Introduction
Mulberry fruit (Morus alba L.) has been
reported to be have beneficial effects in biological events, such
as oxidative stress (1),
hyperlipidemia (1), cancer
(2), neurodegeneration (3) and inflammation (4). Such effects are associated with
polyphenol compounds, such as cyanidin 3-rutinoside, cyanidin
3-glucoside and rutin in mulberry fruit (2,4–6).
Nonetheless, to our knowledge, the inhibitory effects of mulberry
fruit on allergic response have not been reported to date.
Naringinase is known as α-rhamnopyranoside,
possessing the activities of α-L-rhamnosidase (E.C. 3.2.1.40) and
β-D-glucosidase (E.C. 3.2.1.21) (7). Thus, the enzyme is able to hydrolyze
naringin to release L-rhamnose and naringenin (8). The enzyme has been isolated from
plants, yeast, or microorganisms, such as Aspergillus niger
(8), Coniothyrium
diplodiella (9),
Aspergillus terreus (10)
and Penicillium decumbens (11). Moreover, the enzyme can also
hydrolyze rutin, isoquercetin, hesperidin, diosmin and ter-phenyl
glycosides (7,12). Thus, the enzyme has been widely
used in biological engineering processes, such as the debittering
of fruit juices, preparation of antibiotics, biotransformation of
steroids, production of aglycones, production of ginsenosides and
the production of glycolipids (7). Nevertheless, to our knowledge, the
biological activity of mulberry fruit in combination with
naringinase has not been reported to date.
Mast cells play an important role in the initiation
and progression of allergy-related diseases, such as asthma,
psoriasis and arthritis (13),
and cells expressing FcɛRI receptor on the plasma membrane are also
critical cells in the progression of allergic and anaphylactic
reactions (14). As the FcɛRI
receptor is known to be an immunoglobulin E (IgE) high-affinity
receptor, the activation of the FcɛRI receptor can promote the
liberation of various inflammatory mediators, including histamine,
chemotactic factors, cytokines and arachidonate metabolites from
IgE-activated mast cells (15).
In this respect, RBL-2H3 cells, belonging to mast cell lines
(16), have been commonly used
for research on IgE-FcɛRI interactions involving the intracellular
signaling cascade and the degranulation and formation of cytokines
or eicosanoids (17–19).
Previously, mulberry fruit has been shown to have
some beneficial effects in various biological events (1–4),
whereas to our knowledge, the inhibitory effects of mulberry fruit
on allergic response have not been reported to date. In this
respect, we hypothesized that mulberry fruit may exert inhibitory
effects on allergic response in IgE-activated mast cells. Thus, in
this study, we investigated the anti-allergic action of mulberry
fruit extract (MFE) or MFE in combination with naringinase (MFEN)
in IgE-activated RBL-2H3 cells, and investigated the mechanisms
responsible for the anti-allergic effects of MFEN. The data
presented in this study may provide further insight into the
therapeutic application of MFEN or its use as a functional
food.
Materials and methods
Reagents
Minimal essential medium (MEM), penicillin,
streptomycin and fetal bovine serum (FBS) were purchased from
Gibco-Life Technologies (Grand Island, NY, USA).
4-[3-(4-Iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio]-1,3-benzene
disulfonate (WST-1) was obtained from Dojindo Laboratories
(Kumamoto, Japan). Specific antibodies against, Syk, phospho-Syk,
Lyn, phospho-Lyn, linker for activation of T cells (LAT),
phospho-LAT, extracellular signal-regulated protein kinase 1/2
(ERK1/2), phospho-ERK1/2, JNK, phospho-JNK, GRB2-associated binding
protein 2 (Gab2), phospho-Gab2, phosphoinositide-3-kinase (PI3K),
phospho-PI3K, Akt, phospho-Akt, phospholipase C (PLC)γ1,
phospho-PLCγ1, PLCγ2, phospho-PLCγ2, protein kinase C (PKC)δ,
phospho-PKCδ, cytosolic phospholipase A2
(cPLA2), phospho-cPLA2, cyclooxygenase-2 (COX-2) and
β-actin were purchased from Cell Signaling Technology, Inc.
(Beverly, MA, USA). A specific antibody against phospho-Fyn was
obtained from Biorbyt Ltd. (Cambridge, UK). The enzyme-linked
immunosorbent assay (ELISA) kit for tumor necrosis factor-α (TNF-α)
was obtained from eBioscience, Inc. (San Diego, CA, USA). Specific
antibodies against 5-lipoxygenase (5-LO) and phospho-5-LO, as well
as enzyme immunoassay (EIA) kits for prostaglandin D2
(PGD2), leukotriene B4 (LTB4) and
leukotriene C4 (LTC4) were purchased from
Cayman Chemical, Inc. (Ann Arbor, MI, USA). 4-Nitrophenyl
N-acetyl-β-D-glucosaminide (p-NAG), dinitrophenyl (DNP)-IgE,
DNP- human serum albumin (DNP-HSA) and naringinase were obtained
from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals were
of analytical grade.
