Introduction
Epilepsy is a common chronic neurological disorder
characterized by epileptic seizures, which vary in their duration.
These episodes can range from brief and nearly undetectable
seizures to long periods of vigorous shaking (1,2).
In epilepsy, seizures tend to recur, and have no immediate
underlying cause, while seizures that occur due to a specific cause
are not deemed to represent epilepsy (3). In the majority of cases, the cause
of epilepsy is unknown, although some individuals develop epilepsy
as a result of brain injury, stroke, brain cancer or drug and
alcohol misuse, among others. Epileptic seizures are the result of
excessive and abnormal cortical nerve cell activity in the brain
(3).
Genetics are believed to be involved in the majority
of cases of epilepsy, either directly or indirectly. Although some
of the genes involved affect ion channels, other molecules such as
enzymes, gamma-aminobutyric acid (GABA) and G protein-coupled
receptors have been identified as single genes in which defects
cause epilepsy; however, epilepsy may occur due to the interaction
of multiple genes and environmental factors (4). The etiology of epilepsy is
superficially thought to be multifactorial; however, the genetic
factors that may be involved in the pathogenesis of seizures have
not yet been elucidated. To study the gene regulatory networks
involved in epilepsy, a variety of genome-wide studies have been
performed by different groups using various systems and array
platforms. The accumulated functional genomic data are freely
available in the database of Gene Expression Omnibus (GEO,
http://www.ncbi.nlm.nih.gov/geo/)
(5,6), which provides a golden opportunity
for compiling a comprehensive list of genetic factors underlying
the etiology of epilepsy. According to the approach of
differentially expressed gene analysis (DEGA) for studying gene
expression profiles, hundreds of significant genes have been
identified to be associated with epilepsy. However, few studies
have focused on the associated pathways and transcription factors
(TFs), as well as on the co-expression patterns at the multiple
pathways level.
In the present study, we employed a microarray
dataset of genome-wide gene expression profiling from GEO, which is
associated with tumor-induced epileptogenesis. The most well-known
method of gene set enrichment analysis (GSEA) was used to analyze
the genomic data in order to uncover the regulatory mechanisms of
human epilepsy caused by brain tumors at the multiple pathways
level. GSEA is widely used to analyze gene expression profiles,
particularly to identify pre-defined gene sets which exhibit
significant differences in expression between samples from the
control and treatment groups (7–9).
The goal of GSEA is to determine other interesting categories
(pathways) where the constituent genes show coordinated changes in
expression over the experimental conditions, other than in the form
of sets of differentially expressed genes (DEGs). One of the
advantages of GSEA is the ability to highlight genes weakly
connected to the phenotype through pathway analysis, which may be
difficult to detect by using classical univariate statistics
(7).
Materials and methods
Microarray data collection and
pre-processing
We searched the GEO database (www.ncbi.nlm.nih.gov/geo/) for gene expression
profiling studies related to epilepsy. Data were included in our
re-analysis if they met the following criteria: i) the data were
genome-wide; ii) the comparison was conducted between samples with
epilepsy and controls; iii) complete microarray raw or normalized
data were available. Finally, we selected the dataset of GSE32534
for our re-analysis, which was contributed by Niesen et al
(10). In this dataset,
genome-wide gene expression profiling was conducted using the
Affymetrix Human Genome U133 Plus 2.0 Array and the RNA was derived
from formalin-fixed paraffin-embedded (FFPE) peritumoral cortex
tissue slides from 5-paired (seizure vs. non-seizure) low grade
brain tumor patients. There were 5 biological replicates for
epilepsy [samples from GSM805925 to GSM805929, marked with epilepsy
(EP)-1, EP-2, EP-3, EP-4 and EP-5, respectively] and 5 for the
controls [samples from GSM805930 to GSM805934, marked with control
(CT)-1, CT-2, CT-3, CT-4 and CT-5, respectively].
For the assessment of the influence of
pre-processing on the comparison, data pre-processing was performed
using software packages developed in version 2.6.0 of Bioconductor
and R version 2.10.1. Each Affymetrix dataset was background
adjusted, normalized and log2 probe-set intensities were calculated
using the Robust Multichip Averaging (RMA) algorithm in Affy
package, as previously described (11).
GSEA
Our GSEA of pathways and genes was performed using
the Category package in version 2.6.0 of Bioconductor, as
previously described (12). The
goal of GSEA is to determine whether the members of a gene set ‘S’
are randomly distributed throughout the entire reference gene list
‘L’ or are primarily found at the top or bottom of the list. One of
the advantages of GSEA is the relative robustness to noise and
outliers in the data. In our analysis, the gene sets represented by
<10 genes were excluded. The t-statistic mean of the genes was
computed in each Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway. Using a permutation test 1,000 times, the cut-off of the
significance level P-values was selected as 0.05 for the
significant pathways related to epilepsy. Accordingly, the
significant pathways and genes were then identified under the
comparison between the samples with epilepsy and no epilepsy. The
following classification of identified pathways was based on the
KEGG pathway maps br08901 of BRITE Functional Hierarchies in the
KEGG database (http://www.genome.jp/kegg-bin/get_htext?br08901.keg).
The annotation of significant genes in each pathway was performed
by using biomaRt package, BioMart v. 0.8 rc3 (version of 0.8
release candidate 3; http://www.biomart.org/). Subsequently, clustering of
the groups and genes was performed based on the identified gene
expression in each significant pathway using the method of
hierarchical clustering with Pearson correlation coefficient.
Regulatory elements (REs) and TFs of
co-regulated genes
We further employed a web server termed DiRE
(Distant RE of co-expressed genes, http://dire.dcode.org/), based on the enhancer
identification (EI) method, to predict common REs for our input
genes which have co-function in each identified significantly
related pathway (13). It
predicts function-specific REs consisting of clusters of
specifically-associated transcription factor binding sites (TFBSs),
and it also scores the association of individual TFs with the
biological function shared by the group of input genes. We selected
a random set of 5,000 genes in the human genome (hg18) as the
source of background genes. As a result, there are 2 major
parameters of our predicted TFs, including TF occurrence which
denotes the percentage of candidate RE containing a conserved
binding site for a particular TF; and TF importance which denotes
the product of TF occurrence and TF weight. As our candidate
associated TFs with input gene sets, we selected the cut-off value
of TF importance as >0.05.
