Introduction
Currently, comprehensive therapy, including surgery,
chemotherapy and radiotherapy, is the main method for treating
breast cancer, but the curative ratio is low due to frequent
relapse. The resistance of cancer cells to chemotherapeutic drugs
is thought to be the principal cause for relapse. Numerous theories
have attempted to explain this phenomenon, and recently the cancer
stem/initiating cell (CSC) theory has attracted much interest.
In 2006, to evaluate the chemosensitivity of
glioblastoma multiforme (GBM) stem cells, Eramo et al
(1) treated stem cell clones
derived from different GBM patients with eight chemotherapeutic
agents and assessed the rate of cell death in comparison to
chemosensitive Jurkat leukemic cells and primary immature
erythroblast. They observed a marked resistance of GBM stem cells
to all the compounds used, whereas both Jurkat cells and
erythroblasts displayed high rates of cell death. After treatment
with chemotherapeutic agents, GBM stem cells were able to recover
and proliferate. In a study by Phillips et al,
CD24(−/low)/CD44(+) cancer-initiating cells were isolated from
MCF-7 and MDA-MB-231 breast cancer cell lines, and their response
to radiation was investigated. The results showed that the
cancer-initiating cells were more resistant to radiation than the
unisolated cells (2). In another
study which included primary breast cancer patients (n=108) treated
with neoadjuvant chemotherapy consisting of sequential paclitaxel
and epirubicin-based chemotherapy, breast cancer stem cells were
identified by immunohistochemical staining of aldehyde
dehydrogenase 1 (ALDH1). ALDH1-positive tumors were significantly
associated with a low rate of complete response (3). Similar results steadily emerged,
urging researchers to relate drug-resistance to cancer
stem/initiating cells (4).
CSCs which are not killed by chemotherapy, may
become the cause of relapse. However, knowledge concerning the
changes manifested in CSCs which are resistent to chemotherapy is
limited. Answers to questions concerned with the effects of
traditional chemotherapy drugs on CSC, the changes in CSCs treated
by chemotherapy, the relationship between CSCs and drug-resistance
proteins, and the relationship between CSCs and anti-apoptosis
genes may help to understand the mechanism of CSC drug-resistance
and to identify more effective methods of therapy which eventually
kill CSCs.
In the present study, the MDA-MB-468 breast cancer
cell line was cultured and different passages were treated
continually with the traditional chemotherapy drug 5-fluorouracil
(5-FU). For every cell generation, certain markers of CSCs,
drug-resistance proteins and the anti-apoptosis gene were examined
and compared to find a clue with which to resolve the
abovementioned questions. The CSC factors included β-catenin,
Octamer binding factor 3/4 (Oct 3/4), SRY-box 2 (SOX2) and
CD44+/CD24− cells. The markers involved with
drug-resistance proteins and anti-apoptosis genes were multidrug
resistance protein 1 (MRP1), breast cancer resistance protein
(BCRP) and survivin, respectively. Our results suggest that the
drug-resistance of cancer cells is mainly due to CSCs, and
chemotherapy leads to an alteration in the quantity or function of
breast CSCs demonstrating an alternate increase and decrease.
Materials and methods
Cell culture and chemotherapy
The MDA-MB-468 breast cancer cell line was a gift
from the Chinese-United Kingdom Medical Laboratory. The cells were
cultured at 37°C with 5% CO2 in DMEM (HyClone, USA) and
supplemented with 10% fetal calf serum (HyClone), 100 U/ml
penicillin G and 100 μg/ml streptomycin (Gibco, USA). The cells
were cultured in 75 cm2 flasks, and a 0.1 μg/ml
(concentration data were obtained from a preliminary experiment)
terminal concentration of 5-FU was added to the nutrient fluid when
cells grew to a 60–70% base area. Cell cultures were harvested
using 0.05% trypsin/EDTA (Gibco) in PBS and subcultured until
achieving a 80–90% base area. The second generation was cultured,
treated with chemotherapy and passaged in the same manner. Thus,
six generations of cells were cultured. The cells in every
generation were harvested and used in corresponding procedures in
the subsequent experiments. Three independent experiments were
performed. The control group was not treated with chemotherapy.
RNA extraction and semi-quantitative
RT-PCR assay
Cells were harvested and stored at −70°C in
TRIzol® (Invitrogen, USA). Total RNA was isolated by
extracting the TRIzol samples with chloroform followed by
separation and purification according to the manufacturer’s
instructions. RNase inhibitor mix (1 μl) was added per 50 μl total
RNA prior to treatment with DNase (Promega, USA) for 20 min. The
RNA concentration and purity were quality controlled by analysis
using a spectrophotometer (U-3010, Japan). First-strand cDNA was
synthesized from the total RNA (3 μg) using the First-Strand
Synthesis system (Promega) and used as a template for PCR
amplification with Taq polymerase (Qiagen, Germany).
Specific primer sequences are listed in Table I. PCR conditions included an
initial denaturation at 94°C for 5 min, and 35 cycles at 94°C for
30 sec, annealing (Table I) for 45
sec, 72°C for 45 sec and a final extension step at 72°C for 5 min.
RT-PCR products were separated by 1.2% agarose gel electrophoresis
in a 0.5% Tris-acetate buffer and stained with ethidium bromide.
