Introduction
Currently the number of patients suffering from
degenerative bone diseases, such as osteoporosis and osteoarthritis
is increasing due to the aging population (1). In addition, fracture non-union
remains a clinically important issue. Despite substantial advances
in orthopedic surgery, some fractures fail to heal effectively and
result in either delayed union or non-union, causing morbidity,
prolonged hospitalization and increased expenses. To address these
issues, several interventions, including bone autografts have been
exploited to improve the capability of bone tissue regeneration,
leading to the reduction of both the total costs and
hospitalization period (2).
However, the use of autografts is associated with certain major
issues, including donor site morbidity and limited availability
(3). Advances in stem cell
research have prompted the development of cell-based therapy for
bone repair and the treatment of bone diseases (4,5).
Over the past 10 years, tissue engineering using mesenchymal
stromal cells (MSCs) has gained increasing attention in
regenerative medicine, due to the ability of these cells to
differentiate not only into cells of the mesodermal lineage, but
also into endodermal and ectodermal lineages (6,7).
The characteristics of differentiation in a tissue-specific manner
hold great promise for the use of MSCs in the field of regenerative
medicine. MSC therapy has been considered to be a potential
treatment for several bone diseases (2,8).
The use of MSCs for fracture repair has been successfully examined
using animal models (9). A
previous study reported that autologous bone marrow grafting is an
effective and safe method for treating an atrophic tibial
diaphyseal non-union (10).
Although the progress in the process of osteogenic differentiation
of MSCs has been reported, the restricted quantity and quality of
functional osteocytes are the main obstacles for the therapeutic
application. The identification of novel agents that are able to
enhance the osteogenic differentiation capacity of MSCs is thus
necessary.
During normal fracture healing, undifferentiated
MSCs, with the aid of bone morphogenetic proteins (BMPs) and
regulatory cytokines, proliferate, and differentiate into
chondrocytes and osteoblasts, and form bone, thereby repairing the
injury (11). A previous study
reported that BMP-2 can stimulate the entire process of osteogenic
differentiation of bone marrow-derived MSCs (BM-MSCs) in
vitro (12). Clinical
orthopedic studies have shown the benefits of BMP-2 in bone tissue
regeneration. In addition, some studies have supported the use of
BMP-2 in periodontal regeneration, sinus lift bone-grafting, and
non-unions in bone surgery (13,14). Although MSCs derived from
different sources have been assumed to exhibit similar
characteristics to MSCs derived from bone marrow, some differences
at least in terms of the osteogenic differentiation ability have
been reported. MSCs derived from the umbilical cord can be
differentiated into osteoblasts with a phenotypic similarity to
that of BM-MSCs; however, the differentiation ability is not
consistent. In addition, MSCs from the umbilical cord require a
longer period of time to differentiate into osteoblasts (15). Although the use of BMP-2 for bone
tissue regeneration has been extensively investigated (16–18), the BMP-2-induced osteogenic
differentiation of MSCs derived from the umbilical cord has not
been fully examined, in particular as regards the underlying
molecular events governing osteogenic differentiation.
Thus, in this study, we aimed to examine the effect
of BMP-2 on the osteogenic differentiation of MSCs derived from
umbilical cord compared to that of MSCs derived from bone marrow.
The underlining mechanisms, such as the expression of alkaline
phosphatase (ALP) and the changes in the expression of
transcription factors involved in the BMP-2-induced osteogenic
differentiation of these MSCs were also examined. Our data provide
new insight into the effects of BMP-2 on the osteogenic
differentiation of MSCs derived from bone marrow and umbilical
cord, which may lead to the development of advance techniques for
bone tissue regeneration in the future. Our findings also indicate
the potential for using these MSCs as alternative sources for bone
engineering or cell therapy in regenerative medicine.
