-dependent manner. The present study provided the foundation
for further investigation into the anti-inflammatory mechanism of
taraxasterol.
Materials and methods
Drugs and reagents
Taraxasterol (purity, 98% HPLC) was purchased from
Chengdu Herbpurify Co., Ltd. (Chengdu, China). Dextran sodium
sulfate (DSS) was purchased from MP Biomedicals, LLC (Santa Ana,
CA, USA). The ELISA kits for TNF-α (cat. no. A0119), IL-6 (cat. no.
A0107) were purchased from Nanjing Jiancheng Bioengineering
Institute (China). All plastic materials were purchased from Falcon
Labware (Becton, Dickinson and Company, Franklin Lakes, NJ, USA).
RPMI Medium-1640, fetal bovine serum (FBS), and phosphate buffer
saline (PBS) were obtained from Gibco (Thermo Fisher Scientific,
Inc. Waltham, MA, USA). Penicillin G/streptomycin, MTT, and
dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (Merck
KGaA Darmstadt, Germany). Apoptosis Assay kit was purchased from
Nanjing KeyGen Biological Co., Ltd. (Nanjing, China). Rabbit
anti-human Caspase antibody (cat no. 9661; 1:1,000; Cell Signaling
Technology, Inc.), rabbit anti-human BAX antibody (cat no. 2774;
1:1,000; Cell Signaling Technology, Inc.), rabbit anti-human p53
antibody (cat no. 48818; 1:1,000; Cell Signaling Technology, Inc.)
and rabbit anti-human β-actin antibody (cat no. 622101; 1:2,000)
were purchased from BioLegend Inc. (San Diego, CA, USA). Human
colon cancer epithelial cells (HT-29) were purchased from the
Chinese Academy of Sciences Shanghai Cell Bank (Shanghai,
China).
Experimental animals
Adult male C57BL/6 mice (20–24 g) were purchased
from the Laboratory Animal Services Center, The First Affiliated
Hospital of Zhengzhou University (Zhengzhou, China). All animals
were housed in accordance with the National Institutes of Health
Guide for Laboratory animals' use. The study was approved by the
Animal Ethics Committee of The First Affiliated Hospital of
Zhengzhou University. Animals were housed under standard
environment condition of temperature at 20–25°C under a 12-h
dark/light cycle and allowed free access to sterilized water and
standard food.
Colitis mouse model and related
treatments
A total of 60 male C57BL/6 mice were randomly
divided into 5 groups (n=12); control group, model group (DSS
group), low, medium and high dose (25, 50 and 100 mg/kg/day
respectively) taraxasterol groups. Colonic inflammation was
established by oral gavage of 5% DSS solution for 5 days.
Establishment of a successful model was determined based on thin
stools, fecal blood, weight loss and other symptoms and
morphological changes. Intestinal inflammation was graded in a
blinded fashion, according to Cooper et al (9): 0, no inflammation; 1, mild inflammation
of the mucosal intrinsic layer and submucosal layer; 2, severe
inflammation of the mucosal intrinsic layer and submucosal layer;
3, mild inflammation of the whole colon wall; and 4, severe
inflammation of the whole colon wall.
For taraxasterol treatments, C57BL/6 mice were fed
with low, medium or high dose (25, 50 and 100 mg/kg/day)
taraxasterol. Mice in the control group were treated with normal
saline.
Colon histomorphology
Changes in mucosa and serosal surfaces of colonic
tissue were observed via microscope. Fresh colonic tissue was
isolated (size ~1×1×0.2 cm). The embedding box was fixed in 4%
paraformaldehyde for 24 h. The tissue was embedded in paraffin and
stained continuously for 4 µm. Tissue lesions were observed via
microscopic observation through hematoxylin and eosin (HE)
staining. The criteria was as follows: 0, normal colonic mucosa; 1,
crypt defect of 1/3; 2, crypt defect of 2/3; 3, the inherent layer
covers the monolayer epithelium with mild inflammatory cell
infiltration; and 4, erosion and ulcer in mucosa with plentiful
inflammatory cell infiltration.
Immunohistochemistry (IHC)
Paraffin-embedded colonic tissues were continuously
sliced into 4 µm thick sections, at 65°C for 2 h. The
streptavidin-peroxidase (SP) method was used where sections were
dewaxed, hydrated then 0.01 M sodium citrate buffer solution was
used for antigen repair for 15 min (1:300) at 95°C. Sections were
washed and blocked with goat serum (cat. no. ab7481; Abcam) at room
temperature for 20 min. Samples were incubated with primary
antibody anti-p53 (1:300; cat no. 48818; Cell Signaling Technology,
Inc.) at 4°C overnight. Sections were then washed three times for 5
min using PBS with Tween (PBS-T) then incubated with horseradish
peroxidase linked secondary antibody (1:2,000; cat no. 7076; Cell
Signaling Technology, Inc.) at 37°C for 1 h. Following washes, SP
was added at room temperature for 30 min. DAB staining upon the
observation of the slices, and finally the slices were dehydrated,
transparent and sealed. Images were taken using an image scanner
(Zeiss 125 AxioScan.Z1; Zeiss AG).