The bands were photographed using a digital camera (Canon, Japan),
and the absorbance of every band was calculated using Software
Quantity One (Bio-Rad, USA). The absorbances of the inspected bands
were divided by the absorbance of β-actin in the same sample to
obtain the relative value of the marker in the sample. At least six
independent experiments were performed.
 | Table I.Information regarding the PCR
primers. |
Table I.
Information regarding the PCR
primers.
| Primer | NCBI ID | Primer
sequence | Product size
(bp) | Melting temperature
(°C) |
|---|
| β-catenin | NM_001098209.1 | F:
CCCACTAATGTCCAGCGTTT | 382 | 54.0 |
| R:
AACCAAGCATTTTCACCAGG |
| Oct 3/4 | NM_002701.4 | F:
TTCAGCCAAACGACCATC | 484 | 57.4 |
| R:
GGAAAGGGACCGAGGAGTA |
| SOX2 | NM_003106.2 | F:
ACACCAATCCCATCCACACT | 223 | 61.0 |
| R:
CAAACTTCCTGCAAAGCTCC |
| MRP1 | NM_019898.2 | F:
CTGAGTTCCTGCGTACCTAT | 212 | 56.2 |
| R:
TTCTGCGGTGCTGTTGT |
| BCRP | NM_004827.2 | F:
GCCATTCTCCCAGTCA | 494 | 49.7 |
| R:
GGGCGTCTATACACCAT |
| Survivin | NM_001012271.1 | F:
CTTGCCAGAGCCACGAA | 632 | 55.4 |
| R:
GGAACCTCACCCATAGCC |
| β-actin | NM_001101.3 | F:
GCCCTGAGGCACTCTTC | 330 | 54.9 |
| R:
GGCCGGACTCGTCATAC |
Flow cytometry
To identify the CD44+/CD24−
cell proportion in every generation, cells were harvested with
0.05% trypsin/ EDTA (Gibco) and then suspended (2×106
cells/100 μl) in Stain Buffer containing 1% FBS (EBioscience, USA).
Phycoerythrin (PE)-conjugated mouse against human CD44 monoclonal
antibody (EBioscience) and fluorescein isothiocyanate
(FITC)-conjugated mouse anti-human CD24 monoclonal antibody
(EBioscience) were added to the cell suspension at the
concentrations recommended by the manufacturer, and incubation was
carried out at 4°C in the dark for 60 min. Proper isotype controls
were used for each cell labeling experiment. The labeled cells were
fixed in 100% methanol on ice for 5 min. Flow cytometric analysis
was performed in triplicate using a flow cytometer (Partec,
Germany). Four generations were assessed.
Colony formation experiment
Single cells were prepared in each generation and
seeded in culture dishes (60 mm in diameter); each dish initially
had 10×103 cells. The cells were cultured in the same
culture medium as the former (but without 5-FU) for 2–3 weeks and
fixed with methanol when colonies were visible by the naked eye.
The colonies were stained for 2–3 min with haematoxylin and those
that consisted of ≥50 cells were counted under a microscope. Four
generations were assessed. At least six independent experiments
were performed.
Statistical analysis
Data are expressed as the means ± standard deviation
(SD). The statistical significance of differences between groups
was analyzed by one-way analysis of variance, and the relationship
between variables was calculated by two-tailed Pearson correlation
analysis using the Statistical Product and Service Solutions
version 13.0 program for Windows (SPSS, USA). Differences with
p-values <0.05 were considered significant.
Results
mRNA expression of CSC factors,
drug-resistance genes and an anti-apoptosis gene
In the experimental (treated with 5-FU) and control
groups (without 5-FU), the mRNA expression of the markers,
β-catenin, Oct 3/4, SOX2, MRP1, BCRP, survivin and actin, in six
cell generations was investigated.
