Introduction
Peroxisome proliferator-activated receptor δ (PPARδ)
is a ubiquitously expressed member of the ligand-activated nuclear
receptor superfamily. PPARδ has not only been shown to regulate
genes involved in lipid and glucose homeostasis but also seems to
play an important role in inflammation and innate immunity
(1,2). Upon ligand binding, PPARδ is
activated and releases the anti-inflammatory transcriptional
suppressor protein B cell lymphoma-6 (BCL-6), which leads to
suppression of inflammatory gene expression (3,4).
Furthermore, activation of PPARδ has been shown to inhibit tumor
necrosis factor α (TNFα)-induced nuclear factor of κ light
polypeptide gene enhancer in B-cells 1 (NF-κB) activation (5). PPARδ has lately attracted attention
for its involvement in different macrophage phenotypes in mice
where it has been reported to induce a switch from pro-inflammatory
M1 macrophages to the anti-inflammatory M2 phenotype in
metabolically important organs such as liver and adipose tissue
(6,7). The pro-inflammatory M1 macrophage
phenotype is driven by Th1 cytokines e.g., interferon γ (IFNγ) and
TNFα while the anti-inflammatory M2 macrophage phenotype is driven
by Th2 cytokines e.g., interleukin 4 (IL-4) and IL-13. During
atherosclerosis monocytes infiltrate the vessel wall, where these
two subpopulations of macrophages can be detected (8).
Bone marrow-specific deletion of Ppard in
mice renders adipose tissue and liver macrophages incapable of
transition to the M2 phenotype, which in turn causes inflammation
and metabolic derangement in adipocytes as well as hepatic
dysfunction and systemic insulin resistance, respectively (6,7).
These data demonstrate the importance of PPARδ for the inflammatory
response and the dependence of PPARδ for maintaining the
alternative macrophage phenotype in mice. In contrast, a recent
human study investigating PPARδ activation during monocyte
differentiation did not detect any increased expression of PPARδ in
IL-4 induced M2 macrophages compared to untreated macrophages
(9). While research on the
function of PPARδ has mainly focussed on its activation,
considerably less is known about the regulation of expression of
human PPARδ. In the last few years microRNAs (miRNAs) have emerged
as an important class of fine-tuning regulators of gene and protein
expression. These transcripts are endogenous non-coding
single-stranded RNAs ~22 nucleotides in length that bind to the
3′-untranslated region (3′-UTR) of their target mRNA and suppress
expression either by inducing mRNA degradation or inhibiting
protein translation (10). It is
known that ~50% of all mammal protein-coding genes are directly
regulated by miRNAs (11) and
there are over two thousand known human miRNAs described today.
MiRNAs are implicated in processes such as development and
differentiation but many of them have also been reported to be
aberrantly expressed in different forms of cancers. Computational
studies have shown that recurrent networks exist consisting of
specific transcription factors and specific miRNAs that both appear
to regulate one another, thus coupling transcriptional and
post-transcriptional regulation in order to create flexible
expression (12). Identified
miRNAs that have been shown to play key roles in monocytes and/or
macrophages during inflammation include microRNA-9 (miR-9) and
microRNA-155 (miR-155). MiR-9 is involved in the immune response by
fine tuning the expression of a key member of the NF-κB family in
monocytes and polymorphonuclear neutrophils (4) while the expression of miR-155 is
increased during inflammation and has been implicated in macrophage
polarization, where miR-155 modulates the switch between
pro-inflammatory M1 and anti-inflammatory M2 phenotypes. MiR-155
directly inhibits the expression of IL-13 receptor α1 and thereby
downregulates the anti-inflammatory IL-13 pathway in macrophages
(13).
Of the human PPARs, both PPARα and PPARγ have been
shown to be regulated by miRNAs (14–19), but until now no data are available
regarding the regulation of PPARδ by miRNAs. Considering the
important role that PPARδ plays in immunity, we explored the
regulation of human PPARδ expression by miRNAs expressed in primary
human monocytes during the inflammatory response and during
macrophage polarization to M1 and M2 phenotypes, respectively.
Materials and methods
Bioinformatic sequence analysis
Prediction of miRNA binding to the PPARD 3′-UTR was
performed by computer-aided algorithms obtained from TargetScan
(http://www.targetscan.org), PicTar
(http://pictar.mdc-berlin.de), miRanda
http://www.microrna.org) and miRWalk (http://www.umm.uniheidelberg.de/apps/zmf/mirwalk).
Approximately 3 kb of the miR-9 promoter was analyzed for putative
PPREs using the MatInspector software (http://www.genomatix.de).
Reporter gene constructs
Firefly luciferase reporter plasmids containing the
3′-UTR of the PPARδ gene and empty luciferase vector were obtained
from Promega (Madison, WI, USA). The 36 nucleotides of PPARδ 3′-UTR
containing the miR-9 binding site were cloned into psiCHECK-2
(Promega) using the restriction sites XhoI/NotI. The
following oligonucleotides were utilized (the mutated sequences are
shown in bold): miR-9WT-forw: 5′-tcgaTGTCTTCAGAGCAAAAGACTT
GAGCCATCCAAAGAA-3′; miR-9WT-rev: 5′-ggccTTCTTT
GGATGGCTCAAGTCTTTTGCTCTGAAGACA-3′; PPARδ-mir9Mut-forw:
5′-tcgaTGTCTTCAGAGCAAAAGACTTGA GCCATTAGCGTAA-3′;
PPARδ-mir9Mut-rev: 5′-ggccTTAC
GCTAATGGCTCAAGTCTTTTGCTCTGAAGACA-3′. The plasmids were
sequenced and purified using the Endofree Plasmid Maxi kit (Qiagen,
Düsseldorf, Germany).
Cell culture and transfection
HEK293 cells were cultured in DMEM Glutamax (1 g/ml
glucose, Invitrogen) containing 10% FBS, penicillin (100 U/ml) and
streptomycin (100 μg/ml) at 37°C in 5% CO2. For
luciferase assays, HEK293 cells were plated in 24-well
tissue-culture dishes 24 h prior to transfection at a density of
80,000 cells per well. Cells were transfected with the luciferase
reporters, 50 ng per well (Promega), together with pre-miR-9, 10 nM
per well, or miRNA mimics negative control no. 1 (pre-miR-CON,
Ambion, Foster City, CA, USA), 10 nM per well; 50 nM LNA-based
anti-miR-9 (Exiqon, Vedbaek, Denmark) or 50 nM universal LNA-based
negative control (anti-miR-CON) (Exiqon). All transfections were
carried out in triplicates with Lipofectamine 2000 (Invitrogen,
Carlsbad, CA, USA). Cells were lysed 24 h post-transfection using
passive lysis buffer (Promega). Luciferase activity was determined
using Dual Luciferase® Reporter assay system (Promega)
following the manufacturer’s instructions. Relative luciferase
activity was determined by the ratio of renilla luciferase signal
intensity to that of firefly luciferase for normalization.
Human monocytes were isolated from buffy coats as
previously described (20). In
brief, human peripheral blood mononuclear cells (PBMCs) were
isolated from buffy coats by endotoxin-free Ficoll density gradient
centrifugation. Monocytes were then separated from lymphocytes by
high-density hyper-osmotic Percoll density gradient centrifugation
and separated from platelets and dead cells on a low-density
iso-osomotic Percoll density gradient. Monocytes were cultured in
RPMI-1640 medium (Invitrogen) supplemented with
penicillin-streptomycin, L-glutamine (2.05 mM) and 10% human
AB-serum (Invitrogen). Monocytes were transfected with miRNA
oligonucleotides as described above.