A total of 10 non-repeatable fields were selected at
×100 magnification to analyze the % of positively stained cells and
coloring intensity. The intensity was scored as: 0, negative; 1,
weak; 2, medium; 3, strong. The % of positive-stained cancer cells
was scored as: 0, 0–5%; 1, 6–25%; 2, 26–50%; 3, 51–75%; and 4,
76–100%. The intensity number multiplied by extent of staining
number produced the final staining score of p53 (range 0–12).
Cell culture
Human colon cancer epithelium HT-29 cells were
removed from liquid nitrogen, thawed in a water bath at 37°C, then
transferred into a centrifuge tube with RPMI 1640 medium containing
10% fetal bovine serum, and centrifuged at 200 × g for 10 min. The
cell pellet was resuspended with complete medium in a 75 ml
standard flask, then incubated under standard conditions (37°C, 5%
CO2). For LPS and taraxasterol treatments, either LPS
(10 µg/ml) or taraxasterol (2.5, 5 or 10 µg/ml respectively) were
added into the medium.
MTT assay
In the logarithmic growth phase, 5,000 cells/well
were seeded in 5 wells of a 96-well plate. Following 12 h, 1640
medium with 1% serum containing no taraxasterol was added to the
control group and culture medium 1640 with 1% serum containing 2.5,
5 or 10 µg/ml taraxasterol was added to the model group. A total of
100 µl of medium containing MTT (0.5 g/l) was added to each well
then samples were incubated for 4 h under standard conditions
(37°C, 5% CO2). The purple formazan crystals were
dissolved using dimethyl sulfoxide and viability was subsequently
analyzed at a wavelength of 490 nm with a microplate reader
(BIO-RAD680; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Measurement of cytokines
HT-29 cells were treated with different
concentrations of taraxasterol (2.5, 5 or 10 µg/ml) for 24 h. After
digestion, cells were collected and centrifuged at 800 × g for 5
min then the supernatant was collected. For animal experiments,
colon tissue was cut into 2–3 mm samples, homogenized with PBS,
centrifuged at 12,000 × g for 15 min then the supernatant was
collected. Levels of TNF-α and IL-6 in the supernatant were
measured according to the instructions of the kit. The experiment
was repeated at least three times.
Apoptosis analysis in vitro
HT-29 cells were treated with different
concentrations of taraxasterol (2.5, 5 or 10 µg/ml) for 24 h. Cells
were washed with PBS twice, digested and collected by
centrifugation. Cells were resuspended with 500 µl binding-buffer,
followed by 5 µl Annexin-V/fluorescein isothiocyanate (FITC) and 5
µl propidium iodide (PI) for 5 min. Cells were detected by flow
cytometry (BD FACSCanto A) and analyzed using FlowJo v10 (FlowJo
LLC). FITC-/PI- were deemed viable cells, FITC-/PI+ were necrotic
cells, FITC+/PI- were early apoptotic cells, and FITC+/PI+ were
late apoptotic cells. The apoptosis rate was calculated as follows:
Apoptosis rate % = (number of early apoptotic cells + number of
late apoptotic cells)/total number of cells ×100%. The experiment
was repeated at least three times.
Western blot analysis
Proteins were extracted from cell suspension and
tissue homogenates with radioimmunoprecipitation assay buffer
(Beyotime Institute of Biotechnology). The concentration of
proteins were detected via Bicinchoninic acid Protein Assay Kit.
Proteins (30 µg) were separated via for SDS-PAGE on a 12% gel for 2
h, then the proteins were transferred for 2 h to a polyvinylidene
difluoride membrane. Blocking of the membrane with 5% skim milk was
performed at 37°C for 2 h. The membranes were incubated with
primary antibodies anti-p53 (cat no. 48818; 1:1,000; Cell Signaling
Technology, Inc.), BAX (cat no. 2774; 1:1,000; Cell Signaling
Technology, Inc.) and cleaved caspase-3 (cat no. 9661; 1:1,000;
Cell Signaling Technology, Inc.) overnight at 4°C. Membranes were
washed with PBS for 15 min then incubated with secondary antibody
IgG (cat on. 7076; 1:2,000; Cell Signaling Technology, Inc.) for
1.5 h at room temperature. Electrochemiluminescence imaging with
gel imaging analysis system (Bio-Rad Laboratories, Inc.) was used
to measure protein band density value. The experiment was repeated
at least three times with β-actin as the loading control.