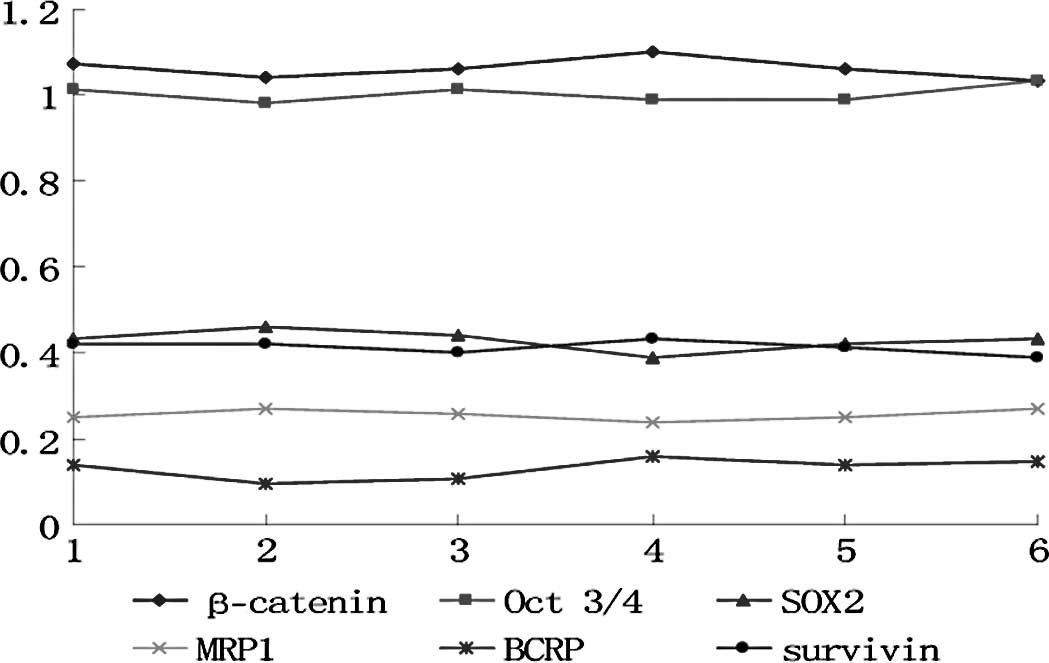
In the control group, each marker exhibited no
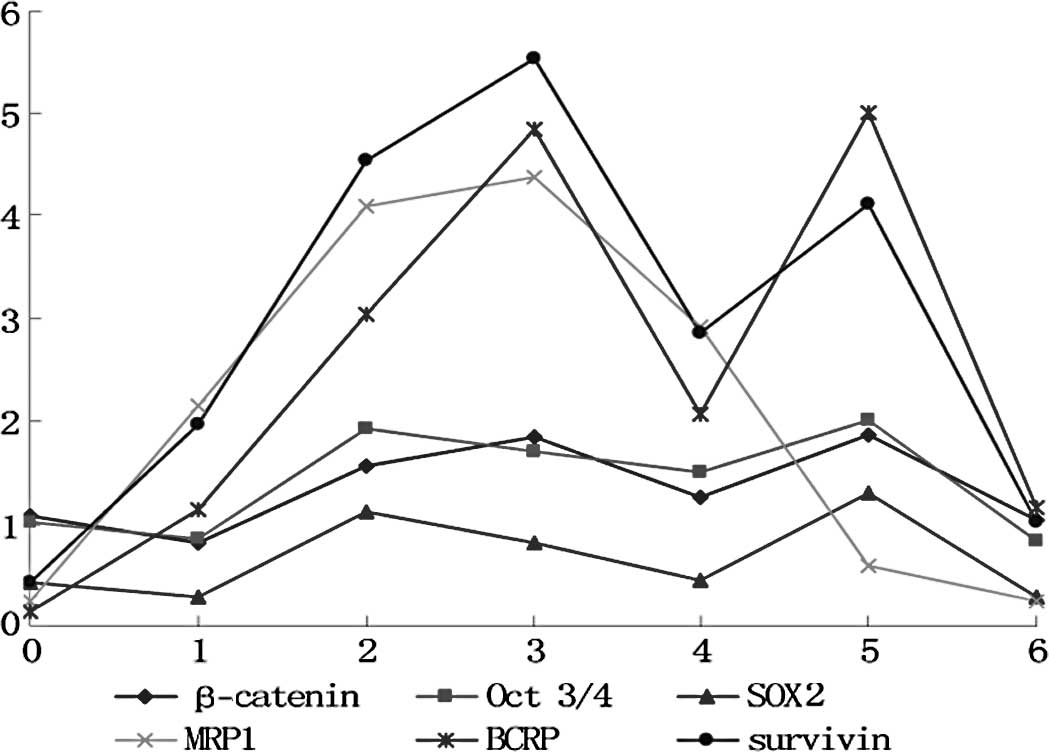
differences among the cell generations (p>0.05) (Fig. 1). In the experimental group, the
markers fluctuated in the different cell generations (p<0.05)
(Figs. 2 and 3). All markers selected in the present
study showed a similar wave of increases and decreases (Fig. 3) The factors related to CSCs
(β-catenin, Oct 3/4 and SOX2) exhibited a fluctuating trend of
increase-increase-further increase-decrease-increase-decrease. The
drug-resistance genes (BCRP and MRP) and the anti-apoptosis gene
(survivin) exhibited waves of increase-further increase-further
increase-decrease-increase (MRP1 decrease)-decrease.
