Introduction
Hepatocellular carcinoma (HCC) is the most common
form of liver cancer and the third leading cause of cancer-related
mortality worldwide (1). The
incidence of HCC has increased in recent years. The Hedgehog (Hh)
signaling pathway plays an important role in embryonic development,
as well as in the regulation of a variety of cellular functions.
Aberrant Hh signaling is associated with a variety of cancers,
including HCC. Glioma-associated oncogene 1 (Gli1) is an important
downstream effector of Hh. It acts as a transcription factor,
participating in the promotion of cell growth and the inhibition of
apoptosis. The transcription of components of the Hh signaling
pathway and related molecules has been reported to be increased in
some cases of HCC (2). Suh et
al (3) demonstrated that the
Hh signaling pathway plays a conserved role in inhibiting fat
formation. Teperino et al (4) also demonstrated that the Hh
signaling pathway stimulates metabolic reprogramming towards a
Warburg-like glycolytic state, and specifically blocks the
adipogenesis of white adipocytes, but not brown adipocytes. This
effect, mediated by a rapid non-canonical Smo-Ca2+-Ampk
signaling arm, causes robust glucose uptake in mouse and human
myocytes, and is induced by several canonical Hh signaling
inhibitors.
5′ Adenosine monophosphate (AMP)-activated protein
kinase(AMPK) plays a crucial role in cellular energy homeostasis,
and is responsive to stimulation by nutrients, stress or exercise
(5–7). The disruption of this balance is
associated with a number of diseases, such as diabetes and cancer
(8,9). AMPK is a heterotrimeric
serine/threonine protein kinase composed of 3 subunits: a catalytic
subunit (α), a scaffolding subunit (β) and an AMP-sensing subunit
(γ). Its kinase activity is controlled by the AMP/ATP ratio and
some upstream kinases, such as liver kinase B1 (LKB1),
TGF-β-activated kinase 1 (TAK1) and
Ca2+/calmodulin-dependent protein kinase kinase (CaMKK)
(10–15). AMPK controls cell metabolism and
growth in response to changes in nutrient availability by
phosphorylating a variety of substrates in cells, including
acetyl-CoA carboxylase (ACC), forkhead box O3 (FOXO3) and tuberous
sclerosis complex 2 (TSC2) (16–18). AMPK also regulates gene
transcription through direct association with chromatin and the
phosphorylation of histone H2B at serine 36 (19). However, it remains unclear as to
whether an interaction exists between the AMPK and Hh pathways.
Since both the AMPK pathway and Hh signaling pathway
affect cellular metabolism, we hypothesized that these two pathways
may interact with each other. The results of the present study
demonstrate that AMPK expression negatively correlates with the
expression of both Sonic hedgehog (Shh) and Gli1 in HCC tissues.
The treatment of HepG2 cells with smoothened agonist (SAG) or
cyclopamine (a specific inhibitor of Hh signaling) resulted in a
negative correlation between AMPK and Gli1 expression, which was
observed in a relatively short period of time (24 h or less).
Furthermore, the overexpression of AMPK induced the downregulation
of Gli1 expression, while the knockdown of AMPK upregulated Gli1
expression in a relatively short period of time (24 h or less).
Thus, AMPK may play an important role in the Hh
signaling pathway. Understanding the relationship between AMPK and
Hh signaling is important in order to elucidate the mechanisms
through which they regulate HCC pathogenesis.
Materials and methods
Cell lines, plasmids, tissue samples,
chemicals and culture media
Cell lines and culture conditions were as follows:
HepG2 (from ATCC, Manassas, VA, USA) were cultured in Dulbecco’s
modified Eagle’s medium (DMEM; HyClone, Logan, UT, USA)
supplemented with 10% (v/v) fetal bovine serum (FBS; Gibco Life
Technologies, Carlsbad, CA, USA); and 293T cells (from the National
Platform of Experimental cell Resources for Sci-Tech, Beijing,
China) were cultured in RPMI-1640 medium (HyClone) supplemented
with 10% (v/v) FBS. AMPKα1 cDNA fragments were PCR-amplified and
cloned into the pS-Flag-SBP (SBP) vector. The human Gli1 expression
vector, pcDNA3-Gli1, and the pIRES2-S-SBP-FLAG plasmid were kindly
provided by Dr Xin Zheng (Deparment of Department of Hepatobiliary
Surgery, the First Affiliated Hospital of Medical College of Xi’an
Jiaotong University, Xi’an, China). Vector PLKO was purchased from
Addgene; it is a replication-incompetent lentiviral vector selected
by the TRC (The RNAi Consortium) for the expression of shRNAs.
GFP-AMPK plasmid was also kindly provided by Dr Xin Zheng, which
had a GFP tag.
A total of 63 patients with HCC were enrolled in
this study between January 2009 and October 2009, including 49
males and 14 females who had not received pre-operative
chemotherapy or embolization. Following routine X-ray, abdominal
ultrasonography and computed tomography, all patients underwent
liver resection, including curative resection for early HCC and
palliative resection for advanced HCC. Tumor tissue and matched
adjacent normal tissue specimens (>2 cm distance to the
resection margin) were collected and immediately stored in liquid
nitrogen for RT-PCR and paraformaldehyde for immunohistochemistry.
Clinical data were obtained from the medical records. The
histopathological Edmonson classification, clinical
tumor-node-metastasis (TNM) grading, maximum tumor diameter and the
adjacent normal tissues were all confirmed by an experienced
pathologist who was blinded to the clinical information The
following clinicopathological parameters of the patients were
investigated: age, hepatitis B virus surface antigen HBsAg (+),
cirrhosis (+), serum alpha fetoprotein (AFP) levels (>400
ng/ml), tumor size (>5 cm), primary tumor (II and III), clinical
tumor-node-metastasis (TNM) grading (III and IV), differentiation,
portal vein tumor thrombosis (PVTT) (+), lymph node invasion (+),
encapsulation (+)”. Univariate and multivariate analyses of the
clinicopathological characteristics were carried out using the Cox
hazard model.
Written informed consent was obtained from all
patients enrolled in this study. The Xi’an Jiaotong University
Ethics Committee approved all protocols according to the Helsinki
Declaration of 1975.
Chemical reagents and antibodies
Anti-AMPKα1 and anti-actin antibodies were purchased
from Cell Signaling Technology (Danvers, MA, USA). Anti-Gli1,
anti-Shh and anti-HA antibodies were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA).