Statistical analysis
Statistical analysis was performed using SPSS
software (version 18.0; SPSS, Inc., Chicago, IL, USA). All results
were presented as mean ± standard error of the mean (SEM).
Comparisons were performed using one-way analysis of variance
followed by Bonferroni's post hoc test. P<0.05 was considered to
indicate significant difference.
Results
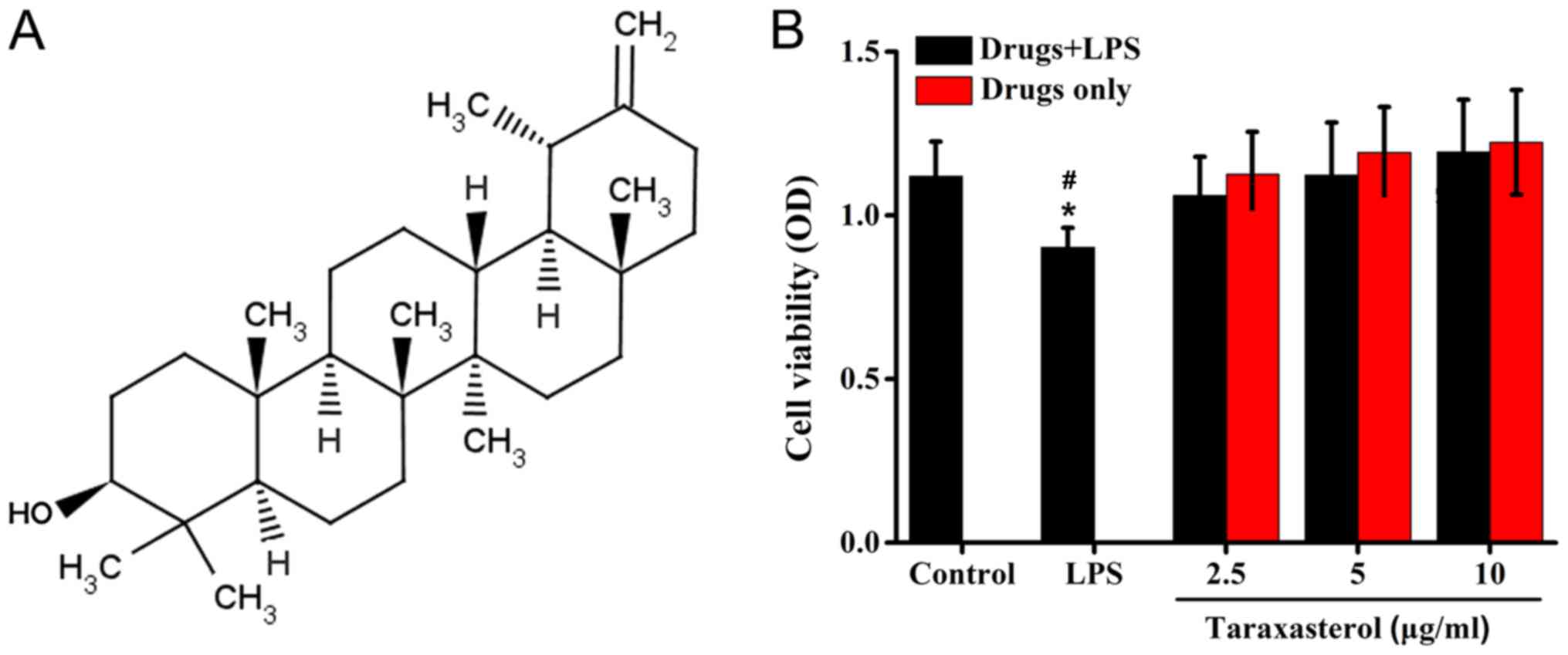
Effect of taraxasterol and LPS on
HT-29 cell toxicity
To obtain the optimal concentration of taraxasterol
and LPS, HT-29 cells were treated with LPS or taraxasterol at
different concentrations, and the cell viability was detected via
MTT assay. The results demonstrated that 10 µg/ml LPS decreased the
cell viability slightly, while the taraxasterol concentrations used
in the experiments did not affect the survival of HT-29 cells
compared with the control group (Fig.
1).