Monocytes were transfected with 50 nM anti-miR-9 or
50 nM anti-miR-CON (Exiqon) using Lipofectamine RNAiMAX
(Invitrogen), following the manufacturer’s instructions. At 24 h
after transfection, monocytes were stimulated with LPS (100 ng/ml)
for 4 h.
Pro-inflammatory M1 and anti-inflammatory M2
macrophages were obtained by stimulating freshly isolated monocytes
with recombinant human IFN-γ (20 ng/ml) and IL-4 (15 ng/ml) for 7
days, respectively. The human embryonic kidney cell line HEK293 was
purchased from ATCC (Rockville, MD, USA). The cells were cultured
in Dulbecco’s modified Eagle’s medium (1 g/ml glucose, Invitrogen)
supplemented with 10% newborn calf serum, 100 U/ml penicillin and
100 μg/ml streptomycin (Invitrogen). All the cells were cultured at
37°C in 5% CO2. All cytokines were purchased from
Peprotech (Rocky Hill, NJ, USA). GW501516 was synthesized by
Synthelec AB, Sweden as described (21).
RNA extraction, reverse transcription and
quantitative real-time PCR
Total RNA was prepared using miRNeasy kit (Qiagen)
according to the manufacturer’s instructions. The RNA concentration
was determined by spectrophotometry. Total RNA (0.5 μg) was reverse
transcribed (RT) into cDNA in a 20-μl reaction by a poly-dT primer
using Superscript III™ (Invitrogen).
Quantification of miRNAs by TaqMan®
real-time PCR was carried out as described by the manufacturer
(Applied Biosystems, Foster City, CA, USA). Briefly, 10 ng of
template RNA was reverse transcribed using the TaqMan MicroRNA
Reverse Transcription kit and miRNA-specific stem-loop primers
(Applied Biosystems). RT product (1.5 μl) was introduced into 20-μl
PCR reactions which were incubated in 96-well plates on the ABI
PRISM® 7900HT Sequence Detection System (Applied
Biosystems) at 95°C for 10 min, followed by 40 cycles of 95°C for
15 sec and 60°C for 1 min. Target gene expression was normalized
between different samples based on the values of U48 RNA
expression.
The cDNA was amplified by real-time PCR as described
(22). For the quantification, 15
ng of cDNA were amplified per reaction in the presence of TaqMan
universal master mix (Applied Biosystems) and TaqMan Gene
Expression Assays for PPARδ (Hs04187066_g1), PLIN2 (Hs00605340_m1),
CPT1A (Hs00912676_m1), ANGPTL4 (Hs01101127_m1), TNFα
(Hs01113624_g1), STAT1 (Hs01013996_m1), MRC1 (Hs00267207_m1) and
STAT6 (Hs00598625_m1), all purchased from Applied Biosystems.
Gene-specific PCR products were measured by means of the ABI
PRISM® 7900HT Sequence Detection System (Applied
Biosystems). Target gene expression was normalized based on the
values of the expression of cyclophilin A, PPIA (Hs04194521_s1) and
18s (Hs99999901_s1) obtained from Applied Biosystems.
Statistical analysis
Wilcoxon rank-sum and signed-rank tests were
employed to determine statistical differences between means of
quantitative real-time PCR and luciferase assay data. P<0.05 was
considered statistically significant.
Results
PPARδ is the direct target of miR-9
To investigate whether the expression of PPARδ in
monocytes could be regulated by miRNAs during the inflammatory
response and human macrophage polarization, bioinformatic analyses
were performed. We screened the human PPARδ 3′-UTR for putative
target sites of candidate miRNAs expressed in monocytes and/or
macrophages (4,13,23). Four different computational
prediction programs; TargetScan, PicTar, miRanda and miRWalk, were
used. One putative miRNA target site for miR-9 was identified by
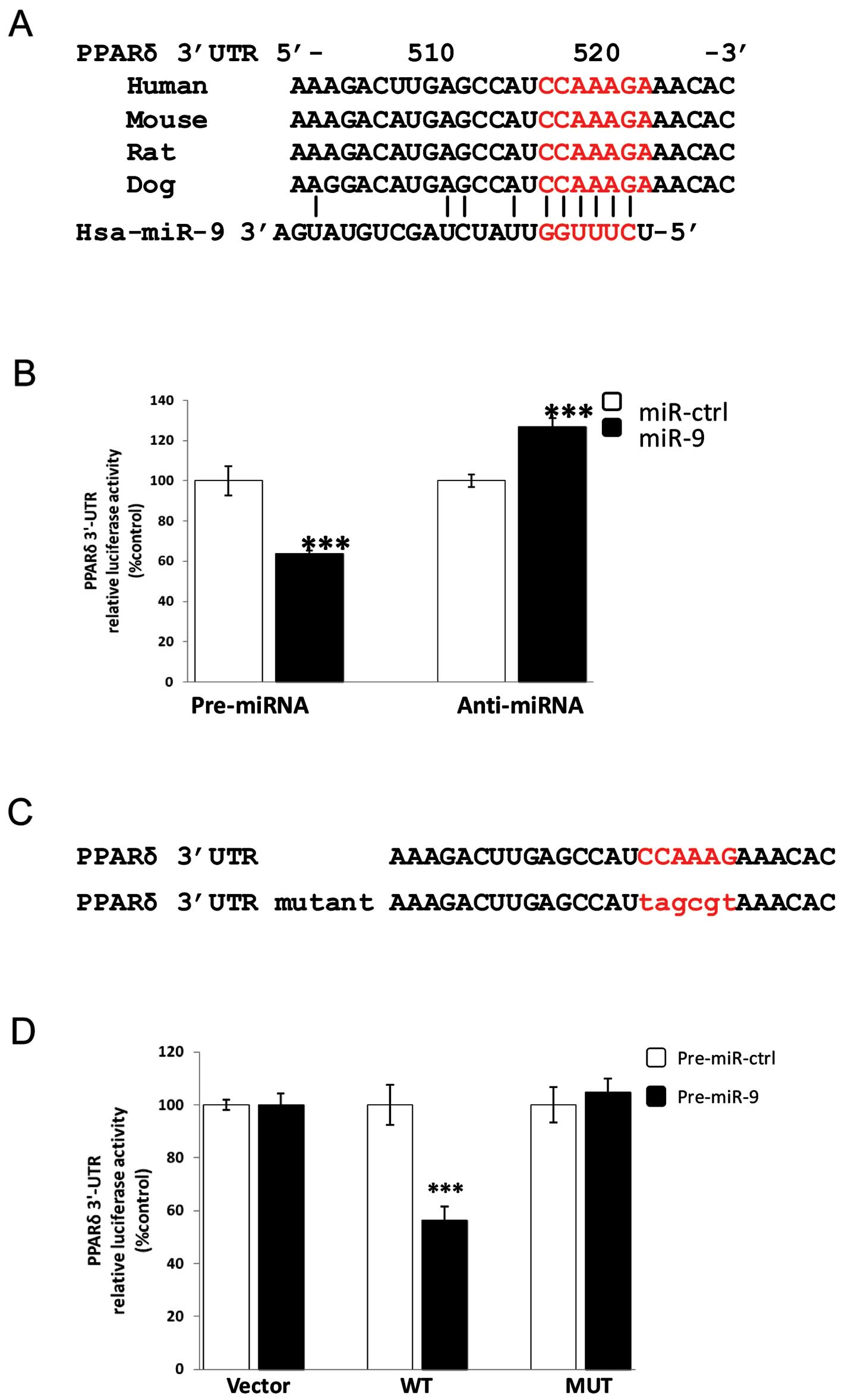
all four programs. The identified target site for miR-9 in PPARδ
3′-UTR is highly conserved in many mammals including human, mouse,
rat and dog (Fig. 1A).