Immunohistochemical staining
Immunohistochemistry was performed on
paraformaldehyde-fixed paraffin sections. The sections were dewaxed
and dehydrated. Following rehydration, endogenous peroxidase
activity was blocked for 30 min using a methanol solution
containing 0.3% hydrogen peroxide. Following antigen retrieval in
citrate buffer, the sections were blocked overnight at 4°C, then
separately incubated with the primary antibodies directed against
Gli1 and AMPK at 4°C overnight. The primary antibody was detected
using biotinylated secondary antibodies (Zhongshan Golden Bridge
Biotechnology Ltd., Co., Beijing, China) according to the
manufacturer’s recommendations. The staining of the sections was
performed using the avidin-biotin-peroxidase complex for Gli1, Shh
and AMPK (SABC method). The sections were visualized using
diaminobenzidine and counterstained with hematoxylin, then
dehydrated in alcohol and xylene and mounted onto glass slides.
All sections were assessed independently by two
experienced pathologists. The staining results for the all proteins
(Gli, Shh and AMPK) were semi-quantitatively expressed by an
immunohistochemical score combined with the percentage of tumor
cells showing specific immunoreactivity. The staining intensity was
expressed in 4 grades: 0, none; 1, weak; 2, oderate; and 3, strong.
The percentage of positive carcinoma cells was expressed in the
following grades: 0, <5%; 1, 6–25%; 2, 26–50%; 3, 51–75%; and 4,
>75%. The staining intensity and average percentage of positive
tumor cells were assayed for 10 independent high magnification
(×400) fields. The total score was calculated by multiplying the
staining intensity and the percentage of positive tumor cells.
Sections with a total score of >1 were defined as exhibiting
positive staining for the above-mentioned proteins.
Cell lysis, immunoprecipitation and
western blot analysis
Cell transfection, protein extract preparation,
immunoprecipitation and western blot analysis were performed as
previously described in the study by Kim et al (20,21). Briefly, for immunoprecipitation,
the cells were lysed with ice-cold NETN buffer (20 mM Tris-HCl, pH
8.0, 100 mM NaCl, 1 mM EDTA and 0.5% Nonidet P-40) containing 10 mM
NaF and 50 mM b-glycerophosphate, and then subjected to sonication
for 12 sec. The supernatants were incubated with he indicated
antibodies and protein G-conjugated Sepharose beads (Amersham
Pharmacia Biotech Inc., Piscataway, NJ, USA). The precipitates were
washed 3 times with NETN, and subjected to SDS-PAGE and western
blot analysis with the indicated antibodies. To examine the AMPK
levels, the cell pellets were lysed with 400 μl NETN100 buffer.
Following centrifugation, the supernatants were named as 100 mM
NaCl samples. The insoluble pellets were collected, washed with
ice-cold PBS, and incubated with 400 μl NETN300 buffer on ice.
Following centrifugation, the supernatants were named as 300 mM
NaCl samples. The remaining pellets were washed twice with ice-cold
PBS and then treated with 200 μl 0.2N HCl. The supernatants were
neutralized with 40 μl 1N NaOH, and named as 0.2N HCl fractions.
Each fraction sample was loaded onto 7.5% SDS-PAGE gels for western
blot analysis with the indicated antibodies.
RNA extraction and RT-qPCR
The expression of Gli1 was determined by the reverse
transcription of total RNA, followed by quantitative PCR analysis.
Total RNA (1 μg) was reverse transcribed with random hexamers using
SuperScript II Reverse Transcriptase (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer’s instructions. Real-time PCR
was performed on a Bio-Rad iCycler using iQ SYBR-Green (Bio-Rad
Laboratories, Hercules, CA, USA) with the following primers: Gli1
forward, 5′-GAAGGTGAAGGTCGGAGT-3′ and reverse,
5′-GTCCAGGCTGGCATCCGACA-3′; and GAPDH forward,
5′-GAAGGTGAAGGTCGGAGT-3′ and reverse, 5′-GAAGA
TGGTGATGGGATTTC-3′.
DNA constructs and mutagenesis
PCR-amplified human AMPKα1 was cloned into the
pS-Flag-SBP (SBP) vector. AMPKα1 shRNA #1-resistant and shRNA
#2-resistant were created using the QuikChange Site-Directed
Mutagenesis kit (Stratagene, La Jolla, CA, USA).
AMPKα1 shRNA #1 was generated with GATGGAATAT
GTCTCAGGAGG. AMPKα1 shRNA #2 was generated with
ATGATGTCAGATGGTGAATTT. We used the following primers: AMPKα1 shRNA
#1-resistant forward, 5′-GATAT
TTTCATGGTaATGGAgTAcGTgagtGGtGGAGAGCTATT TGA and reverse,
5′-TCAAATAGCTCTCCaCCactcACg TAcTCCATtACCATGAAAATATC; and shRNA
#2-resistant forward, 5′-GGTCTTTCAAACATGATGagaGAaGGaGAgT
TcTTAAGAACAAGTTG and reverse, 5′-CAACTTGTTCTT
AAgAAcTCtCCtTCtctCATCATGTTTGAAAGACC.
GST-pull down assay
For the GST pull-down assay, 1 μg of GST-AMPK or GST
as a control was incubated with the cell lysates from the 293T
cells overexpressing HA-tagged Gli1. Glutathione beads were then
added and followed by incubation for 2 h. The bound proteins were
eluted with sample loading buffer and analyzed by immunoblotting
with HA antibodies. For endogenous immunoprecipitation, the 293T
cell lysates were immunoprecipitated with normal mouse IgG as a
control, followed by incubation with protein A beads. The bound
proteins were subjected to immunoblot analysis with AMPK
antibody.
Transfection
Tgh 293T cells were grown in DMEM containing 10%
FBS. The Cells (2×106) were seeded in 10-cm dishes 24 h
prior to transfection and transfected with 5 μg of the AMPK plasmid
using the Lipofectamine 2000 reagent (Invitrogen) according to
manufacturer’s instructions.
Statistical analysis
Throughout the study, the distribution of data
points is expressed as the mean ± standard error of the mean. The
Mann-Whitney U test was used for statistical analysis unless
otherwise indicated. The relationship between the
clinicopathological characteristics of the patients and Gli1
protein expression was analyzed using the Cox hazard model.
Additionally, the Spearman rank test was also used to analyze the
correlation between AMPK protein expression and the expression of
Shh and Gli1. Values of P<0.05 were considered to indicate
statistically significant differences.