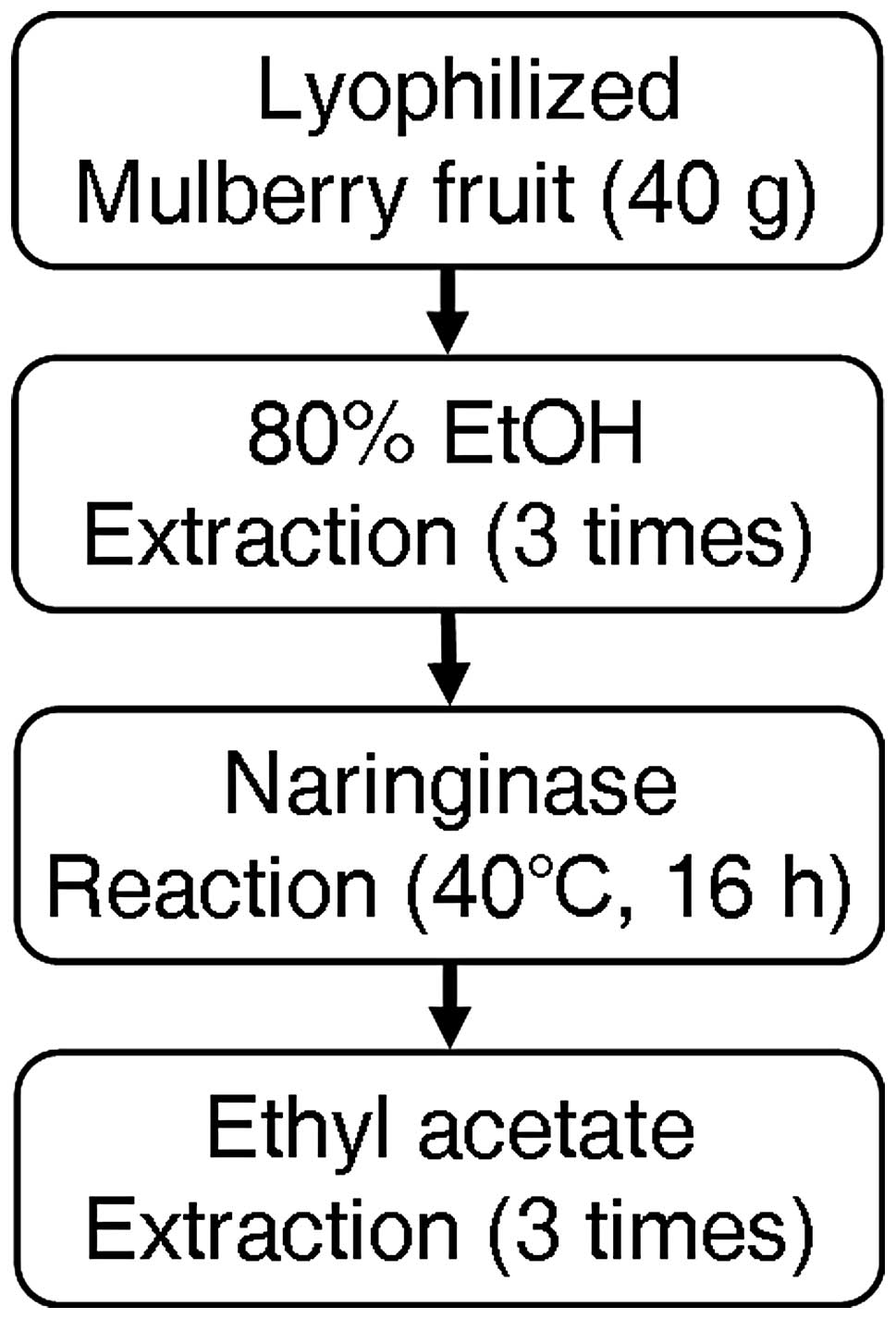
Preparation of MFE
Lyophilized mulberry fruit was obtained from
S&D, Inc. (Yeongi, Korea). Lyophilized mulberry fruit (40 g)
was extracted with 80% ethanol (1,000 ml) in a bath sonicator for 3
days, and the mixture was filtered by Whatman no. 3 filter paper.
The total filtrate was lyophilized, and then the residue of MFE (20
g) was completely dissolved in 0.1 M sodium acetate buffer (pH 4.5)
containing 2.5 g naringinase, and then incubated for 18 h at 40°C
(20). The solution was added to
ethyl acetate (1:1v/v), and then the layer of ethyl acetate was
separated and evaporated (evaporator, MG-2100; Buchi, Flawil,
Switzerland). The dried residue of ethyl acetate extract (1 g) was
dissolved in ethanol or suspended in water.
Analytical methods
High-performance liquid chromatography (HPLC)
analysis was carried out as previously described (5,6).
To analyze flavonoids, HPLC analysis was performed using a
Perkin-Elmer Flexar (Perkin-Elmer, Inc., Waltham, MA, USA) and a
Capcell PAK C18 column (4.6×250 mm, 5 μm; Shiseido,
Tokyo, Japan). The flavonoids were eluted in gradient system
composed of solvent A (methanol:water:acetic acid, 5:92.5:2.5;
v/v/v) and solvent B (methanol:water:acetic acid, 95:2.5:2.5;
v/v/v). The gradient was 10-50-50-60-10-10% of solvent B at
gradient time (tG, 0-12-17-30-30.1-35 min), oven temperature
was 40°C and the flow rate was 1.0 ml/min; an injection volume of
10 μl was applied. Perkin-Elmer Flexar UV/Vis detector was set at a
wavelength of 280 and 350 nm. The resulting data and
chromatographic profiles were evaluated using the
Chromera® Chromatography Data System (Perkin-Elmer,
Inc.).
To analyze anthocyanins, HPLC analysis was performed
using an Agilent Technologies 1200 series (Agilent Technologies,
Wilmington, DE, USA) a Synergi 4μ Polar-RP 80A (250×4.6 mm, 4 μm)
and a guard column (AQ C18 4×3.0 mm) (both from
Phenomenex, Inc., Torrance, CA, USA). The anthocyanins were eluted
in gradient system composed of solvent A (water:formic acid, 95:5;
v/v) and solvent B (acetonitrile:formic acid, 95:5; v/v).
The gradient was 0-10-13-15-15-5-5% of solvent B at gradient time
(tG =0-8-15-18-25-25.1-35 min), oven temperature was 40°C
and the flow rate was 1.0 ml/min; an injection volume of 10 μl was
applied. The UV/Vis detector was set at a wavelength of 520 nm. The
resulting data and chromatographic profiles were evaluated using
Analyst software (version 1.4.2; Applied Biosystems, Foster City,
CA, USA). Separately, to identify flavonoid compounds using the
liquid chromatography-electrospray tandem mass spectrometry
(LC-ESI/MS/MS), LC-ESI/MS/MS analysis was evaluated following a
previously described method (21). The LC-ESI/MS/MS system consisted
of an Shimadzu 20AD-XR HPLC system (Shimadzu, Kyoto, Japan) and an
API 3200 Q-TRAP LC-MS/MS system equipped with a Turbo V Ion Spray™
source (Applied Biosystems) operated in the negative ion mode. The
sample injection volume was 10 μl and the separation was performed
on a XTerra™ 3.5μ C18 column (2.1×50 mm i.d.; Waters,
Milford, MA, USA) with a SecurityGuard™ C18 guard column
(2.0×4.0 mm i.d.; Phenomenex, Inc.). The samples were analyzed via
multiple reaction monitoring (MRM). Quantification was performed by
MRM of the [M-H]− ion and the related production for
quercetin, using an internal standard to establish peak area
ratios.
Acquisition and data analysis were performed using
Analyst™ software (version 1.5.2; Applied Biosystems). Calibration
standards (3.9–1,000 nM) were prepared in blank matrices
pre-treated with ice-cold acetonitrile containing
4-methylumbelliferone (internal standard); the pre-treatment of
blank matrices was necessary due to the instability of the analytes
in the matrices. Calibration curves constructed using linear
least-squares regression were linear over the concentration range
of the standards used (r2 >0.999).
Relative standard deviation (RSD) of the measured concentrations
was used to assess the precision. A comparison of the mean measured
concentration versus the corresponding nominal concentration was
used to assess the accuracy. Both the accuracy (80–120%) and
precision (RSD <20%) of the assay were acceptable.
Cell culture
RBL-2H3 cells were cultured in MEM containing 5%
(v/v) FBS, 100 U/ml penicillin and 100 μg/ml streptomycin at 37°C
in a humidified atmosphere of 5% CO2 as previously
described (22).