In order to verify whether PPARδ is a direct target
of miR-9, we performed 3′-UTR luciferase activity assays.
Accordingly, 36 nucleotides encompassing the putative miR-9 binding
site in the 3′-UTR of the PPARδ gene were cloned into a reporter
plasmid containing the renilla luciferase gene. This reporter
construct was transiently co-transfected with miR-9 mimic
(pre-miR-9) or the specific inhibitor of miR-9 (anti-miR-9) as well
as their respective control oligonucleotides into HEK293 cells and
relative luciferase activities were determined 24 h after
transfection (Fig. 1B).
Overexpression of miR-9 reduced the luciferase activity down to 64%
(P<0.001), whereas inhibition of endogenous miR-9 increased the
luciferase activity up to 130% (P<0.001) compared with scrambled
pre-miR or anti-miR control nucleotides, respectively (Fig. 1B), indicating that miR-9 directly
targets PPARδ.
To examine whether the effects on transcription are
mediated by the predicted miR-9 target site in the 3′-UTR of PPARδ,
we changed 6 nucleotides within the miR-9 seed-matching sequence of
the 3′-UTR of PPARδ to generate a construct named MUT (Fig. 1C). Mutation of the miR-9
seed-matching sequence led to a complete restoration of luciferase
activity and reversed the inhibitory effect of miR-9 in the 3′-UTR
of PPARδ (Fig. 1D), which shows
that the effects are mediated through the identified miR-9 target
site. Taken together, these results demonstrate that miR-9 directly
regulates PPARδ expression by binding to the target site in the
3′-UTR of PPARδ mRNA.
PPARδ mRNA expression is regulated by
miR-9 in monocytes after LPS treatment
Since miR-9 has been shown to play an important role
in the inflammatory response in monocytes, where it serves as a
feedback controller of inflammation by suppressing NFκB1 signaling
(4), the relevance of miR-9
regulation in relation to PPARδ expression was investigated in
monocytes. To this end, primary monocytes were isolated from PBMC
obtained from healthy donors followed by stimulation with LPS for
different time periods. Expression of miR-9 and PPARδ was evaluated
by qRT-PCR. To estimate the corresponding effects due to changes in
human PPARδ protein expression, we analysed PPARδ target genes,
such as perilipin-2 (PLIN2), carnitine palmitoyltransferase
1α (CPT1A) and angiopoietin-related protein 4
(ANGPTL4), none of which had putative miR-9 target sites as
evaluated by bioinformatic analyses (24).
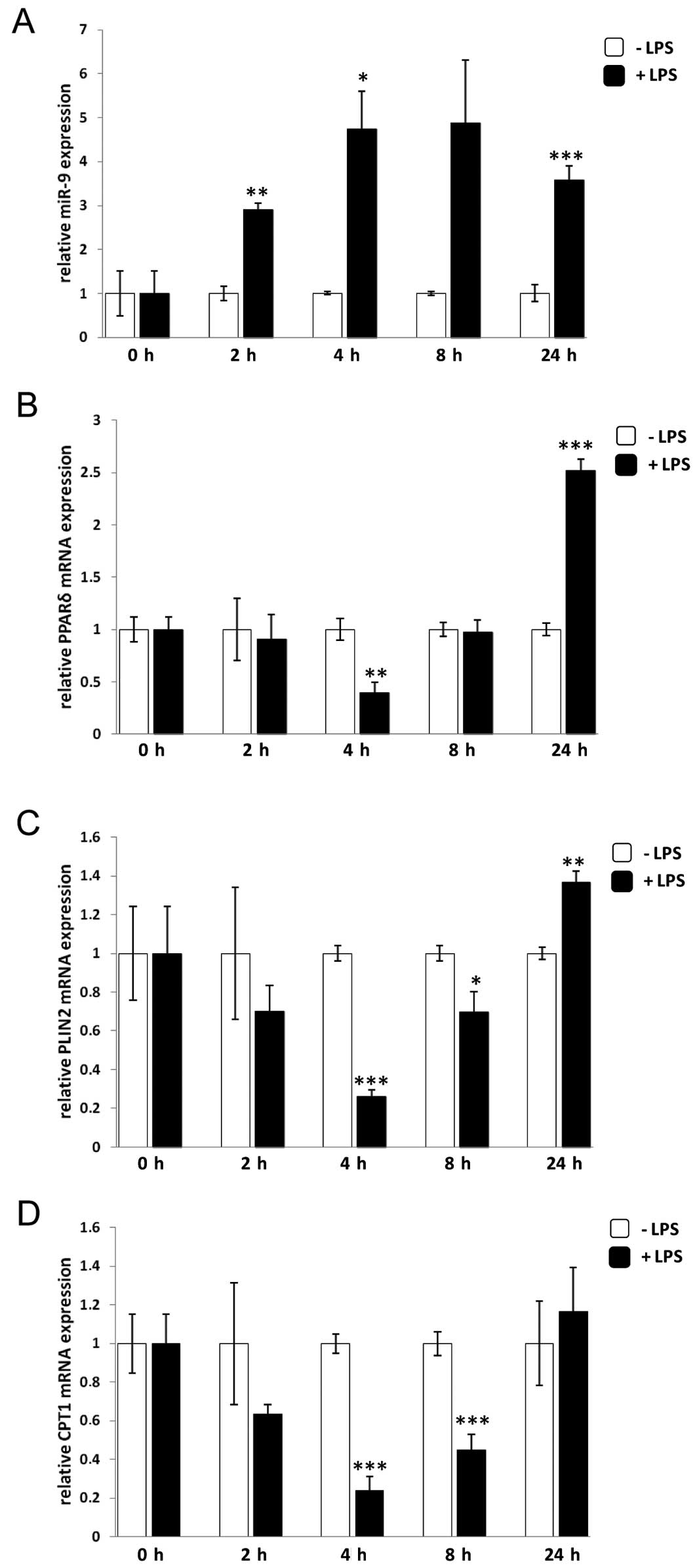
In line with the findings by Bazzoni et
al(4), miR-9 levels increased
rapidly after 2 h and remained increased until 24 h after treatment
with LPS (Fig. 2A). The PPARδ
mRNA level, on the other hand, was unchanged until 4 h after LPS
treatment when it was suppressed by 60% compared to control cells
treated with vehicle only. The suppression was abolished after 8 h
and increased at 24 h of LPS stimulation (Fig. 2B). Analysis of the PPARδ target
genes PLIN2 and CPT1A showed 80 and 76%, respectively, reduced
expression levels compared to unstimulated control cells after 4 h
of LPS treatment and this reduction remained until 8 h, followed by
an increase at 24 h (Fig. 2C and
D). Analysis of another PPARδ target gene, ANGPTL4, showed the
same trend as PLIN2 and CPT1A (data not shown). Thus, PLIN2
was chosen as the PPARδ target gene in the following analyses due
to its relatively high abundance. Taken together, the results
suggest a model in which upregulation of miR-9 upon LPS stimulation
in monocytes results in a decrease in PPARδ expression thereby
suppressing its corresponding target genes at an early time-point
(4–8 h), indicating that PPARδ is regulated by miR-9 in monocytes
after LPS treatment.