Results
Expression of Gli1, Shh and AMPK in
HCC
To evaluate Gli1 expression in HCC, a total of 63
pairs of HCC and adjacent tissues from HCC patients were examined
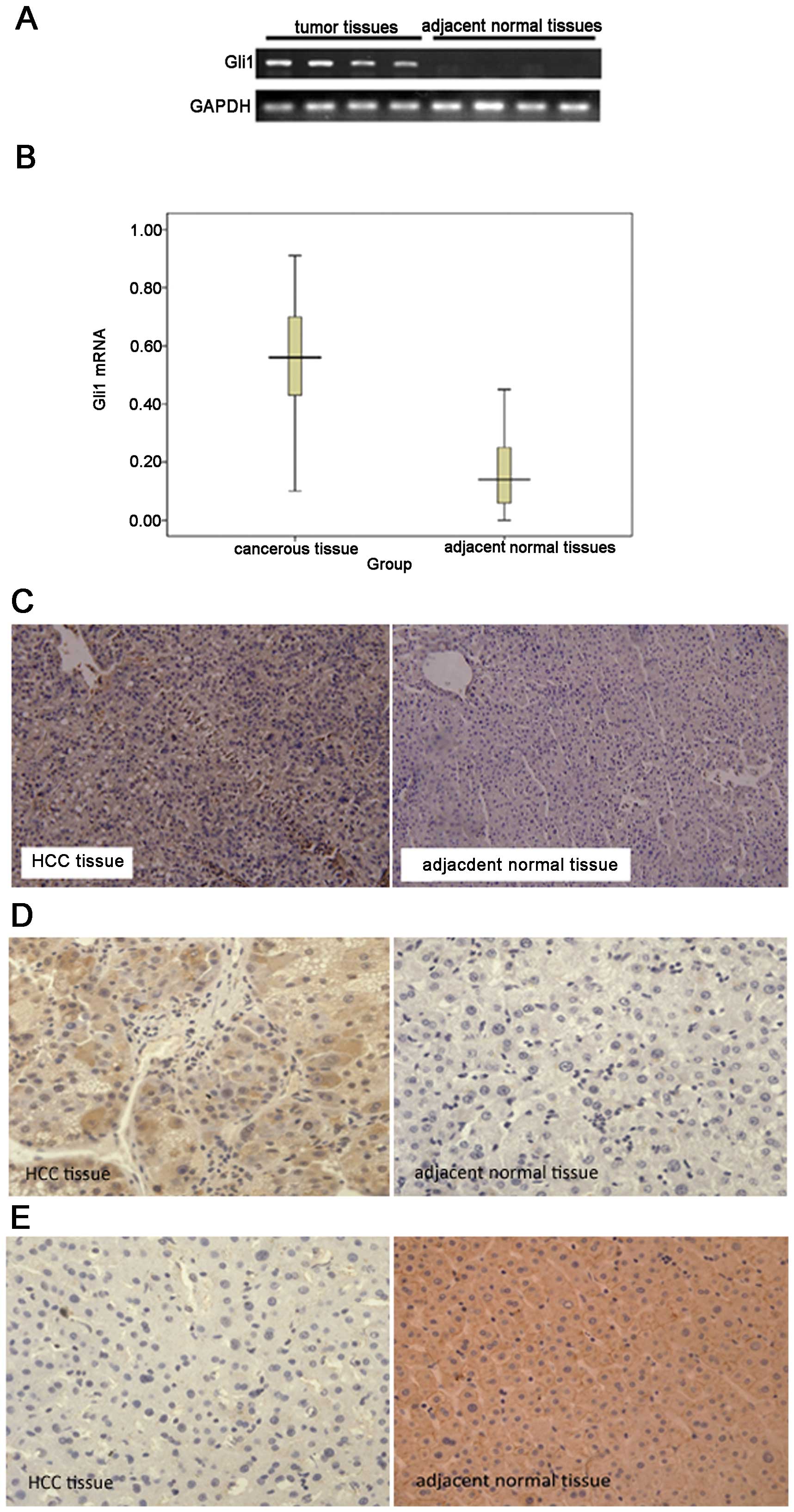
by RT-qPCR and immunohistochemistry. Gli1 mRNA was detected in 52
HCC samples, but only in 16 of the adjacent normal samples
(Fig. 1A). The expression of Gli1
mRNA in the HCC tissues was significantly higher than that in the
adjacent normal tissues (Fig.
1B). Gli1 protein levels in the HCC and adjacent normal tissues
were measured using a Gli1-specific antibody. Gli1 protein was
detected in 57 of the 63 tumor tissues (90.47%), including 17
(26.98%) highly positive (+++) cases. By contrast, Gli1 protein
expression was only observed in 21 of the 63 normal tissues
(33.33%), and none of these normal tissues was scored as highly
positive (Fig. 1C, Table I).
 | Table IExpression of Gli1 protein in the HCC
tumor tissues and adjacent normal tissues. |
Table I
Expression of Gli1 protein in the HCC
tumor tissues and adjacent normal tissues.
| | Gli1 expression
level | | | |
|---|
| |
| | | |
|---|
| Pathological
type | n | − | + | + + | + + + | Positive n (%) | χ2 | P-value |
|---|
| HCC tissues | 63 | 6 | 8 | 32 | 17 | 57 (90.47) | | |
| Adjacent normal
tissues | 63 | 42 | 18 | 3 | 0 | 21 (33.33) | 43.6 | <0.05 |
The protein expression of Shh was examined by
immunohistochemistry. As shown in Fig. 1D, Shh was mainly expressed in the
cytoplasm. Quantitative analysis indicated that the level of Shh in
the HCC tissues was significantly higher than that in the adjacent
normal tissues (P<0.001). In addition, a negative correlation
between AMPK and Shh expression was confirmed in the HCC tissues
using the Spearman rank test (r=−0.574, P<0.05).
Similarly, the expression of AMPK protein was
determined by immunohistochemistry. As shown in Fig. 1E, AMPK was mainly located on the
cytomembrane. Quantitative analysis revealed that the level of AMPK
in the HCC tissues was significantly lower than that in the
adjacent normal tissues (4.40±3.68 vs. 0.47±1.01; P<0.05).
Furthermore, the Spearman rank test revealed a
negative correlation between Gli1 and AMPK expression levels
(r=−0.429, P<0.05).
