Introduction
Nonalcoholic fatty liver disease (NAFLD) is
characterized by increased fat deposits in the liver and can
precede more severe diseases, including nonalcoholic
steatohepatitis, cirrhosis and in certain cases, hepatocellular
carcinoma (1). According to a
previous epidemiological survey, NAFLD is highly prevalent,
affecting ~20% of the general population (2). In addition, NAFLD is strongly
associated with metabolic syndrome and associated obesity, type 2
diabetes mellitus, arterial hypertension and dyslipidemia. NAFLD is
considered to be the major hepatic component of metabolic syndrome.
Despite large involvement in key human health crises, the
pathogenesis of NAFLD is not completely understood; although the
pathogenesis is considered to be multifactorial, including genetic,
environmental and behavioral variables (3). As a result, current treatments for
patients with fatty liver disease include lifestyle alterations and
targeting the reduction of elevated lipolysis and high circulating
free fatty acid (FFA) levels (4,5).
Preventing increased circulating FFA levels in the liver, which can
trigger a series of biological changes in hepatic lipid metabolism,
may be an effective therapy for multiple stages of NAFLD (6). Cellular FFA loading is commonly
utilized to develop in vitro models of steatosis and these
models can reliably reproduce key features of hepatic steatosis in
humans (7,8).
5′-AMP-activated protein kinase (AMPK) is a critical
modulator of pathways involved in hepatic lipid metabolism. This
kinase plays a central role in lipid metabolism regulation by
activating fatty acid oxidation pathways and inhibiting lipid
synthesis (7). AMPK is a
heterotrimeric protein consisting of one catalytic subunit (α) and
two non-catalytic subunits (β and γ). In response to an elevated
cellular AMP/ATP ratio, AMPK is physiologically activated by
phosphorylation at the threonine 172 (Thr172) residue in the
α-subunit (9). AMPK activation is
responsible for metabolic modifications (10) via the phosphorylation of downstream
substrates, including acetyl-CoA carboxylase (ACC), the
rate-limiting enzyme in fatty acid biosynthesis. Cholesterol
production in the liver is also inhibited by AMPK activation via
the suppression of HMG-CoA reductase (11). AMPK inhibits de novo fatty
acid synthesis by inactivating ACC and promoting fatty acid
oxidation by upregulating the gene expression levels of carnitine
palmitoyltransferase-1 (CPT-1) and peroxisome
proliferator-activated receptor (PPAR)-α (12). Fatty acid synthase (FASN) is a
rate-limiting enzyme involved in fatty acid biosynthesis,
catalyzing the final step in this pathway.
As an effective medical therapy for NAFLD is yet to
be established, developing therapeutic agents for NAFLD is
critical. Previous studies have focused on Chinese herbal remedies
that can suppress hepatic lipid accumulation, including Cirsium
japonicum (CJ) (13). The
positive biological effects are wide-ranging, with CJ functioning
as a cancer chemopreventive agent, a powerful anti-inflammatory and
an antioxidant. Although numerous beneficial roles of CJ have been
hypothesized, there have been no studies investigating the role of
CJ in the regulation of genes associated with hepatocellular lipid
metabolism. If CJ is hepatoprotective, understanding the genetic
effects that the extract has may yield new insight into potential
therapies for a variety of liver diseases. Thus, the present study
investigated whether CJ can attenuate hepatic lipid accumulation
through activating AMPK utilizing human HepG2 cells.
Materials and methods
Materials and reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), penicillin-streptomycin and trypsin-EDTA were
purchased from Gibco-BRL (Carlsbad, CA, USA). TRIzol reagent and
dimethyl sulfoxide (DMSO) were purchased from Invitrogen Life
Technologies (Carlsbad, CA, USA). Oleic acid (OA) and palmitic acid
(PA) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Polyclonal antibodies against AMPK, phospho-AMPK, ACC, phospho-ACC,
CPT-1, FASN and β-actin were obtained from Cell Signaling
Technology, Inc., (Danvers, MA, USA). Assay kits for triglyceride
(TG), total cholesterol (TC) and Oil Red O staining were obtained
from the Jiancheng Institute of Biotechnology (Nanjing, China).
Other reagents and chemicals used were of the highest grade
commercially available.
Preparation of ethanol extract from
CJ
Authentic plant material was purchased from Guo Yi
Tang Chinese Herbal medicine store (Fujian, China). The stock
solution of CJ in the cell-based experiments was prepared by
dissolving pure CJ ethanol extract in 50% DMSO and 50%
phosphate-buffered saline (PBS) to a concentration of 500 mg/ml.
Diluting the stock solution in cell culture medium established the
working concentration. The final DMSO concentration in the medium
for all the cell experiments was <0.1%.