Suppression of miR-9 upregulates the mRNA
expression of PPARδ and its target gene PLIN2 in human primary
monocytes
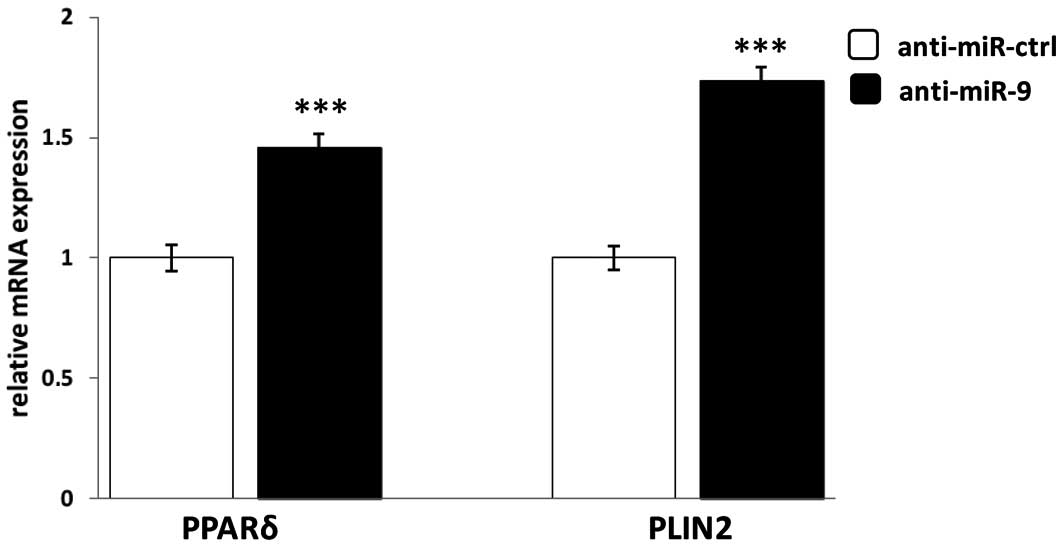
The influence of miR-9 on PPARδ expression in
monocytes during the inflammatory response is unknown. Thus, we
further evaluated regulation by miR-9 of PPARδ expression in human
primary monocytes stimulated with the pro-inflammatory agent LPS.
Human primary monocytes were transfected with the specific
anti-miR-9 or its control oligonucleotides for 24 h, followed by
LPS stimulation for 4 h. In order to measure the effect of miR-9
the mRNA expression of PPARδ and its target gene PLIN2 was
quantified by qRT-PCR. As shown in Fig. 3, specific inhibition of miR-9
significantly increased PPARδ and PLIN2 mRNA expression 40 and 70%,
respectively, compared to scrambled control oligonucleotides
(anti-miR-ctrl). This result suggests that miR-9 downregulates the
expression of PPARδ and its target gene PLIN2 and further confirms
the regulatory link between PPARδ and miR-9.
Both PPARδ mRNA and miR-9 expression are
higher in M1 than in M2 macrophages
PPARδ has been suggested to play a role in the
switch from pro-inflammatory M1 phenotype to anti-inflammatory M2
phenotype in macrophages. To study whether miR-9 expression was
involved in the polarization of M1 and M2 macrophage phenotypes
through modulation of PPARδ expression, human primary monocytes
were differentiated into M1 and M2 macrophages, respectively.
Hence, human primary monocytes were cultured in the presence of
either IFNγ to promote a shift to pro-inflammatory M1 macrophages,
or IL-4 to polarize the macrophages into the anti-inflammatory M2
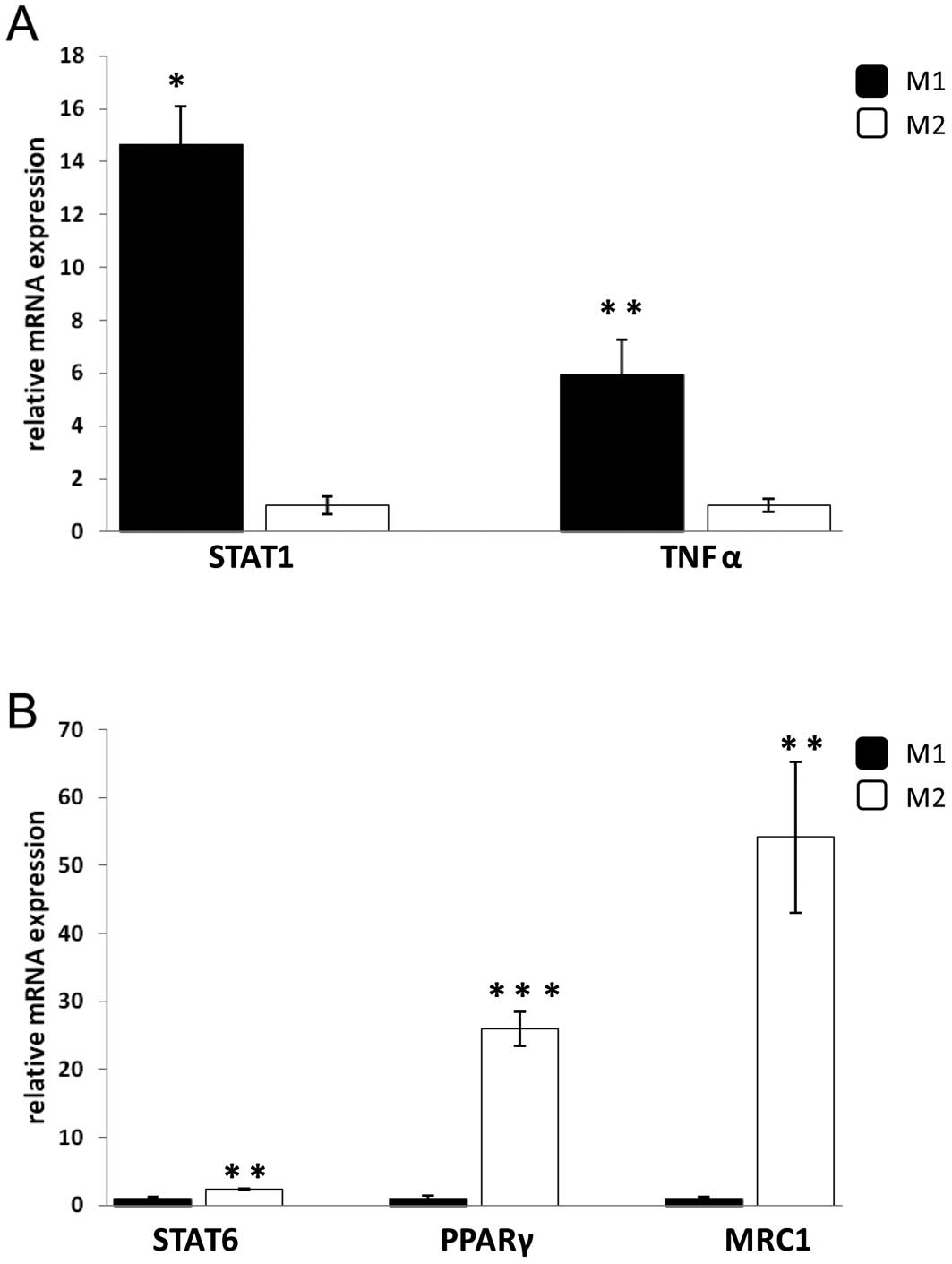
phenotype. As expected the M2 marker, mannose receptor C type 1
(MRC1), showed higher expression in M2 compared with M1 or
untreated macrophages using FACS analysis (data not shown). TaqMan
analysis revealed that the expression of TNFα and signal transducer
and activator of transcription 1 (STAT1), which are markers of M1,
were higher in M1 compared to M2 macrophages while the mRNA
expression of MRC1 and STAT6 were higher in M2 compared to M1
macrophages (Fig. 4). In
addition, PPARγ mRNA was also analysed since its expression has
been shown to correlate with markers for M2 macrophages. As shown
in Fig. 4B, PPARγ mRNA levels
were significantly increased in M2 compared with M1 macrophages.