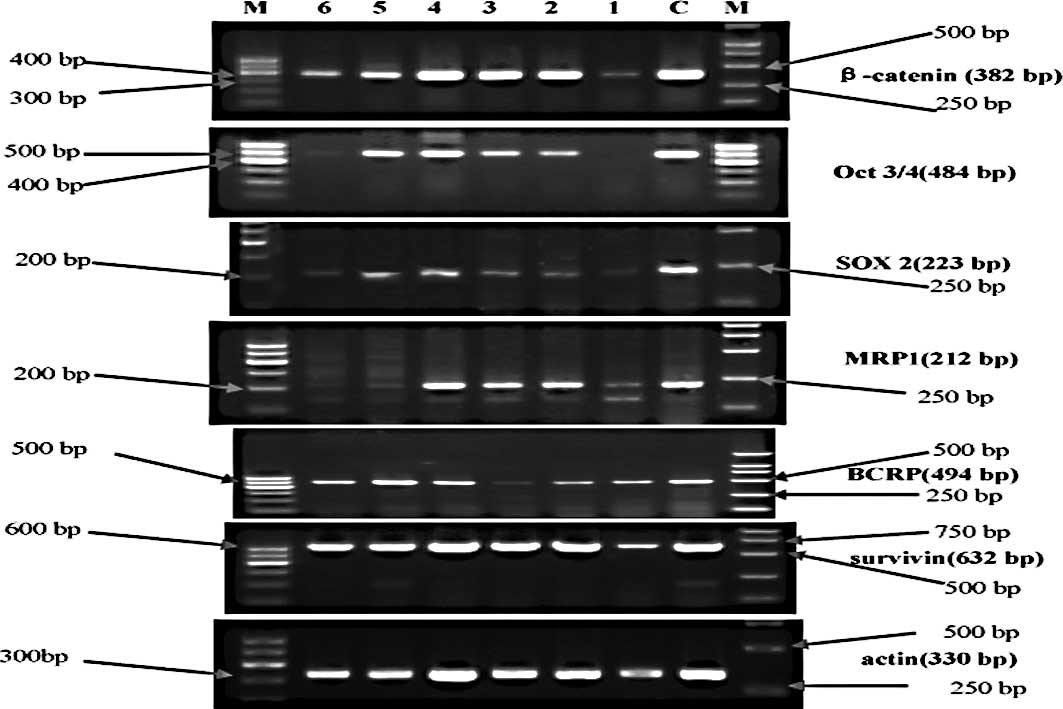 | Figure 2.Electrophorogram of mRNA expression of
the relative markers in each cell generation in the experimental
group. M, molecular weight marker; the left marker is the 100-bp
ladder, the right marker is DL2000 (apart from Oct 3/4, whose
marker was the 100-bp ladder). C, control group. Lanes 1, 2, 3, 4,
5 and 6 represent the first, second, third, fourth, fifth and sixth
cell generation, respectively. |
The correlationship among the markers was also
analyzed and most exhibited a positive correlation (Table II).
| Table II.Correlation among the
stem/initiating-related factors, chemoresistance and anti-apoptotic
factors. |
Table II.
Correlation among the
stem/initiating-related factors, chemoresistance and anti-apoptotic
factors.
| Factors | | β-catenin | Oct 3/4 | SOX2 | MRP1 | BCRP | Survivin |
|---|
| β-catenin | r-value | 1 | 0.922 | 0.882 | 0.389 | 0.936 | 0.859 |
| p-value | | 0.003 | 0.009 | 0.389 | 0.002 | 0.013 |
| Oct 3/4 | r-value | 0.922 | 1 | 0.933 | 0.486 | 0.861 | 0.860 |
| p-value | 0.003 | | 0.002 | 0.269 | 0.013 | 0.013 |
| SOX2 | r-value | 0.882 | 0.933 | 1 | 0.279 | 0.824 | 0.759 |
| p-value | 0.009 | 0.002 | | 0.544 | 0.023 | 0.048 |
| MRP1 | r-value | 0.389 | 0.486 | 0.279 | 1 | 0.436 | 0.753 |
| p-value | 0.389 | 0.269 | 0.544 | | 0.329 | 0.051 |
| BCRP | r-value | 0.936 | 0.861 | 0.824 | 0.436 | 1 | 0.917 |
| p-value | 0.002 | 0.013 | 0.023 | 0.329 | | 0.004 |
