Introduction
The role of adventitial cells in restenosis after
percutaneous transluminal angioplasty (PTA) has not been
investigated and no related results were reported until the middle
to late 1990s (1–3). Upon arterial injury after PTA or
intravascular stenting, the vascular adventitia is involved in the
pathological repair or restenosis of blood vessels. Several studies
have shown that restenosis following PTA involves the migration of
not only vascular smooth muscle cells (VSMCs) in the media but
cells of the vascular adventitia (4,5).
That is, fibroblasts in the vascular adventitia may migrate from
the adventitia into the intima through the media, and the newly
generated intima contains not only VSMCs but myofibroblasts that
were transformed from fibroblasts in the adventitia (6,7).
However, the migration of cells in the adventitia has still not
been confirmed in vascular repair following injury. Several studies
have even yielded contradicting results, to the effect that the
migration of fibroblasts from the adventitia into the media is
absent (8,9). A number of researchers have even
postulated that a stem cell pool is present in the vascular
adventitia. Following vascular injury, cells may produce the recall
response that occurs in embryonic development. Non-muscle cells
(stem cells in the adventitia) may transform into myofibroblasts
and further into VSMCs (10). In
the present study, the phenotypic transformation and migration of
adventitial cells were observed and analyzed by morphological
examination and detection of protein expression. Thus, we may
provide evidence on the role of cell migration in vascular
restenosis following PTA.
Materials and methods
Animal model of common carotid artery
restenosis following PTA
We establishment an animal model of common carotid
artery restenosis using a modified wire method. A total of 24 male
SD rats weighing 250–300 g were divided into 4 groups and
sacrificed at 3, 7, 14 and 28 days after vascular injury. For
preparation of the arterial injury model, animals were
intraperitoneally anesthetized with 10% chloral hydrate (350 mg/kg)
and then a midline incision was made in the neck. Following
separation of subcutaneous tissues, the bilateral common carotid
artery (CCA) and external carotid artery (ECA) were exposed. A
metal wire 0.038 inches in diameter was introduced into one of the
CCAs through the ECA and the wire was forwarded and withdrawn 3–4
times to injure the vascular intima and all the layers of the blood
vessels, which is similar to the vascular injury following balloon
injury or intravascular stenting. The rats with injured CCAs served
as the experimental group and those with intact CCAs were the
control group.
Preparation of the antigen marker and
labeling
5-Bromo-2′-deoxyuridine (BrdU) is involved in the
synthesis of DNA in proliferating cells. After BrdU was integrated
into proliferating adventitial cells, immunohistochemistry was
performed to detect the number of BrdU-positive cells. First, the
BrdU powder was dissolved in PBS at a final concentration of 25%
(0.25 mg/ml). Then, pluronic biogum powder was mixed together with
the above BrdU solution at a final concentration of 2.5% (2.5
mg/ml). The injured and uninjured arteries were washed with
sterilized normal saline and the liquid was removed. The BrdU
solution was mixing by shaking before use, and then, 0.2 ml of this
solution was used to smear the bilateral CCAs.
Sample collection
Tissues were collected, embedded in paraffin and cut
into 5-μm consecutive sections which were adherent to the
APES-treated slides. The sections were heated at 58–60°C for 30–60
min and the slides were allowed to cool to room temperature for
further use.
Double immunohistochemistry for BrDU and
α-actin
Immunohistochemistry was performed using a
traditional protocol. The sections were treated with BrDU
monoclonal antibody to stain the proliferating and migrating cells
in the different vascular layers. Following BrDU staining, these
sections were treated with α-actin monoclonal antibody for double
immunohistochemistry. BrDU and BrDU monoclonal antibody were
purchased from Sigma Chemical Co. and α-actin monoclonal antibody,
the ABC detection kit and DAB substrate kit were from Wuhan Boster
Biotech Co., Ltd.
Determination of the number of BrDU-
and/or α-actin-positive cells
Immunohistochemistry for BrDU was carried out alone
and BrDU-positive cells displayed brown granules in the nucleus.
Immunohistochemistry was also carried out for both BrDU and
α-actin, and the BrDU-positive cells had a dark blue nucleus while
α-actin-positive cells had brown cytoplasm. Cells positive for both
BrDU and α-actin displayed a dark blue nucleus and brown
cytoplasm.
Counting of BrDU-positive cells
Four fields were selected at 4 quadrants of the
blood vessel wall at a high magnification by 2 researchers blinded
to the study details. The number of BrDU-positive cells in the
vascular intima, media and adventitia were counted and the average
eas calculated. The number of BrDU-positive cells in the blood
vessels was the sum of these cells in the different vessel layers.
The ratio of the number of BrDU-positive cells in each layer to the
total number of BrDU-positive cells was calculated. This ratio was
used to monitor changes in the number of positive cells in the
blood vessels.