Cytotoxicity assay
Cell respiration, an indicator of cell viability,
was determined by measuring the mitochondrial-dependent reduction
of WST-1 to water-soluble tetrazolium salt, as previously described
(23). Briefly, the RBL-2H3 cells
were seeded on a 96-well plate (1×104 cells/well) in MEM
with 5% FBS at 37°C overnight. The cells were washed, and then
incubated with DNP-IgE (1 μg/ml) for 24 h. The IgE-sensitized cells
were incubated with MFEN at various concentrations (0–200 μg/ml)
for 1 h. Both DNP-HSA (25 ng/ml) and WST-1 reagent (10 μl) were
simultaneously added to the above, and the mixture was incubated
for a further 4 h. To measure cell viability, the absorbance was
measured at 450 nm using a microplate reader (Emax; Molecular
Devices Inc., Sunnyvale, CA, USA).
β-hexosaminidase release activity
The RBL-2H3 cells were incubated in a 24-well plate
(1×105 cells/well) at 37°C overnight. The above cells
were washed with 1× PBS, and then incubated with DNP-IgE for 24 h.
IgE-sensitized cells were incubated with MFEN (0–200 μg/ml) for 1
h, spiked with DNP-HSA, and then incubated for a further 4 h. To
measure the amount of β-hexosaminidase activity released from the
cells, the culture medium was transferred and centrifuged (17,000 ×
g for 10 min) at 4°C. The supernatant (25 μl) was mixed with 50 μl
p-NAG (10 mM) in 0.1 M sodium citrate buffer (pH 4.5) into a
96-well plate, and then incubated for 1 h at 37°C. The reaction was
terminated by stop buffer (0.1 M Na2CO3
buffer, pH 10.0). The β-hexosaminidase activity was determined by
measuring the difference in absorbance at 405 nm.
ELISA of TNF-α
To measure the TNF-α level in the culture medium,
all culture media were centrifuged (17,000 × g for 10 min) at 4°C,
and the samples were stored at −80°C until use. The concentration
of TNF-α was determined using ELISA kits (eBioscience, Inc.)
according to the manufacturer’s instructions.
EIA of PGD2, LTB4
and LTC4
To determine the levels of PGD2,
LTB4 and LTC4 in the culture medium, all
culture media were centrifuged (17,000 × g for 10 min) at 4°C, and
the supernatant was stored at −80°C until use. The concentrations
of PGD2, LTB4 and LTC4 were
determined using EIA kits (Cayman Chemical, Inc.) according to the
manufacturer’s instructions.
Immunoblot analysis
Immunoblot analysis was carried out according to a
previously described method (24). The membranes were then incubated
with a 1:1,000 dilution of specific antibodies against
phospho-PLCγ1, PLCγ1, phospho-PLCγ2, PLCγ2, phospho-PKCδ, PKCδ,
phospho-Fyn, phospho-Lyn, Lyn, phospho-Syk, Syk, phospho-LAT, LAT,
phospho-ERK1/2, ERK1/2, phospho-JNK, JNK, phospho-Gab2,
phospho-PI3K, PI3K, phospho-Akt, Akt, phospho-cPLA2,
cPLA2, COX-2 and β-actin (Cell Signaling Technology,
Inc.), and antibodies against phospho-5-LO and 5-LO (Cayman
Chemical, Inc.). The blots were washed with TBS-T, and then
incubated with a 1:5,000 dilution of horseradish
peroxidase-conjugated IgG secondary antibody (Cell Signaling
Technology, Inc.). The proteins on the membranes were detected
using a chemiluminescent reaction (ECL plus kit), followed by the
exposure of the membranes to Hyperfilm ECL (both from Amersham
Pharmacia Biotech, Buckinghamshire, UK). The levels of the target