Results and Discussion
Identification of significant pathways
associated with epilepsy
In the study of Niesen et al (10), a number of DEGs between the 2
groups (epilepsy and the control) were identified using both the
parametric unpaired Student’s t-test (345 probe sets representing
296 genes with fold-changes ≥2 plus P≤0.05) and the non-parametric
rank product [377 probe sets representing 344 genes with a false
discovery rate (FDR) of ≤0.3]. Seven DEGs, i.e., C1QB,
CALCRL, CCR1, KAL1, SLC1A2,
SSTR1 and TYRO3 were validated by qRT-PCR. Moreover,
the pathway analysis using DAVID bioinformatics resources revealed
that these DEGs were mainly enriched in focal adhesion,
extracellular matrix (ECM)-receptor interaction and cell adhesion
molecule (CAM) pathways.
Compared to the approach of DEGs, the strategy of
GSEA we used in this study is likely to be more powerful than
conventional single-gene methods in the study of complex diseases
in which many genes make subtle contributions. According to our
GSEA on the dataset of 10 samples by the comparison of epilepsy to
the controls, in total, there were 72 significant pathways
associated with epilepsy, whose P-values were <0.05, including 4
downregulated and 68 upregulated pathways. The 3 main pathways
identified by Niesen et al (10) were also included in our results.
Moreover, based on the KEGG pathway maps in the database of KEGG
(http://www.genome.jp/kegg/), these 72
significant pathways could be mainly mapped into 6 functional
classes, including cellular processes, environmental information
processing, genetic information processing, human diseases, as well
as metabolism and organismal systems. The details of the involved
pathways in each class are presented in Tables I–V. The details of the associated genes in
each significant pathway are available upon request (data not
shown), including the information on probe set id and gene
symbol.
 | Table ISignificant pathways associated with
epilepsy in the functional class of cellular processes. |
Table I
Significant pathways associated with
epilepsy in the functional class of cellular processes.
| Pathways | Map B | P-value | No. of genes | No. of TFs |
|---|
| 04510: Focal
adhesion | Cell
communication | 0.009 | 199 | 33 |
| 04540a: Gap junction | Cell
communication | 0.034 | 86 | 27 |
| 04210: Apoptosis | Cell growth and
death | <0.001 | 87 | 28 |
| 04115: p53 signaling
pathway | Cell growth and
death | 0.018 | 68 | 31 |
| 04110: Cell
cycle | Cell growth and
death | 0.035 | 122 | 37 |
| 04810: Regulation of
actin cytoskeleton | Cell motility | 0.048 | 211 | 37 |
| 04146:
Peroxisome | Transport and
catabolism | <0.001 | 78 | 29 |
| 04142: Lysosome | Transport and
catabolism | 0.014 | 111 | 26 |