Cell culture and viability assay
HepG2 cells, a human hepatoma cell line, were
obtained from a cell bank of the Chinese Academy of Science
(Shanghai, China). Cells were cultured in DMEM supplemented with
antibiotics (100 U/ml penicillin A and 100 U/ml streptomycin) and
10% heat-inactivated FBS, and were maintained at 37°C in a
humidified incubator containing 5% CO2. Cells were
subcultured at 80–90% confluence. Cell viability was determined
using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) assay. In brief, HepG2 cells were seeded at a density
of 3×104 cells/well in a 96-well plate. The following
day, the culture medium was replaced with fresh medium containing
various concentrations of CJ, which was maintained for 24 h. Cells
were subsequently exposed to 1 mmol/l HFFA (OA and PA at a 2:1
ratio) or equal DMSO in fresh medium for 24 h. Following treatment,
10 ml MTT (5 mg/ml in PBS) was added to each well and the samples
were incubated for an additional 4 h at 37°C. The purple-blue MTT
formazan precipitate was dissolved in 100 ml DMSO and the
absorbance was measured at 570 nm using an ELISA reader (ELx800;
BioTek Instruments, Inc., Winooski, VT, USA).
Oil Red O staining
HepG2 cells were grown in six-well plates at a
density of 7×104 cells/well for 24 h. Stock solutions of
0.6 mmol/l OA and 0.6 mmol/l PA were prepared in DMSO. Following CJ
and HFFA (OA:PA, 2:1) treatment for 24 h, the cells were washed
twice with PBS and stained with Oil Red O for 15 min. Cell nuclei
were labeled by brief exposure to hematoxylin solution, and were
washed with PBS. The samples were observed under a microscope at a
magnification of ×200 and ×400.
Intracellular TG and TC measurements
To induce cellular fat-overloading, HepG2 cells at
60% confluence were cultured with or without 1.00 mmol/l HFFA
(OA:PA, 2:1) in the presence or absence of CJ at 0.5 or 1 mg/ml.
After 24 h, the cells were washed twice with 1 ml cold PBS.
Intracellular TG and TC levels were measured in the HepG2 cell
lysates and the concentrations were determined using commercial
kits based on phosphoglycerol oxidase/peroxidase enzymatic
reactions, according to the manufacturer’s instructions.
RNA extraction and quantitative
polymerase chain reaction (PCR) analysis
Total RNA was extracted using TRIzol reagent,
according to the manufacturer’s instructions. To obtain cDNA, 1 μg
total RNA was reverse-transcribed using PrimeScript® II
1st strand cDNA Synthesis kit (Takara Biotechnology Co., Ltd.,
Dalian, China). Real-time fluorescent quantitative PCR was
performed using a SYBR-Green premix (Applied Biosystems, Carlsbad,
CA, USA), according to the manufacturer’s instructions, under the
following cycling parameters: 1 cycle, 95°C for 7 min; 40 cycles,
95°C for 15 sec and 60°C for 1 min. The following primer sequences
were used: Human sterol regulatory element-binding protein-1c
sense, 5′-CCACTTCATCAAGGCAGACTCG-3′ and antisense,
5′-CAAGATGGTTCCGCCACTCAC-3′; human FASN sense,
5′-GCTTCCGAGATTCCATCCTACG-3′ and antisense,
5′-GCAGTCAGGCTCACAAACGA-3′; human ACC sense,
5′-GTTATGTGAAAGATGTGGATGA-3′ and antisense,
5′-TGTCTGAAGAGATTAGGGAAGT-3′; human PPAR-α sense,
5′-TCCATCGGCGAGGATAGTTCT-3′ and antisense,
5′-GGTGAAAGCGTGTCCGTGAT-3′; and human CPT-1 sense,
5′-AAATCAATCGGACTCTGGAAACG-3′ and antisense,
5′-TCTTGGTGGCACGACTCACTT-3′.
Cycling conditions were as follows: 95°C for 30 sec,
followed by 40 cycles at 95°C for 5 sec and 59°C for 30 sec.
Relative PCR product levels were determined based on the threshold
cycle value. Glyceraldehyde 3-phosphate dehydrogenase was used as a
normalized reference gene. Following amplification, melting curve
analysis was completed to verify the accuracy of the amplicon. The
2−ΔΔCT method was used to calculate the relative
fold-changes in mRNA expression.
Western blot analysis
Following CJ treatment, HepG2 hepatocytes were
washed with ice-cold PBS and lysed using radioimmunoprecipitation
assay buffer with protease and phosphatase inhibitors (Roche
Diagnostics, Mannheim, Germany) on ice for 30 min. Insoluble
proteins were removed by centrifugation at 14,000 × g at 4°C for 20
min. The cell lysate protein concentration was measured using a
bicinchoninic acid protein assay kit (Pierce Biotechnology, Inc.,
Rockford, IL, USA). Next, 50-μg samples of total protein were
loaded and separated by 10% SDS polyacrylamide gel electrophoresis
under denaturing and non-reducing conditions. The samples were then
transferred to polyvinylidine fluoride membranes (Millipore
Corporation, Billerica, MA, USA). The membranes were blocked with
5% non-fat milk in Tris-buffered saline and Tween-20 (TBST) at room
temperature for 2 h, which was followed by incubation with a
primary antibody at 4°C overnight. Following three TBST washes, the
membranes were incubated with secondary antibodies for 1 h at room
temperature. Proteins were detected with a chemiluminescence
detection system (Pierce Biotechnology, Inc.) and visualized with a
LuminoImager (LAS-3000 Bio Imaging analysis system; Fujifilm Co,
Ltd., Tokyo, Japan).