| Survivin | r-value | 0.859 | 0.860 | 0.759 | 0.753 | 0.917 | 1 |
| p-value | 0.013 | 0.013 | 0.048 | 0.051 | 0.004 | |
Proportion of
CD44+/CD24− cells
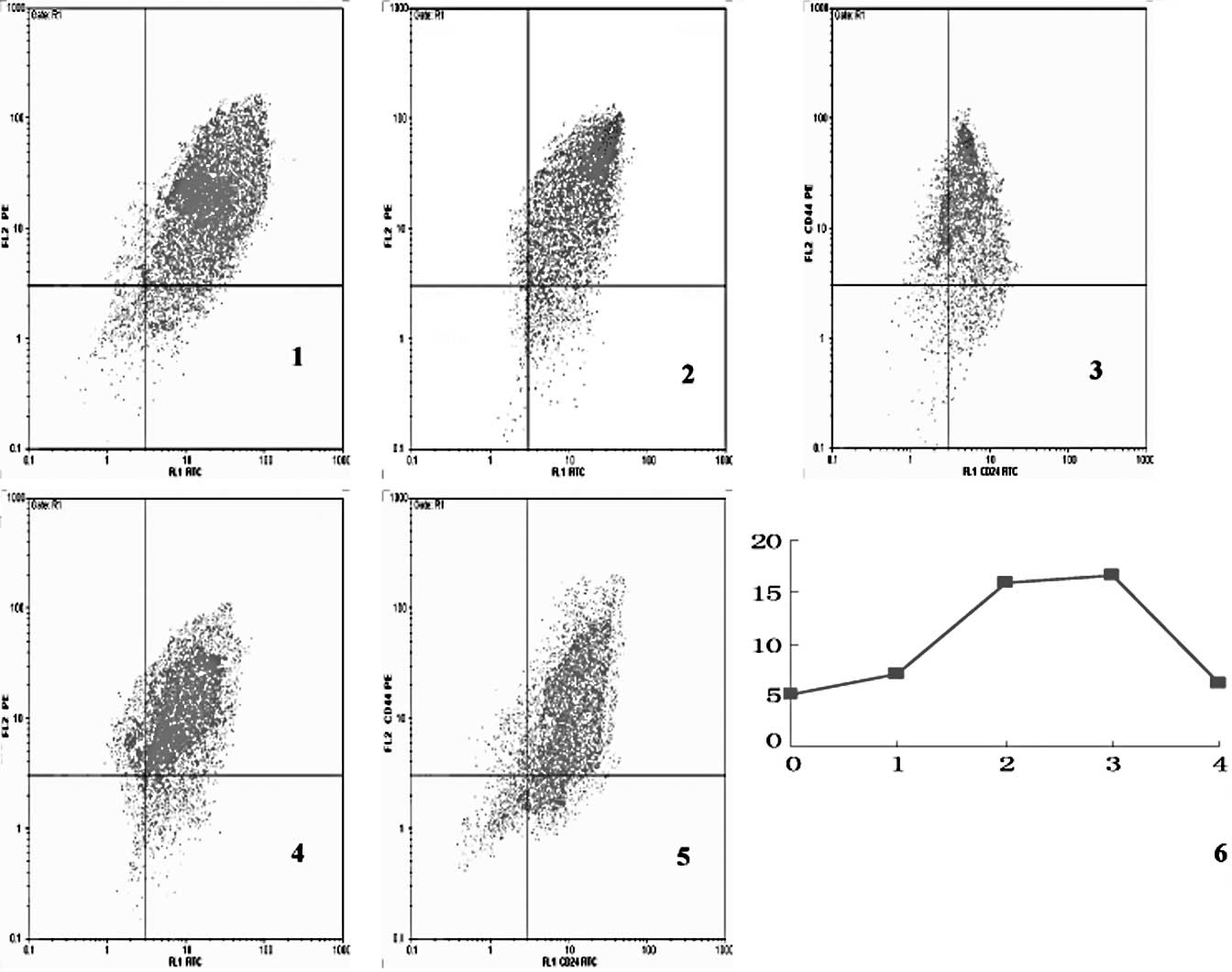
The proportion of CD44+/ CD24−
cells in the experimental group also exhibited a trend of decrease
and increase in each cell generation (p<0.05) (Fig. 4). The proportion of
CD44+/CD24− cells in each cell generation in
the control group showed no difference among the different cell
generations (p>0.05; data not shown).
The proportion of CD44+/CD24−
cells in the experimental group showed a positive correlation with
other markers (Table III).
| Table III.Correlation between the proportion of
CD44+/CD24− cells and the other factors. |
Table III.
Correlation between the proportion of
CD44+/CD24− cells and the other factors.
| β-catenin | Oct 3/4 | SOX2 | MRP1 | BCRP | Survivin |
|---|
| Proportion of
CD44+/CD24− | | | | | | |
| r-value | 0.857 | 0.804 | 0.889 | 0.846 | 0.891 | 0.921 |
| p-value | 0.063 | 0.101 | 0.043 | 0.071 | 0.042 | 0.026 |
Colony formation
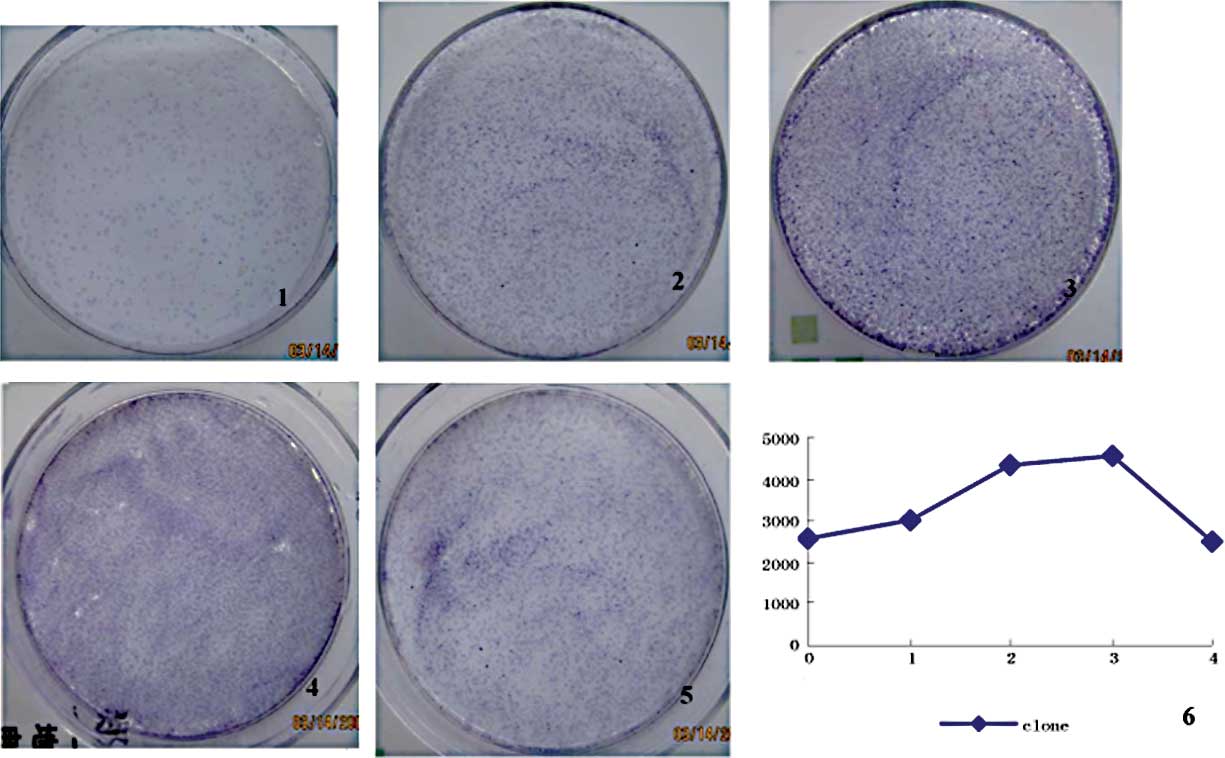
In the different cell generations, the colonies
formed per 104 seeded cells varied (p<0.05) (Fig. 5). The decrease and increase
fluctuation was very similar to that shown in Fig. 4. The number of colonies formed in
the experimental group exhibited a positive correlation with
several of the other markers (Table
IV).