Materials and methods
Cell isolation and culture
The present study was approved by the Human Ethics
Committee of Thammasat University No. 1 (Faculty of Medicine;
MTU-EC-DS-1-061-57). All subjects participated in the study after
providing written informed consent. Bone marrow (BM) was aspirated
from healthy volunteers (n=5). Mononuclear cells (MNCs) were
isolated using Ficoll-Hypaque solution. BM-MNCs were then cultured
in Dulbecco's modified Eagle's medium (DMEM; Gibco-BRL, Carlsbad,
CA, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen,
Carlsbad, CA, USA), 2 mM L-glutamine, 100 U/ml penicillin and 100
µg/ml streptomycin (both from Gibco-BRL). The medium was
changed every 3–4 days. The plastic adherent fibroblast-like cells
or so-called BM-MSCs (approximately 80–90% confluence) were
sub-cultured using 0.25% trypsin-EDTA (Gibco-BRL) and replated at
density of 1×104 cell/cm2 for further
expansion.
The umbilical cord tissues from full-term
pregnancies (n=5) were minced into small sections and digested with
1.6 mg/ml collagenase XI and 200 mg/ml deoxyribonuclease I (both
from Sigma-Aldrich, St. Louis, MO, USA) for 4 h at 37°C.
Subsequently, the cells were washed twice with washing buffer and
cultured in complete medium containing DMEM supplemented with FBS,
2 mM L-glutamine, 100 U/ml penicillin and 100 µg/ml
streptomycin. The medium was changed every 3–4 days and
non-adherent cells were removed. The adherent cells were further
cultured under the same conditions until obtaining the colonies of
fibroblast-like cells. For further expansion, the cells were
sub-cultured using 0.25% trypsin-EDTA and replated at density of
1×104 cells/cm2.
Characterization of MSCs using flow
cytometry
The MSCs derived from bone marrow and umbilical cord
were trypsinized and re-suspended with phosphate-buffered saline
(PBS). In each sample, 5×105 MSCs in 50 µl of PBS
were incubated with 2.5 µl of fluorescein isothiocyanate
(FITC) or phycoerythrin (PE) conjugated antibodies against human
antigens, including FITC-CD45 antibody (Cat. no. 304006), FITC-CD90
antibody (Cat. no. 328108) (both from BioLegend, San Diego, CA,
USA), PE-CD105 antibody (Cat. no. 560839; BD Bioscience, Franklin
Lakes, NJ, USA), PE-CD34 (Cat. no. 343506) antibody and PE-CD73
antibody (Cat. no. 344004) (both from BioLegend) for 30 min at 4°C
in the dark. After washing with PBS, the cells were fixed with 1%
paraformaldehyde in PBS. At least 10,000 labeled cells were
acquired and analyzed using a flow cytometer (FACSCalibur™) and
CellQuest® software (both from Becton-Dickinson,
Franklin Lakes, NJ, USA)
Osteogenic and adipogenic differentiation
assay
For osteogenic differentiation, the MSCs were seeded
in 35-mm2 dishes at a density of 5×103
cells/cm2 and allowed to adhere to the dishes overnight.
Subsequently, the cells were washed with PBS and 2 ml of osteogenic
differentiation medium [DMEM supplemented with 10% FBS, 100 U/ml
penicillin, 100 µg/ml streptomycin, 0.5 mM
isobutyl-methylxanthine, 0.1 µM dexamethasone, 50
µg/ml ascorbic acid (all from Sigma-Aldrich)] was added. On
day 7 of culture, 10 µM β-glycerophosphate (Sigma-Aldrich),
was added. On day 21 of culture, the cells were harvested for
Alizarin Red S staining (Sigma-Aldrich) to determine bone matrix
mineralization.
For adipogenic differentiation, the MSCs were
cultured with adipogenic differentiation medium [DMEM supplemented
with 10% FBS, 100 U/ml penicillin, 100 µg/ml streptomycin,
0.5 mM isobutyl-methylxanthine, 1 µM dexamethasone, 10
µM insulin, 100 µM indomethacin (all from
Sigma-Aldrich)] at a density of 7.8×102
cells/cm2. The medium was changed every 4 days. On day
21 of culture, the cells were fixed in 10% buffered formalin for 30
min at room temperature, washed with PBS and incubated with 2% Oil
Red O (Sigma-Aldrich) for 1 h. The cells were washed with distilled
H2O and observed under an inverted microscope (Nikon
TS100; Nikon, Tokyo, Japan). Adipogenic cells were visualized as
red-stained lipid droplets. Control cultures without the
differentiation stimuli were carried out in parallel to the
experiments and stained in the same manner.