These results demonstrate that two distinct macrophage phenotypes,
M1 and M2, were obtained.
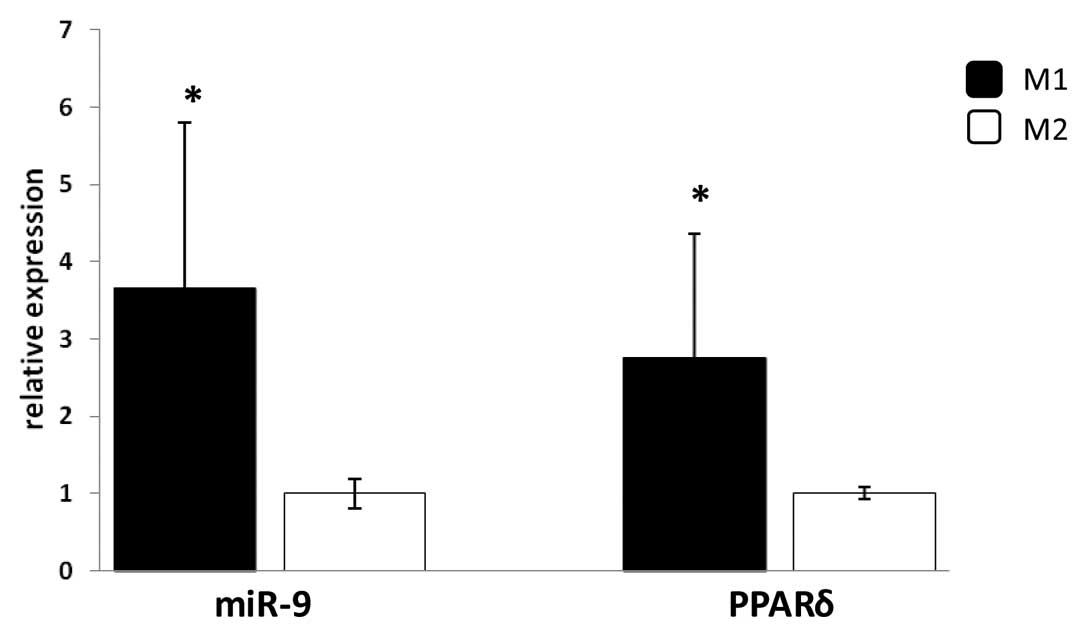
Analyses of the expression of PPARδ mRNA and miR-9
showed that their levels are significantly increased in M1 compared
with M2 macrophages (Fig. 5),
which suggest that PPARδ and miR-9 might be of importance in
modulating the pro-inflammatory M1 human macrophage phenotype.
Expression of miR-9 in relation to PPARδ
agonist treatment
The polarization towards the M1 phenotype is
important for macrophages exposed to pro-inflammatory stress. To
further investigate the interplay between PPARδ and miR-9 in
monocytes and macrophages during the inflammatory response, we
examined whether miR-9 can be regulated by PPARδ. Since pri-mir-9-1
is the only primary miR-9 transcript induced by LPS (4), we set out to analyse whether a PPAR
response element (PPRE) exists in the pri-mir-9-1 promoter region.
In silico analysis of the promoter region of pri-mir-9-1
revealed five putative PPREs within 2.5-kb upstream of the
transcriptional start site (data not shown).
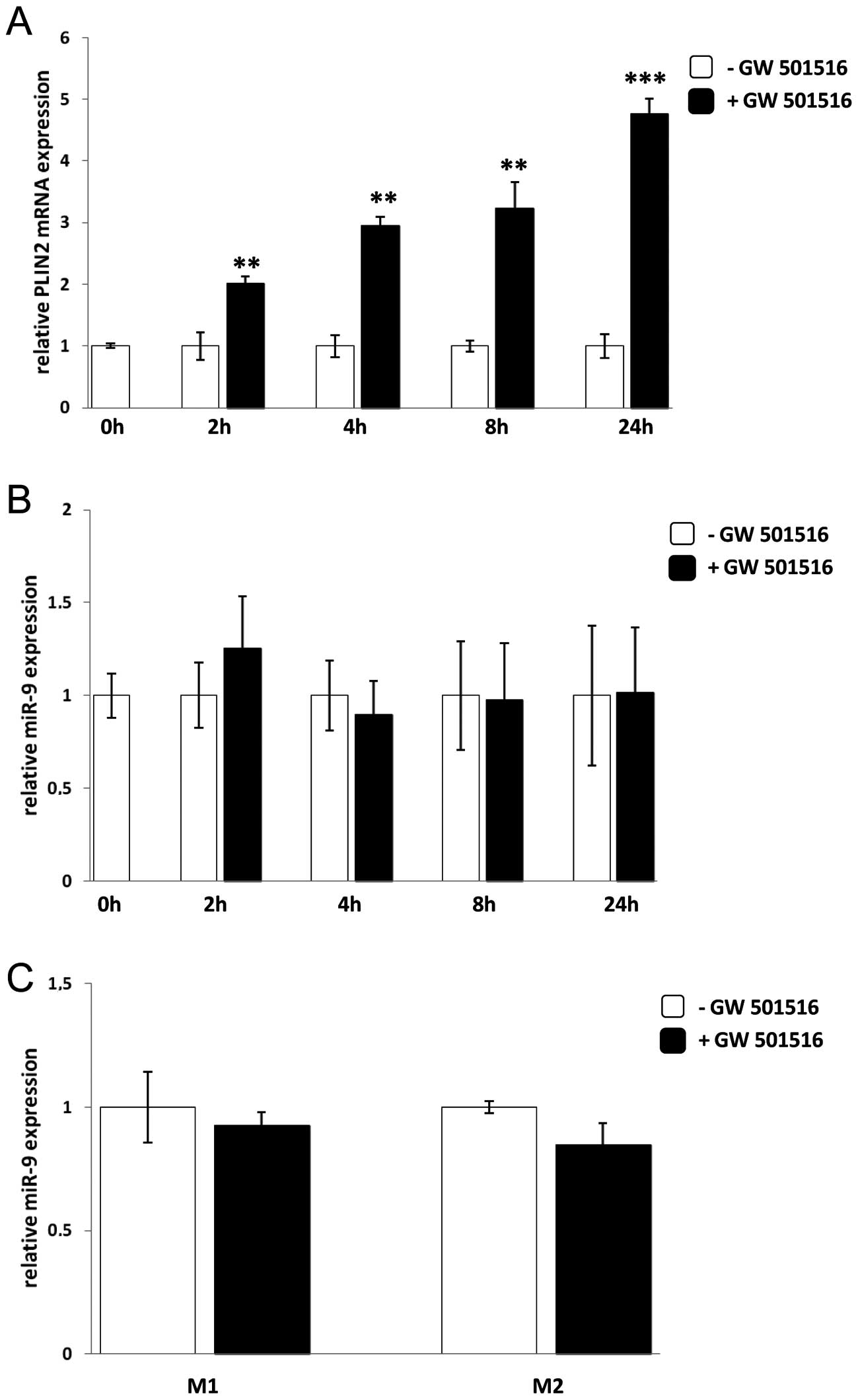
Next, we explored whether PPARδ activation could
induce miR-9 expression in human monocytes and/or macrophages using
the specific PPARδ agonist, GW501516. Human monocytes were isolated
from PBMC and stimulated with GW501516 for 2, 4, 8 and 24 h. Since
PLIN2 gene expression is regulated by PPARδ, PLIN2 mRNA levels were
measured at the different time-points to serve as a positive
control and to ensure that GW501516 had activated PPARδ. As
expected PLIN2 mRNA expression was upregulated at the different
time-points analysed after adding GW501516 (Fig. 6A). However, the miR-9 expression
was not induced at any of the different time-points of treatment
with GW501516 (Fig. 6B), nor was
there any difference of miR-9 expression in M1 or M2 macrophages
after GW501516 treatment (Fig.