| Table IV.Correlation between the rate of clone
formation and the other stem cell-related factors. |
Table IV.
Correlation between the rate of clone
formation and the other stem cell-related factors.
| β-catenin | Oct 3/4 | SOX2 | MRP1 | BCRP | Survivin |
CD44+/CD24− |
|---|
| Clone
formation | | | | | | | |
| r-value | 0.811 | 0.722 | 0.850 | 0.792 | 0.858 | 0.880 | 0.992 |
| p-value | 0.096 | 0.168 | 0.068 | 0.110 | 0.063 | 0.049 | 0.001 |
Discussion
In the present study, three factors were selected to
represent the existence or quantity/functional change in CSCs: mRNA
expression of β-catenin, Oct 3/4 and SOX2. β-catenin is the key
component in the Wnt/β-catenin signaling pathway. The Wnt/β-catenin
signaling pathway plays an important role in the differentiation
and proliferation of stem cells (5). Experiments have confirmed that
activation of the Wnt/β-catenin signaling pathway maintains stem
cell renewal capacity and restrains stem cell differentiation in
normal tissue (6–9), whereas it prompts tumor stem cell
amplification in tumor tissue (9).
Oct 3/4 is an embryo-specific transcription factor, which maintains
totipotency (10) of the embryo
stem cells and participates in the renewal of stem cells (11). Oct 3/4 is thought to be one of the
markers of totipotency or stem cells. Its expression was found to
be extremely low or absent in differentiated cells, but was
increased in normal stem and cancer stem cells (12,13).
SOX2 was found to be expressed in totipotency cells and early
multipotential cells in the embryo, regulating the self-renewal
process (14,15) of embryo stem cells. SOX2 has been
associated with the maintainence of cell pluripotency (16,17)
and is also considered as a marker of stem cells (18). In light of the above findings, the
mRNA expression of β-catenin, Oct 3/4 and SOX2 may be considered to
represent the quantity of activated stem/initiating cells.
The other two markers of stem/initiating cells are
CD44+/ CD24− cells and the rate of colony
formation. According to a study by Al-Hajj et al (19) as well as other previous studies
(20–23), CD44+/CD24−
cells in breast cancer tissue exhibit certain stem cell-like
properties, such as self-renewal and multi-directional
differentiation, therefore they are considered to be breast cancer
stem/initiating cells. The capacity to form colonies reflects
renewal ability, so under equivalent seeding conditions, the rate
of colony formation reflects the proportion of stem/initiating
cells in the primitive seeded cells. In the present study, these
two markers represented the proportion of stem/initiating
cells.
In the experimental group, the mRNA expression of
the above markers in the six generations exhibited an evident
fluctuation with statistical significance, whereas the mRNA
expression in the control group did not fluctuate. The results
indicate that chemotherapy surely affects the quantity or function
of CSCs.
The mRNA expression of three markers representing
the quantity of CSCs (β-catenin, Oct 3/4 and SOX2) exhibited the
following trend of fluctuation: decrease-increase-further
increase-decrease-increase-decrease (1–6 generation). The two
markers representing the proportion of CSCs exhibited the following
fluctuating trend: increase-further increase-further
increase-decrease (1–4 generation). In the first cell generation,
the quantity of CSCs decreased slightly since some of them were
killed by 5-FU (even though they had drug-resistance properties).
More non-stem/initiating cells were killed compared to CSCs,
therefore the proportion of CSCs increased in the first generation
cells. CSCs are activated and proliferate through stimulation of
5-FU, a decrease in cell number, and factors released from dead
cells. Thus, the quantity and proportion of CSCs increased
significantly in the second and third generations. In the fourth
generation, most of the CSCs differentiated to relatively mature
tumor cells, leading to the quantity and proportion of CSCs to
decrease. The differentiated tumor cells had no or low
drug-resistance, so a large portion of them were killed and the
CSCs were again activated and proliferated, leading to an increase
in the quantity and proportion of CSCs in the fifth generation
cells. The situation in the sixth generation was similar to the
fourth generation, reflecting the alteration resulting from CSC
differentiation.
The results indicate that the quantity and
proportion of CSCs increase and decrease in turn in different cell
generations under a condition of persistent chemotherapy. This
finding has not been previously reported.