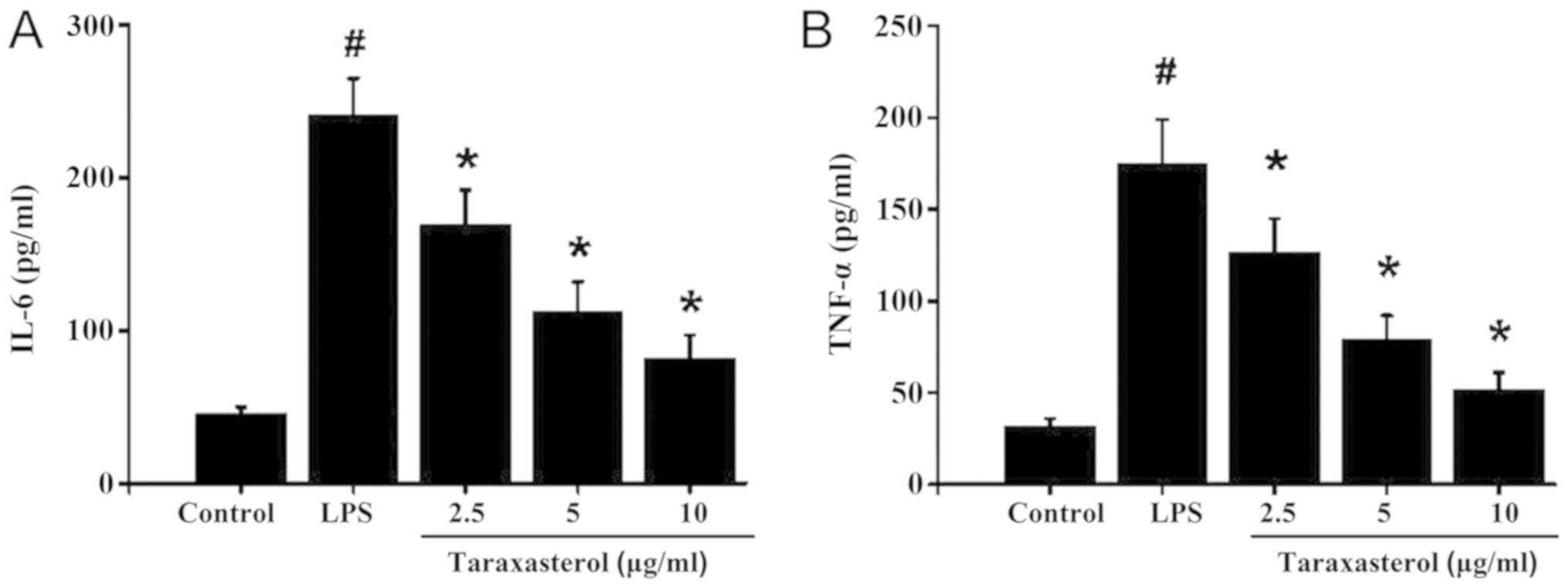
Effect of taraxasterol on expression
of inflammatory factors TNF-α and IL-6 in vitro
Next, the effect of taraxasterol on the expression
of inflammation factors was investigated. To this effect, HT-29
cells were treated with LPS to induce an inflammatory response. LPS
stimulation significantly increased TNF-α and IL-6 levels in the
supernatant (Fig. 2A and B).
Taraxasterol treatment decreased the LPS-induced TNF-α and IL-6
expression in a dose-dependent manner which suggested that
taraxasterol exerted a significant anti-inflammatory effect.