| 04145: Phagosome | Transport and
catabolism | 0.031 | 141 | 29 |
| Table VSignificant pathways associated with
epilepsy in the functional class of organismal systems. |
Table V
Significant pathways associated with
epilepsy in the functional class of organismal systems.
| Pathways | Map B | P-value | No. of genes | No. of TFs |
|---|
| 04380: Osteoclast
differentiation | Development | 0.004 | 125 | 45 |
| 04730a: Long-term depression | Nervous system | 0.004 | 65 | 46 |
| 04920:
Adipocytokine signaling pathway | Endocrine
system | 0.027 | 67 | 31 |
| 04912a: GnRH signaling pathway | Endocrine
system | 0.029 | 94 | 25 |
| 03320: PPAR
signaling pathway | Endocrine
system | 0.045 | 68 | 21 |
| 04650: Natural
killer cell mediated cytotoxicity | Immune system | 0.004 | 125 | 31 |
| 04670: Leukocyte
transendothelial migration | Immune system | 0.009 | 113 | 46 |
| 04610: Complement
and coagulation cascades | Immune system | 0.013 | 67 | 25 |
| 04672: Intestinal
immune network for IgA production | Immune system | 0.013 | 44 | 30 |
| 04640:
Hematopoietic cell lineage | Immune system | 0.014 | 84 | 31 |
| 04612: Antigen
processing and presentation | Immune system | 0.017 | 63 | 21 |
| 04666: Fc gamma
R-mediated phagocytosis | Immune system | 0.018 | 90 | 31 |
| 04620: Toll-like
receptor signaling pathway | Immune system | 0.024 | 99 | 30 |
| 04662: B cell
receptor signaling pathway | Immune system | 0.028 | 74 | 43 |
| 04621: NOD-like
receptor signaling pathway | Immune system | 0.032 | 58 | 20 |
| 04623: Cytosolic
DNA-sensing pathway | Immune system | 0.033 | 53 | 17 |
| 04660: T cell
receptor signaling pathway | Immune system | 0.048 | 108 | 35 |
In the functional class of cellular processes, there
was 1 significantly downregulated and 8 significantly upregulated
pathways associated with epilepsy (Table I). These pathways were involved in
cell communication, cell growth and death, cell motility, as well
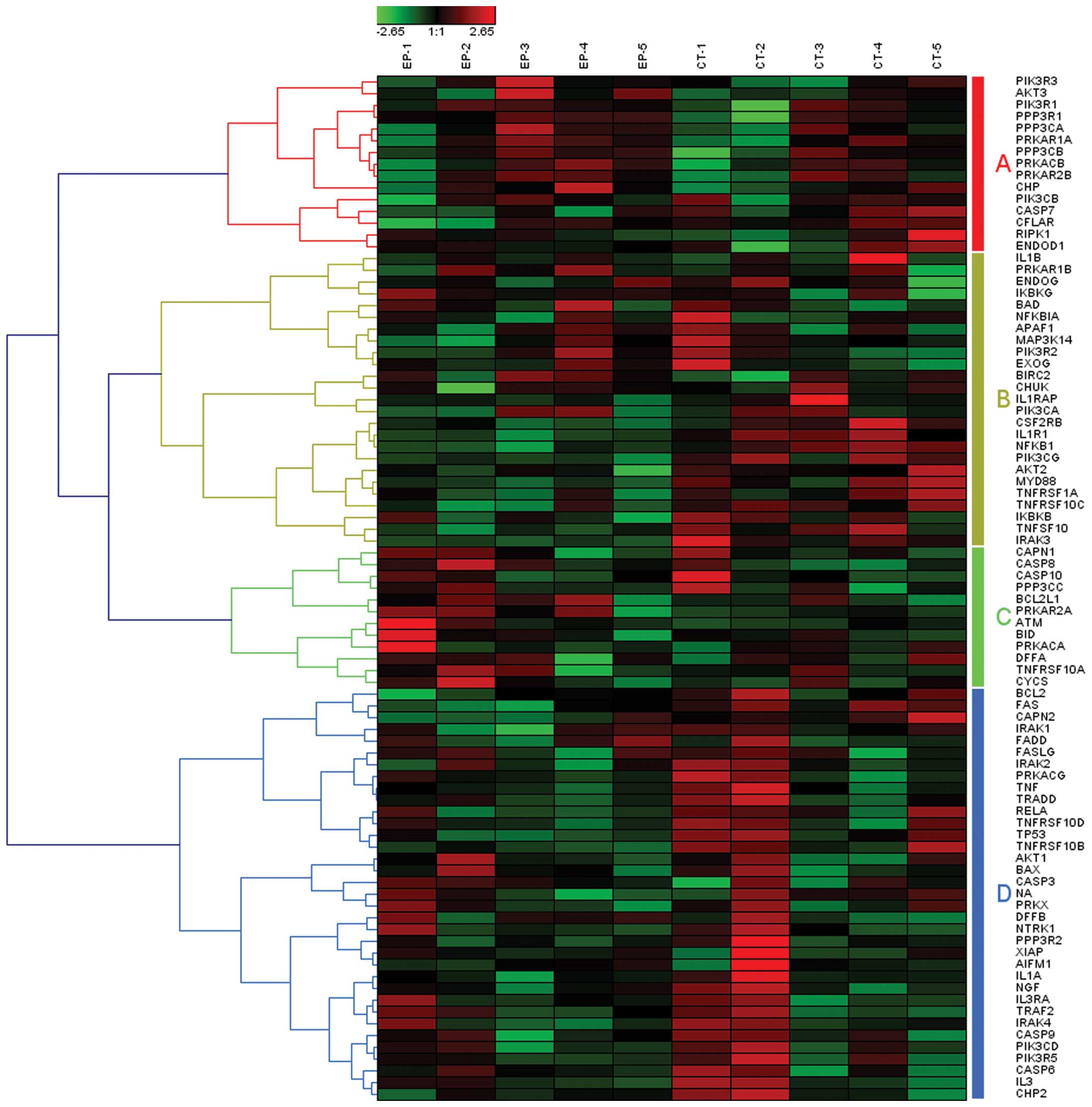
as transport and catabolism. Among these, apoptosis was one of the
most significant pathways (P<0.001), which was classified into
the functional group of cell growth and death. As is known,
high-frequency stimulation (HFS) of the hippocampus may be a
promising method in the treatment of epilepsy. It has been
indicated that hippocampal HFS can protect hippocampal neurons
against kainic acid (KA) neurotoxicity, and that the
neuroprotective effects of HFS may be mediated through the
inhibition of apoptosis (14). In
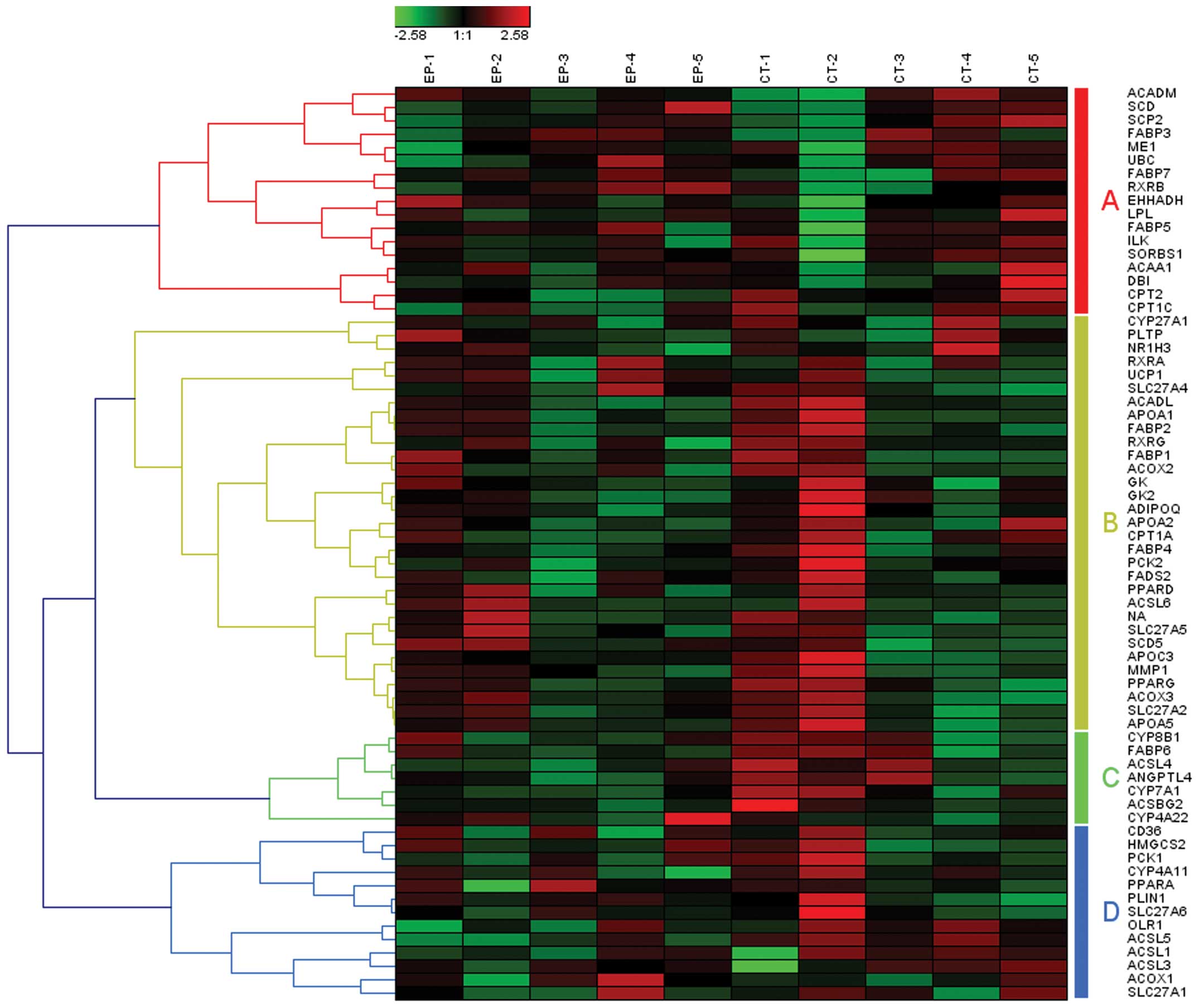
total, there were 87 involved genes in the pathway of apoptosis
related to epilepsy, which may be clustered into 4 gene set groups
based on hierarchical clustering with Pearson correlation
coefficient (Fig. 1, groups
A–D).