Electron microscopy
The 1-mm3 samples of tissues were fixed
in 2.5% glutaraldehyde in 0.1 mol/l sodium cacodylate buffer for 1
h. After dehydration, embedding, sectioning and staining with
uranyl acetate, the tissues were ultra-sectioned using an LKB-V
ultramicrotome (Sweden) and the myofibroblasts in the different
layers were observed under a transmission electron microscope
(Hitachi H-7500, Japan).
Statistical analysis
Statistical analysis was carried out with SPSS
version 10.0, and the t-test and analysis of variance were
performed for comparisons. A value of P<0.05 was considered to
indicate statistical significance.
Results
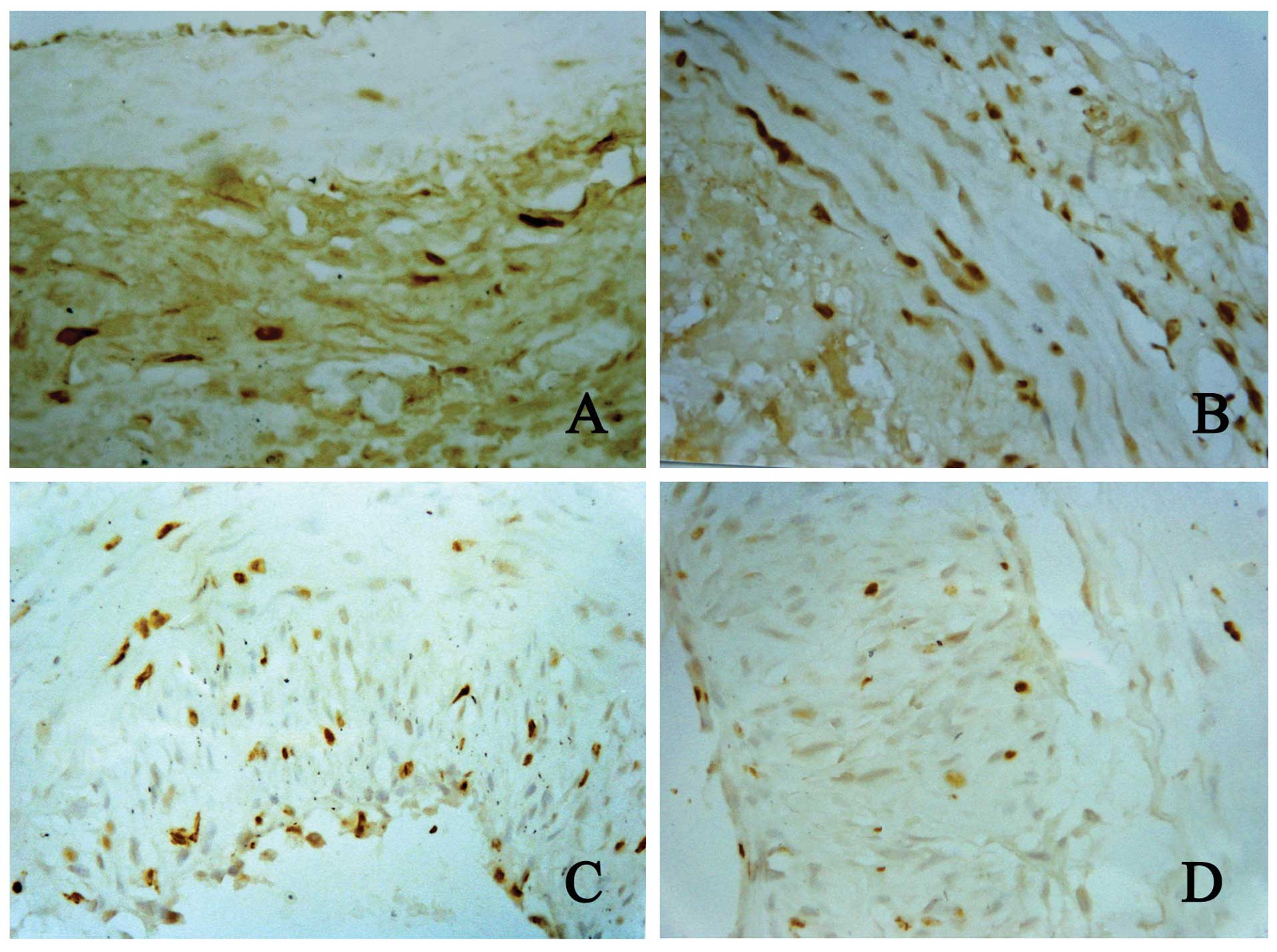
Immunohistochemistry for BrDU
Three days after PTA, at high magnification, the
number of adventitial cells increased and many cells had a brown
nucleus indicative of proliferating cells. In the experimental
group, the number of BrDU-positive cells was higher than that in
the control group (cells having a brown nucleus were not found in
the adventitia). In the media, an extremely small number of
proliferating cells had a brown nucleus. In the media close to the
lumen, cells with a brown nucleus were not present. Shedding of
endothelial cells was noted but newly generated intima was absent.