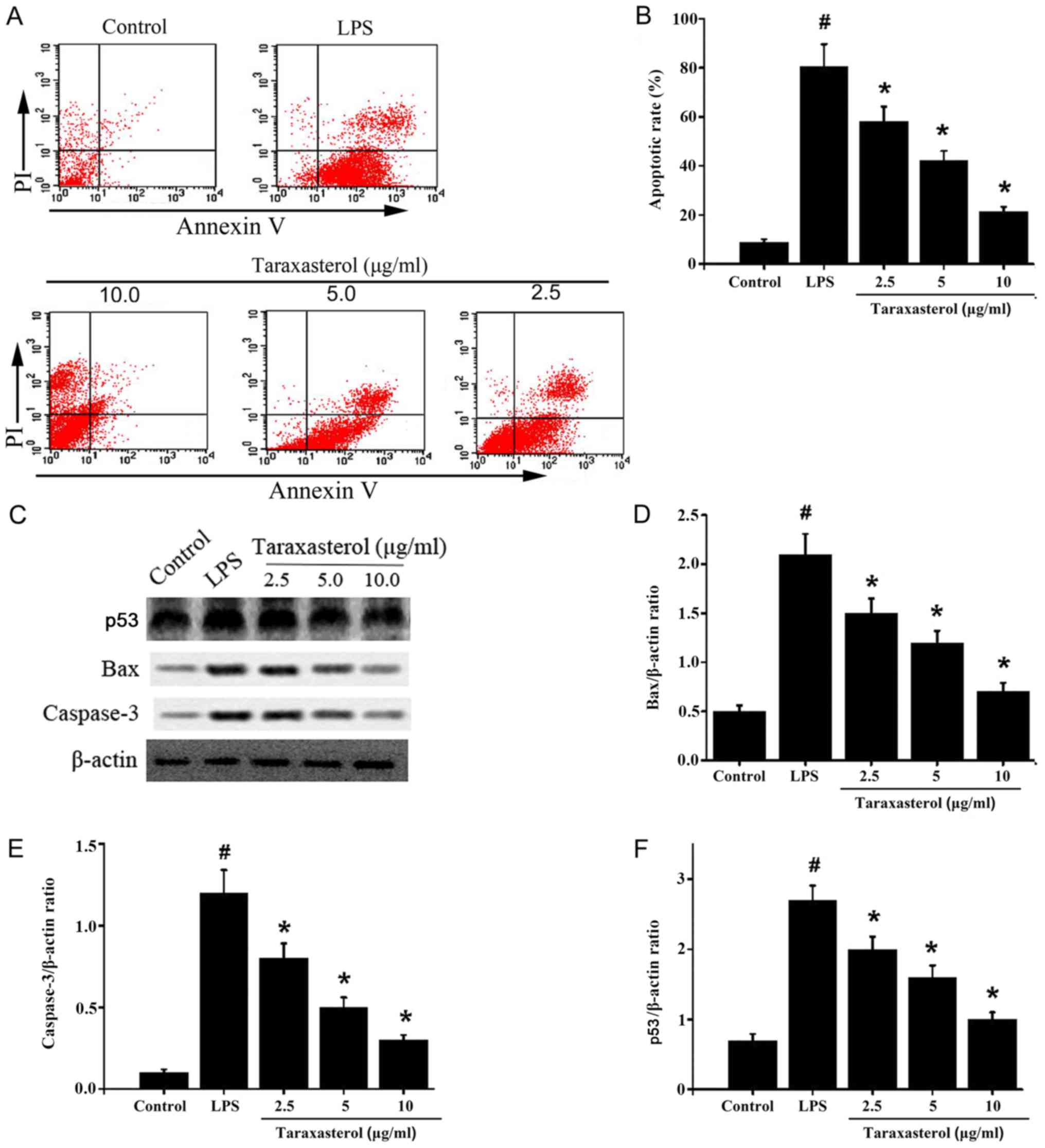
Effect of taraxasterol on LPS-induced
apoptosis
Since taraxasterol could reduce the expression of
inflammatory factors, the effect of taraxasterol on apoptosis
activity was investigated. HT-29 cells were treated with LPS or
taraxasterol, flow cytometry was used to detect the apoptosis
rates. The results demonstrated that LPS induced apoptosis in HT-29
cells. In comparison, taraxasterol significantly decreased
apoptosis in HT-29 cells (Fig 3A and
B). Next, the expression levels of apoptosis-related proteins
were detected via western blotting. The results demonstrated that
expression of p53, BAX and cleaved caspase-3 were significantly
higher in the LPS-treated group compared with the control group
(Fig. 3C-F). Taraxasterol
significantly decreased p53, BAX and cleaved caspase-3 protein
levels in a dose dependent manner (Fig.
3C-F), which suggests that taraxasterol could reduce
LPS-induced apoptosis in vitro.