Gli1 expression correlates with the
prognosis of HCC patients
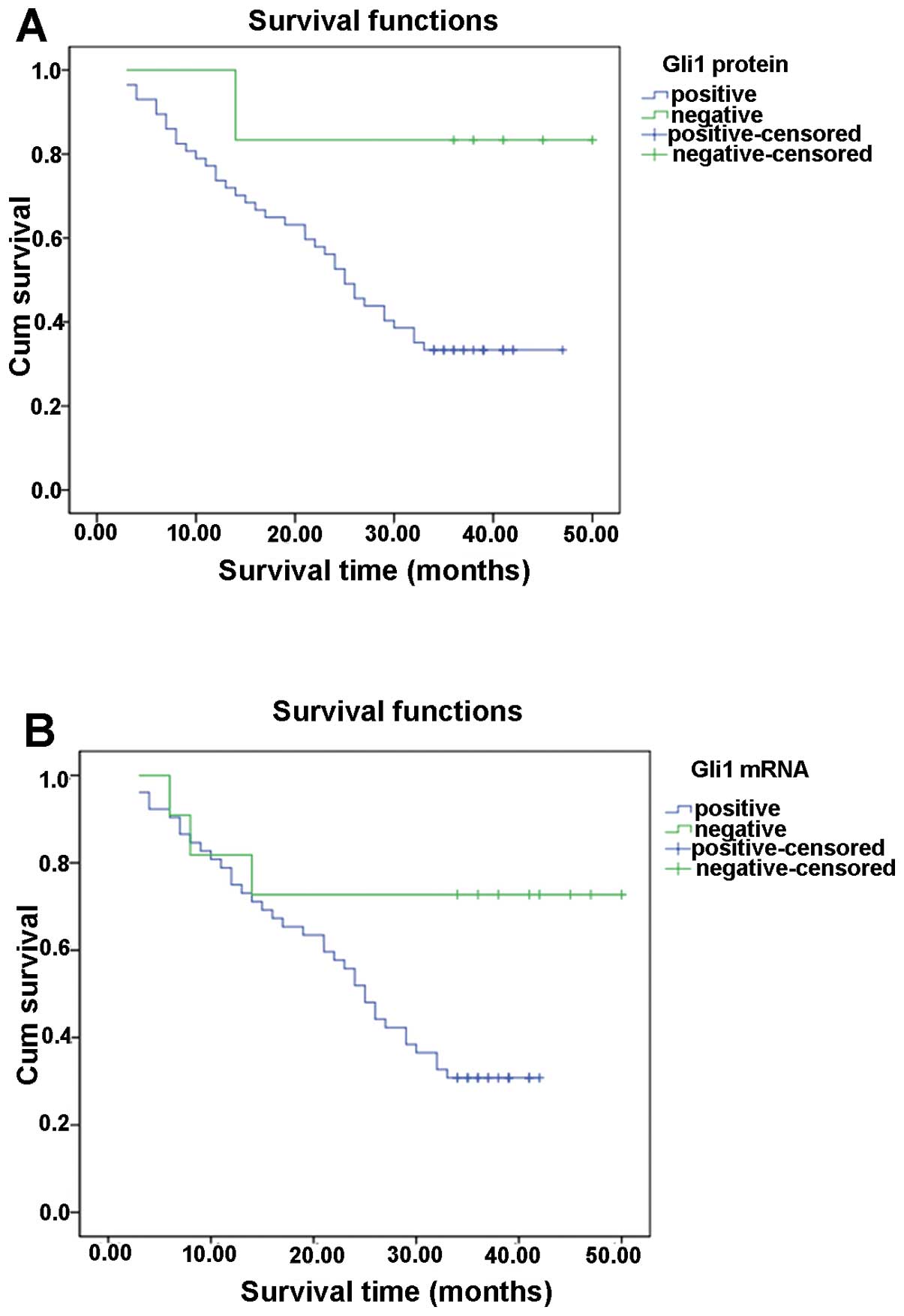
We also analyzed the correlation between Gli1
expression and patient survival, which was carried out by
univariate and multivariate analyses adjusted for
clinicopathological parameters. Cox regression analysis revealed
that there was a significant correlation between Gli1 expression
and tumor invasiveness, including histological differentiation,
portal vein tumorous thrombogenesis, lymph node invasion and TNM
stage (Table II). Using
Kaplan-Meier analysis, we found that the patients with positive
Gli1 expression had poor overall survival prognosis (Fig. 2, Table III). These results suggest that
Gli1 is a useful prognostic marker for HCC patients.
| Table IIPrognostic factors in the Cox
proportional hazards model. |
Table II
Prognostic factors in the Cox
proportional hazards model.
| Parameters | RR | 95% CI | Wald | P-value |
|---|
| Gender | 0.819 | 0.127–5.274 | 0.044 | 0.834 |
| Age | 1.033 | 0.205–5.196 | 0.002 | 0.968 |
| HBsAg | 0.310 | 0.057–1.675 | 1.851 | 0.174 |
| Cirrhosis | 0.237 | 0.033–1.721 | 2.027 | 0.155 |
| Serum AFP (>400
ng/ml) | 0.530 | 0.062–4.494 | 0.339 | 0.560 |
| Tumor size (>5
cm) | 0.728 | 0.181–2.923 | 0.200 | 0.655 |
|
Differentiation | 15.197 | 2.039–113.291 | 7.048 | 0.008a |
| PVTT | 6.041 | 1.395–26.162 | 5.784 | 0.016a |
| Lymph node
invasion | 0.032 | 0.003–0.369 | 7.627 | 0.006a |
| Encapsulation | 2.484 | 0.435–14.180 | 1.048 | 0.306 |
| Primary tumor (II
and III) | 3.105 | 0.395–24.435 | 1.159 | 0.282 |
| TNM stage (III and
IV) | 75.634 | 2.757–2075E3 | 6.554 | 0.010a |
| Gli1 mRNA | 22.298 | 2.110–235.510 | 6.663 | 0.010a |
| Table IIIUnivariate prognostic factors in
Kaplan-Meier survival analysis. |
Table III
Univariate prognostic factors in
Kaplan-Meier survival analysis.