The fact that the mRNA expression of β-catenin, Oct
3/4 and SOX2 exhibits a nearly perfect positive correlation
(p<0.01) suggests that they come from the same population of
cells, proving their reliability to represent CSCs. The percentage
of CD44+/CD24− cells exhibited nearly a
perfect positive correlation (r=0.99, p<0.01) with the rate of
colony formation, confirming their reliability as indicators of the
proportion of CSCs.
Another key point in the present study is the
drug-resistance of CSC. The MRP1 gene is a member of the
superfamily of ATP-binding cassette (ABC) transporters. This full
transporter is a member of the MRP subfamily which is involved in
multi-drug resistance (24–27).
MRP proteins markedly increase in side population (SP) cells
(28–30), in which CSCs are rich. The BCRP
gene is also included in the superfamily of ABC transporters. BCRP
has been proven to be involved in the multi-drug resistance of many
types of cancers (particularly breast cancer) (31–33)
and in CSCs (24–27).
mRNA expression of BCRP fluctuated along with an
increase and decrease in the mRNA expression of β-catenin, Oct 3/4
and SOX2 in the second, third, fourth, fifth and sixth generation,
showing a positive correlation between them. The results suggest
that BCRP expression is a property of CSCs, and there is low BCRP
expression in differentiated cells. This finding is consistent with
that of Doyle et al (32),
who argued that the number of SP cells (abundant in CSCs) in
MCF-7/AdrVp (a multidrug-resistant human breast cancer subline) was
higher than in parental MCF-7 cells, and BCRP was overexpressed in
the former. A similar study (34)
showed that CD133-positive cells (assumed markers of glioblastoma
cancer stem cells) isolated from human glioblastoma expressed
higher levels of BCRP1 mRNA. MRP1 did not demonstrate a positive
correlation with CSC-related factors; thus, MRP1 may be expressed
in both CSCs and differentiated cells.
Survivin is a member of the inhibitor of apoptosis
gene family, which encodes negative regulatory proteins that
prevent apoptotic cell death (35,36).
In the present study, the expression of survivin mRNA was
positivlye correlated with factors of CSCs, suggesting that the
expression of survivin is associated with the drug-resistance of
CSCs. In fact, overexpression of anti-apoptosis genes in cancer
stem cells has been previously reported (34,37,38).
‘Survivor cells’ surviving chemotherapy in acute myelocytic
leukemia, for example, were found to be enriched in
CD34+ stem cells and accompanied by overexpression of
many anti-apoptosis genes. BCRP may cooperate with survivin to
maintain CSC survival.
In summary, the overexpression of drug-resistance
genes and anti-apoptosis genes in CSCs is an important cause of
traditional chemotherapy failure and cancer relapse. To cure
cancer, effective ways to eliminate CSCs must be developed.
References
|
1.
|
Eramo A, Ricci-Vitiani L, Zeuner A, et al:
Chemotherapy resistance of glioblastoma stem cells. Cell Death
Differ. 13:1238–1241. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Phillips TM, McBride WH and Pajonk F: The
response of CD24(−/low)/CD44+ breast cancer-initiating cells to
radiation. J Natl Cancer Inst. 98:1777–1785. 2006.
|
|
3.
|
Tanei T, Morimoto K, Shimazu K, et al:
Association of breast cancer stem cells identified by aldehyde
dehydrogenase 1 expression with resistance to sequential paclitaxel
and epirubicin-based chemotherapy for breast cancers. Clin Cancer
Res. 15:4234–4241. 2009. View Article : Google Scholar
|
|
4.
|
Dean M, Fojo T and Bates S: Tumour stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar
|
|
5.
|
Mimeault M, Hauke R, Mehta PP and Batra
SK: Recent advances in cancer stem/progenitor cell research:
therapeutic implications for overcoming resistance to the most
aggressive cancers. J Cell Mol Med. 11:981–1011. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Reguart N, He B, Taron M, You L, Jablons
DM and Rosell R: The role of Wnt signaling in cancer and stem
cells. Future Oncol. 1:787–797. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Mishra L, Shetty K, Tang Y, Stuart A and
Byers SW: The role of TGF-beta and Wnt signaling in
gastrointestinal stem cells and cancer. Oncogene. 24:5775–5789.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
He B, Barg RN, You L, et al: Wnt signaling
in stem cells and non-small-cell lung cancer. Clin Lung Cancer.
7:54–60. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Kruger JA, Kaplan CD, Luo Y, et al:
Characterization of stem cell-like cancer cells in immune-competent
mice. Blood. 108:3906–3912. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Scholer HR, Dressler GR, Balling R,
Rohdewohld H and Gruss P: Oct-4: a germline-specific transcription
factor mapping to the mouse t-complex. EMBO J. 9:2185–2195.
1990.PubMed/NCBI
|
|
11.
|
Scholer HR, Ruppert S, Suzuki N, Chowdhury
K and Gruss P: New type of POU domain in germ line-specific protein
Oct-4. Nature. 344:435–439. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Odorico JS, Kaufman DS and Thomson JA:
Multilineage differentiation from human embryonic stem cell lines.
Stem Cells. 19:193–204. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Hubner K, Fuhrmann G, Christenson LK, et
al: Derivation of oocytes from mouse embryonic stem cells. Science.