proteins were compared to those of a loading control (β-actin or
non-phosphorylated protein), and then the density of the resolved
bands was evaluated using ImageJ software.
Statistical analysis
The experimental results are expressed as the means
± SD. One-way analysis of variance (ANOVA) was used for multiple
comparisons (GraphPad Prism version 4.03 for Windows; GraphPad
Software, San Diego, CA, USA). If there was a significant variation
between the treatment groups, the Dunnett test was applied. Values
of P<0.05 and P<0.01 were considered to indicate
statistically significant differences.
Results
Inhibitory effects of MFE or MFEN on
IgE-mediated allergic response in RBL-2H3 cells
First, to determine the effects of MFE on
IgE-antigen complex reaction, the IgE-sensitized RBL-2H3 cells were
exposed to MFE at various concentrations (0–75 mg/ml) for 1 h, and
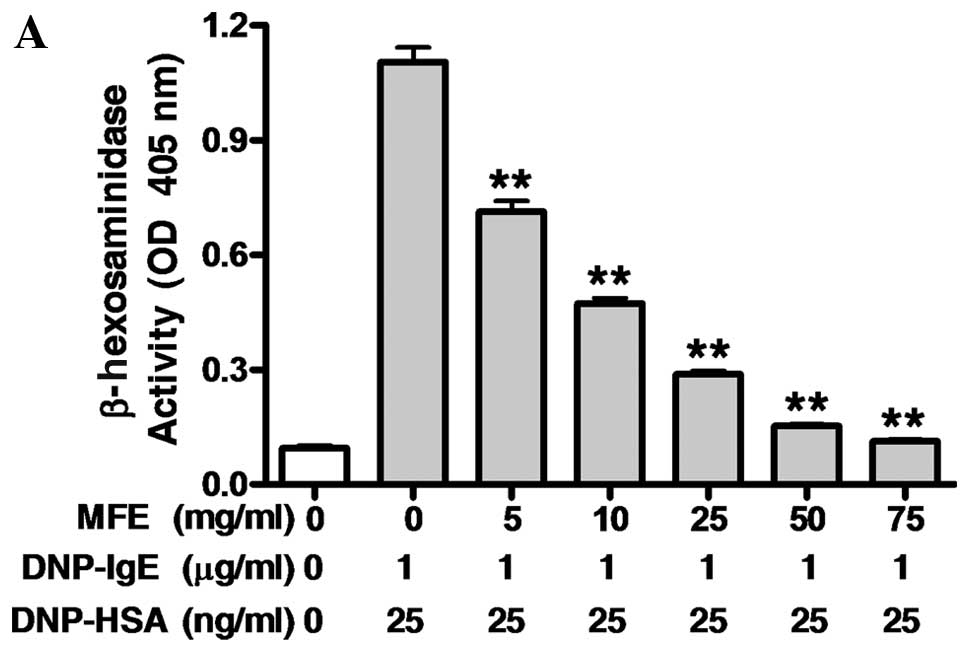
then stimulated with 25 ng/ml of DNP-HSA for 4 h. MFE markedly
inhibited the release of both β-hexosaminidase (IC50,
10.59 mg/ml), a general biomarker of degranulation, and TNF-α
(IC50, 4.87 mg/ml), a pro-inflammatory cytokine
(Fig. 2). When the IgE-sensitized
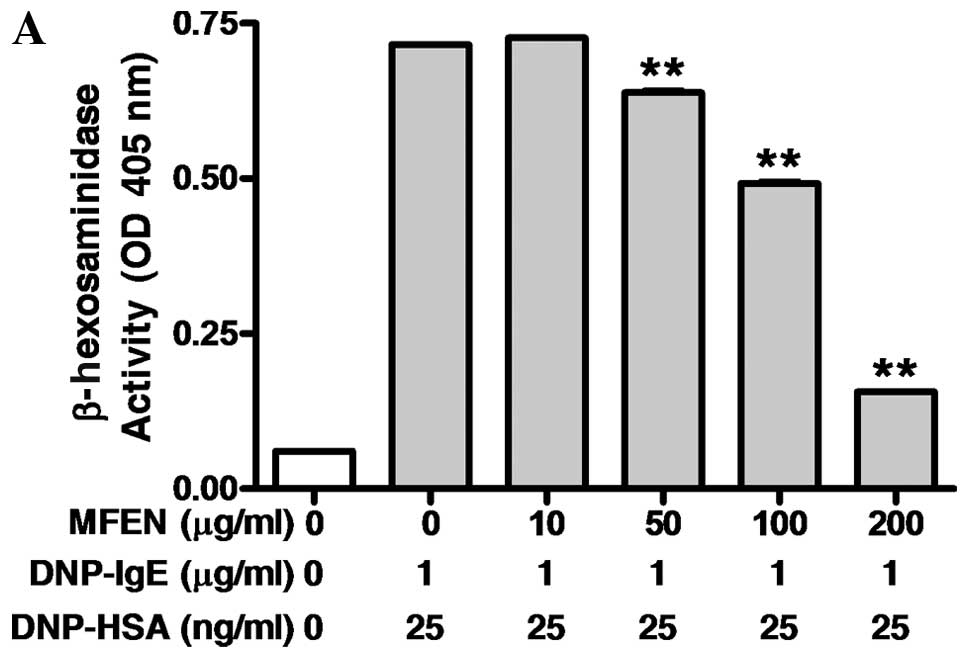
RBL-2H3 cells were exposed to MFEN, MFE in combination with
naringinase (Fig. 1) at various
concentrations (0–200 μg/ml) for 1 h prior to antigen challenge,
(Fig. 3A and B), MFEN markedly
suppressed the release of both β-hexosaminidase (IC50,
123.10 μg/ml) and TNF-α (IC50, 65.01 μg/ml). Moreover,
MFEN had no significant cytotoxicity at the concentrations used to
inhibit degranulation (Fig. 3C).
Taken together, these results indicate that MFE possesses
anti-allergic activity, and that the anti-allergic activity of MFE
may be derived from certain bioactive components, such as cyanidin
3-rutinoside, cyanidin 3-glucoside or rutin. In addition, treatment
with naringinase enhanced the inhibitory effects on allergic
response in IgE-activated mast cells. These effects of MFEN may be
associated with aglycones, which are released from glycosides.
Inhibitory effects of MFEN on the
formation of pro-inflammatory lipid mediators
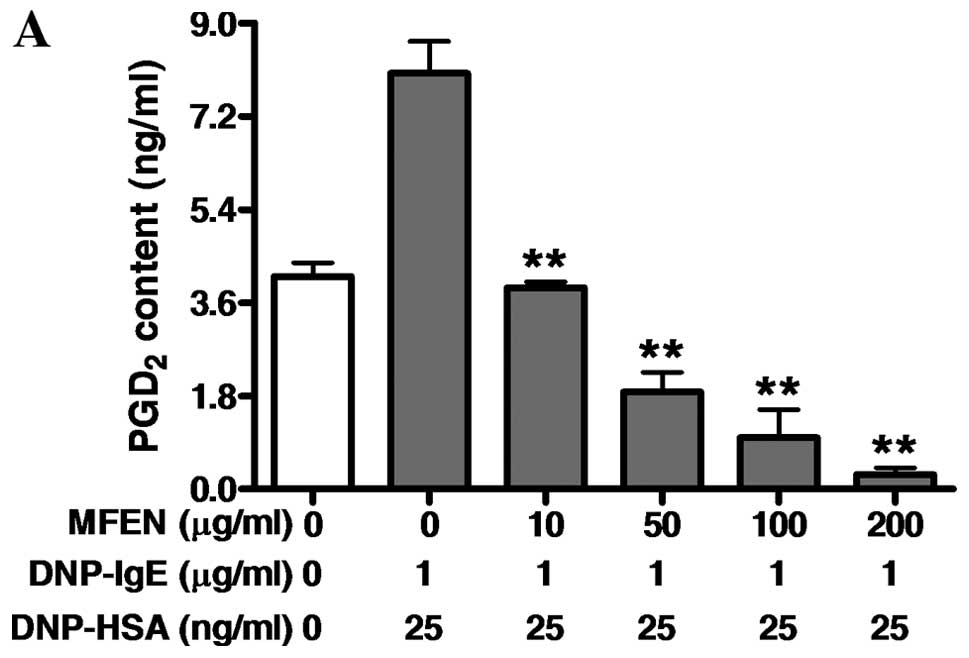
We then examined the effects of MFEN on the
formation of pro-inflammatory lipid mediators, such as
PGD2, LTB4 and LTC4 associated
with allergic response (16,25–28), since the activation of the
arachidonate cascade is involved in FcɛRI activation in
IgE-activated mast cells (29).
RBL-2H3 cells were pre-incubated with MFEN (0–200 μg/ml) prior to
antigen challenge, and the formation of PGD2,
LTB4 or LTC4 was then assessed. MFEN markedly
inhibited the formation of both PGD2 (IC50,
6.47 μg/ml) and LTC4 (IC50, 0.31 μg/ml),
whereas it only slightly suppressed the formation of
LTB4 (IC50, 25.75 μg/ml) (Fig. 4). Taken together, these results
suggest that MFEN suppresses allergic inflammation induced by
PGD2, LTC4 or LTB4. Thus, this
indicates that MFEN directly inhibits an enzyme involved in the
biosynthesis of prostaglandins or leukotrienes. In particular, our
results indicate that MFEN potently inhibits the formation of
LTC4, and MFEN may contain a specific inhibitor against
LTC4 synthase.