|
Characteristics | N | Deaths | Median/M | SE | Log-rank | P-value |
|---|
| Gender |
| Male | 49 | 31 | 26.00 | 3.50 | | |
| Female | 14 | 8 | 25.58 | 3.74 | 0.124 | 0.725 |
| Age (years) |
| ≤45 | 15 | 10 | 24.00 | 7.24 | | |
| >45 | 48 | 29 | 26.00 | 2.47 | 0.201 | 0.654 |
| HBsAg |
| Positive | 50 | 33 | 25.00 | 2.80 | | |
| Negative | 13 | 6 | 29.00 | 3.73 | 0.933 | 0.334 |
| Cirrhosis |
| Positive | 51 | 36 | 24.00 | 2.40 | | |
| Negative | 11 | 3 | 30.00 | 2.83 | 5.466 | 0.019a |
| Serum
AFP/(ng/ml) |
| ≤400 | 16 | 7 | 31.00 | 4.27 | | |
| >400 | 47 | 32 | 25.00 | 2.06 | 2.500 | 0.114 |
| Tumor size
(cm) |
| ≤5 | 21 | 13 | 28.00 | 6.29 | | |
| >5 | 42 | 26 | 26.00 | 2.69 | 0.004 | 0.953 |
|
Differentiation |
| Well and moderate
(I + II) | 49 | 25 | 33.00 | 2.46 | | |
| Poor and none (III
+ IV) | 14 | 14 | 12.00 | 2.49 | 18.669 | 0.000a |
| PVTT |
| Positive | 21 | 18 | 21.00 | 5.34 | | |
| Negative | 42 | 21 | 29.00 | 2.63 | 4.924 | 0.026a |
| Lymph node
invasion | | | | | | |
| Positive | 27 | 22 | 19.00 | 4.33 | | |
| Negative | 36 | 17 | 26.00 | 2.84 | 6.870 | 0.009a |
| Encapsulation |
| Positive | 40 | 29 | 24.00 | 2.37 | | |
| Negative | 23 | 10 | 28.00 | 3.11 | 3.645 | 0.056 |
| Primary tumor |
| T1+T2 | 19 | 6 | 40.00 | 3.51 | | |
| T3+T4 | 44 | 33 | 23.00 | 3.27 | 10.548 | 0.001a |
| TNM stage |
| I+II | 19 | 2 | 45.00 | 2.75 | | |
| III+IV | 44 | 37 | 21.00 | 3.32 | 23.612 | 0.000a |
| Gli1 protein |
| Positive | 57 | 38 | 24.00 | 2.15 | | |
| Negative | 6 | 1 | 41.00 | 3.19 | 3.996 | 0.046a |
| Gli1 mRNA |
| Positive | 52 | 36 | 25.00 | 1.80 | | |
| Negative | 11 | 3 | 38.00 | 2.81 | 4.230 | 0.040a |
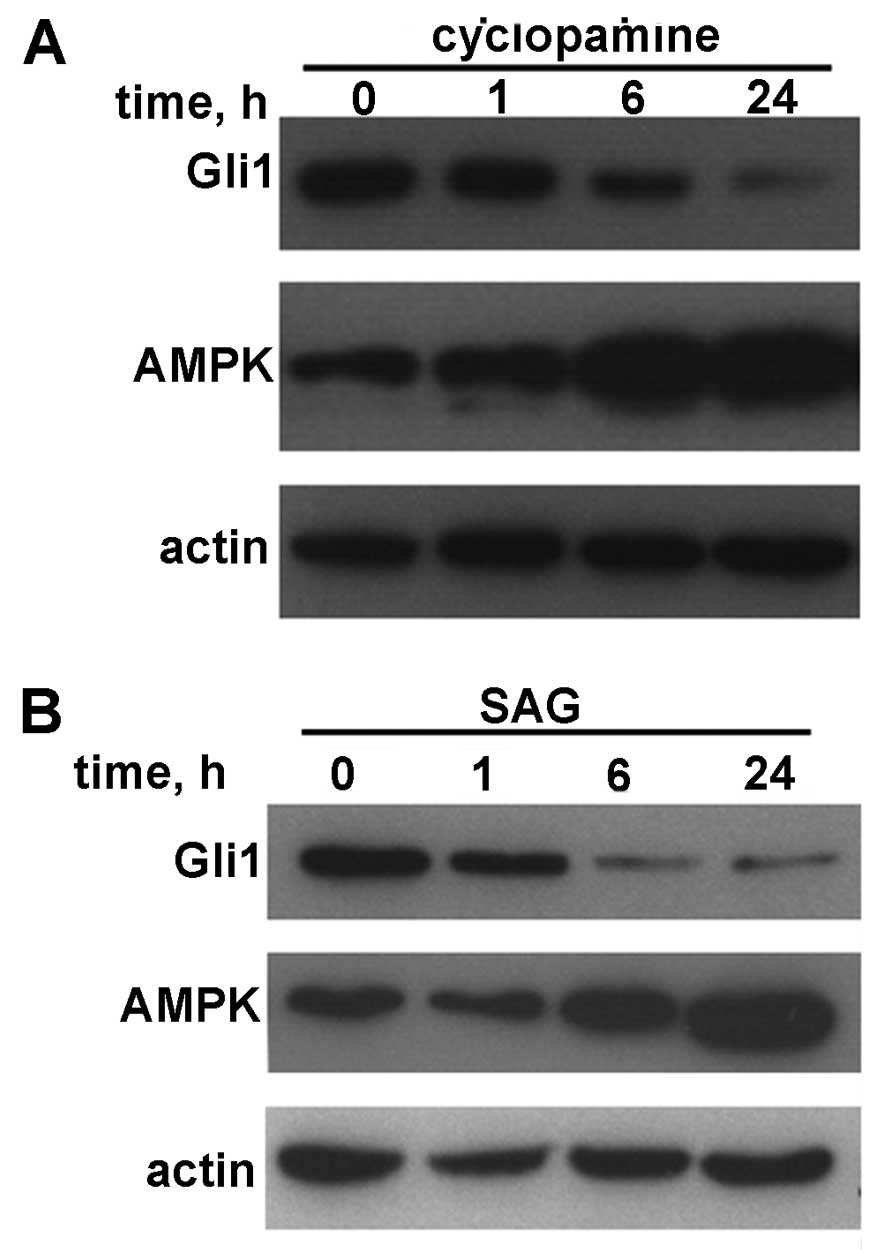
Correlation between AMPK and Gli1 is
reversed by treatment with cyclopamine or SAG
The HepG2 cells were treated with cyclopamine (a
specific inhibitor of Hh signaling) or SAG. The cell lysates were
then immunoblotted with Gli1 and AMPK antibodies. As shown in
Fig. 3, a negative correlation
between AMPK and Gli1 was observed in a relatively short period of
time (24 h or less).