Seven days after PTA, the number of adventitial cells were markedly
increased when compared with those 3 days after PTA, and the number
of proliferating cells with a brown nucleus also increased. In the
media, some proliferating cells had a brown nucleus and were
distributed around the external elastic lamina. In the newly
generated intima, a few new intimal cells with a brown nucleus were
noted. Fourteen days after PTA, the number of adventitial cells was
dramatically reduced when compared with the number of adventitial
cells 3 and 7 days after PTA, and the number of proliferating cells
displaying a brown nucleus also declined. In addition, many
proliferating cells with a brown nucleus were noted in the media.
In the relatively thick newly generated intima, a large number of
proliferating cells with a brown nucleus was noted. At 28 days
after PTA, only a few adventitial cells were found when compared
with those 3, 7 and 14 days after PTA, and these cells were
long-spindle-like while proliferating cells with a brown nucleus
were not found. In the experimental group, the findings in the
adventitia were similar to those in the control group. The media
was thickened and proliferating cells with a brown nucleus were
occasionally noted. The thick newly generated intima had a few
proliferating cells with a brown nucleus (Fig. 1).
At high magnification, the cells in the different
layers of the blood vessels were observed at different time points.
Results showed that the number of BrDU-positive cells changed
significantly in the experimental group. In the adventitia, median
and intima, the number of BrDU-positive cells reached a maximal
level at 7 and 14 days after PTA, respectively. The peak number of
BrDU-positive cells in the different layers occurred alternatively
in a chronological sequence (Fig.
1). Three and 28 days after PTA, the number of positive cells
in the adventitia, media and intima was similar to those in the
control group. At these two time points, proliferating cells
(BrDU-positive) with a brown nucleus were rarely found. The number
of BrDU-positive cells in the different layers at different time
points is shown in Table I.
 | Table I.Number of BrDU-positive cells in the
different vascular layers at different time points. |
Table I.
Number of BrDU-positive cells in the
different vascular layers at different time points.
| BrDU-positive cells,
mean ± SD
|
|---|
| Layers | 3 days | 7 days | 14 days | 28 days | Control group |
|---|
| Adventitia | 161.3±52.18 | 247.2±33.72 | 80.0±9.34 | 4.3±1.97a | 2.1±2.99a |
| Media | 8.8±4.96b | 52.8±25.20 | 164.3±28.39 | 8.2±4.79b | 1.3±8.944b |
| Intima | 0.00±0.00c | 165.3±2.86c | 274.5±53.69 | 45.5±13.78 | 1.0±2.56c |
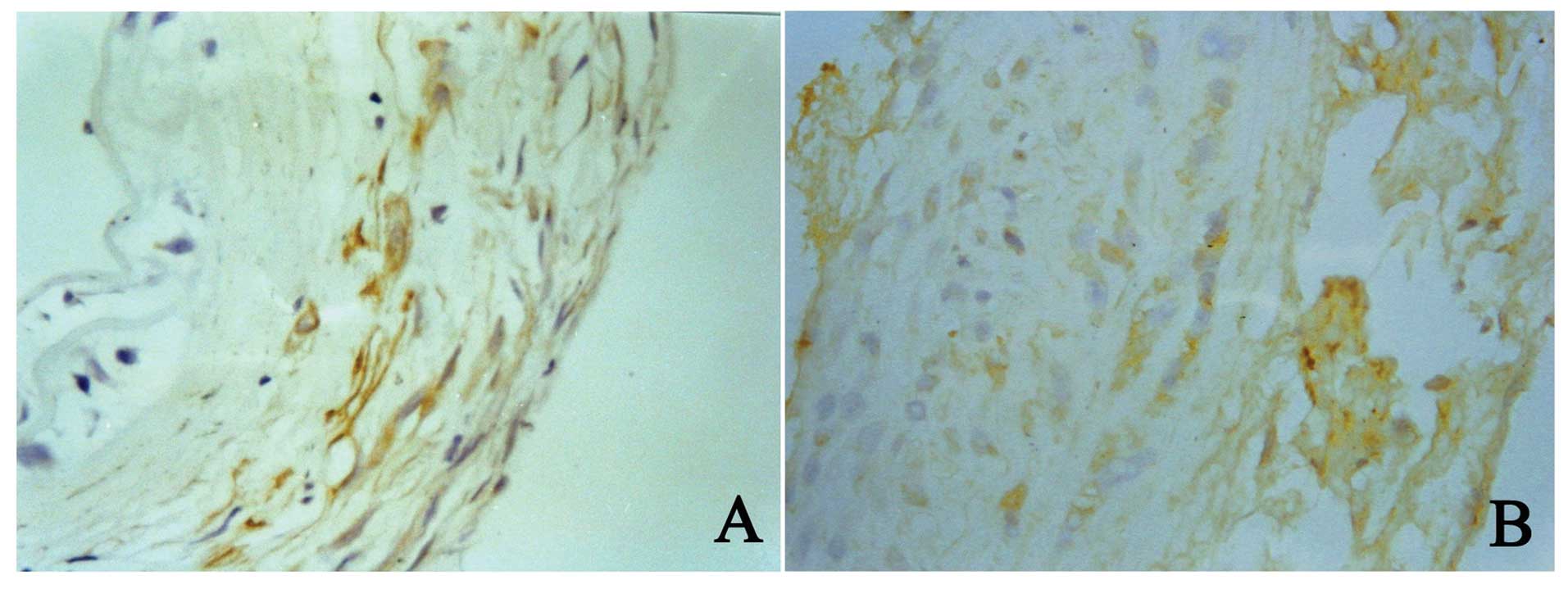
Double immunohistochemistry for BrDU and
α-actin
Seven days after PTA, the number of adventitial
cells increased, the media was thickened and newly generated intima
was barely noticeable. At 14 days after PTA, the intima was
significantly thickened. Thus, in the experimental group, samples
collected at 7 and 14 days were used for further double
immunohisto-chemistry for BrDU and α-actin. The nucleus was stained
by BrDU and BrDU-positive cells had a blue nucleus.