6C).
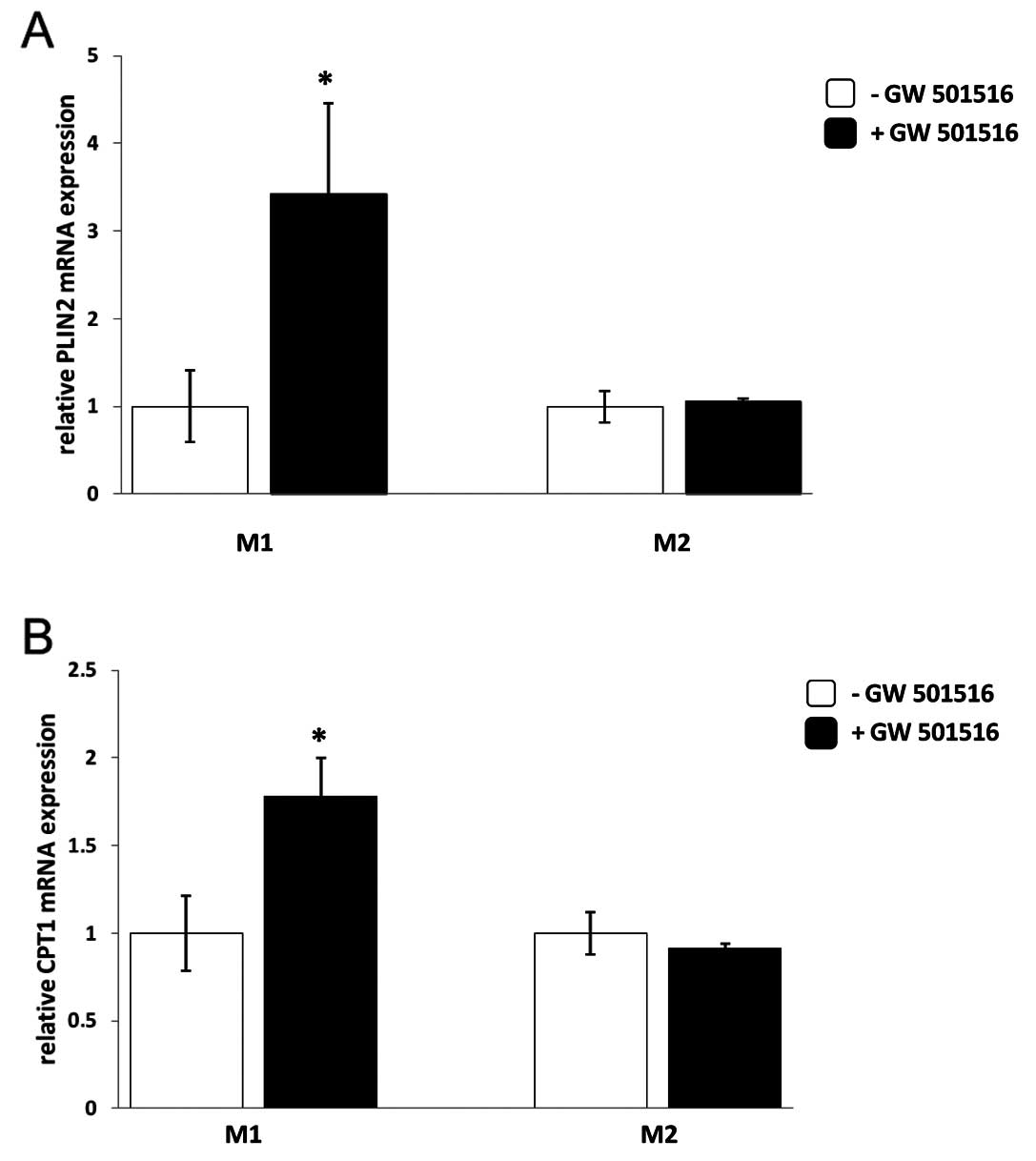
PPARδ agonist treatment results in
upregulation of PPARδ target genes in M1 but not in M2
macrophages
In order to evaluate the influence of PPARδ on the
expression of its target genes, M1 and M2 macrophages were
subjected to GW501516 treatment for 4 h. Human primary monocytes
that were differentiated into macrophages of the pro-inflammatory
M1 phenotype responded to the treatment with GW501516 by
upregulation of PLIN2 and CPT1A mRNA while macrophages
differentiated into the anti-inflammatory M2 phenotype did not
(Fig. 7). Another PPARδ target
gene that was analysed, ANGPTL4, also showed induction of mRNA
expression upon GW501516 treatment only in M1 but not in M2
macrophages (data not shown).
Discussion
It has been suggested that the primary regulation of
the human PPARδ gene is not at the transcriptional level. Instead,
PPARδ is known to be regulated at the post-translational level by
the presence of ligands along with specific cofactors that are of
major importance for PPARδ activation and function. In this study,
we show that PPARδ is also regulated at the post-transriptional
level by miR-9. Upregulation of miR-9 results in the direct
repression of PPARδ, the mRNA levels of which are found to be
higher in pro-inflammatory M1 than in anti-inflammatory M2
macrophages.
MiR-9 has been shown to be of importance during the
immune response (4),
post-traumatic stress (25),
neuronal differentiation (26),
different forms of cancers (27,28) and exocytosis of insulin from
pancreatic islets (29). Here, we
show that miR-9 is upregulated in human monocytes after LPS
treatment while PPARδ and its target genes PLIN2, CPT1A and ANGPTL4
are downregulated 4–8 h after LPS-treatment compared with untreated
cells. The suppression, intriguingly, was abolished after 8–24 h of
LPS stimulation despite the presence of continued high levels of
miR-9. One possible explanation might be that the effect and
activity of miR-9 are influenced due to interactions with
RNA-binding proteins (30).
Another reason might be that LPS triggers other factors that in
turn could affect the expression of PPARδ. Indeed, Tan et
al(31) have shown that
pro-inflammatory cytokines, such as TNFα, can both increase PPARδ
expression via the stress kinase signaling pathway and trigger the
production of ligands for this receptor, which is in agreement with
our findings that PPARδ is upregulated after 24 h of LPS treatment.
The inhibition of PPARδ by miR-9 at the early time-points might be
a mechanism to delay the effect of PPARδ action early in
inflammation to prevent PPARδ from suppression of NF-κB since NF-κB
and PPARδ have been shown to be able to crosstalk and inhibit the
function of each other (32). The
inhibitory effect by miR-9 on PPARδ expression was confirmed by
transfection of anti-miR-9 that sequesters mature miR-9 thus
inhibiting its biologic function into monocytes, which resulted in
induction of PPARδ and PLIN2 mRNA levels.