Regulatory effects of MFEN on the FcɛRI
signaling pathway
We then investigated the mechanisms responsible for
the anti-allergic effects of MFEN. The activation of the FcɛRI
receptor is known to be associated with the phosphorylation of Lyn
and Syk, which mediate the induction of degranulation in mast cells
(27). Quercetin, an aglycone of
isoquercetin or rutin, is known to inhibit the degranulation of
IgE-activated RBL-2H3 cells (30). Thus, we hypothesized that MFEN can
affect the phosphorylation of Lyn or Syk, as well as the FcɛRI
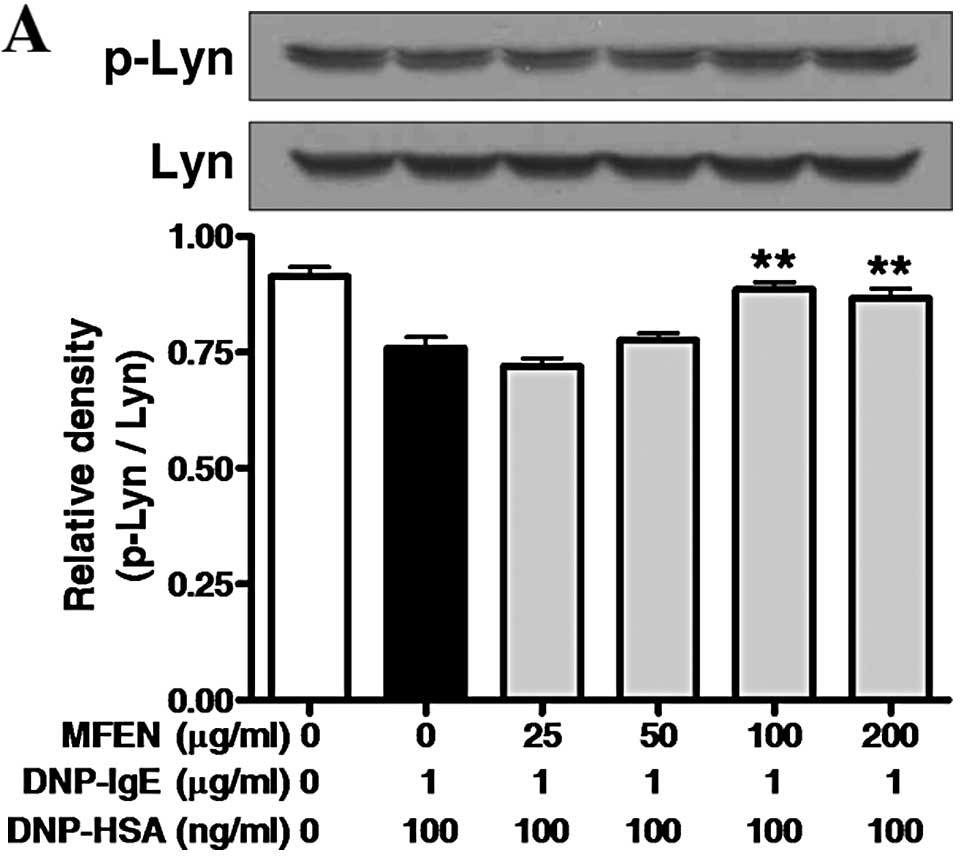
cascade. When the IgE-sensitized RBL-2H3 cells were pre-incubated
with MFEN (0–200 μg/ml) for 1 h prior to antigen challenge, and
then the incubation was extended for a further 10 min, MFEN
markedly inhibited Syk phosphorylation, but not that of Lyn or Fyn
(Fig. 5A). MFEN dose-dependently
suppressed the activation of PLCγ1/2 and PKCδ, implicated in the
degranulation process in mast cells (28) (Fig.
5B). Additionally, MFEN reduced the phosphorylation of Gab2,
PI3K, Akt, LAT, ERK1/2 and JNK (Fig.
5C and D). These findings indicate that MFEN may directly block
the activation of Syk. Consequently, MFEN also inhibited the
activation of other targets, such as PLCγ1/2, PKCδ, Gab2, PI3K,
Akt, LAT, ERK1/2 and JNK on the FcɛRI cascade in IgE-activated mast
cells.
 | Figure 5Effects of mulberry fruit extract in
combination with naringinase (MFEN) on the early stage of the FcɛRI
signaling cascade in immunoglobulin E (IgE)-activated RBL-2H3
cells. IgE-sensitized RBL-2H3 cells were exposed to MFEN (0–200
μg/ml) for 1 h, and then stimulated by dinitrophenyl-human serum
albumin (DNP-HSA) (100 ng/ml) for 10 min. The cells were rinsed
with 1× PBS, and lysed with cell lysis buffer. The expression of
(A) phoshoprylated (p-)Lyn, Lyn, p-Fyn, p-Syk, Syk, (B) p-PLCγ1,
PLCγ1, p-PLCγ2, PLCγ2, p-PKCδ, PKCδ, (C) p-Gab2, p-PI3K, PI3K,
p-Akt, Akt, (D) p-LAT, LAT, p-ERK1/2, ERK1/2, p-JNK, JNK or β-actin
was determined as described in Materials and methods. Similar
results were obtained in 3 independent experiments.
*P<0.05 and **P<0.01 vs.
DNP-HSA-treated group. |
Regulatory effects of MFEN on the enzymes
associated with the arachidonate cascade
We additionally investigated the anti-allergic
effects of MFEN on the activation of enzymes associated with the
arachidonate cascade. The activation of the arachidonate cascade
has been implicated in the activation of the FcɛRI receptor in
IgE-activated mast cells (27).