There were 4 and 3 significantly upregulated
pathways in the functional class of environmental and genetic
information processing, respectively which were associated with
epilepsy (Table II). The
environmental information processing pathways of ECM-receptor
interaction (P=0.004) and CAMs (P=0.020) were related to the
functions of signaling molecules and interaction, and the ABC
transporter pathway (P=0.030) was related to the function of
membrane transport, and the Jak-STAT signaling pathway (P=0.046)
was related to the function of signal transduction. The genetic
information processing pathways of mismatch repair (P=0.031) and
DNA replication (P=0.048) were related to the functions of
replication and repair, and the RNA polymerase pathway (P=0.008)
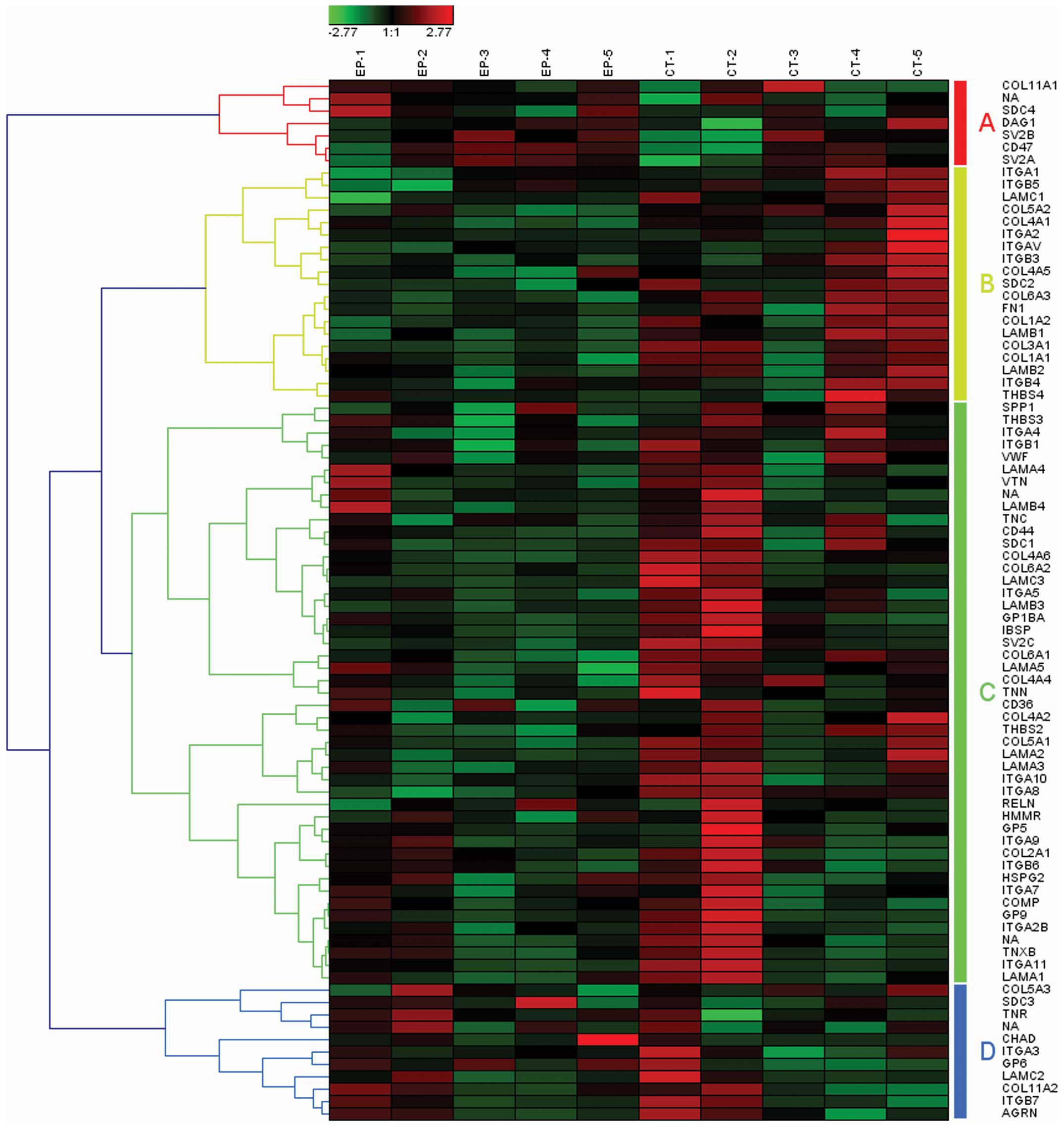
was transcription related. Among these, the pathway of ECM-receptor
interaction was the most significant in this class, which has also
been identified as one of the DEG enriched pathways by Niesen et
al (10). The ECM is known to
regulate important processes in neuronal cell development, activity
and growth. The remodeling of the ECM during both development and
following injury to the central nervous system has been shown to
affect neuronal guidance, synaptic plasticity and their
regenerative responses. The functions and potential therapeutic
value of several key ECM molecules in epileptogenesis and dementia
have been extensively investigated in previous studies (15,16). The 84 genes involved in the
pathway of ECM-receptor interaction may also be clustered into 4
gene set groups (Fig. 2, groups
A–D).
| Table IISignificant pathways associated with
epilepsy in the functional class of environmental and genetic
information processing. |
Table II
Significant pathways associated with
epilepsy in the functional class of environmental and genetic
information processing.