300:1251–1256. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Chew JL, Loh YH, Zhang W, et al:
Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the
Oct4/Sox2 complex in embryonic stem cells. Mol Cell Biol.
25:6031–6046. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Park IH, Zhao R, West JA, et al:
Reprogramming of human somatic cells to pluripotency with defined
factors. Nature. 451:141–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Maruyama M, Ichisaka T, Nakagawa M and
Yamanaka S: Differential roles for Sox15 and Sox2 in
transcriptional control in mouse embryonic stem cells. J Biol Chem.
280:24371–24379. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Masui S, Nakatake Y, Toyooka Y, et al:
Pluripotency governed by Sox2 via regulation of Oct3/4 expression
in mouse embryonic stem cells. Nat Cell Biol. 9:625–635. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Ginis I, Luo Y, Miura T, et al:
Differences between human and mouse embryonic stem cells. Dev Biol.
269:360–380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Ponti D, Costa A, Zaffaroni N, et al:
Isolation and in vitro propagation of tumorigenic breast cancer
cells with stem/progenitor cell properties. Cancer Res.
65:5506–5511. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Balic M, Lin H, Young L, et al: Most early
disseminated cancer cells detected in bone marrow of breast cancer
patients have a putative breast cancer stem cell phenotype. Clin
Cancer Res. 12:5615–5621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Mylona E, Giannopoulou I, Fasomytakis E,
et al: The clinicopathologic and prognostic significance of
CD44+/CD24(-/low) and CD44−/CD24+
tumor cells in invasive breast carcinomas. Hum Pathol.
39:1096–1102. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Wright MH, Calcagno AM, Salcido CD,
Carlson MD, Ambudkar SV and Varticovski L: Brca1 breast tumors
contain distinct CD44+/CD24− and
CD133+ cells with cancer stem cell characteristics.
Breast Cancer Res. 10:R102008. View
Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Stride BD, Valdimarsson G, Gerlach JH,
Wilson GM, Cole SP and Deeley RG: Structure and expression of the
messenger RNA encoding the murine multidrug resistance protein, an
ATP-binding cassette transporter. Mol Pharmacol. 49:962–971.
1996.PubMed/NCBI
|
|
25.
|
D’Hondt V, Caruso M and Bank A:
Retrovirus-mediated gene transfer of the multidrug
resistance-associated protein (MRP) cDNA protects cells from
chemotherapeutic agents. Hum Gene Ther. 8:1745–1751.
1997.PubMed/NCBI
|
|
26.
|
Lautier D, Canitrot Y, Deeley RG and Cole
SP: Multidrug resistance mediated by the multidrug resistance
protein (MRP) gene. Biochem Pharmacol. 52:967–977. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Abaan OD, Mutlu PK, Baran Y, Atalay C and
Gunduz U: Multidrug resistance mediated by MRP1 gene overexpression
in breast cancer patients. Cancer Invest. 27:201–205. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Dean M: ABC transporters, drug resistance,
and cancer stem cells. J Mammary Gland Biol Neoplasia. 14:3–9.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Chuthapisith S, Eremin J, El-Sheemey M and
Eremin O: Breast cancer chemoresistance: emerging importance of
cancer stem cells. Surg Oncol. 19:27–32. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Dave B and Chang J: Treatment resistance
in stem cells and breast cancer. J Mammary Gland Biol Neoplasia.
14:79–82. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Allikmets R, Schriml LM, Hutchinson A,
Romano-Spica V and Dean M: A human placenta-specific ATP-binding
cassette gene (ABCP) on chromosome 4q22 that is involved in
multidrug resistance. Cancer Res. 58:5337–5339. 1998.PubMed/NCBI
|
|
32.
|
Doyle LA, Yang W, Abruzzo LV, et al: A
multidrug resistance transporter from human MCF-7 breast cancer
cells. Proc Natl Acad Sci USA. 95:15665–15670. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Ross DD, Yang W, Abruzzo LV, et al:
Atypical multidrug resistance: breast cancer resistance protein
messenger RNA expression in mitoxantrone-selected cell lines. J
Natl Cancer Inst. 91:429–433. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Liu G, Yuan X, Zeng Z, et al: Analysis of
gene expression and chemoresistance of CD133+ cancer
stem cells in glioblastoma. Mol Cancer. 5:672006. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Khan S, Aspe JR, Asumen MG, et al:
Extracellular, cell-permeable survivin inhibits apoptosis while
promoting proliferative and metastatic potential. Br J Cancer.
100:1073–1086. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Tamm I, Wang Y, Sausville E, et al:
IAP-family protein survivin inhibits caspase activity and apoptosis
induced by Fas (CD95), Bax, caspases, and anticancer drugs. Cancer
Res. 58:5315–5320. 1998.PubMed/NCBI
|
|
37.
|
Wang J, Guo LP, Chen LZ, Zeng YX and Lu
SH: Identification of cancer stem cell-like side population cells
in human nasopharyngeal carcinoma cell line. Cancer Res.
67:3716–3724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Kornblau SM, Qiu YH, Bekele BN, et al:
Studying the right cell in acute myelogenous leukemia: dynamic
changes of apoptosis and signal transduction pathway protein
expression in chemotherapy resistant ex-vivo selected ‘survivor
cells’. Cell Cycle. 5:2769–2777. 2006.PubMed/NCBI
|