Hence, the miR-9 mediated inhibition of PPARδ
expression in monocytes may constitute a negative feedback loop,
modulating the levels of the receptor during inflammation. This
finding regarding miR-9 expression in monocytes is in agreement
with a previous study, which showed that the expression of miR-9 is
dramatically increased after treatment with LPS (4). Bazzoni et al showed that the
miR-9 targeting of NF-κB at the mRNA level constitutes a feedback
loop of the inflammatory response. Other studies have confirmed the
regulatory function of miR-9 on the mRNA of NF-κB in both ovarian
and gastric cancers where miR-9 has been shown to act as a tumor
suppressor (33,34). Of note, in the monocytic cell line
THP1, miR-9 is abundantly expressed and there is no induction of
miR-9 upon LPS stimulation (unpublished data), therefore all the
current experiments were carried out in human primary monocytes or
macrophages.
Here, the regulation of PPARδ by miR-9 was examined
in relation to the pro-inflammatory M1 and anti-inflammatory M2
macrophage phenotypes. The role of PPARδ in the pro-inflammatory M1
macrophages has not been investigated before while PPARδ has been
suggested to be of importance in macrophages with the
anti-inflammatory M2 phenotype, both in adipose tissue and liver of
mice (6,7) although this seems not to be the case
in humans (9).
Monocytes were differentiated to macrophages of the
M1 and M2 phenotypes by stimulation with IFNγ and IL-4,
respectively, for 7 days. To confirm this, several markers that are
commonly used to distinguish between the two distinct M1 and M2
macrophage phenotypes were analyzed. In agreement with other
reports the expression of STAT1 and TNFα were higher in M1 than M2
macrophages while STAT6, mannose receptor type C and PPARγ were
higher in M2 than in M1 macrophages (35).
In this study, both the expression of PPARδ and
miR-9 were higher in pro-inflammatory M1 than in anti-inflammatory
M2 macrophages, indicating the potential involvement of PPARδ and
miR-9 in modulating the M1 macrophage phenotype. As expected TNFα
expression was higher in M1 than in M2 macrophages and the PPARδ
ligand, GW501516, did not influence the expression of miR-9 in M1
macrophages. These results indicate that the pro-inflammatory
cytokines are responsible for the induction of both miR-9 and PPARδ
expression, which is in agreement with previous studies (4,36,37). In addition, the expression of
PPARδ target genes was induced by GW501516 in macrophages of the
M1, but not the M2 phenotype, which probably is due to the fact
that PPARδ is mainly expressed in M1 macrophages while PPARγ is
predominantly expressed in M2 macrophages in humans (6–9).
Our data show the regulation of PPARδ by miR-9 in
monocytes. Whether this regulation might play a role in foam cell
formation and initiation of atherosclerosis remains to be
elucidated. However, activation of PPARδ does not seem to promote
the polarization into M2 macrophages (unpublished data). In
contrast to the studies by Kang et al and Odegaard et
al(6,7), our study of human macrophages showed
a higher PPARδ mRNA expression in M1 macrophages compared to M2
macrophages. This result supports the data from Bouhlel et
al(9), who showed that PPARδ
expression was lower in M2 macrophages compared to resting
(untreated) macrophages. These data highlight differences between
human and mouse macrophages and suggest that they may be regulated
by different factors.
The importance of miR-9 in monocytes and macrophages
as a response to proatherogenic factors is further demonstrated by
a recent report showing that miR-9 is significantly upregulated in
monocytes and macrophages after exposure to oxLDL (38). One purpose of the inhibition of
PPARδ by miR-9 in monocytes could be to induce a block in monocyte
expansion. Whether PPARδ inhibits or stimulates cell proliferation
is still debated (39), but
during the inflammatory response cells must focus on repair and
disposal of causal pathogens rather than undergoing cell divisions.
In this context, PPARδ has been shown to function as a sensor of
apoptotic cells in macrophages by promoting the clearance of
apoptotic cells and suppressing autoreactive immune responses
(40).
Importantly, a miRNA does not only target one gene,
but frequently rather modestly regulates a number of genes
belonging to the same network. The long list of putative miR-9
targets identified by computational prediction programs contains
genes working in a close network with PPARδ, for example PPARα,
RXRα, PGC1α, FOXO and BCL-6. If all these genes are miR-9
regulated, the outcome of miR-9 regulation would be due to effects
on the whole network rather than only to PPARδ. Putative target
genes of PPARδ can also be found on the miR-9 target list, such as
ABCA1, ABCD1, GOT1 and PDK4. The effects of targeting these genes
would cause similar results as targeting of PPARδ itself, but
resulting in a larger net effect. To draw firm conclusions,
however, a functional approach for each gene would be required in
order to validate whether these genes are true miR-9 targets, which
although relevant, was outside the scope of this study.
In conclusion, we have identified miR-9 as a
regulator of PPARδ expression in monocytes through direct targeting
of a specific sequence with the 3′-UTR of PPARδ. These data show
that miR-9 and PPARδ are involved in central signalling pathways
during the inflammatory response in monocytes. Interestingly,
activation of PPARδ by GW501516 showed that the PPARδ target genes
expressed in the pro-inflammatory M1 macrophages were induced while
the expression of the same genes were unaffected in the
anti-inflammatory M2 macrophages, which may suggest that PPARδ is
of importance in M1 pro-inflammatory macrophages.
Acknowledgements
This study was supported by the Swedish Research
Council, the Swedish Heart-Lung Foundation, the Swedish Diabetes
Foundation, the Novo Nordisk Foundation, the Swedish Cancer
Foundation, Radiumhemmets Forskningsfonder and the Karolinska
Institutet Funds. P.T. was supported by a doctoral research
fellowship from the Swedish Heart-Lung Foundation and E.E. was
supported by a senior researcher fellowship from the Swedish
Heart-Lung Foundation.
References
|
1
|
Ehrenborg E and Krook A: Regulation of
skeletal muscle physiology and metabolism by peroxisome
proliferator-activated receptor δ. Pharmacol Rev. 61:373–393.
2009.
|
|
2
|
Yessoufou A and Wahli W: Multifaceted
roles of peroxisome proliferator-activated receptors (PPARs) at the
cellular and whole organism levels. Swiss Med Wkly.
140:w130712010.PubMed/NCBI
|
|
3
|
Barish GD, Narkar VA and Evans RM: PPARδ:
a dagger in the heart of the metabolic syndrome. J Clin Invest.
116:590–597. 2006.
|
|
4
|
Bazzoni F, Rossato M, Fabbri M, et al:
Induction and regulatory function of miR-9 in human monocytes and
neutrophils exposed to proinflammatory signals. Proc Natl Acad Sci
USA. 106:5282–5287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barroso E, Eyre E, Palomer X and
Vázquez-Carrera M: The peroxisome proliferator-activated receptor
β/δ (PPARβ/δ) agonist GW501516 prevents TNF-α-induced NF-κB
activation in human HaCaT cells by reducing p65 acetylation through
AMPK and SIRT1. Biochem Pharmacol. 81:534–543. 2011.
|
|
6
|
Kang K, Reilly SM, Karabacak V, et al:
Adipocyte-derived Th2 cytokines and myeloid PPARδ regulate
macrophage polarization and insulin sensitivity. Cell Metab.