Statistical analysis
All data are expressed as the mean ± SE from three
measurements. Statistical significance of the differences among
multiple treatment groups was determined through one-way analysis
of variance. All the data were analyzed using the SPSS package for
Windows (version 18.0; SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
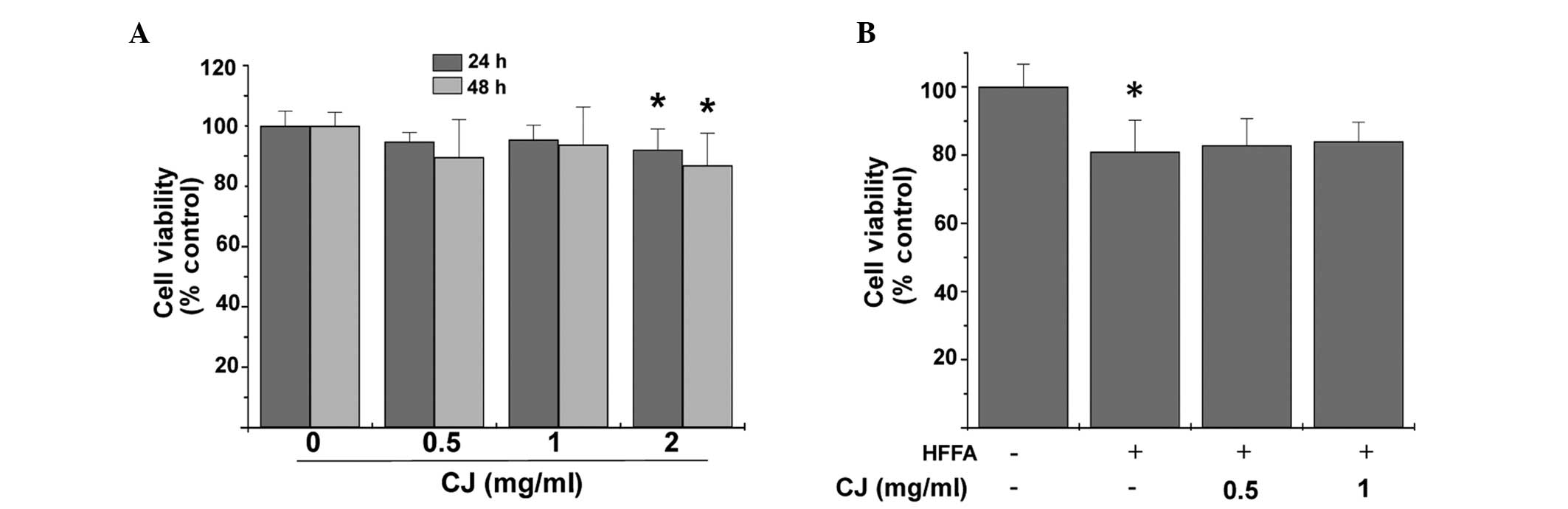
CJ effects on cell viability
The concentration dependence of the potential
cytotoxicity of CJ in HepG2 cells in the absence or presence of 1
mmol/l FFA for 24 h was firstly determined using an MTT assay. CJ
exhibited between 1.7 and 12.4% cytotoxicity in HepG2 cells at the
0–2 mg/ml concentration range when incubated for 24 and 48 h, as
shown in Fig. 1A. When treated
with HFFA for 24 h to induce conditions of hepatic steatosis, 14.3%
cytotoxicity was observed in the cells. Nevertheless, no
significant cytotoxicity was imparted by CJ, as demonstrated in
Fig. 1B.