α-actin-positive cells had brown cytoplasm. The results showed, 7
days after PTA, that the BrDU-positive cells in the adventitia were
also positive for α-actin, suggesting that the proliferating cells
also expressed α-actin. In addition, these cells aggregated around
the external elastic lamina. At 14 days after PTA, the number of
cells positive for both BrDU and α-actin were markedly reduced but
these cells in the newly generated intima increased dramatically
(Fig. 2). Double
immunohistochemistry revealed that the phenotype of the adventitial
cells changed after PTA, and that the adventitial cells had
transformed into myofibroblasts.
Electron microscopy
Three days after PTA, the fibroblasts in the
adventitia exhibited a mature phenotype in the control group. The
cells and nuclei were long-spindle-shaped and the cells had less
pseudopod-like processes and less cytoplasm. After arterial injury,
the spindle-shaped fibroblasts were found in the adventitia and the
collagen and pseudopod-like processes increased. Seven days after
PTA, cells with different shapes and sizes were found and the
nuclei were irregular and spindle-like. These cells were noted to
be myofibroblasts. These cells had processes with varied sizes and
the number of processes increased and formed lamellipodia
stretching to the external elastic lamina and crossing the fenestra
of the external elastic lamina. These cells had a tendency to
migrate from the adventitia into the media. Fourteen days after
PTA, the intima was markedly thickened and spindle-like and oval
myofibroblasts were noted. The longitudinal axis of cell growth was
vertical to the internal elastic lamina. Changes in the
ultrastructures of the myofibroblasts were similar to those 7 days
after PTA, but the distribution of intrinsic structures in the
artery was different. At 14 days after PTA, the number of cells in
the media and intima increased and the fenestra of internal elastic
lamina enlarged. In the intima, myofibroblasts formed several
processes and the lamellipodia stretched to and crossed the
external elastic lamina. These cells had a tendency to migrate into
the intima (Fig. 3). Twenty-eight
days after PTA, the myofibroblasts in the different vascular layers
transformed reversely into fibroblasts which were characterized by
small nuclei, less nucleoplasm, small but obvious nucleoli and less
cytoplasm. In addition, the number of pinocytotic vesicles was
reduced and the lamellipodia almost disappeared in these cells.