Treatment of MSCs with BMP-2
To examine the effect of BMP-2 on the osteogenic
differentiation of MSCs, 9.5×103 MSCs at passage 4 were
cultured in 24-well plate (Corning, Inc., Corning, NY, USA) with
osteogenic differentiation medium supplemented with 100 ng/ml of
BMP-2 (R&D Systems, Minneapolis, MN, USA). MSCs cultured with
MSC complete medium and osteogenic differentiation medium were used
as controls. Following incubation for 3, 7, 14, 21 and 28 days, the
cell differentiation was measured by qualitative cytochemical
staining for ALP using 5-bromo-4-chloro-3-indolylphosphate/nitro
blue tetrazolium (BCIP/NBT; Sigma-Aldrich) as a substrate.
ALP activity assay
For quantitative ALP activity, the MSCs were
cultured with osteogenic differentiation medium supplemented with
100 ng/ml of BMP-2 for 3, 7, 14, 21 and 28 days. MSCs cultured with
MSC complete medium and osteogenic differentiation medium were used
as controls. The colorimetric ALP activity assay was performed
using the SensoLyte® pNPP Alkaline Phosphatase assay kit
(AnaSpec, Inc., Fremont, CA, USA) according to the manufacturer's
instructions. Briefly, the cells were washed twice and
permeabilized with 0.2% Triton X-100. Subsequently, the cells were
incubated with p-nitrophenyl phosphate (pNPP) chromogenic substrate
solution for 45 min at room temperature. The final solution yielded
a yellow-colored product. The stop solution was added and the
absorbance was measured at 405 nm using a micro-plate reader
(BioTex, Winooski, VT, USA). The total protein concentration
(mg/ml) was determined biochemically using the bicinchoninic acid
(BCA) assay kit (Sigma-Aldrich). A standard curve was prepared
using bovine serum albumin (BSA). ALP levels were normalized
against the total protein content. The measured ALP activity was
expressed as ng/mg protein.
Gene expression analysis by reverse
transcription-quantitative PCR (RT-qPCR)
To examine the expression of osteogenic
differentiation markers following treatment with BMP-2, total RNA
was extracted from the MSCs cultured with osteogenic
differentiation medium supplemented with 100 ng/ml of BMP-2 for 3,
7, 14, 21 and 28 days using TRIzol® reagent
(Invitrogen). Two micrograms of total RNA were reverse transcribed
into cDNA using SuperScript® III Reverse Transcriptase
(Invitrogen). The synthesized cDNAs were subjected to qPCR using
the ABI 7500 Real-time PCR system (Applied Biosystems, Foster City,
CA, USA). The PCR conditions were as follows: initial denaturation
at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C
for 10 sec, annealing at 60°C for 10 sec, and extension at 72°C for
40 sec. Finally, all PCR product quantifications were normalized to
the endogenous control gene, glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) using the StepOne™ Software version
2.2 (Applied Biosystems) and presented as the relative mRNA
expression level. MSCs cultured with MSC complete medium and
osteogenic differentiation medium were used as controls. The primer
sequences are listed in Table
I.
 | Table ISequences of the primers used for PCR
and the product size. |
Table I
Sequences of the primers used for PCR
and the product size.
| Gene | Forward primer | Reverse primer | Product size
(bp) |
|---|
| Runx2 |
5′-GACAGCCCCAACTTCCTGT-3′ |
5′-CCGGAGCTCAGCAGAATAAT-3′ | 159 |
| Osterix |
5′-TGCTTGAGGAGGAAGTTCAC-3′ |
5′-CTGCTTTGCCCAGAGTTGTT-3′ | 114 |
|
Osteocalcin |
5′-CTCACACTCCTCGCCCTATT-3′ |
5′-TCAGCCAACTCGTCACAGTC-3′ | 245 |
| GAPDH |
5′-CAATGACCCCTTCATTGACC-3′ |
5′-TTGATTTTGGAGGGATCTCG-3′ | 159 |
Statistical analysis
The data were presented as the means ± standard
error of mean (SEM). Statistical comparisons were performed using
the unpaired t-test. A P-value <0.05 was considered to indicate
a statistically significant difference.