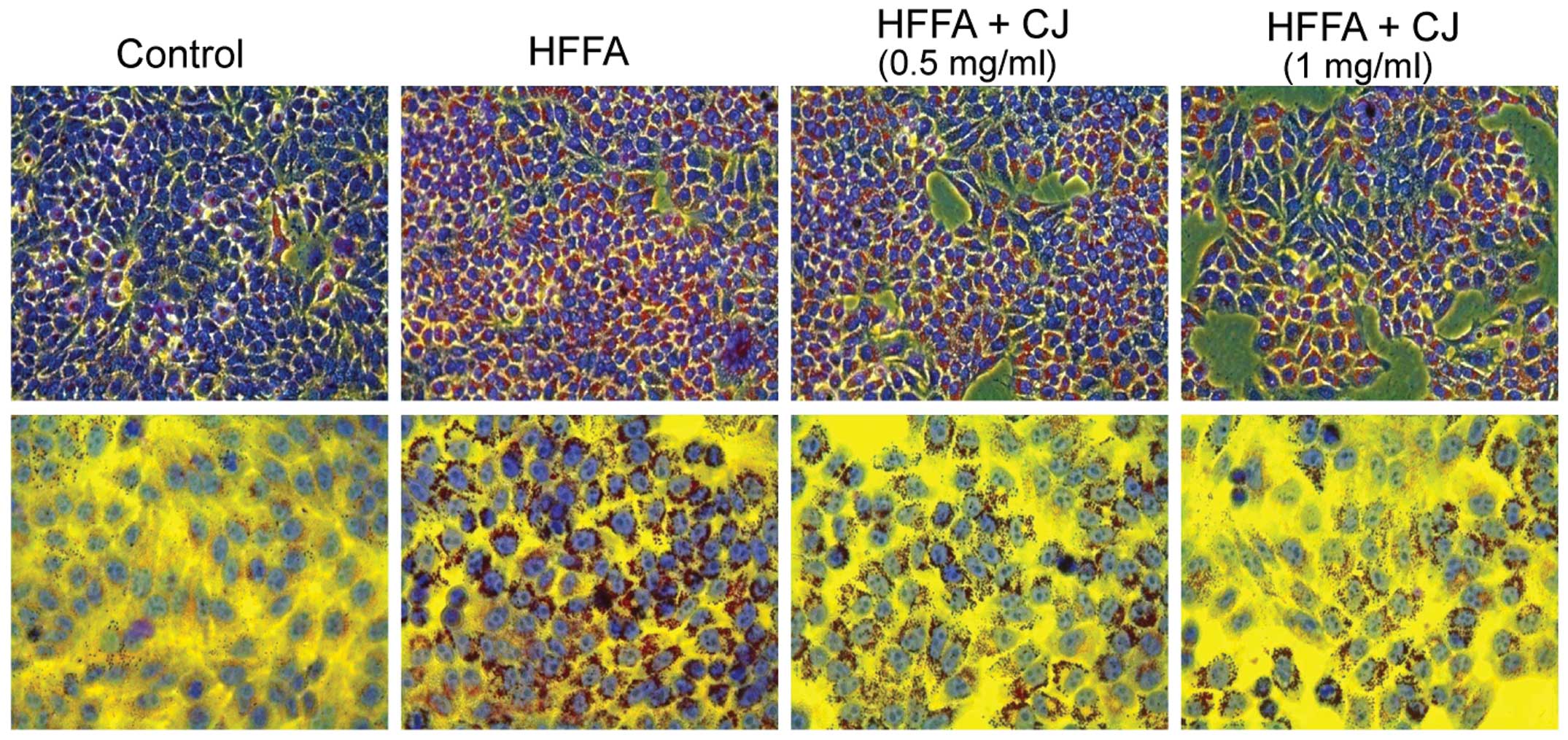
CJ decreases lipid accumulation in
HFFA-induced steatotic HepG2 cells
Microscopic examination revealed that HepG2 cells
treated with HFFA exhibited significant morphological changes in
lipid droplet formation. When CJ was simultaneously administered
for 24 h, hepatic lipid accumulation significantly decreased
(Fig. 2).
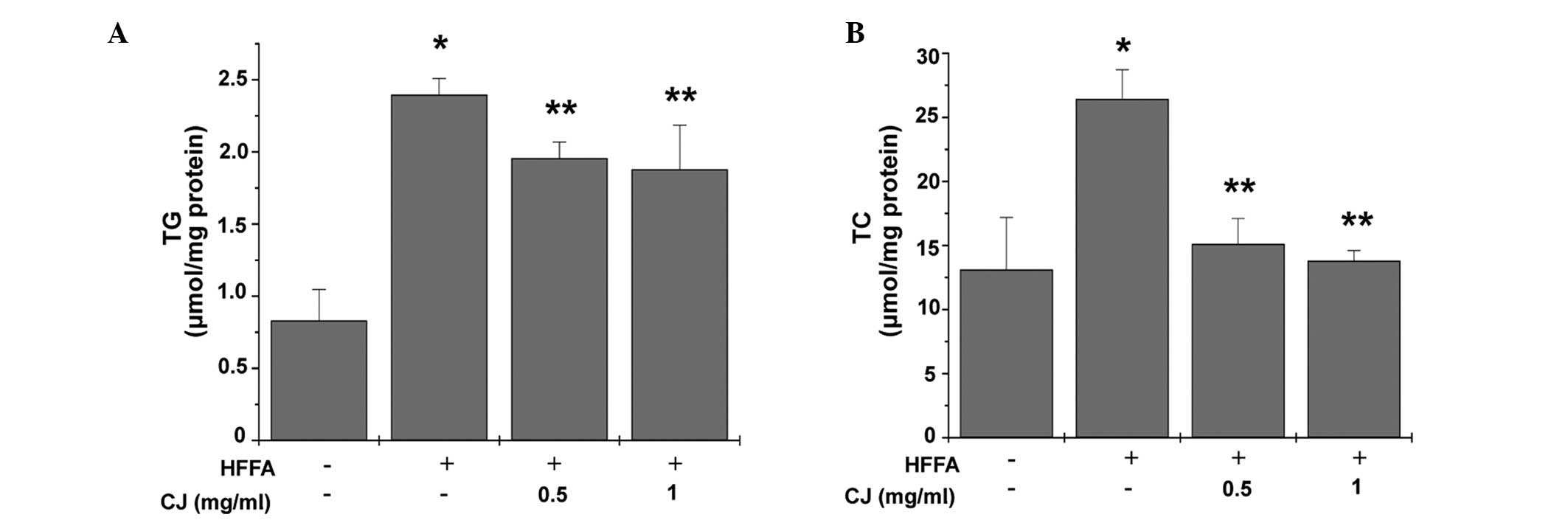
CJ decreases intracellular TG and TC
levels
Following stimulation with FFA for 24 h,
intracellular TG and TC levels significantly increased (Fig. 3). However, upon treatment with CJ,
TC and TG levels decreased significantly.