| Pathways | Map B | P-value | No. of genes | No. of TFs |
|---|
| 04512: ECM-receptor
interaction | Signaling molecules
and interaction | 0.004 | 84 | 28 |
| 04514: Cell adhesion
molecules (CAMs) | Signaling molecules
and interaction | 0.020 | 128 | 41 |
| 02010: ABC
transporters | Membrane
transport | 0.030 | 43 | 17 |
| 04630: Jak-STAT
signaling pathway | Signal
transduction | 0.046 | 153 | 33 |
| 03020: RNA
polymerase | Transcription | 0.008 | 27 | 20 |
| 03430: Mismatch
repair | Replication and
repair | 0.031 | 21 | 20 |
| 03030: DNA
replication | Replication and
repair | 0.048 | 35 | 11 |
Twenty-five significantly associated pathways were
classified into the functional class of human diseases, including 5
upregulated and 1 downregulated cancer-related pathways, 4
upregulated cardiovascular disease-related pathways, 1 upregulated
endocrine and metabolic disease-related pathway, 6 upregulated
immune disease-related pathways and 8 upregulated infectious
disease-related pathways (Table
III). It has been reported that cardiac changes may accompany
epilepsy, which may lead to significant seizure-associated cardiac
performance decreases (17).
These data are consistent with the results from our study which
identidied 4 significant cardiovascular disease-related pathways
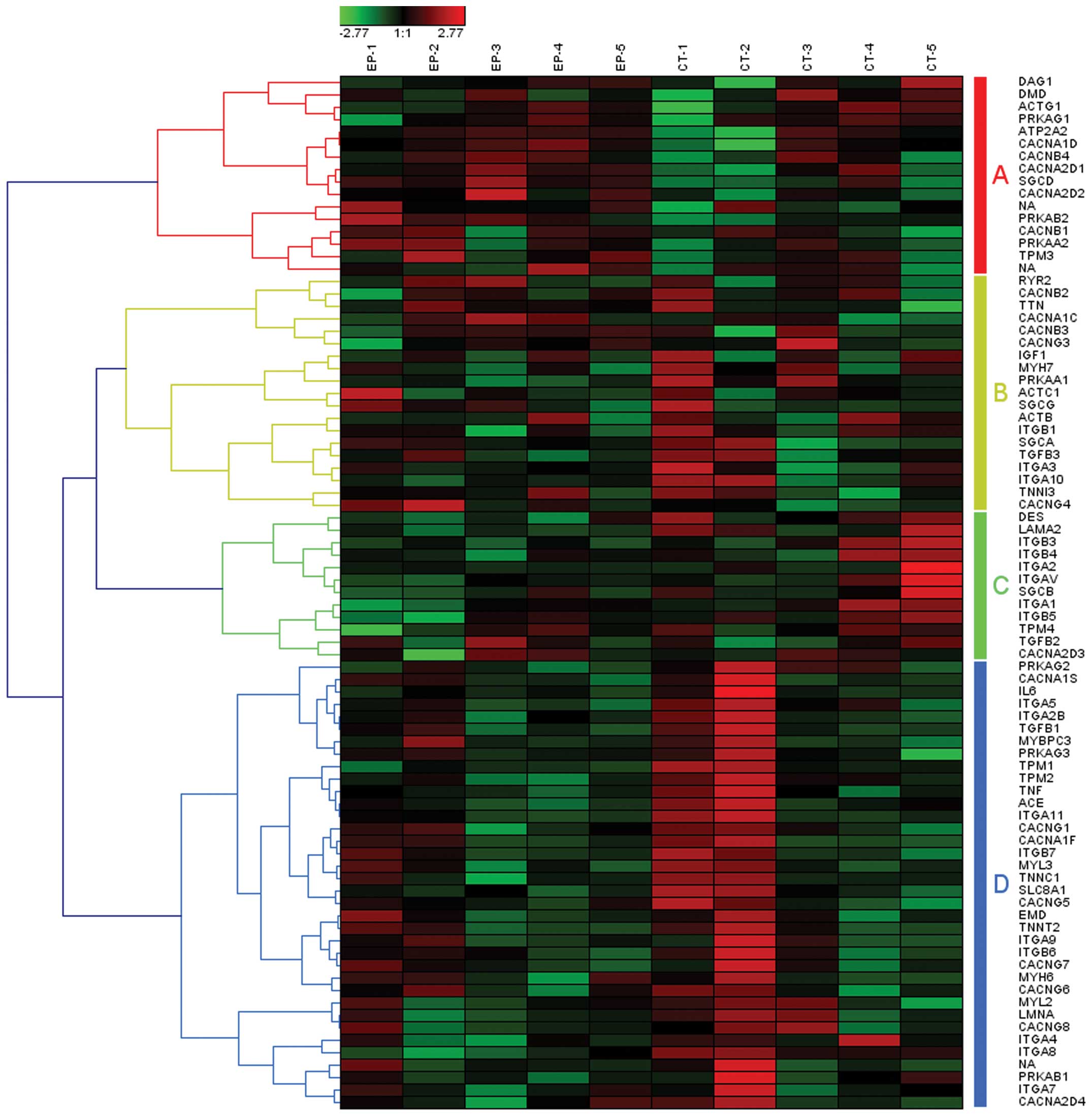
associated with epilepsy. Among these, the pathway of hypertrophic
cardiomyopathy (HCM) was one of the most significant cardiovascular
disease-related pathways (P=0.004). The association between these 2
diseases has been revealed in a clinical case report (18). In total, there were 83 genes
involved in the pathway of HCM associated with epilepsy, which may
be clustered into 4 gene set groups (Fig. 3, groups A–D).
| Table IIISignificant pathways associated with
epilepsy in the functional class of human diseases. |
Table III
Significant pathways associated with
epilepsy in the functional class of human diseases.