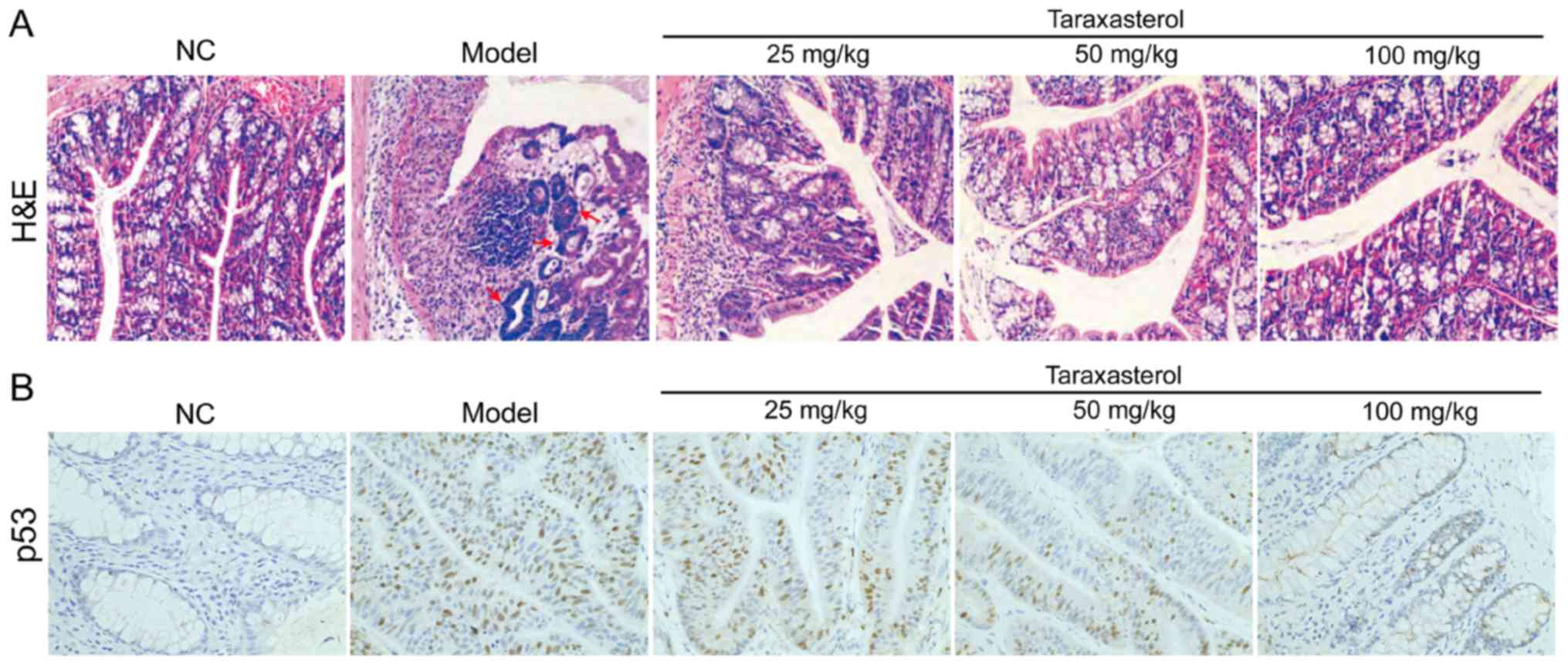
Effect of taraxasterol on colon
histopathological changes induced by DSS
Animal experiments were used to identify the effects
of taraxasterol on UC in vivo, after DSS was used to
establish a UC mouse model. The structure of the control group
colonic tissue was distinct with smooth intact mucosa and no
erosion of the tissue or inflammatory cell infiltration (Fig. 4A). In the UC model group, mucosal
erosion, granulation tissue hyperplasia and glandular enlargement
was observed in Fig. 4A (indicated
by the red arrows). In the taraxasterol groups, there was no
obvious erosion in the colonic mucosa of mice with minimal
inflammatory cell infiltration (Fig.
4A). Colonic pathological changes in the 100 mg/kg taraxasterol
dosage group were significantly reduced compared with the 50 mg/kg
or 25 mg/kg taraxasterol treatment groups (Fig. 4A). The histopathological scores of
the model group and taraxasterol treated groups were significantly
higher compared with the control group (Table I; P<0.05). The histopathological
score of the 100 mg/kg group (1.5±0.2) was significantly lower
compared with the 25 mg/kg group (2.7±0.3; P<0.05).
 | Table I.Effect of taraxasterol on
histopathological score, p53 IHC score and expression of TNF-α and
IL-6 in vivo. |
Table I.
Effect of taraxasterol on
histopathological score, p53 IHC score and expression of TNF-α and
IL-6 in vivo.
| Group | n | Histopathological
score | p53 IHC score | IL-6 (pg/ml) | TNF (pg/ml) |
|---|
| Model | 12 | 9.12±1.12 | 16.27±2.12 | 271.36±24.28 | 342.182±24.283 |
| 25 mg/kg | 12 | 7.43±1.02 | 13.28±1.87 |
215.37±24.28c |
254.281±24.283c |
| 50 mg/kg | 12 | 5.38±0.93 | 10.27±1.68 |
162.27±24.28b,c |
182.472±24.283b,c |
| 100 mg/kg | 12 | 4.58±0.98 | 9.12±1.57 |
135.21±24.28a–c |
152.811±16.482a–c |
| Control | 12 | 1.95±0.47 | 5.27±1.20 |
69.26±11.45a |
83.127±9.281a |
Effect of taraxasterol on p53 protein
expression in colon tissues
Immunohistochemical assay was used to determine p53
protein levels in colon tissues. The results demonstrated that
positive p53 staining (brown) was mainly located within nuclei
(Fig. 4B). Expression of p53 in
colonic mucosa was higher in the UC model groups and the
taraxasterol treated groups compared to the control groups
(Table I; P<0.01). Treatment with
100 mg/kg taraxasterol significantly decreased p53 levels compared
with the UC model group (Fig. 4B).