Effect of CJ on the expression levels of
genes associated with lipid metabolism
Relative mRNA expression levels of lipid metabolism
markers were determined using quantitative PCR. FASN and ACC mRNA
expression levels were shown to increase relative to β-actin mRNA
expression in the HFFA-treated cells (Fig. 4). CJ exhibited an inhibitory effect
on the mRNA expression levels of FASN and ACC in the HepG2 cells.
By contrast, mRNA expression levels of PPAR-α and CPT-1, regulators
of lipolysis and fatty acid transport, were significantly elevated
when HepG2 cells were treated with CJ in a concentration-dependent
manner.
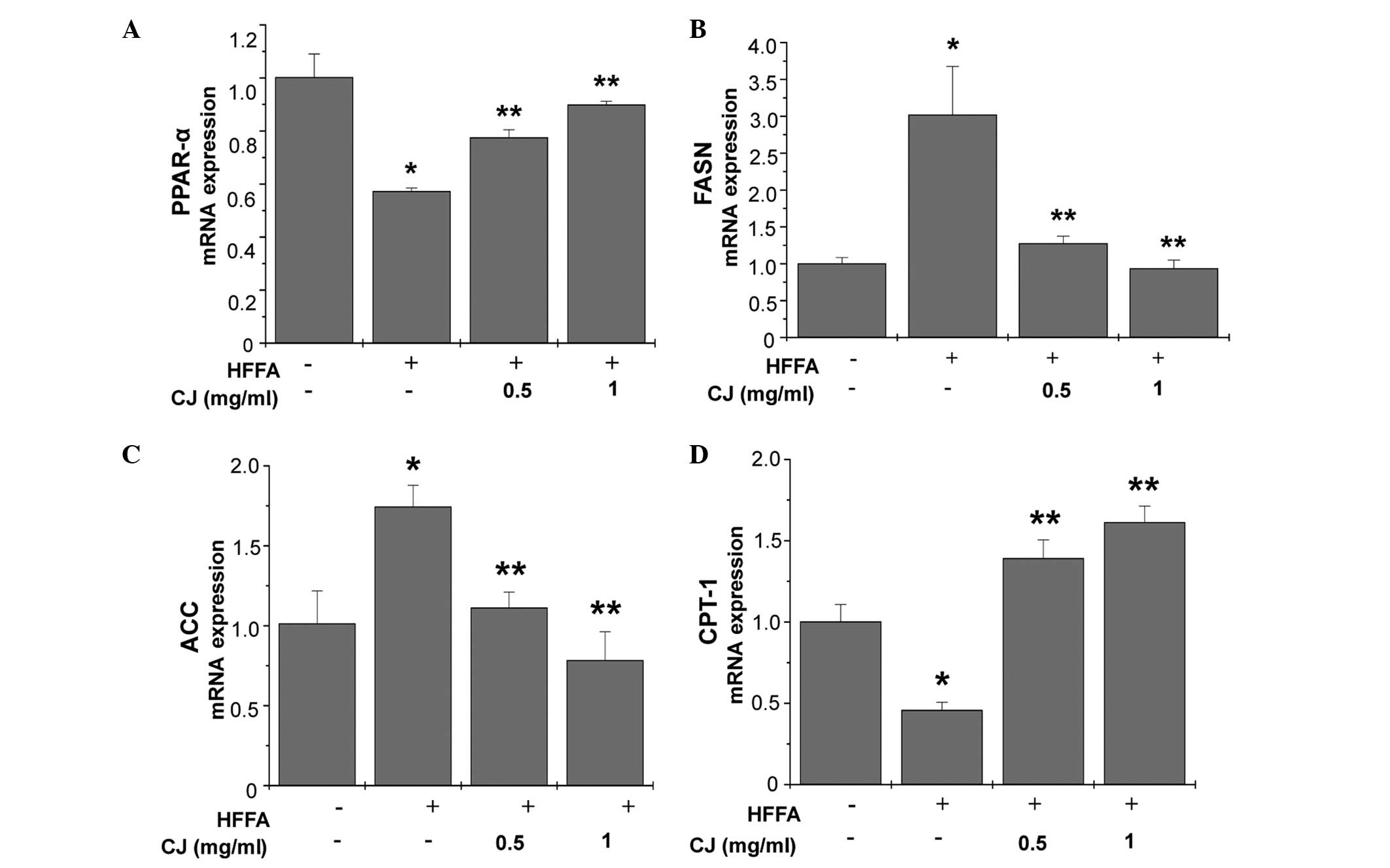 | Figure 4Effects of CJ on lipid
metabolism-associated genes in HepG2 cells. Cells were treated with
CJ in a concentration-dependent manner for 24 h and (A) PPAR-α, (B)
FASN, (C) ACC and (D) CPT-1 mRNA expression levels were determined.