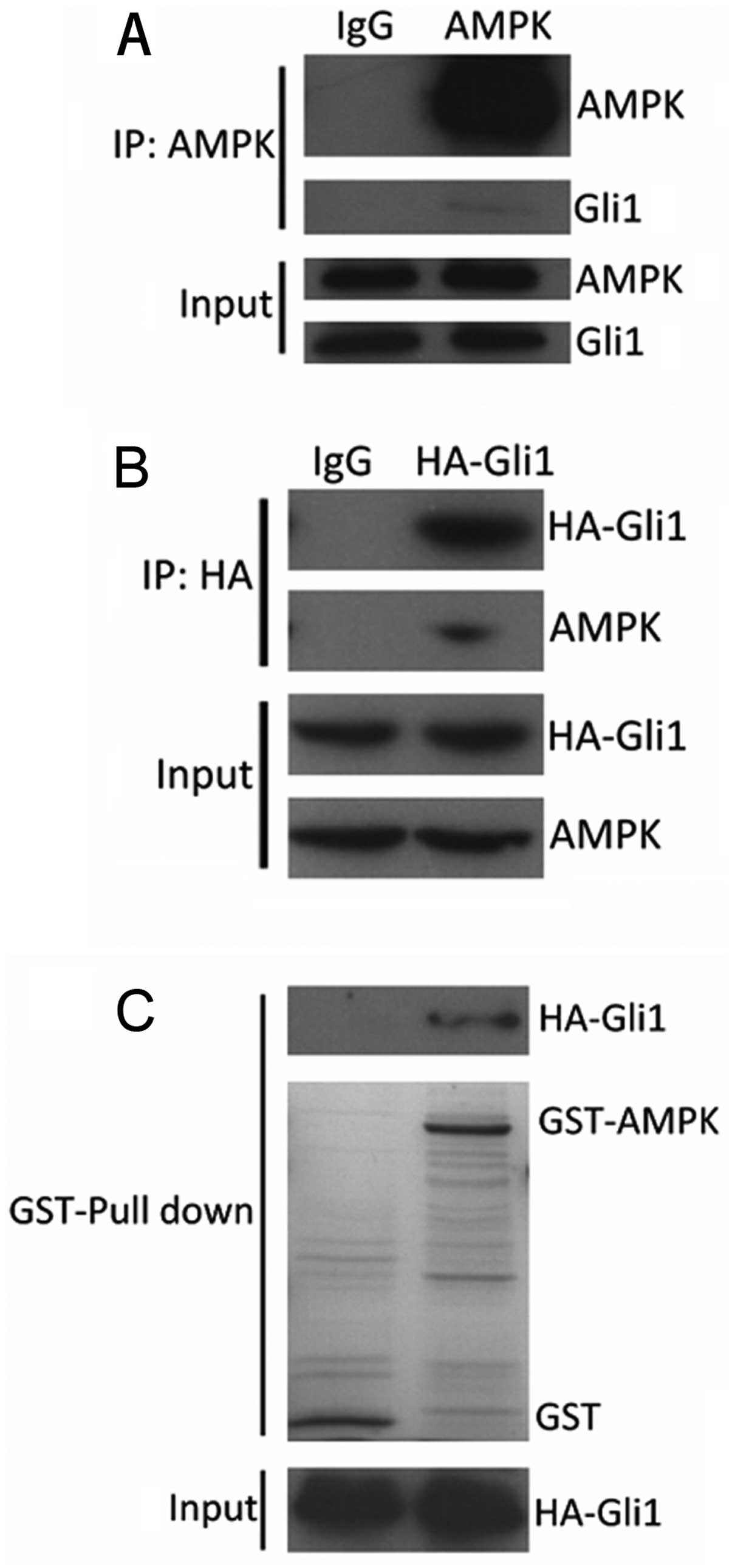
Gli1 directly interacts with AMPK
To further understand the functional relationship
between AMPK and Gli1, we examined the physical interaction between
these two molecules using immunoprecipitation. As shown in Fig. 4A, Gli1 protein was precipitated
from the 293T cells lysates using AMPK-specific antibody. We also
examined whether AMPK can be precipitated by Gli1 antibody. As
Gli1-specific antibody for immunoprecipitation is not available, we
transfected the 293T cells with HA-Gli1. The cell lysates were
subjected to incubation with HA antibody and immunoblotted with
AMPK antibody. We found that HA-Gli1 interacted with AMPK (Fig. 4B). To examine the direct
interaction between AMPK and Gli1, GST-AMPK and HA-Gli1 were
purified for the GST pull-down assay. As shown in Fig. 4C, GST-AMPK interacted with HA-Gli1
in the 293T cells. These results indicated that AMPK interacted
with Gli1 in vitro.
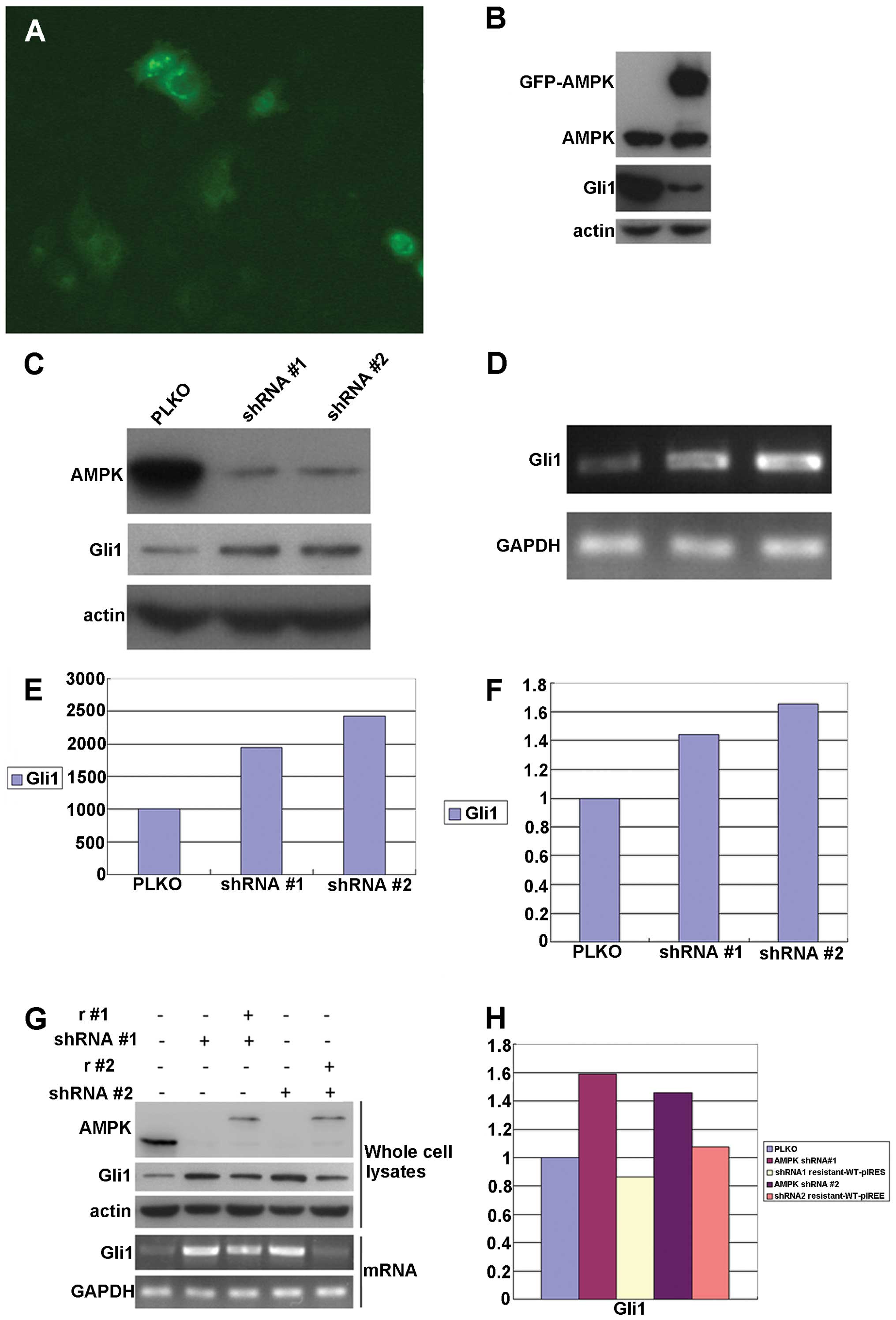
AMPK regulates Gli1 expression
To investigate whether AMPK regulates Gli1
expression, AMPK was overexpressed in the HepG2 cells by
transfecting the cells with the pIRES-S-SBP-FLAG-AMPK vector. Gli1
expression was examined in the transfected cells (Fig. 5A). It was found that Gli1
expression was significantly downregulated in the
AMPK-overexpressing cells (Fig.