7:485–495. 2008.PubMed/NCBI
|
|
7
|
Odegaard JI, Ricardo-Gonzalez RR, Red
Eagle A, et al: Alternative M2 activation of kupffer cells by PPARδ
ameliorates obesity-induced insulin resistance. Cell Metab.
7:496–507. 2008.PubMed/NCBI
|
|
8
|
Chinetti-Gbaguidi G, Baron M, Bouhlel MA,
et al: Human atherosclerotic plaque alternative macrophages display
low cholesterol handling but high phagocytosis because of distinct
activities of the PPARgamma and LXRalpha pathways. Circ Res.
108:985–995. 2011. View Article : Google Scholar
|
|
9
|
Bouhlel MA, Brozek J, Derudas B, et al:
Unlike PPARgamma, PPARalpha or PPARdelta activation does not
promote human monocyte differentiation toward alternative
macrophages. Biochem Biophys Res Commun. 386:459–462. 2009.
View Article : Google Scholar
|
|
10
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chekulaeva M and Filipowicz W: Mechanisms
of miRNA-mediated post-transcriptional regulation in animal cells.
Curr Opin Cell Biol. 21:452–460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dahan O, Gingold H and Pilpel Y:
Regulatory mechanisms and networks couple the different phases of
gene expression. Trends Genet. 27:316–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Martinez-Nunez RT, Louafi F and
Sanchez-Elsner T: The interleukin 13 (IL-13) pathway in human
macrophages is modulated by microRNA-155 via direct targeting of
interleukin 13 receptor alpha1 (IL13Ralpha1). J Biol Chem.
286:1786–1794. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martinelli R, Nardelli C, Pilone V, et al:
miR-519d overexpression is associated with human obesity. Obesity.
18:2170–2176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou J, Wang K-C, Wu W, et al: MicroRNA-21
targets peroxisome proliferator-activated receptor-alpha in an
autoregulatory loop to modulate flow-induced endothelial
inflammation. Proc Natl Acad Sci USA. 108:10355–10360. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tong JL, Zhang CP, Nie F, et al: MicroRNA
506 regulates expression of PPAR alpha in
hydroxycamptothecin-resistant human colon cancer cells. FEBS Lett.
585:3560–3568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Karbiener M, Fischer C, Nowitsch S, et al:
microRNA miR-27b impairs human adipocyte differentiation and
targets PPARgamma. Biochem Biophys Res Commun. 390:247–251. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim SY, Kim AY, Lee HW, et al: miR-27a is
a negative regulator of adipocyte differentiation via suppressing
PPARgamma expression. Biochem Biophys Res Commun. 392:323–328.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee EK, Lee MJ, Abdelmohsen K, et al:
miR-130 suppresses adipogenesis by inhibiting peroxisome
proliferator-activated receptor gamma expression. Mol Cell Biol.
31:626–638. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Repnik U, Knezevic M and Jeras M: Simple
and cost-effective isolation of monocytes from buffy coats. J
Immunol Methods. 278:283–292. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Glinghammar B, Skogsberg J, Hamsten A and
Ehrenborg E: PPARδ activation induces COX-2 gene expression and
cell proliferation in human hepatocellular carcinoma cells. Biochem
Biophys Res Commun. 308:361–368. 2003.
|
|
22
|
Lundell K, Thulin P, Hamsten A and
Ehrenborg E: Alternative splicing of human peroxisome
proliferator-activated receptor delta (PPARdelta):effects on
translation efficiency and trans-activation ability. BMC Mol Biol.
8:702007. View Article : Google Scholar
|
|
23
|
Contreras J and Rao DS: MicroRNAs in
inflammation and immune responses. Leukemia. 26:404–413. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wagner K-D and Wagner N: Peroxisome
proliferator-activated receptor beta/delta (PPARδ) acts as
regulator of metabolism linked to multiple cellular functions.
Pharmacol Ther. 125:423–435. 2010.
|
|
25
|
Zhang Y, Liao Y, Wang D, et al: Altered
expression levels of miRNAs in serum as sensitive biomarkers for
early diagnosis of traumatic injury. J Cell Biochem. 112:2435–2442.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Krichevsky AM, Sonntag K-C, Isacson O and
Kosik KS: Specific microRNAs modulate embryonic stem cell–derived
neurogenesis. Stem Cells. 24:857–864. 2006.
|
|
27
|
Nie K, Gomez M, Landgraf P, et al:
MicroRNA-mediated down-regulation of PRDM1/Blimp-1 in
Hodgkin/Reed-Sternberg cells: a potential pathogenetic lesion in
Hodgkin lymphomas. Am J Pathol. 173:242–252. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma L, Young J, Prabhala H, et al: miR-9, a
MYC/MYCN-activated microRNA, regulates E-cadherin and cancer
metastasis. Nat Cell Biol. 12:247–256. 2010.PubMed/NCBI
|
|
29
|
Plaisance V, Abderrahmani A, Perret-Menoud
V, Jacquemin P, Lemaigre F and Regazzi R: MicroRNA-9 controls the
expression of granuphilin/Slp4 and the secretory response of
insulin-producing cells. J Biol Chem. 281:26932–26942. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Agami R: microRNAs, RNA binding proteins
and cancer. Eur J Clin Invest. 40:370–374. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tan NS, Michalik L, Noy N, et al: Critical
roles of PPAR beta/delta in keratinocyte response to inflammation.
Genes Dev. 15:3263–3277. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Delerive P, De Bosscher K, Besnard S, et
al: Peroxisome proliferator-activated receptor alpha negatively
regulates the vascular inflammatory gene response by negative
cross-talk with transcription factors NF-kappaB and AP-1. J Biol
Chem. 274:32048–32054. 1999. View Article : Google Scholar
|
|
33
|
Guo L-M, Pu Y, Han Z, et al: MicroRNA-9
inhibits ovarian cancer cell growth through regulation of NF-κB1.
FEBS J. 276:5537–5546. 2009.PubMed/NCBI
|
|
34
|
Wan H-Y, Guo L-M, Liu T, Liu M, Li X and
Tang H: Regulation of the transcription factor NF-kappaB1 by
microRNA-9 in human gastric adenocarcinoma. Mol Cancer. 9:162010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wolfs IM, Donners MM and de Winther MP:
Differentiation factors and cytokines in the atherosclerotic plaque
micro-environment as a trigger for macrophage polarisation. Thromb
Haemost. 106:763–771. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee CH, Chawla A, Urbiztondo N, et al:
Transcriptional repression of atherogenic inflammation: modulation
by PPARdelta. Science. 302:453–457. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kharroubi I, Lee CH, Hekerman P, et al:
BCL-6: a possible missing link for anti-inflammatory PPAR-δ
signalling in pancreatic beta cells. Diabetologia. 49:2350–2358.
2006.PubMed/NCBI
|
|
38
|
Chen T, Huang Z, Wang L, et al:
MicroRNA-125a-5p partly regulates the inflammatory response, lipid
uptake, and ORP9 expression in oxLDL-stimulated
monocyte/macrophages. Cardiovasc Res. 83:131–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peters JM and Gonzalez FJ: Sorting out the
functional role(s) of peroxisome proliferator-activated
receptor-β/δ (PPARβ/δ) in cell proliferation and cancer. Biochim
Biophys Acta. 1796:230–241. 2009.PubMed/NCBI
|
|
40
|
Mukundan L, Odegaard JI, Morel CR, et al:
PPAR-[delta] senses and orchestrates clearance of apoptotic cells
to promote tolerance. Nat Med. 15:1266–1272. 2009.
|