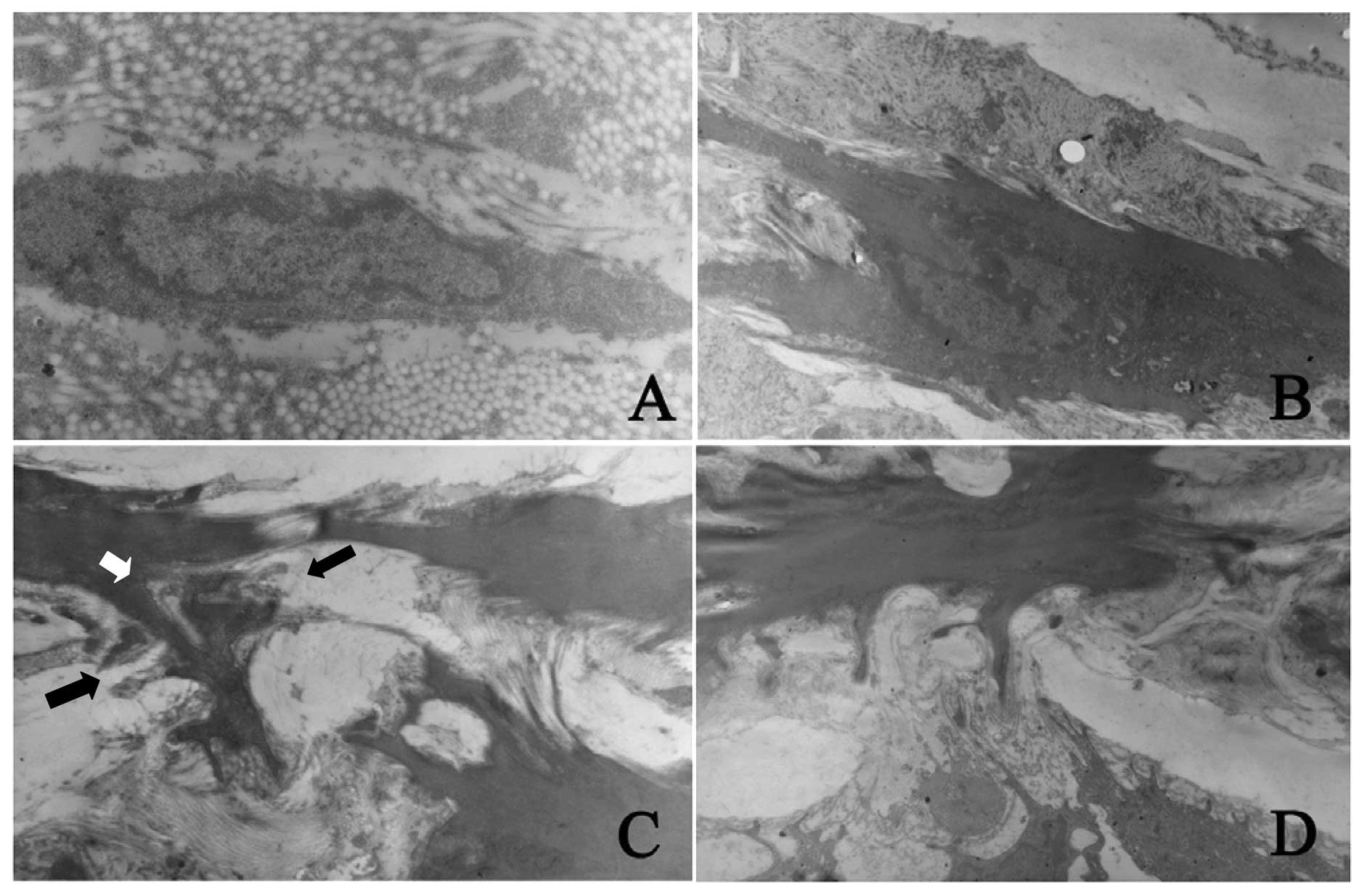 | Figure 3.Detection of morphological changes and
migration of fibroblasts after PTA by scanning electron microscopy.
(A) The adventitial fibroblasts in normal blood vessels had a
contractile quiescent phenotype. These cells had long-spindle
nuclei but had less pseudopod-like processes (magnification,
x15,000); (B) 7 days after PTA, the myofibroblasts in adventitia
found processes with different sizes and the processes increased
(magnification, x7,000); (C) 7 days after PTA, the myofibroblasts
in adventitia formed a lot of processes and the lamellipodia (white
arrow) crossed the fenestra of external elastic lamina (black
arrow). Cells had a trend to migrate from adventitia into media
(magnification, x10,000); (D) 14 days after PTA, the fenestra of
internal elastic lamina enlarged and the myofibroblasts in the
media formed several processes. The lamellipodia crossed the
fenestra of internal elastic lamina (as in C). Cells had a trend to
migrate from media into intima (magnification, x10,000). |
Discussion
Cells in the adventitia are related to
vascular restenosis following PTA and their characteristics are
described
In mammals and humans, the vascular adventitia
mainly consists of fibroblasts. Studies have shown that fibroblasts
are involved in vascular repair following arterial injury. Soon
after PTA, proliferating cells are present in the vascular
adventitia or even in the media. These cells are large,
spindle-like and similar to VSMCs but different from fibroblasts in
functions (11). Patel et
al found in an in vitro experiment that non-muscle cells
in the vascular adventitia were highly adherent and proliferative
and active in the synthesis of collagen. Thus, some researchers
postulate that non-muscle cells in the adventitia are derived from
fibroblasts and are also named myofibroblasts. However, the source
and characteristics of myofibroblasts are still controversial.
Immunohistochemistry has demonstrated that these cells synthesize
and secrete α-actin. α-actin has been regarded as a specific marker
of muscular cells. That is to say, the spindle-like cells which
synthesize and secret α-actin are muscular cells (MCs). Thus,
myofibroblasts in blood vessels were previously referred to as
VSMCs in different environments (12). However, Skalli et al
(13) speculated that these cells
are possibly derived from fibroblasts. In studies on myofibroblasts
in human normal and pathological tissues, antibodies against desmin
or vimentin or allogeneic different actin antibodies were used.