In this respect, we hypothesized that MFEN, which showed
anti-allergic effects, would affect the activation of
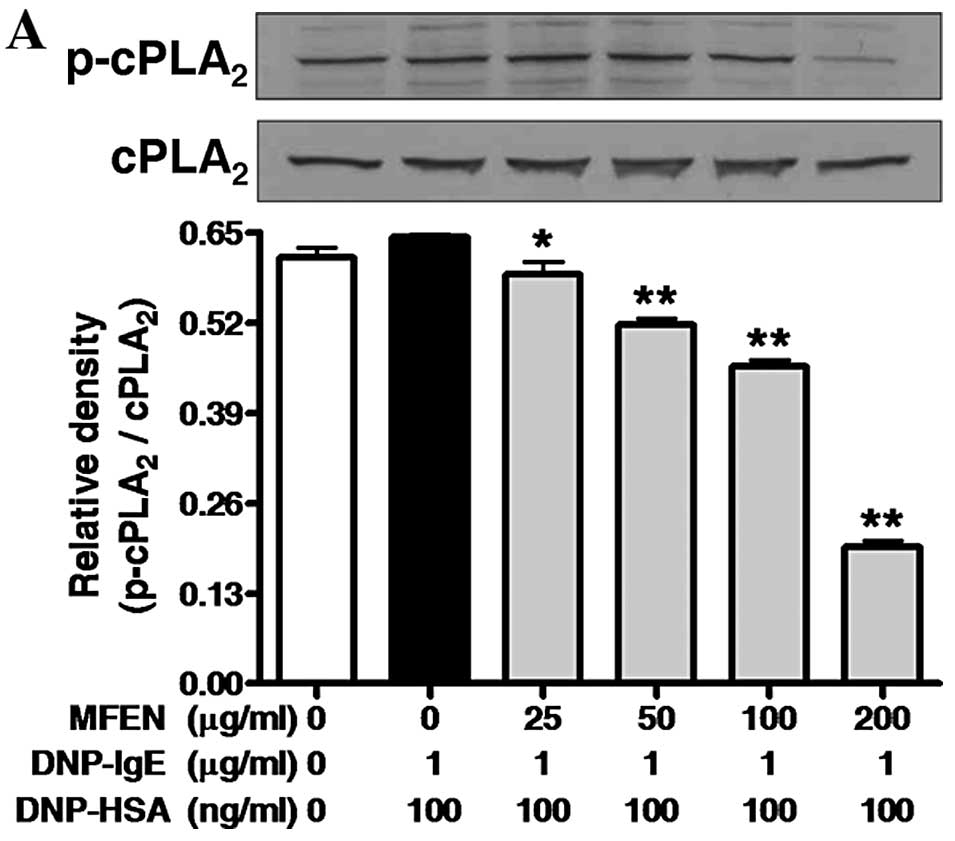
cPLA2, 5-LO or COX-2. When the IgE-sensitized RBL-2H3
cells were exposed to MFEN at various concentrations for 1 h prior
to stimulation with the antigen, and then the incubation was
extended for a further 4 h, MFEN suppressed the phosphorylation of
cPLA2 and 5-LO, as well as the expression of COX-2
(Fig. 6). These findings indicate
that MFEN suppresses the activation of cPLA2, 5-LO and
COX-2. In particular, since MFEN at 50 μg/ml strongly inhibited the
activation of 5-LO, the composition of MFEN may include a specific
inhibitor against 5-LO.
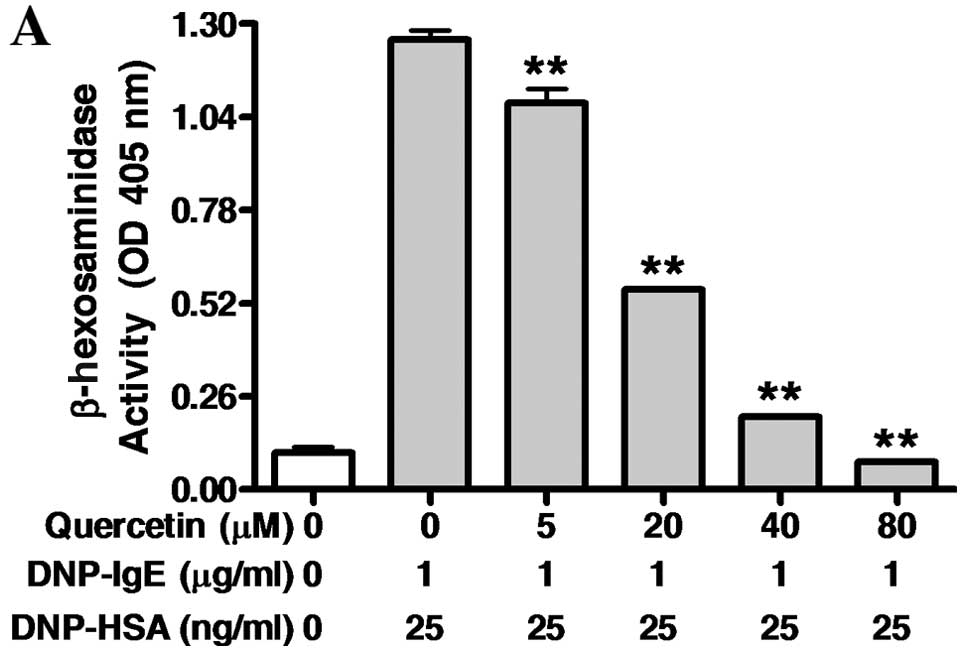
Inhibitory effects of quercetin on
IgE-mediated allergic response and flavonoid profiles of MFEN
Finally, to substantiate that MFEN includes
quercetin, we analyzed the chemical profiles for anthocyanins and
quercetin or its glycosides, such as isoquercetin and rutin using
HPLC and LC-ESI/MS/MS. MFE contained cyanidin 3-glucoside, cyanidin
3-rutinoside, isoquercetin and rutin, but not quercetin (Table I). On the other hand, MFEN showed
an increase in the amount of quercetin, and all the amounts of
glycoside compounds, including isoquercetin and rutin were reduced
by treatment with naringinase. In addition, to confirm the
anti-allergic effects of quercetin, found in high quantities in
MFEN, we examined the IgE-sensitized RBL-2H3 cells exposed to
quercetin at various concentrations (0–80 μM) for 1 h, and then
stimulated with 25 ng/ml of DNP-HSA for 4 h. Quercetin markedly
inhibited the release of both β-hexosaminidase (IC50,
17.58 μM) and TNF-α (IC50, 4.80 μM) (Fig. 7). Taken together, these results
indicate that MFEN contains a high level of quercetin, whereas the
glycoside compounds in MFEN are reduced by naringinase.
Consequently, the anti-allergic effects of MFEN may be derived from
quercetin.
 | Table IComposition of anthocyanins and
flavonoids of MFE or MFEN. |
Table I
Composition of anthocyanins and
flavonoids of MFE or MFEN.
| Anthocyanins | Flavonoids |
|---|
|
|
|
|---|
| C3G | C3R | Cyanidin | Isoquercetin | Rutin | Quercetin |
|---|
| MFE | 6.16±0.08 | 4.11±0.04 | 0.04±0.00 | 3.67±0.03 | 3.67±0.03 | ND |
| MFEN | 0.12±0.01 | ND | ND | ND | ND | 19.60±6.35 |
Discussion
Previously, mulberry fruit has been reported to
possess some biological properties, such as antioxidant (1), anti-hyperlipidemic (1), anti-cancer (2), anti-neurodegenerative (3) and anti-inflammatory properties
(4). Mulberry fruit is also known
to be rich in polyphenolic compounds, such as cyanidin
3-rutinoside, cyanidin 3-glucoside and rutin, associated with such
beneficial effects (2,4–6).
Nevertheless, to our knowledge, the inhibitory effects of mulberry
fruit on allergic response have not been reported to date.
Separately, naringinase is known to possess the
activities of α-L-rhamnosidase (EC 3.2.1.40) and β-D-glucosidase
(EC 3.2.1.21) (7). Thus, the
enzyme can also hydrolyze cyanidin 3-glucoside, cyanidin
3-rutinoside, isoquercetin or rutin, which found in rich quantities
in mulberry fruit, to aglycones with glucose or rutinose (7,12).