There was no significant difference in IHC score between 100 mg/kg
and 50 mg/kg taraxasterol treatment groups, however both had lower
IHC scores than the 25 mg/kg treatment groups (Table I; P<0.01).
Effect of taraxasterol on the
expression of TNF-a and IL-6 in vivo
The expression levels of TNF-α and IL-6 in the
supernatant of colonic mucosa tissues was determined. TNF-α and
IL-6 expression levels were lower in taraxasterol-treated groups
compared with UC model groups (Table
I; P<0.01). The levels of TNF-α and IL-6 in the
taraxasterol-treated groups were significantly higher compared with
the control group (Table I;
P<0.05). In taraxasterol-treated groups, the levels of TNF-α and
IL-6 in the 100 mg/kg treatment group were significantly lower than
those in the 25 mg/kg treatment group (Table I; P<0.05). These results suggested
that taraxasterol significantly decreased the expression of TNF-α
and IL-6 in vivo.
Discussion
Cytokines are immunomodulators that are released in
response to injury or inflammation with inflammatory cytokines
serving an important role in the development of UC. The present
study investigated the anti-inflammatory and anti-apoptosis
activity of taraxasterol in relation to UC. Taraxasterol is an
active compound extracted from dandelion with a molecular structure
resembling a steroid hormone and has been demonstrated to have
anti-inflammatory effects in vivo and in vitro
(8). LPS is the specialized
structural component of bacteria cell walls, a major virulence
factor of gram-negative bacteria (10) and can induce production of a variety
of proinflammatory cytokines, including TNF-α, IL-1 and IL-6
(11). TNF-α is a prototypical
proinflammatory cytokine that has been implicated in carcinogenesis
due to its participation in chronic inflammatory diseases (12). IL-6 is another proinflammatory
cytokine with a typical protumorigenic effect, with elevated IL-6
serum levels detected in patients with systemic cancers (13). Previous studies have reported that
TNF-α and IL-6 are closely associated with UC (14).
In the present study, first, the optimal
concentrations of LPS and taraxasterol on HT-29 cells were
determined, and the results demonstrated that the different
concentrations of taraxasterol and LPS examined had no obvious
cytotoxic effects on HT-29 cells. The levels of TNF-α and IL-6 were
increased both in the in vitro UC model induced by LPS and
in the in vivo UC model induced by DSS. Of note,
taraxasterol significantly decreased the inflammatory cytokines
TNF-α and IL-6 secretion in vivo and in vitro. These
results revealed that taraxasterol could reduce inflammation
through inhibiting TNF-α and IL-6 secretion.
Apoptosis is programmed cell death and maintains the
balance between healthy cell survival and death. Numerous studies
have identified that apoptosis in intestinal epithelial cells is
driven by increased cytokine activity such as TNF, IL, and
interferon family members (15).
Increased apoptosis of intestinal epithelial cells has been
observed at acute UC inflammatory sites which can disrupt
intestinal mucosal integrity and barrier function eventually
leading to inflammation (16).
Identifying how best to decrease colonic epithelial cell apoptosis
and how to repair mucosal tissues has become the focus of therapy
of UC (17). Studies have identified
p53, BAX, caspase-3 as essential proteins involved in apoptosis
(18). In the present study, results
demonstrated that LPS significantly increased apoptosis as well as
the protein expression levels of p53, BAX and caspase-3.
Taraxasterol treatment decreased apoptosis rate, and decreased the
protein levels of these apoptosis-related proteins. In addition,
taraxasterol treatment effectively reduced pathological injury of
colonic epithelial tissues in UC mouse models. Furthermore, p53
expression in colonic tissue was decreased and TNF-α and IL-6
levels were reduced following taraxasterol treatment.
In summary, the present study identified that
taraxasterol significantly inhibited secretion of TNF-α and IL-6
in vivo and in vitro, and decreased apoptosis and
associated protein levels. Taken together, these findings suggest
that taraxasterol could decrease inflammation and apoptosis
associated with UC.