| Pathways | Map B | P-value | No. of genes | No. of TFs |
|---|
| 05222: Small cell
lung cancer | Cancers | 0.008 | 85 | 38 |
| 05213a: Endometrial cancer | Cancers | 0.008 | 52 | 30 |
| 05212: Pancreatic
cancer | Cancers | 0.011 | 69 | 40 |
| 05218:
Melanoma | Cancers | 0.012 | 71 | 40 |
| 05200: Pathways in
cancer | Cancers | 0.022 | 323 | 30 |
| 05219: Bladder
cancer | Cancers | 0.023 | 41 | 16 |
| 05410: Hypertrophic
cardiomyopathy (HCM) | Cardiovascular
diseases | 0.004 | 83 | 37 |
| 05412:
Arrhythmogenic right ventricular cardiomyopathy (ARVC) | Cardiovascular
diseases | 0.004 | 74 | 33 |
| 05416: Viral
myocarditis | Cardiovascular
diseases | 0.034 | 64 | 21 |
| 05414: Dilated
cardiomyopathy | Cardiovascular
diseases | 0.049 | 90 | 37 |
| 04940: Type I
diabetes mellitus | Endocrine and
metabolic diseases | 0.013 | 39 | 24 |
| 05323: Rheumatoid
arthritis | Immune
diseases | 0.004 | 83 | 19 |
| 05322: Systemic
lupus erythematosus | Immune
diseases | 0.013 | 82 | 24 |
| 05320: Autoimmune
thyroid disease | Immune
diseases | 0.013 | 47 | 20 |
| 05330: Allograft
rejection | Immune
diseases | 0.013 | 33 | 14 |
| 05332:
Graft-versus-host disease | Immune
diseases | 0.013 | 33 | 21 |
| 05310: Asthma | Immune
diseases | 0.020 | 26 | 27 |
| 05100: Bacterial
invasion of epithelial cells | Infectious
diseases: Bacterial | 0.009 | 70 | 36 |
| 05150:
Staphylococcus aureus infection | Infectious
diseases: Bacterial | 0.013 | 48 | 19 |
| 05146:
Amoebiasis | Infectious
diseases: Parasitic | <0.001 | 106 | 25 |
| 05145:
Toxoplasmosis | Infectious
diseases: Parasitic | 0.008 | 120 | 35 |
| 05140:
Leishmaniasis | Infectious
diseases: Parasitic | 0.008 | 66 | 25 |
| 05144: Malaria | Infectious
diseases: Parasitic | 0.024 | 48 | 28 |
| 05142: Chagas
disease (American trypanosomiasis) | Infectious
diseases: Parasitic | 0.036 | 101 | 41 |
| 05160: Hepatitis
C | Infectious
diseases: Viral | 0.018 | 131 | 38 |
In the functional class of metabolism, there were 14
significantly upregulated pathways associated with epilepsy
(Table IV). These were involved
in 6 different types of metabolism, including amino acid
metabolism, carbohydrate metabolism, glycan biosynthesis and
metabolism, lipid metabolism, metabolism of co-factors and
vitamins, as well as nucleotide metabolism. Among these, the
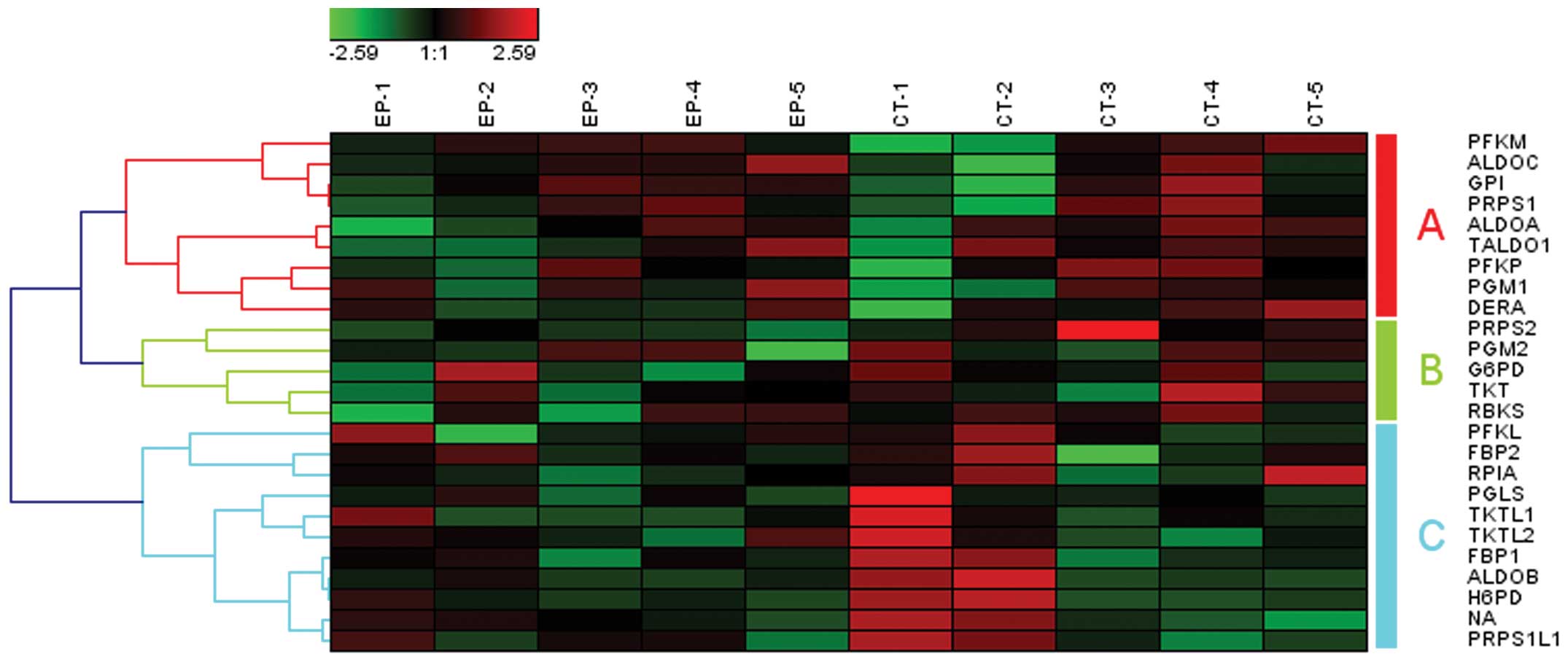
pentose phosphate pathway was one of the most significant pathways
(P=0.007), which was classified into the functional group of
carbohydrate metabolism. A variety of observations suggested that
decreasing glycolysis and increasing the levels of reduced
glutathione, generated by the metabolism of glucose through the
pentose phosphate pathway, may have an anticonvulsant effect.