In support of this, MFE contains cyanidin 3-glucoside, cyanidin
3-rutinoside, isoquercetin and rutin, but not quercetin. In our
study, when MFE was incubated with naringinase, the amount of
quercetin in MFEN was highly elevated, whereas the amount of
isoquercetin or rutin was not detected. Moreover, the anti-allergic
effects of MFEN were more potent than those of MFE. In this
respect, cyanidin or quercetin, released from cyanidin 3-glucoside
or cyanidin 3-rutinoside, isoquercetin or rutin, respectively, may
be core chemicals with anti-allergic properties. Although mulberry
fruit has been known to contain richer quantities of cyanidin
3-glucoside or cyanidin 3-rutinoside than isoquercetin or rutin
(5), cyanidin or cyanidin
3-glucoside have been known to poorly inhibit allergic response in
IgE-activated mast cells (31).
In addition, in our study, cyanidin 3-glucoside did not inhibit
degranulation and TNF-α release in IgE-activated RBL-2H3 cells
(data not shown). Quercetin is known to exert a more potent
inhibitory effect than cyanidin on allergic response in
IgE-activated mast cells (30).
In support of this, in our study, MFEN and quercetin inhibited
degranulation and TNF-α release in IgE-activated RBL-2H3 cells.
Furthermore, the combination of naringinase and mulberry fruit
enhanced the anti-allergic activity. This may also enhance other
beneficial effects, and such effects may be derived from
quercetin.
Concerning the mechanism responsible for the
anti-allergic activity of MFEN, one possible mechanism may involve
the direct inhibition of the FcɛRI cascade. The IgE-induced
degranulation in mast cells is related to the activation of the
FcɛRI receptor, and this activation induces the release of various
inflammatory mediators containing TNF-α, leukotrienes and
prostaglandins through the phosphorylation of the Lyn or Fyn/Syk
pathway (28,32). Consequently, the activation of Syk
leads to the increase in intracellular Ca2+ levels and
the activation of the MAP kinase family (28). Thus, Lyn, Fyn and Syk are
important intracellular mediators in the early signaling pathway of
FcɛRI receptor activation. In the present study, the inhibition of
Syk by MFEN may support the notion that a primary target of MFEN
may be Syk. In support of this, MFEN significantly reduced the
phosphorylation of Gab2, PI3K, Akt, LAT, ERK1/2 and JNK, which
belongs to a downstream protein in the FcɛRI receptor cascade
(28). In addition, MFEN
suppressed the activation of PLCγ1/2 and PKCδ, which are involved
in the process of IgE-mediated degranulation in mast cells
(33).
Although MFEN did not completely suppress both
cPLA2 phosphorylation and COX-2 expression, it markedly
reduced the levels of PGD2, corresponding to the COX-2
product. Thus, these findings suggest that MFEN directly inhibits
COX-2 activity or PGD2 synthase in prostaglandin
biosynthesis. MFEN at 50 μg/ml completely inhibited the
phosphorylation of 5-LO, whereas MFEN had more potent inhibitory
effects on the formation of LTB4 and LTC4,
corresponding to the 5-LO product, than 5-LO activation. Thus,
these findings suggest that MFEN directly inhibits 5-LO activation,
as well as LTB4 hydrolase or LTC4 synthase.
Therefore, the inhibitory effects of MFEN on the formation of
PGD2, LTB4 and LTC4 may be
involved in its anti-allergic action. PGD2 is known to
cause bronchoconstriction, vasodilation, increase capillary
permeability and mucous production in asthma (25). LTB4 is a potent
chemoattractant and activator of neutrophils and other immune cells
in severe asthma (34,35); LTC4 is known to be a
potent spasmogenic and chemotactic biochemical and an agonist of
cysteinyl-LT receptors, which is known to induce chronic
inflammatory reactions in allergic diseases (26). Taken together, our results
suggested that MFEN inhibited allergic reaction through the
suppression of Syk, PLCγ1/2, PKCδ, Gab2, PI3K, Akt, LAT, ERK1/2 and
JNK, as well as the activation of cPLA2, 5-LO and COX-2.
Furthermore, such an effect of MFEN may be extended to its
anti-inflammatory effects on other cells or tissues. Several
cytokines may play critical roles in allergic inflammation
(13). In particular, TNF-α,
secreted from mast cells in IgE-antigen complex reaction (36), is known to play an important role
in allergic reaction (13).
Moreover, the expression of TNF-α is related to the JNK or ERK1/2
activation of the FcɛRI cascade in IgE-activated mast cells
(28,33). In the present study, MFEN markedly
inhibited the activation ERK1/2 and JNK, and reduced the TNF-α
level. Therefore, the reduction of TNF-α formation by MFEN may
provide an additional advantage of MFEN as an anti-allergic
food.
In conclusion, the present study demonstrates that
MFEN possesses anti-allergic functions in IgE-activated RBL-2H3
cells. These findings reveal a novel feature of MFEN in allergic
response. The mechanisms responsible for its anti-allergic effects
may involve multiple targets, including Syk, PLCγ1/2, PKCδ, Gab2,
PI3K, Akt, LAT, ERK1/2, JNK, cPLA2, 5-LO and COX-2. Such
effects may be derived from the presence of quercetin, which is
released from isoquercetin or rutin by naringinase, and provide
further insight into the application of MFEN as a therapeutic
reagent or a functional food.
Acknowledgements
This study was financially supported by the Basic
Science Research Program through the National Research Foundation
of Korea (NRF) funded by the Ministry of Education, Science and
Technology (2011-0013606).
References
|
1
|
Yang X, Yang L and Zheng H: Hypolipidemic
and antioxidant effects of mulberry (Morus alba L.) fruit in
hyperlipidaemia rats. Food Chem Toxicol. 48:2374–2379. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen PN, Chu SC, Chiou HL, Kuo WH, Chiang
CL and Hsieh YS: Mulberry anthocyanins, cyanidin 3-rutinoside and
cyanidin 3-glucoside, exhibited an inhibitory effect on the
migration and invasion of a human lung cancer cell line. Cancer
Lett. 235:248–259. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim HG, Ju MS, Shim JS, et al: Mulberry
fruit protects dopaminergic neurons in toxin-induced Parkinson’s
disease models. Br J Nutr. 104:8–16. 2010.PubMed/NCBI
|
|
4
|
Liu LK, Lee HJ, Shih YW, Chyau CC and Wang
CJ: Mulberry anthocyanin extracts inhibit LDL oxidation and
macrophage-derived foam cell formation induced by oxidative LDL. J
Food Sci. 73:H113–H121. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pawlowska AM, Oleszek W and Braca A:
Quali-quantitative analyses of Flavonoids of Morus nigra L. and
Morus alba L. (Moraceae) fruits. J Agric Food Chem.