HFFA significantly increased the mRNA expression levels of ACC and
FASN, while CJ significantly reversed these increases. By contrast,
HFFA significantly downregulated the mRNA expression levels of
PPAR-α and CPT-1. The effect of CJ cotreatment was a significant
reversal and increase in these specific mRNA expression levels in
HepG2 cells. Each bar represents the mean ± SE of three independent
experiments. *P<0.05, vs. control;
**P<0.05, vs. HFFA group. CJ, Cirsium
japonicum; HFFA, free fatty acid; PPAR-α, peroxisome
proliferator-activated receptor-α; CPT-1 carnitine
palmitoyltransferase-1; ACC, acetyl-CoA carboxylase; FASN, fatty
acid synthase. |
CJ inhibits hepatic lipid accumulation
via the activation of the AMPK signaling pathway
To investigate whether CJ-mediated lipid reduction
proceeded via AMPK signaling, the Thr172-phosphorylation status of
AMPK, an essential marker of AMPK activity, was determined. As
shown in Fig. 5, CJ treatment
significantly increased AMPK phosphorylation, as well as the
phosphorylation of its direct substrate, ACC, in a
concentration-dependent manner. To determine whether AMPK
activation affected the downstream target genes, the products of
ACC activity were investigated due to their essential role in the
regulation of fatty acid metabolism. Consistent with the gene
expression profiles, CJ was shown to increase the protein
expression levels of CPT-1 and PPAR-α, which are associated with
fatty acid oxidation. In addition, CJ-facilitated downregulation of
FASN, one of the key enzymes involved in lipid synthesis, was
observed, which can catalyze acetyl coenzyme A and malonyl coenzyme
A synthesis of TGs.
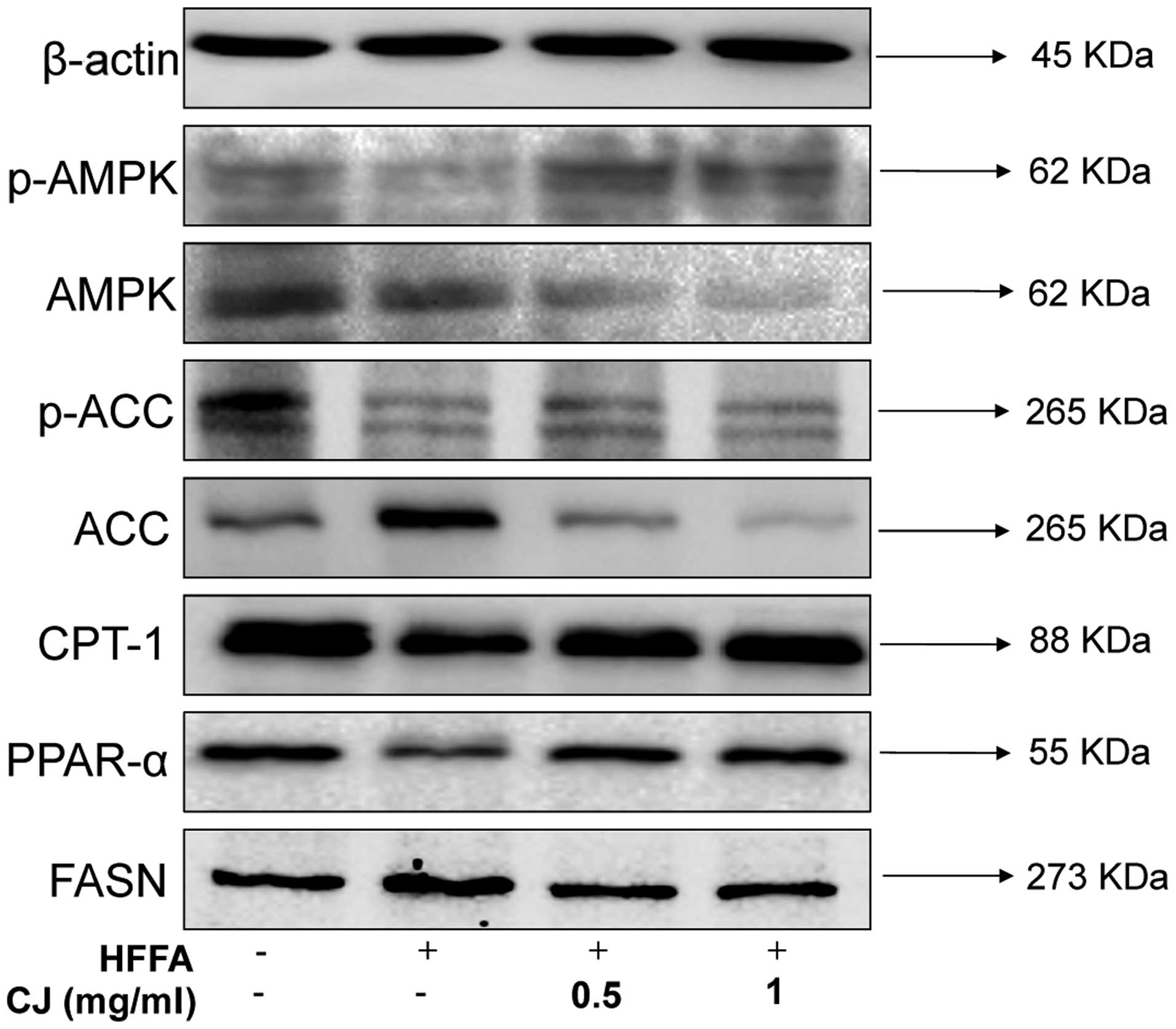 | Figure 5Representative western blot analysis
of AMPK and ACC phosphorylation and downstream protein expression
levels in HepG2 cells. Cells were cultured with or without 1.00
mmol/l HFFA (OA:PA, 2:1) in the presence or absence of CJ at 0.5 or
1 mg/ml for 24 h. The blots shown are representative of three
independent experiments. CJ, Cirsium japonicum; HFFA, free
fatty acid; AMPK, AMP-activated protein kinase; ACC, acetyl-CoA
carboxylase; CPT-1, carnitine palmitoyltransferase-1; PPAR-α,
proliferator-activated receptor-α; FASN, fatty acid synthase; OA,
oleic acid; PA, palmitic acid. |
Discussion
NAFLD is the most common symptomless liver disease
worldwide, characterized by steatosis or fat accumulation in the
liver (14). It is estimated that
the prevalence of NAFLD in the general USA population is ~20% with
anticipated annual increases (15). Despite the high prevalence of NAFLD
and its potential for serious liver damage, currently available
therapies based on the modifications of diet and physical activity
(16) may be insufficient to
address the disease in all patients. Natural products, including
traditional Chinese medicines (TCMs), have been applied clinically
for the treatment of various metabolic syndromes and diseases,
including NAFLD. The general effects of such therapies are
hypothesized to result from the broad targeting of multiple
molecular pathways. In addition, TCMs have relatively few adverse
side effects. In this category of herbal therapies, CJ is a