Acknowledgements
The authors would like to thank the members of The
First Affiliated Hospital of Zhengzhou University, for providing
helpful discussions and technical support concerning the present
study.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HJL analyzed and interpreted the main data regarding
the cell function study and immunofluorescence. XYL was responsible
for design and drafting of the manuscript. DBJ was responsible for
statistical analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institute
Research Medical Ethics Committee of The First Affiliated Hospital
of Zhengzhou University (Zhengzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Scarpa M, Castagliuolo I, Castoro C, Pozza
A, Scarpa M, Kotsafti A and Angriman I: Inflammatory colonic
carcinogenesis: A review on pathogenesis and immunosurveillance
mechanisms in ulcerative colitis. World J Gastroenterol.
20:6774–6785. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hisamatsu T, Kanai T, Mikami Y, Yoneno K,
Matsuoka K and Hibi T: Immune aspects of the pathogenesis of
inflammatory bowel disease. Pharmacol Ther. 137:283–297. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rosenberg L, Nanda KS, Zenlea T, Gifford
A, Lawlor GO, Falchuk KR, Wolf JL, Cheifetz AS, Goldsmith JD and
Moss AC: Histologic markers of inflammation in patients with
ulcerative colitis in clinical remission. Clin Gastroenterol
Hepatol. 11:991–996. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mankowska-Wierzbicka D, Swora-Cwynar E,
Poniedzialek B, Adamski Z, Dobrowolska A and Karczewski J:
Usefulness of selected laboratory markers in ulcerative colitis.
Eur Cytokine Netw. 26:26–37. 2015.PubMed/NCBI
|
|
5
|
Wei C, Tan CK, Xiaoping H and Junqiang J:
Acanthoic acid inhibits LPS-induced inflammatory response in human
gingival fibroblasts. Inflammation. 38:896–901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kitajima S, Takuma S and Morimoto M:
Changes in colonic mucosal permeability in mouse colitis induced
with dextran sulfate sodium. Exp Anim. 48:137–143. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Herz W and Mirrington RN: Identification
of pyrethrol with taraxasterol. J Pharm Sci. 55:1041966. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Piao T, Ma Z, Li X and Liu J: Taraxasterol
inhibits IL-1β-induced inflammatory response in human
osteoarthritic chondrocytes. Eur J Pharmacol. 756:38–42. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cooper HS, Murthy SN, Shah RS and
Sedergran DJ: Clinicopathologic study of dextran sulfate sodium
experimental murine colitis. Lab Invest. 69:238–249.
1993.PubMed/NCBI
|
|
10
|
Kinoshita T, Imamura R, Kushiyama H and
Suda T: NLRP3 mediates NF-κB activation and cytokine induction in
microbially induced and sterile inflammation. PLoS One.
10:e01191792015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Frost RA, Nystrom GJ and Lang CH:
Lipopolysaccharide regulates proinflammatory cytokine expression in
mouse myoblasts and skeletal muscle. Am J Physiol Regul Integr Comp
Physiol. 283:R698–R709. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liverani E, Rico MC, Yaratha L, Tsygankov
AY, Kilpatrick LE and Kunapuli SP: LPS-induced systemic
inflammation is more severe in P2Y12 null mice. J Leukoc Biol.
95:313–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Popa C, Netea MG, van Riel PL, van der
Meer JW and Stalenhoef AF: The role of TNF-alpha in chronic
inflammatory conditions, intermediary metabolism, and
cardiovascular risk. J Lipid Res. 48:751–762. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gupta RA, Motiwala MN, Mahajan UN and
Sabre SG: Protective effect of Sesbania grandiflora on acetic acid
induced ulcerative colitis in mice by inhibition of TNF-α and IL-6.
J Ethnopharmacol. 219:222–232. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mochizuki D, Adams A, Warner KA, Zhang Z,
Pearson AT, Misawa K, McLean SA, Wolf GT and Nör JE: Anti-tumor
effect of inhibition of IL-6 signaling in mucoepidermoid carcinoma.
Oncotarget. 6:22822–22835. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Edelblum KL, Yan F, Yamaoka T and Polk DB:
Regulation of apoptosis during homeostasis and disease in the
intestinal epithelium. Inflamm Bowel Dis. 12:413–424. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Di Sabatino A, Ciccocioppo R, Luinetti O,
Ricevuti L, Morera R, Cifone MG, Solcia E and Corazza GR: Increased
enterocyte apoptosis in inflamed areas of Crohn's disease. Dis
Colon Rectum. 46:1498–1507. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Al-Assaf AH, Alqahtani AM, Alshatwi AA,
Syed NA, Shafi G and Hasan TN: Mechanism of cadmium induced
apoptosis in human peripheral blood lymphocytes: The role of p53,
Fas and Caspase-3. Environ Toxicol Pharmacol. 36:1033–1039. 2013.
View Article : Google Scholar : PubMed/NCBI
|