Fructose-1,6-bisphosphate (F1,6BP) may have anticonvulsant activity
in models of acute seizures in adult rats, which shifts the
metabolism of glucose from glycolysis to the pentose phosphate
pathway (19). The total of 25
genes involved in the pentose phosphate pathway may be clustered
into 3 gene set groups (Fig. 4,
groups A–C).
| Table IVSignificant pathways associated with
epilepsy in the functional class of metabolism. |
Table IV
Significant pathways associated with
epilepsy in the functional class of metabolism.
| Pathways | Map B | P-value | No. of genes | No. of TFs |
|---|
| 00360:
Phenylalanine metabolism | Amino acid
metabolism | 0.031 | 17 | 26 |
| 00380: Tryptophan
metabolism | Amino acid
metabolism | 0.039 | 42 | 21 |
| 00030: Pentose
phosphate pathway | Carbohydrate
metabolism | 0.007 | 25 | 39 |
| 00520: Amino sugar
and nucleotide sugar metabolism | Carbohydrate
metabolism | 0.018 | 46 | 29 |
| 00010:
Glycolysis/gluconeogenesis | Carbohydrate
metabolism | 0.024 | 63 | 25 |
| 00511: Other glycan
degradation | Glycan biosynthesis
and metabolism | 0.014 | 16 | 22 |
| 00604:
Glycosphingolipid biosynthesis - ganglio series | Glycan biosynthesis
and metabolism | 0.043 | 15 | 26 |
| 00071: Fatty acid
metabolism | Lipid
metabolism | 0.021 | 42 | 22 |
| 00100: Steroid
biosynthesis | Lipid
metabolism | 0.032 | 18 | 21 |
| 00860: Porphyrin
and chlorophyll metabolism | Metabolism of
cofactors and vitamins | 0.009 | 30 | 23 |
| 00770: Pantothenate
and CoA biosynthesis | Metabolism of
cofactors and vitamins | 0.013 | 16 | 20 |
| 00480: Glutathione
metabolism | Metabolism of other
amino acids | 0.022 | 47 | 19 |
| 00230: Purine
metabolism | Nucleotide
metabolism | <0.001 | 155 | 26 |
| 00240: Pyrimidine
metabolism | Nucleotide
metabolism | 0.029 | 92 | 43 |
In the last functional class of organismal systems,
there were 2 significantly downregulated and 15 significantly
upregulated pathways associated with epilepsy (Table V). These were involved in
development, the nervous system, the endocrine system, as well as
the immune system. Among these, the peroxisome
proliferator-activated receptor (PPAR) signaling pathway was one of
the most associated pathways (P=0.045), which was classified into
the functional group of endocrine system. PPAR-γ/mitochondrial
uncoupling protein 2 signaling has been reported to protect neurons
against seizure-induced neuronal cell death in the hippocampus
following experimental status epilepticus (20). The activation of the mammalian
target of rapamycin or PPAR-γ pathways has been considered as one
of the pre-clinical models for the anti-epileptogenic activity of a
diverse range of potential therapies (21). Furthermore, there were 68 genes in
total which were involved in the PPAR signaling pathway associated
with epilepsy, which may be clustered into 4 gene set groups
(Fig. 5, groups A–D).
Candidate TF selection related to
epilepsy
To predict the TFs potentially involved in the
regulation of epilepsy, we performed the analysis of TFBSs and the
prediction of TFs using the significant genes in each identified
pathway. Based n the cut-off value of TF importance, we identified
the candidate TFs related to epilepsy with potential target genes
which were co-regulated in each of the 72 pathways identified. The
details are available upon request.
Among these, the TF of PPAR-γ appeared in several
pathways, including fatty acid metabolism, pyrimidine metabolism,
the PPAR signaling pathway, lysosome, amoebiasis and melanoma (data
available upon request). PPARs belong to the nuclear hormone
receptor superfamily. They play critical physiological roles as
lipid sensors and regulators of lipid metabolism and are activated
by fatty acids (22). Previous
studies have shown that PPAR-γ has anti-inflammatory effects in
seizure animal models (20,23). The anticonvulsant effects of acute
pioglitazone on pentylenetetrazol (PTZ)-induced seizures in mice
have been demonstrated to be mediated through the PPAR-γ
receptor-mediated pathway (24).
One of the PPAR-γ agonists, rosiglitazone, has been proven to
protect the central nervous system from oxidative damage in
epileptic rats, which may be a potential neuroprotective agent for
epilepsy (25). Our data also
support the idea that PPAR-γ may be one of the essential targets
for the management of epilepsy due to brain tumors.
Another essential TF of the
phosphatidylethanolamine-binding protein (PEBP) was identified in 3
of 4 downregulated pathways, including the GnRH signaling pathway,
gap junction and long-term depression pathways. PEBP is
alternatively named Raf-1 kinase inhibitor protein, the precursor
of the hippocampal cholinergic neurostimulating peptide (HCNP)
corresponding to its natural N-terminal fragment, which has been
previously described to be released by hippocampal neurons
(26–28). The crystal structure of human PEBP
(hPEBP) suggests that the ligand-binding site may accommodate the
phosphate head groups of membrane lipids, therefore allowing the
protein to adhere to the inner leaf of bilipid membranes where it
would be ideally positioned to relay signals from the membrane to
the cytoplasm (29). With the
previous evidence of calpain dysregulation in Alzheimer’s disease
(AD), PEBP has been confirmed as a novel in vitro and in
situ calpain substrate using an in vitro proteomics
approach or serial analysis of gene expression (SAGE) (30,31). In particular, during brain
development, the N-terminal part of mammalian PEBP has been
reported to be specifically cleaved and the resulting 11 amino acid
peptides may stimulate the growth and activity of acetylcholinergic
neurons (32).
Acknowledgements
We acknowledge financial support by the Scientific
Research Foundation and Academic and Technology Leaders
Introduction Project, and the 211 project of Anhui University
(10117700023).
References
|
1
|
Chang BS and Lowenstein DH: Epilepsy. N
Engl J Med. 349:1257–1266. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Magiorkinis E, Sidiropoulou K and
Diamantis A: Hallmarks in the history of epilepsy: epilepsy in
antiquity. Epilepsy Behav. 17:103–108. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fisher RS, van Emde Boas W, Blume W, et
al: Epileptic seizures and epilepsy: definitions proposed by the
International League Against Epilepsy (ILAE) and the International
Bureau for Epilepsy (IBE). Epilepsia. 46:470–472. 2005. View Article : Google Scholar
|
|
4
|
Mulley JC, Scheffer IE, Harkin LA,
Berkovic SF and Dibbens LM: Susceptibility genes for complex
epilepsy. Hum Mol Genet. 14:R243–R249. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Edgar R, Domrachev M and Lash AE: Gene
Expression Omnibus: NCBI gene expression and hybridization array
data repository. Nucleic Acids Res. 30:207–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barrett T, Wilhite SE, Ledoux P, et al:
NCBI GEO: archive for functional genomics data sets - update.