Results
Characteristic of the MSCs
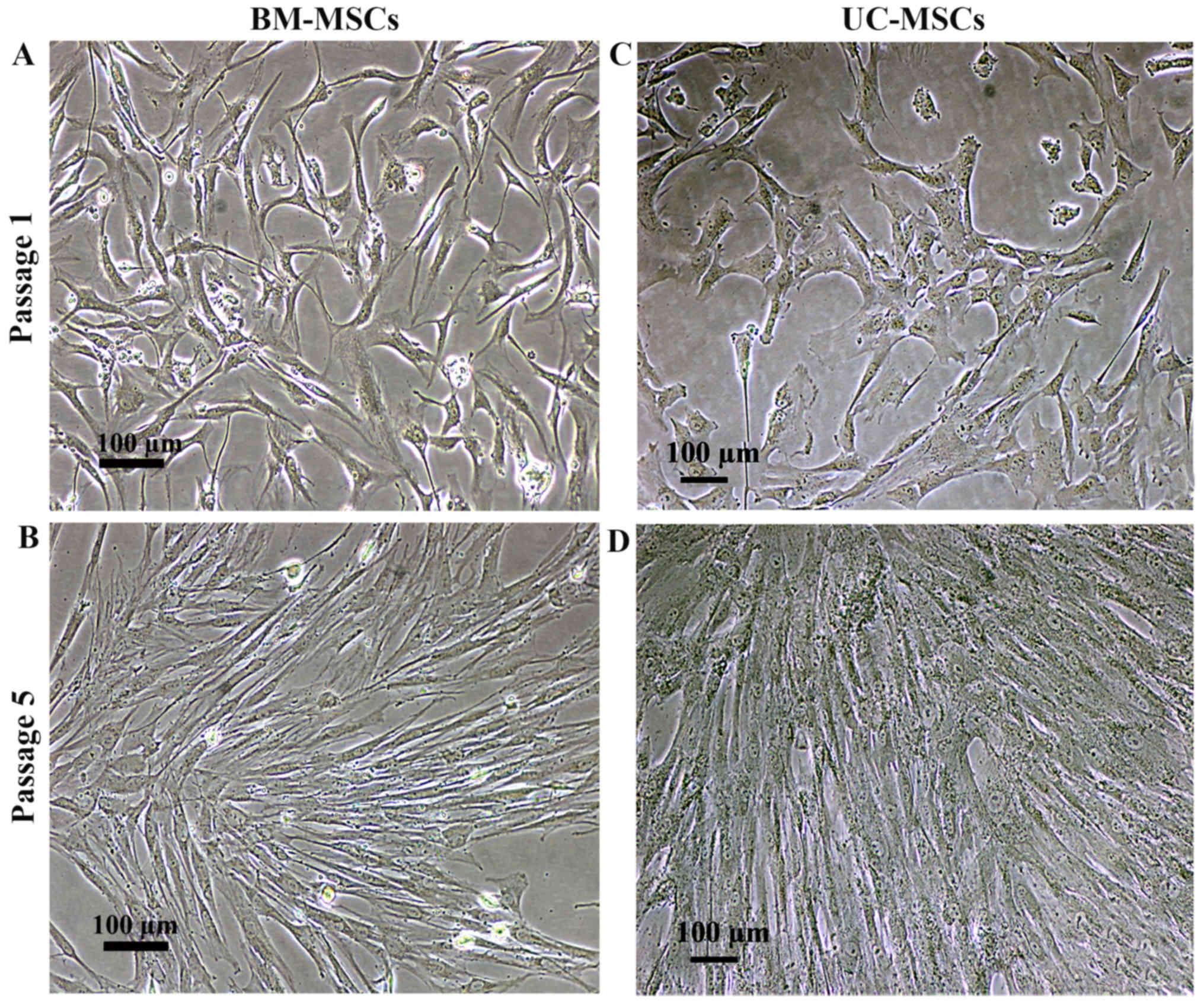
The cells isolated from bone marrow (BM-MSCs) and
umbilical cord (UC-MSCs) were cultured in DMEM supplemented with
10% FBS. Similar to the BM-MSCs, the cultured UC-MSCs exhibited a
spindle-shaped morphology with a high nuclear to cytoplasmic ratio
following culture for 7 days (Fig. 1A
and C). The culture medium was changed every 3 days and the
non-adherent cells were removed. Further passages were carried out
whenever the cell density reached 90% confluence, mostly on day 3
following sub-culture. There was no visible difference in the
morphology of the UC-MSCs and BM-MSCs (Fig. 1B and D).