Results revealed that general myofibroblasts express vimentin and
cytoplasmic actin. Thus, researchers postulated that these cells
were derived from fibroblasts. In pathological tissues (such as
fibromatosis), the myofibroblasts have various characteristics:
some express only vimentin (type V) and some express both vimentin
and α-actin (type VA). Although components of the cytoskeleton are
different, vimentin is consistently found in the cytoskeleton.
These cells may have the same source (such as fibroblasts), and
they express cytoskeletal proteins found in smooth muscles
following specific stimulation (14). Thus, under normal conditions,
fibroblasts express vimentin alone and do not express α-actin or
desmin. After arterial injury, fibroblasts in the adventitia were
found to express α-actin and their morphology was also altered.
Scott et al defined these cells with changes in morphology
in the adventitia and media by immunohistochemistry: myofibroblasts
do not express desmin, caldesmon (a marker of well-differentiated
VSMCs) and myosin but inductively express α-actin (4). These characteristics suggest that
myofibroblasts are different from VSMCs. However, both fibroblasts
and myofibroblasts can express vimentin. Thus, we speculate that
both cells have a common source: fibroblasts can transform into
myofibroblasts. In the present study, a large amount of collagen
bundle was found in the fibroblasts in the vascular adventitia
under electron microscopy and the morphology was similar to that of
fibroblasts. Cells had several processes, which were different from
VSMCs. Immunohistochemistry indicated actin expression. Based on
the findings of the morphological examination and
immunohistochemistry, we can speculate that fibroblasts undergo
phenotypic transformation and transform into myofibroblasts.
Phenotypic transformation of cells in the
adventitia
A majority of researchers postulate that
proliferating cells at an early stage following PTA are VSMCs in
several regions of the vascular media. VSMCs have been found to
proliferate at 24–48 h after vascular injury (15) and their proliferation reaches a
peak at 7–14 days after injury. These proliferating cells are
mainly found in the vascular media. Following PTA, VSMCs in the
vascular media auto-regulate and the VSMC phenotype switches from a
contractile quiescent to a proliferative motile phenotype
(synthetic phenotype). These cells are star-like in morphology and
organelles increased. However, these cells lack contraction while
proliferation, migration and secretion are significantly enhanced.
In vascular restenosis, the phenotype of VSMCs reverses
approximately 2 weeks after arterial injury, and the VSMC phenotype
switches from a proliferative motile to a contractile quiescent
phenotype. At 4–6 months after injury, VSMCs are in a relatively
quiescent state when the restenosis has formed (16). However, our findings were
inconsistent with those described above. Under light microscopy, at
3–7 days after PTA, the cells in the vascular adventitia were
significantly altered and a large amount of short-spindle-like
fibroblasts were noted. They had large nuclei and full cytoplasm
and were in a proliferative state. Analysis of BrDU-positive cells
in all layers showed that cell proliferation was active in the
vascular adventitia, which was consistent with the findings of
Scott et al who observed that cell proliferation was mainly
found in the adventitia at an early stage following PTA and the
proliferation of adventitial cells was more obvious than those in
the media. These findings differed from previous results that early
cell proliferation was predominantly found in the vascular media.
Double immunohisto chemistry showed, 7 days after PTA, that the
BrDU-positive cells began to express α-actin. The myofibroblasts
were transformed from fibroblasts located near the external elastic
lamina. The α-actin expression was also found in the newly
generated intima 14 days after PTA. Twenty-eight days after PTA,
the α-actin expression was absent in the intima and adventitia.
This suggests that cells migrating into the newly generated intima
lose the ability to express α-actin at a late stage following PTA
and these cells were transformed into fibroblasts. Wilcox and Scott
(5) found that, at an early stage
after arterial injury, proliferating adventitial cells were
negative for α-actin. Phenotypic transformation occurred at 14 days
after PTA and cells were positive for α-actin. Our results
indicated that the time when fibroblasts underwent phenotypic
transformation and expressed α-actin was different from that noted
in the study of Wilcox and Scott. Our findings also confirmed that
fibroblasts in the vascular adventitia underwent phenotypic
transformation after arterial injury (6). The fibroblasts were transformed into
myofibroblasts which possess the ability to proliferate,
synthesize, migrate and specifically secrete and express α-actin.
Thus, not only VSMCs in the media but fibroblasts in the adventitia
have throwback after PTA. That is to say, the phenotype of these
cells becomes primitive (17),
which is shared by fibroblasts and VSMCs and is also a premise of
the proliferation and migration of adventitial cells. In the
present study, as noted by electron microscopy, myofibroblasts had
many mitochondria and active Golgi body and the granules on rough
endoplasmic reticulum increased, which suggest that the cells
acquired the functions of synthesis and secretion following
phenotypic transformation.