5B). We also examined the effects of AMPK downregulation by
shRNA knockdown. The HepG2 cells were transfected with recombinant
lentiviruses expressing either AMPK shRNA #1 or shRNA #2 or vector
PLKO as a control. Both ‘targeted’ shRNAs decreased AMPK expression
in the HepG2 cells (Fig. 5C;
>80% knockdown). AMPK knockdown significantly increased the
levels of Gli1 mRNA and protein (Fig.
5C–F).
To further verify the specificity of AMPK shRNAs and
the regulatory role of AMPK in Gli1 expression, a mutated version
of AMPK was expressed in the HepG2 cells. This AMPK mutant is
resistant to AMPK shRNAs, but can express functional AMPK in HepG2
cells. We found that Gil1 expression in the HepG2 cells was
completely rescued by the expression of AMPK mutant. The effects
were observed at both the mRNA and protein level (Fig. 5G and H). These results confirmed
that Gli1 was specifically regulated by AMPK.
Discussion
The Hh signaling pathway plays an important role in
embryonic development, as well as in the regulation of a variety of
cellular functions (22–25). Aberrant Hh signaling is associated
with a variety of cancers, including glioma (26,27), glioblastoma (28,29), breast cancer (30,31) and pancreatic adenocarcinoma
(32,33) among others (34–36).
In mammals, the Hh signaling pathway mainly consists
of Hh ligands [Desert hedgehog (Dhh), Indian hedgehog (Ihh) and
Shh], two transmembrane proteins [patched 1 (Ptch) and smoothened,
frizzled class receptor (Smo)], the nuclear transcription factor,
Gli (Gli1, Gli2 and Gli3), and downstream target genes. When this
pathway is activated, ligand binding to the inhibitory cell surface
receptor, Ptch, initiates Smo translocation to the tip of the
cilium and liberates the zinc finger family transcription factors,
Gli1, Gli2 and Gli3, from proteasomal degradation and inhibitory
tethering, leading to the activation of Gli target genes (37–41), which exert further effects on cell
growth, proliferation and differentiation.
In the present study, using immunohistochemical
analysis, we first confirmed that the Shh and Gli1 proteins were
expressed at significantly higher levels in the HCC tissues than in
the adjacent normal tissues. This result suggests that Hh
signaling, which plays a role in liver embryonic development and
differentiation, was aberrantly activated and that this may lead to
tumorgenesis in HCC.
In addition, we analyzed the correlation between
Gli1 expression and tumor pathological characteristics and patient
prognosis. The results revealed that Gli1 expression in the highly
aggressive (including histological differentiation, portal vein
tumorous thrombogenesis, lymph node invasion and TNM stage) HCC
group was significantly higher than that in the less aggressive HCC
group. Furthermore, there was a significant negative correlation
between Gli1 expression and patient prognosis. These results
suggest that Hh signaling plays a critical role in
hepatocarcinogenesis. However, the exact molecular mechanisms of Hh
signaling in HCC progression remain unclear.
It is known that cancer cells show enhanced
glycolysis and inhibition of oxidative phosphorylation, even in the
presence of sufficient oxygen (aerobic glycolysis) (42). AMPK is a key regulator of cellular
metabolism and is involved in the pathogenesis of several diseases
(8,9). However, to the best of our
knowlwedge, there is no report to date on the possible interactions
between AMPK and Hh signaling.
Recent studies have indicated that Hh signaling
rewires cellular metabolism. There is a cilium-dependent
Smo-Ca2+-Ampk axis that triggers rapid Warburg-like
metabolic reprogramming within minutes of activation, which is
required for proper metabolic selectivity and flexibility. In a
previous study, Teperino et al (4) demonstrated that Smo modulators can
uncouple the Smo-Ampk axis from the canonical signaling pathway,
and identified cyclopamine as one of the new class of ‘selective
partial agonists’, capable of the concomitant inhibition of
canonical Hh signaling and the activation of non-canonical Hh
signaling.
The present study revealed a negative correlation
between AMPK and Gli1 expression. Of note, when the HepG2 cells
were treated with cyclopamine or SAG, the Gli1 expression level
decreased, but the AMPK expression level increased in a relatively
short period of time (24 h or less). Furthermore, the
overexpression of AMPK downregulated Gli1 expression, while the
knockdown of AMPK upregulated Gli1 expression in a relatively short
period of time (24 h or less). On the basis of these findings, we
suggest that AMPK may play a role in the Hh signaling pathway by
interacting with Shh and Gli1, and this interaction may affect
cellular metabolism and tumorgenesis. Further studies are required
to elucidate the molecular mechanisms involved in this process.
References
|
1
|
Yang JD and Roberts LR: Hepatocellular
carcinoma: a global view. Nat Rev Gastroenterol Hepatol. 7:448–458.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Patil MA, Zhang J, Ho C, Cheung ST, Fan ST
and Chen X: Hedgehog signaling in human hepatocellular carcinoma.
Cancer Biol Ther. 5:111–117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Suh JM, Gao X, McKay J, McKay R, Salo Z
and Graff JM: Hedgehog signaling plays a conserved role in
inhibiting fat formation. Cell Metab. 3:25–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Teperino R, Amann S, Bayer M, et al:
Hedgehog partial agonism drives Warburg-like metabolism in muscle
and brown fat. Cell. 151:414–426. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sun Y, Connors KE and Yang DQ: AICAR
induces phosphorylation of AMPK in an ATM-dependent,
LKB1-independent manner. Mol Cell Biochem. 306:239–245. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stockebrand M, Sauter K, Neu A, Isbrandt D
and Choe CU: Differential regulation of AMPK activation in leptin-
and creatine-deficient mice. FASEB J. 27:4147–4156. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McFadden SA, Menchella JA, Chalmers JA,
Centeno ML and Belsham DD: Glucose responsiveness in a novel
adult-derived GnRH cell line, mHypoA-GnRH/GFP: involvement of
AMP-activated protein kinase. Mol Cell Endocrinol. 377:65–74. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Faubert B, Boily G, Izreig S, et al: AMPK
is a negative regulator of the Warburg effect and suppresses tumor
growth in vivo. Cell Metab. 17:113–124. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hardie DG: AMPK: a target for drugs and
natural products with effects on both diabetes and cancer.