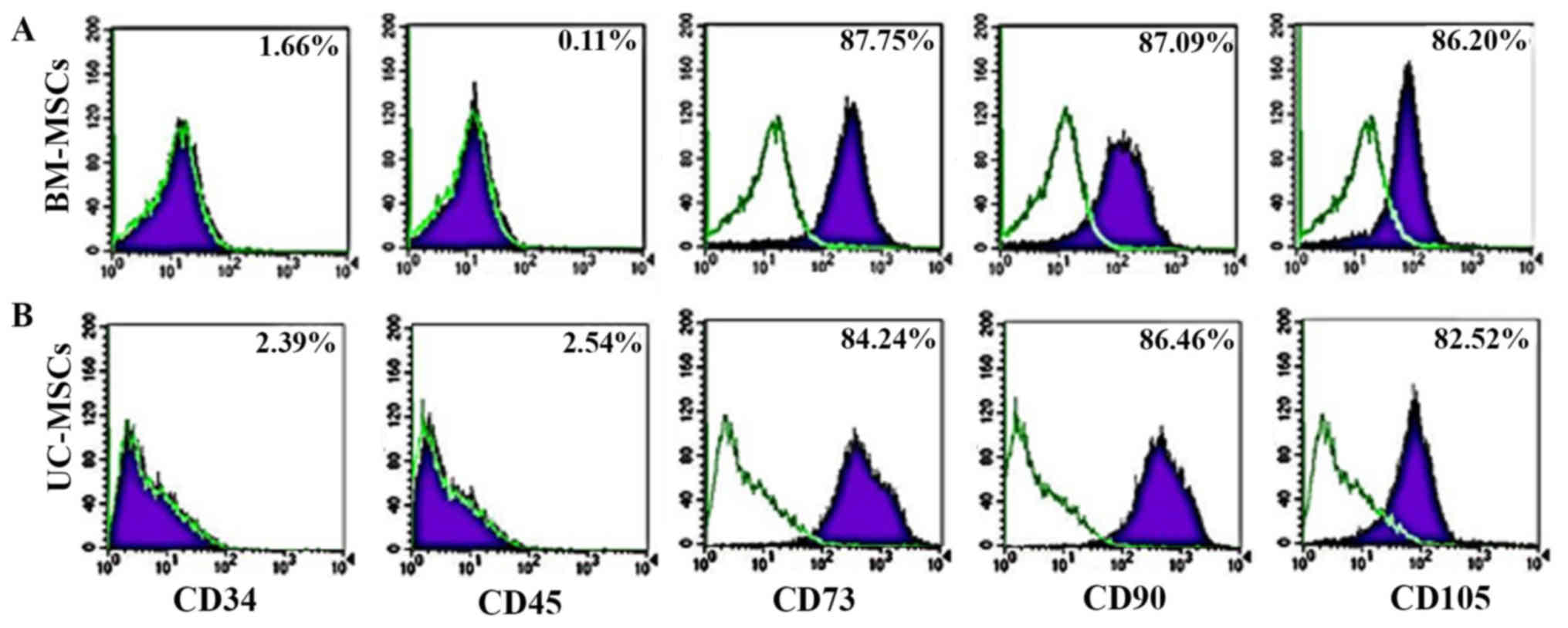
For immunophenotyping, both the UC-MSCs and BM-MSCs
expressed typical surface markers associated with MSCs, including
CD73, CD90 and CD105, and did not express the hematopoietic cell
markers, CD34 and CD45. There was no statistically significant
difference in the expression of MSC surface markers between the
UC-MSCs and BM-MSCs (Fig. 2).