Migration of adventitial cells
The migration of proliferating adventitial cells
into the intima following arterial injury is still controversial
(18–23). Our findings suggest that
proliferating adventitial cells migrate into the intima and are the
main source of newly generated intima. At 3 days after PTA, the
proliferating cells were mainly found in the adventitia. Seven days
after PTA, the number of proliferating cells in the adventitia
reached a maximal level and these cells were mainly found around
the external elastic lamina and the proliferating cells in the
media began to increase. At 14 days after PTA, the proliferating
cells in the adventitia reduced but those in the media and intima
were markedly increased. The number of BrDU-positive cells in both
the intima and media reached a peak at this time point and the
thickness of the intima was maximized. At 28 days after PTA, the
BrDU-positive cells in the adventitia, media and the newly
generated intima were significantly reduced and the proliferating
cells were almost absent in the adventitia and media. Analysis of
the BrDU-positive cells in the different vascular layers showed
that the number of BrDU-positive cells increased sequentially and
alternatively: the increase in BrDU-positive cells in the media and
newly generated intima occurred when the proliferating cells in the
adventitia began to decrease. This may be explained by the fact
that the myofibroblasts in the vascular adventitia migrated into
the media and intima. These findings are consistent with those in a
study by Scott et al. Smearing CCAs with BrDU solution can
directly and effectively label myofibroblasts. Of note, in the
present study, smearing CCAs with BrDU immediately after PTA only
labeled the adventitial cells and labeling was not performed later.
Thus, we must address the question of the origin of the the
BrDU-positive cells in the newly generated intima 7–14 days after
PTA. We speculate that the BrDU cells in the vascular adventitia
proliferated and migrated from the adventitia into the intima,
which is also supported by other studies (24). In addition, as noted at high
magnification under a light microscope, a large amount of
BrDU-positive cells in the newly generated intima with thickening
were short-spindle-like and had full cytoplasm. These morphological
features significantly resembled those of long-spindle-like VSMCs
in the media, and suggest that these cells are similar to
myofibroblasts in the vascular adventitia. Results in other studies
also support our findings; following VSMC activation under in
vitro control, stimulation of the vascular adventitia could
induce the migration of characteristic fibroblasts in the
adventitia (25). Following
transfection with a retrovirus carrying the β-galactosidase LacZ
gene, primary fibroblasts in the adventitia were inoculated into
the adventitia of rat CCAs to induce LacZ expression. In
situ hybridization was employed to monitor the myofibroblasts.
Results showed LacZ expression in the injured intima. In newly
generated intima, the morphology of LacZ-positive fibroblasts was
similar to adventitial cells indirectly labeled with BrDU. These
findings indicate that the newly generated intima had cells derived
from the adventitia (26,27). In addition, we also observed
findings similar to those in the study of Scott et al
(4). The nuclei of BrDU-positive
cells in adventitia, media and intima became light from 3 to 28
days after PTA. This implies that, from 3 days after PTA, the
BrDU-labeled myofibroblasts in the adventitia migrated into the
newly generated intima in which they underwent division,
replication and proliferation. Thus, these cells lose BrDU-labeled
DNA. These findings suggest that proliferating adventitial cells
migrate from the adventia and cross the external elastic lamina and
become the main cells in the newly generated intima. Additionally,
electron microscopy showed, 7 days after PTA, that the
myofibroblasts in the adventitia formed lamellipodia which
stretched to the external elastic lamina and crossed the fenestra
of the external elastic lamina. These cells had a tendency to
migrate into the media. At 14 days after PTA, the fenestra of the
internal elastic lamina in the media enlarged. The myofibroblasts
formed lamellipodia with several processes which stretched to the
internal elastic lamina and crossed the fenestra of the internal
elastic lamina. These cells had a tendency to migrate into the
intima.
Taken together, in vascular restenosis following
PTA, adventitial fibroblasts undergo phenotypic transformation and
migration. These cells migrate into the newly generated intima,
transform into myofibroblasts and thus are involved in vascular
restenosis.
Acknowledgements
Some of the results of the study were
from a doctoral dissertation for Tianjin Medical University and we
thank He N.S. for assistance with the research study.
References
|
1
|
Gutterman DD: Adventitia-dependent
influences on vascular function. Am J Physiol. 277:H1265–H1272.
1999.PubMed/NCBI
|
|
2
|
Barker SG, Tilling LC, Miller GC, et al:
The adventitia and atherogenesis: removal initiates intimal
proliferation in the rabbit which regresses on generation of a
‘neoadventitia’. Atherosclerosis. 105:131–144. 1994.PubMed/NCBI
|
|
3
|
Schneider DB, Sassani AB, Vassalli G,
Driscoll RM and Dichek DA: Adventitial delivery minimizes the
proinflammatory effects of adenoviral vectors. J Vasc Surg.
29:543–550. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scott NA, Cipolla GD, Ross CE, Dunn B,
Martin FH, Simonet L and Wilcox JN: Identification of a potential
role for the adventitia in vascular lesion formation after balloon
overstretch injury of porcine coronary arteries. Circulation.
93:2178–2187. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wilcox JN and Scott NA: Potential role of
the adventitia in arteritis and atherosclerosis. Int J Cardiol.