Nucleic Acids Res. 41:D991–D995. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Subramanian A, Tamayo P, Mootha VK, et al:
Gene set enrichment analysis: a knowledge-based approach for
interpreting genome-wide expression profiles. Proc Natl Acad Sci
USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He K, Chen Z, Ma Y and Pan Y:
Identification of high-copper-responsive target pathways in Atp7b
knockout mouse liver by GSEA on microarray data sets. Mamm Genome.
22:703–713. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao H, Wang Q, Bai C, He K and Pan Y: A
cross-study gene set enrichment analysis identifies critical
pathways in endometriosis. Reprod Biol Endocrinol. 7:942009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Niesen CE, Xu J, Fan X, et al:
Transcriptomic profiling of human peritumoral neocortex tissues
revealed genes possibly involved in tumor-induced epilepsy. PLoS
One. 8:e560772013. View Article : Google Scholar
|
|
11
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: affy - analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chiaretti S, Li X, Gentleman R, et al:
Gene expression profile of adult T-cell acute lymphocytic leukemia
identifies distinct subsets of patients with different response to
therapy and survival. Blood. 103:2771–2778. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gotea V and Ovcharenko I: DiRE:
identifying distant regulatory elements of co-expressed genes.
Nucleic Acids Res. 36:W133–W139. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen N, Gao Y, Yan N, et al:
High-frequency stimulation of the hippocampus protects against
seizure activity and hippocampal neuronal apoptosis induced by
kainic acid administration in macaques. Neuroscience. 256:370–378.
2014. View Article : Google Scholar
|
|
15
|
Soleman S, Filippov MA, Dityatev A and
Fawcett JW: Targeting the neural extracellular matrix in
neurological disorders. Neuroscience. 253:194–213. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Heck N, Garwood J, Loeffler JP, Larmet Y
and Faissner A: Differential upregulation of extracellular matrix
molecules associated with the appearance of granule cell dispersion
and mossy fiber sprouting during epileptogenesis in a murine model
of temporal lobe epilepsy. Neuroscience. 129:309–324. 2004.
View Article : Google Scholar
|
|
17
|
Naggar I, Lazar J, Kamran H, Orman R and
Stewart M: Relation of autonomic and cardiac abnormalities to
ventricular fibrillation in a rat model of epilepsy. Epilepsy Res.
108:44–56. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Iniesta I, Yotti R and Garcia-Pastor A:
Transient loss of consciousness with convulsions in two young
adults with potentially fatal underlying heart disease: syncope
versus seizures. BMJ Case Rep. 2009:pii: bcr06.2008.0285. 2009.
|
|
19
|
Lian XY, Khan FA and Stringer JL:
Fructose-1,6-bisphosphate has anticonvulsant activity in models of
acute seizures in adult rats. J Neurosci. 27:12007–12011. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chuang YC, Lin TK, Huang HY, et al:
Peroxisome proliferator-activated receptors γ/mitochondrial
uncoupling protein 2 signaling protects against seizure-induced
neuronal cell death in the hippocampus following experimental
status epilepticus. J Neuroinflammation. 9:1842012.
|
|
21
|
Kaminski RM, Rogawski MA and Klitgaard H:
The potential of antiseizure drugs and agents that act on novel
molecular targets as antiepileptogenic treatments.
Neurotherapeutics. 11:385–400. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hihi AK, Michalik L and Wahli W: PPARs:
transcriptional effectors of fatty acids and their derivatives.
Cell Mol Life Sci. 59:790–798. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fuenzalida K, Quintanilla R, Ramos P, et
al: Peroxisome proliferator-activated receptor gamma up-regulates
the Bcl-2 anti-apoptotic protein in neurons and induces
mitochondrial stabilization and protection against oxidative stress
and apoptosis. J Biol Chem. 282:37006–37015. 2007. View Article : Google Scholar
|
|
24
|
Adabi Mohazab R, Javadi-Paydar M, Delfan B
and Dehpour AR: Possible involvement of PPAR-gamma receptor and
nitric oxide pathway in the anticonvulsant effect of acute
pioglitazone on pentylenetetrazole-induced seizures in mice.
Epilepsy Res. 101:28–35. 2012.PubMed/NCBI
|
|
25
|
Yu X, Shao XG, Sun H, et al: Activation of
cerebral peroxisome proliferator-activated receptors gamma exerts
neuroprotection by inhibiting oxidative stress following
pilocarpine-induced status epilepticus. Brain Res. 1200:146–158.
2008. View Article : Google Scholar
|
|
26
|
Goumon Y, Angelone T, Schoentgen F, et al:
The hippocampal cholinergic neurostimulating peptide, the
N-terminal fragment of the secreted
phosphatidylethanolamine-binding protein, possesses a new
biological activity on cardiac physiology. J Biol Chem.
279:13054–13064. 2004. View Article : Google Scholar
|
|
27
|
Yeung K, Seitz T, Li S, et al: Suppression
of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature.
401:173–177. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Frayne J, Ingram C, Love S and Hall L:
Localisation of phosphatidylethanolamine-binding protein in the
brain and other tissues of the rat. Cell Tissue Res. 298:415–423.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Banfield MJ, Barker JJ, Perry AC and Brady
RL: Function from structure? The crystal structure of human
phosphatidylethanolamine-binding protein suggests a role in
membrane signal transduction. Structure. 6:1245–1254. 1998.
View Article : Google Scholar
|
|
30
|
Chen Q, Wang S, Thompson SN, Hall ED and
Guttmann RP: Identification and characterization of PEBP as a
calpain substrate. J Neurochem. 99:1133–1141. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
George AJ, Gordon L, Beissbarth T, et al:
A serial analysis of gene expression profile of the Alzheimer’s
disease Tg2576 mouse model. Neurotox Res. 17:360–379.
2010.PubMed/NCBI
|
|
32
|
Vallée BS, Coadou G, Labbé H, Sy D,
Vovelle F and Schoentgen F: Peptides corresponding to the N- and
C-terminal parts of PEBP are well-structured in solution: new
insights into their possible interaction with partners in vivo. J
Pept Res. 61:47–57. 2003.PubMed/NCBI
|