traditional Chinese perennial herb with a number of known
pharmacological applications. The main constituents of CJ include
flavonoids, essential oils, triterpene and sterol. Although CJ has
a long history of use in TCM, the molecular mechanisms underlying
the biological effects have been understudied.
AMPK, an important metabolic regulator, functions as
a ‘master switch’ by phosphorylating specific target enzymes
(17). Activation of AMPK in the
liver results in the stimulation of fatty acid oxidation and the
inhibition of lipogenesis, glucose production and protein
synthesis. In addition, liver AMPK activation causes the direct
phosphorylation and inactivation of ACC, which leads to decreased
conversion of acetyl-CoA to malonyl-CoA and downregulated fatty
acid synthesis (18).
Alternatively, AMPK positively regulates fatty acid oxidation by
activating PPAR-α and PPAR-γ coactivator-1 (19). In light of these effects,
identifying pharmacological agents that stimulate AMPK activity in
hepatocytes may provide effective treatment options for fatty liver
disease.
A number of herbal medicines, including resveratrol,
Artemisia scoparia and berberine, have demonstrated efficacy
in treating and preventing obesity and associated metabolic
disorders via the regulation of AMPK activity (20–22).
Considering the key role of AMPK activation in regulating lipid
metabolism, we hypothesized that AMPK may play a vital role in
mediating the effects of CJ against hepatic lipid accumulation.
In the present study, HepG2 cells were supplemented
with pathophysiological levels of HFFA to mimic the influx of
excess FFAs in hepatocytes, with the result of hepatic steatosis.
Upon establishing this successful hepatic steatosis model, the
observations demonstrated that exposure of HepG2 cells to HFFA
increased the levels of intracellular TG and TC. Addition of CJ
decreased HFFA-stimulated hepatic TG and TC accumulation. To gain a
better understanding of the potential molecular mechanisms
underlying the effects of CJ in the in vitro model, the
influence of CJ on AMPK activation and the associated genes in
fatty acid metabolism was investigated. CJ triggered an increase in
the phosphorylation of AMPK and its direct substrate, ACC,
indicating that CJ activated AMPK. Furthermore, CJ treatment
modulated the expression levels of genes associated with lipid
metabolism, including FASN, PPAR-α and CPT-1. To the best of our
knowledge, the results of the present study are the first to
hypothesize that CJ may serve as a therapeutic tool to attenuate
hepatic steatosis by targeting the hepatic AMPK system.
In conclusion, the present study demonstrated that
CJ treatment reduced the levels of TC and TG in steatotic HepG2
cells and promoted the phosphorylation of AMPK and ACC. From these
results, we hypothesize that a major mechanism underlying the
beneficial effects of CJ in reducing hepatic lipid accumulation is
via activating AMPK. Thus, CJ possesses great potential for
preventing NAFLD and nonalcoholic steatohepatitis. Future research
is likely to provide further insight into the benefits and
mechanisms of CJ in the treatment of NAFLD and other liver
diseases.