54(Suppl): S21–S35. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi Y, O’Brien JE, Fard A, et al:
Adventitial myofibroblasts contribute to neointimal formation in
injured porcine coronary arteries. Circulation. 94:1655–1664. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Haurani MJ, Cifuentes ME, Shepard AD, et
al: Nox4 oxidase overexpression specifically decreases endogenous
Nox4 mRNA and inhibits angiotensin II-induced adventitial
myofibroblast migration. Hypertension. 52:143–149. 2008. View Article : Google Scholar
|
|
8
|
Hecto DL, Jeremy DO, Kathy KG, et al:
Adventitial cells do not contribute to neointimal mass after
balloon angioplasty of the rat common carotid artery. Circulation.
104:1591–1593. 2001.PubMed/NCBI
|
|
9
|
Fleenor BS and Bowles DK: Negligible
contribution of coronary adventitial fibroblasts to neointimal
formation following balloon angioplasty in swine. Am J Physiol
Heart Circ Physiol. 296:H1532–H1539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Faggin E, Puato M, Zardo L, et al: Smooth
muscle-specific SM22 protein is expressed in the adventitial cells
of balloon-injured rabbit carotid artery. Arterioscler Thromb Vasc
Biol. 19:1393–1404. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Patel S, Shi Y, Niculescu R, Chung EH,
Martin JL and Zalewski A: Characteristics of coronary smooth muscle
cells and adventitial fibroblasts. Circulation. 101:524–532. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sipes JM, Guo N, Negre E, et al:
Inhibition of fibronectin binding and fibronectin-mediated cell
adhesion to collagen by a peptide from the second type I repeat of
thrombospondin. J Cell Biol. 121:469–477. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Skalli O, Darby I and Gabbiani G: Smooth
muscle actin is transiently expressed in by myofibroblasts during
experimental wound healing. Lab Invest. 63:21–29. 1990.PubMed/NCBI
|
|
14
|
Gabbiani G, Rungger-Brandle E, de
Chastonay C, et al: Vimentin-containing smooth muscle cells in
aortic intimal thickening after endotherlial injury. Lab Invest.
20:196–202. 1982.PubMed/NCBI
|
|
15
|
Marnur JD, Rossikhina M, Guha A, Fyfe B,
et al: Tissue factor is rapidly induced in arterial smooth muscle
after balloon injury. J Clin Invest. 91:2253–2259. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Reidy MA, Fingerle J and Linddner V:
Factors controlling the development of arterial lesions after
injury. Circulation. 86(Suppl 6): III43–III46. 1992.PubMed/NCBI
|
|
17
|
Hogeman B, Gillessen A, Bocker W, et al:
Myofibroblast-like cells produce mRNA for type I and III
procollagens in chronic active hepatitis. Scan J Gastroenterol.
28:591–594. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wexberg P, Muck K, Windberger U, et al:
Adventitial response to intravascular brachytherapy in a rabbit
model of restenosis. Wien Klin Wochenschr. 116:190–195. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kingsley K, Huff JL, Rust WL, et al:
ERK1/2 mediates PDGF-BB stimulated vascular smooth muscle cell
proliferation and migration on laminin-5. Biochem Biophys Res
Commun. 293:1000–1006. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Z and Newman WH: Smooth muscle cell
migration stimulated by interleukin 6 is associated with
cytoskeletal reorganization. J Surg Res. 111:261–266. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oparil S, Chen SJ, Chen YF, et al:
Estrogen attenuates the adventitial contribution to neointima
formation in injured rat carotid arteries. Cardiovasc Res.
44:608–614. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mallawaarachchi CM, Weissberg PL and Siow
RC: Smad7 gene transfer attenuates adventitial cell migration and
vascular remodeling after balloon injury. Arterioscler Thromb Vasc
Biol. 25:1383–1387. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Torsney E, Hu Y and Xu Q: Adventitial
progenitor cells contribute to arteriosclerosis. Trends Cardiovasc
Med. 15:64–68. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Damrauer SM, Fisher MD, Wada H, et al: A20
inhibits post-angioplasty restenosis by blocking macrophage
trafficking and decreasing adventitial neovascularization.
Atherosclerosis. 211:404–408. 2010. View Article : Google Scholar
|
|
25
|
Li G, Chen YF, Greene GL, et al: Estrogen
inhibits vascular smooth muscle cell-dependent adventitial
fibroblast migration in vitro. Circulation. 100:1639–1645. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li G, Chen SJ, Oparil S, Chen YF and
Thompson JA: Direct in vivo evidence demonstrating neointimal
migration of adventitial fibroblasts after ballooninjury of rat
carotid arteries. Circulation. 101:1362–1365. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Appleby CE and Kingston PA: Gene therapy
for restenosis - what now, what next? Curr Gene Ther. 4:153–182.
2004. View Article : Google Scholar : PubMed/NCBI
|