56:3377–3380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Song W, Wang HJ, Bucheli P, Zhang PF, Wei
DZ and Lu YH: Phytochemical profiles of different mulberry
(Morus sp) species from China. J Agric Food Chem.
57:9133–9140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ribeiro MH: Naringinases: occurrence,
characteristics, and applications. Appl Microbiol Biotechnol.
90:1883–1895. 2011. View Article : Google Scholar
|
|
8
|
Bram B and Solomons GL: Production of the
enzyme naringinase by Aspergillus niger. Appl Microbiol.
13:842–845. 1965.PubMed/NCBI
|
|
9
|
Nomura D: Studies on naringinase produced
by Coniothyrium diplodiella. I The properties of naringinase
and the removal of co-existing pectinase from the enzyme
preparation. Enzymologia. 29:272–282. 1965.PubMed/NCBI
|
|
10
|
Soria F, Ellenrieder G, Grasselli M,
Navarro del Cañizo AA and Cascone O: Fractionation of the
naringinase complex from Aspergillus terreus by dye affinity
chromatography. Biotechnol Lett. 26:1265–1268. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Magario I, Vielhauer O, Neumann A,
Hausmann R and Syldatk C: Kinetic analysis and modeling of the
liquid-liquid conversion of emulsified di-rhamnolipids by
Naringinase from Penicillium decumbens. Biotechnol Bioeng.
102:9–19. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vila-Real H, Alfaia AJ, Bronze MR, Calado
AR and Ribeiro MH: Enzymatic synthesis of the flavone glucosides,
prunin and isoquercetin, and the aglycones, naringenin and
quercetin, with selective alpha-L-rhamnosidase and
beta-D-glucosidase activities of naringinase. Enzyme Res.
2011:6926182011. View Article : Google Scholar
|
|
13
|
Theoharides TC and Kalogeromitros D: The
critical role of mast cells in allergy and inflammation. Ann NY
Acad Sci. 1088:78–99. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Theoharides TC, Alysandratos KD, Angelidou
A, et al: Mast cells and inflammation. Biochim Biophys Acta.
1822:21–33. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gilfillan AM and Tkaczyk C: Integrated
signalling pathways for mast-cell activation. Nat Rev Immunol.
6:218–230. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Siraganian RP, McGivney A, Barsumian EL,
Crews FT, Hirata F and Axelrod J: Variants of the rat basophilic
leukemia cell line for the study of histamine release. Fed Proc.
41:30–34. 1982.PubMed/NCBI
|
|
17
|
Ortega E, Schweitzer-Stenner R and Pecht
I: Possible orientational constraints determine secretory signals
induced by aggregation of IgE receptors on mast cells. EMBO J.
7:4101–4109. 1988.PubMed/NCBI
|
|
18
|
Funaba M, Ikeda T and Abe M: Degranulation
in RBL-2H3 cells: regulation by calmodulin pathway. Cell Biol Int.
27:879–885. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ikawati Z, Wahyuono S and Maeyama K:
Screening of several Indonesian medicinal plants for their
inhibitory effect on histamine release from RBL-2H3 cells. J
Ethnopharmacol. 75:249–256. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ni H, Chen F, Cai H, Xiao A, You Q and Lu
Y: Characterization and preparation of Aspergillus niger
naringinase for debittering citrus juice. J Food Sci. 77:C1–C7.
2012.
|
|
21
|
Lee SY, Lee JY, Kang W, et al: Cytochrome
P450-mediated herb-drug interaction potential of Galgeun-tang. Food
Chem Toxicol. 51:343–349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Morita Y and Siraganian RP: Inhibition of
IgE-mediated histamine release from rat basophilic leukemia cells
and rat mast cells by inhibitors of transmethylation. J Immunol.
127:1339–1344. 1981.PubMed/NCBI
|
|
23
|
Ishiyama M, Tominaga H, Shiga M, Sasamoto
K, Ohkura Y and Ueno K: A combined assay of cell viability and in
vitro cytotoxicity with a highly water-soluble tetrazolium salt,
neutral red and crystal violet. Biol Pharm Bull. 19:1518–1520.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoo JM, Park ES, Kim MR and Sok DE:
Inhibitory effect of N-Acyl dopamines on IgE-mediated allergic
response in RBL-2H3 cells. Lipids. 48:383–393. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Arima M and Fukuda T: Prostaglandin
D2 and T(H)2 inflammation in the pathogenesis
of bronchial asthma. Korean J Intern Med. 26:8–18. 2011.
|
|
26
|
Nettis E, D’Erasmo M, Di Leo E, et al: The
employment of leukotriene antagonists in cutaneous diseases
belonging to allergological field. Mediators Inflamm.
2010:6281712010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kawakami Y, Kitaura J, Satterthwaite AB,
et al: Redundant and opposing functions of two tyrosine kinases,
Btk and Lyn, in mast cell activation. J Immunol. 165:1210–1219.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Roth K, Chen WM and Lin TJ: Positive and
negative regulatory mechanisms in high-affinity IgE
receptor-mediated mast cell activation. Arch Immunol Ther Exp
(Warsz). 56:385–399. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim Y, Lee YS, Hahn JH, et al: Hyaluronic
acid targets CD44 and inhibits FcepsilonRI signaling involving
PKCdelta, Rac1, ROS, and MAPK to exert anti-allergic effect. Mol
Immunol. 45:2537–2547. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morimoto Y, Yasuhara T, Sugimoto A, et al:
Anti-allergic substances contained in the pollen of Cryptomeria
japonica possess diverse effects on the degranulation of
RBL-2H3 cells. J Pharmacol Sci. 92:291–295. 2003.PubMed/NCBI
|
|
31
|
Han SJ, Ryu SN, Trinh HT, et al:
Metabolism of cyanidin-3-O-beta-D-glucoside isolated from black
colored rice and its antiscratching behavioral effect in mice. J
Food Sci. 74:H253–H258. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gomez G, Gonzalez-Espinosa C, Odom S, et
al: Impaired FcepsilonRI-dependent gene expression and defective
eicosanoid and cytokine production as a consequence of Fyn
deficiency in mast cells. J Immunol. 175:7602–7610. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Metcalfe DD: Mast cells and mastocytosis.
Blood. 112:946–956. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ford-Hutchinson AW, Bray MA, Doig MV,
Shipley ME and Smith MJ: Leukotriene B, a potent chemokinetic and
aggregating substance released from polymorphonuclear leukocytes.
Nature. 286:264–265. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tager AM and Luster AD: BLT1 and BLT2: the
leukotriene B(4) receptors. Prostaglandins Leukot Essent Fatty
Acids. 69:123–134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gordon JR and Galli SJ: Mast cells as a
source of both preformed and immunologically inducible
TNF-alpha/cachectin. Nature. 346:274–276. 1990.PubMed/NCBI
|