References
|
1
|
Koo SH: Nonalcoholic fatty liver disease:
molecular mechanisms for the hepatic steatosis. Clin Mol Hepatol.
19:210–215. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gentile CL and Pagliassotti MJ: The role
of fatty acids in the development and progression of nonalcoholic
fatty liver disease. J Nutr Biochem. 19:567–576. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kaila B and Raman M: Obesity: a review of
pathogenesis and management strategies. Can J Gastroenterol.
22:61–68. 2008.PubMed/NCBI
|
|
4
|
Angulo P: Obesity and nonalcoholic fatty
liver disease. Nutr Rev. 65:S57–S63. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marra F, Gastaldelli A, Svegliati Baroni
G, Tell G and Tiribelli C: Molecular basis and mechanisms of
progression of non-alcoholic steatohepatitis. Trends Mol Med.
4:72–81. 2008. View Article : Google Scholar
|
|
6
|
Parekh S and Anania FA: Abnormal lipid and
glucose metabolism in obesity: implications for nonalcoholic fatty
liver disease. Gastroenterology. 132:2191–2207. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gómez-Lechón MJ, Donato MT,
Martínez-Romero A, Jiménez N, Castell JV and O’Connor JE: A human
hepatocellular in vitro model to investigate steatosis. Chem Biol
Interact. 165:106–116. 2007.PubMed/NCBI
|
|
8
|
Ricchi M, Odoardi MR, Carulli L, et al:
Differential effect of oleic and palmitic acid on lipid
accumulation and apoptosis in cultured hepatocytes. J Gastroenterol
Hepatol. 24:830–840. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stapleton D, Mitchelhill KI, Gao G, et al:
Mammalian AMP-activated protein kinase subfamily. J Biol Chem.
271:611–614. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Misra P: AMP activated protein kinase: a
next generation target for total metabolic control. Expert Opin
Ther Targets. 12:91–100. 2008.PubMed/NCBI
|
|
11
|
Viollet B, Mounier R, Leclerc J, Yazigi A,
Foretz M and Andreelli F: Targeting AMP-activated protein kinase as
a novel therapeutic approach for the treatment of metabolic
disorders. Diabetes Metab. 33:395–402. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saha AK, Avilucea PR, Ye JM, Assifi MM,
Kraegen EW and Ruderman NB: Pioglitazone treatment activates
AMP-activated protein kinase in rat liver and adipose tissue in
vivo. Biochem Biophys Res Commun. 314:580–585. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Park JC, Hur JM, Park JG, Kim SC, Park JR,
Choi SH and Choi JW: Effects of methanol extract of Cirsium
japonicum var. ussuriense and its principle,
hispidulin-7-O-neohesperidoside on hepatic alcohol-metabolizing
enzymes and lipid peroxidation in ethanol-treated rats. Phytother
Res. 18:19–24. 2004. View
Article : Google Scholar
|
|
14
|
Clark JM, Brancati FL and Diehl AM:
Nonalcoholic fatty liver disease. Gastroenterology. 122:1649–1657.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ruhl CE and Everhart JE: Epidemiology of
nonalcoholic fatty liver. Clin Liver Dis. 8:501–519. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ha SK, Kim J and Chae C: Role of
AMP-activated protein kinase and adiponectin during development of
hepatic steatosis in high-fat diet-induced obesity in rats. J Comp
Pathol. 145:88–94. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Federico A, Niosi M, Vecchio Blanco CD and
Loguercio C: Emerging drugs for nonalcoholic fatty liver disease.
Expert Opin Emerg Drugs. 13:145–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Viollet B, Foretz M, Guigas B, et al:
Activation of AMP-activated protein kinase in the liver: a new
strategy for the management of metabolic hepatic disorders. J
Physiol. 574:41–53. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee WJ, Kim M, Park HS, et al: AMPK
activation increases fatty acid oxidation in skeletal muscle by
activating PPARalpha and PGC-1. Biochem Biophys Res Commun.
340:291–295. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lagouge M, Argmann C, Gerhart-Hines Z, et
al: Resveratrol improves mitochondrial function and protects
against metabolic disease by activating SIRT1 and PGC-1alpha. Cell.
127:1109–1122. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang ZQ, Zhang XH, Yu Y, et al:
Artemisia scoparia extract attenuates non-alcoholic fatty
liver disease in diet-induced obesity mice by enhancing hepatic
insulin and AMPK signaling independently of FGF21 pathway.
Metabolism. 62:1239–1249. 2013. View Article : Google Scholar
|
|
22
|
Quan HY, Kim do Y, Kim SJ, Jo HK, Kim GW
and Chung SH: Betulinic acid alleviates non-alcoholic fatty liver
by inhibiting SREBP1 activity via the AMPK-mTOR-SREBP signaling
pathway. Biochem Pharmacol. 85:1330–1340. 2013. View Article : Google Scholar : PubMed/NCBI
|