Diabetes. 62:2164–2172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakagawa K, Uehata Y, Natsuizaka M, et al:
The nuclear protein Artemis promotes AMPK activation by stabilizing
the LKB1-AMPK complex. Biochem Biophys Res Commun. 427:790–795.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Momcilovic M, Hong SP and Carlson M:
Mammalian TAK1 activates Snf1 protein kinase in yeast and
phosphorylates AMP-activated protein kinase in vitro. J Biol Chem.
281:25336–25343. 2006. View Article : Google Scholar
|
|
12
|
Hawley SA, Pan DA, Mustard KJ, et al:
Calmodulin-dependent protein kinase kinase-beta is an alternative
upstream kinase for AMP-activated protein kinase. Cell Metab.
2:9–19. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Herrero-Martin G, Hoyer-Hansen M,
Garcia-Garcia C, et al: TAK1 activates AMPK-dependent
cytoprotective autophagy in TRAIL-treated epithelial cells. EMBO J.
28:677–685. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Polizio AH, Chinchilla P, Chen X, Kim S,
Manning DR and Riobo NA: Heterotrimeric Gi proteins link Hedgehog
signaling to activation of Rho small GTPases to promote fibroblast
migration. J Biol Chem. 286:19589–19596. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cantó C and Auwerx J: Calorie restriction:
is AMPK a key sensor and effector? Physiology. 26:214–224.
2011.PubMed/NCBI
|
|
16
|
Sato A, Sunayama J, Okada M, et al:
Glioma-initiating cell elimination by metformin activation of FOXO3
via AMPK. Stem Cells Transl Med. 1:811–824. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gwinn DM, Shackelford DB, Egan DF, et al:
AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol
Cell. 30:214–226. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi YK and Park KG: Metabolic roles of
AMPK and metformin in cancer cells. Mol Cells. 36:279–287. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bungard D, Fuerth BJ, Zeng PY, et al:
Signaling kinase AMPK activates stress-promoted transcription via
histone H2B phosphorylation. Science. 329:1201–1205. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim JE, Chen J and Lou Z: DBC1 is a
negative regulator of SIRT1. Nature. 451:583–586. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim H, Chen J and Yu X: Ubiquitin-binding
protein RAP80 mediates BRCA1-dependent DNA damage response.
Science. 316:1202–1205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ruiz i Altaba A, Sanchez P and Dahmane N:
Gli and hedgehog in cancer: tumours, embryos and stem cells. Nat
Rev Cancer. 2:361–372. 2002.PubMed/NCBI
|
|
23
|
Ingham PW and McMahon AP: Hedgehog
signaling in animal development: paradigms and principles. Genes
Dev. 15:3059–3087. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chari NS and McDonnell TJ: The sonic
hedgehog signaling network in development and neoplasia. Adv Anat
Pathol. 14:344–352. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Elia D, Madhala D, Ardon E, Reshef R and
Halevy O: Sonic hedgehog promotes proliferation and differentiation
of adult muscle cells: involvement of MAPK/ERK and PI3K/Akt
pathways. Biochim Biophys Acta. 1773:1438–1446. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Clement V, Sanchez P, de Tribolet N,
Radovanovic I and Ruiz i Altaba A: HEDGEHOG-GLI1 signaling
regulates human glioma growth, cancer stem cell self-renewal, and
tumorigenicity. Curr Biol. 17:165–172. 2007. View Article : Google Scholar
|
|
27
|
Zbinden M, Duquet A, Lorente-Trigos A,
Ngwabyt S-N, Borges I and i Altaba AR: NANOG regulates glioma stem
cells and is essential in vivo acting in a cross-functional network
with GLI1 and p53. EMBO J. 29:2659–2674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lo HW, Zhu H, Cao X, Aldrich A and
Ali-Osman F: A novel splice variant of GLI1 that promotes
glioblastoma cell migration and invasion. Cancer Res. 69:6790–6798.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rossi M, Magnoni L, Miracco C, et al:
β-catenin and Gli1 are prognostic markers in glioblastoma. Cancer
Biol Ther. 11:753–761. 2011.
|
|
30
|
Goel HL, Pursell B, Chang C, et al: GLI1
regulates a novel neuropilin-2/α6β1 integrin based autocrine
pathway that contributes to breast cancer initiation. EMBO Mol Med.
5:488–508. 2013.PubMed/NCBI
|
|
31
|
ten Haaf A, Bektas N, von Serenyi S, et
al: Expression of the glioma-associated oncogene homolog (GLI) 1 in
human breast cancer is associated with unfavourable overall
survival. BMC Cancer. 9:2982009.PubMed/NCBI
|
|
32
|
Nolan-Stevaux O, Lau J, Truitt ML, et al:
GLI1 is regulated through Smoothened-independent mechanisms in
neoplastic pancreatic ducts and mediates PDAC cell survival and
transformation. Genes Dev. 23:24–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Inaguma S, Riku M, Hashimoto M, Saga S,
Ikeda H and Kasai K: GLI1 interferes with the DNA mismatch repair
system in pancreatic cancer through BHLHE41-mediated suppression of
MLH1. Cancer Res. 73:7313–7323. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pasca di Magliano M and Hebrok M: Hedgehog
signalling in cancer formation and maintenance. Nat Rev Cancer.
3:903–911. 2003.PubMed/NCBI
|
|
35
|
Liao X, Siu MK, Au CW, et al: Aberrant
activation of hedgehog signaling pathway in ovarian cancers: effect
on prognosis, cell invasion and differentiation. Carcinogenesis.
30:131–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rubin LL and de Sauvage FJ: Targeting the
Hedgehog pathway in cancer. Nat Rev Drug Discov. 5:1026–1033. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hui CC and Angers S: Gli proteins in
development and disease. Annu Rev Cell Dev Biol. 27:513–537. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ingham PW, Nakano Y and Seger C:
Mechanisms and functions of Hedgehog signalling across the metazoa.
Nat Rev Genet. 12:393–406. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Riobo N and Manning D: Pathways of signal
transduction employed by vertebrate Hedgehogs. Biochem J.
403:369–379. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rohatgi R and Scott MP: Patching the gaps
in Hedgehog signalling. Nat Cell Biol. 9:1005–1009. 2007.
View Article : Google Scholar
|
|
41
|
Ruiz i Altaba A, Mas C and Stecca B: The
Gli code: an information nexus regulating cell fate, stemness and
cancer. Trends Cell Biol. 17:438–447. 2007.
|
|
42
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|