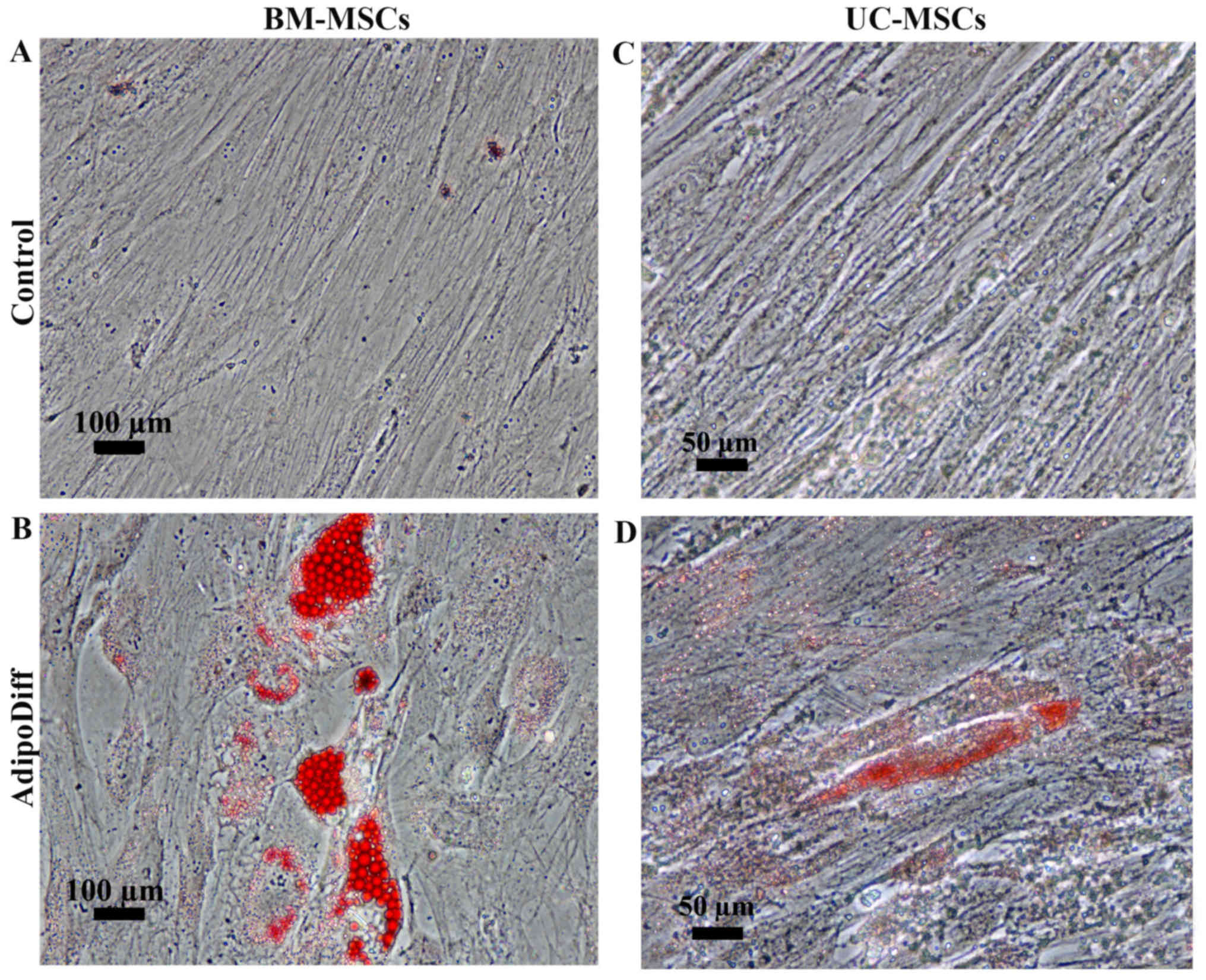
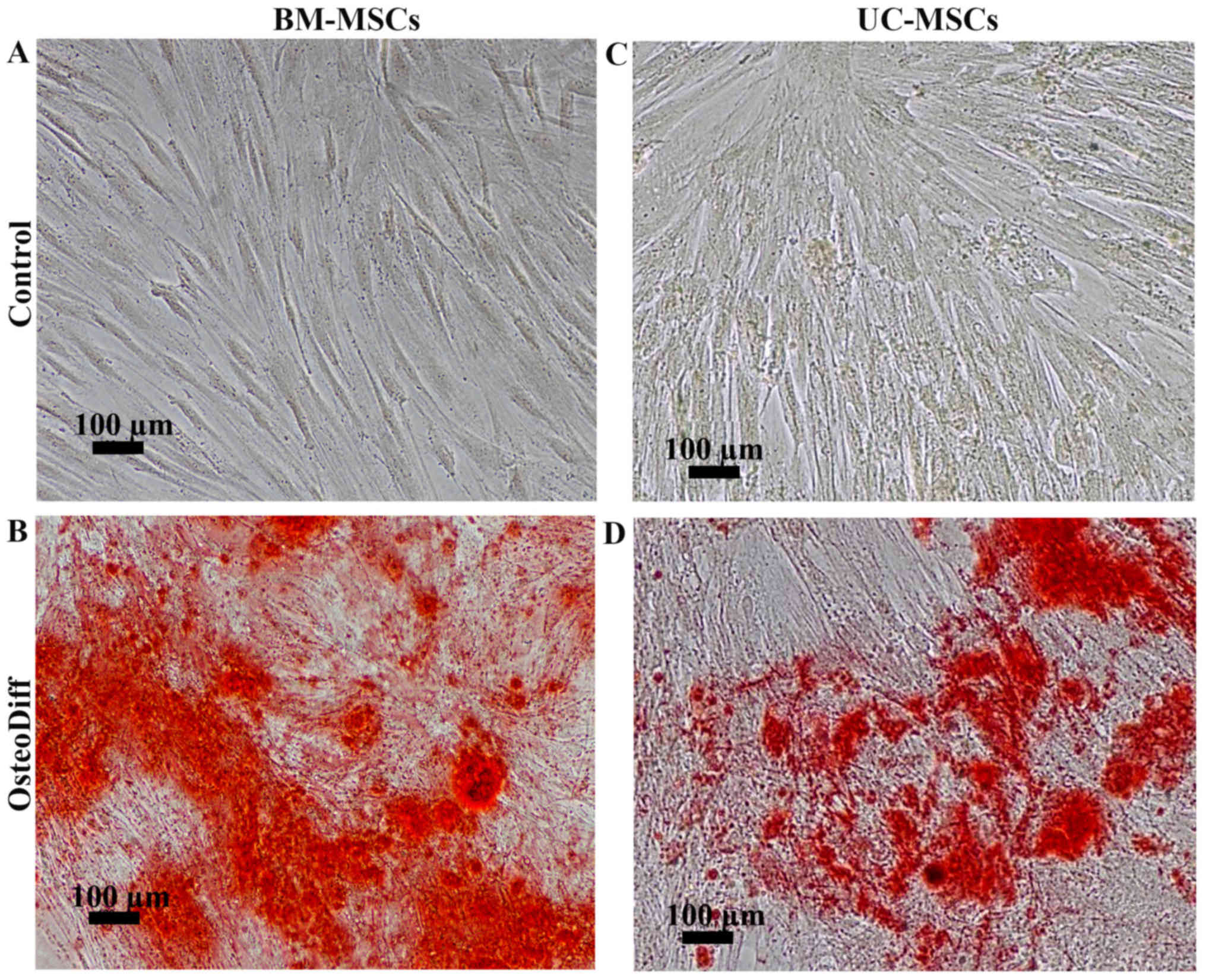
To examine the differentiation potential, the
UC-MSCs and BM-MSCs were induced to differentiate into adipocytes
or osteoblasts by culturing the cells in adipogenic or osteogenic
differentiation media. After 3 weeks of adipogenic induction, the
BM-MSCs became large cells which contained numerous lipid droplets
in their cytoplasm. In contrast to the BM-MSCs, the differentiated
UC-MSCs exhibited small amounts of lipid droplets. However, these
lipid droplets were positive for Oil Red O staining (Fig. 3B and D). The control cultures did
not have any lipid droplets in their cytoplasm and were negative
for Oil Red O staining (Fig. 3A and
C). For osteogenic differentiation, the mineralized matrix was
detected in the BM-MSCs and UC-MSCs following induction for 21 and
28 days (Fig. 4B and D). The
control cultures were negative for Alizarin Red staining (Fig. 4A and C). Of note, the MSCs from
both sources had the capacity to differentiate into osteoblasts and
adipocytes; however, the UC-MSCs required a longer induction period
than the BM-MSCs.
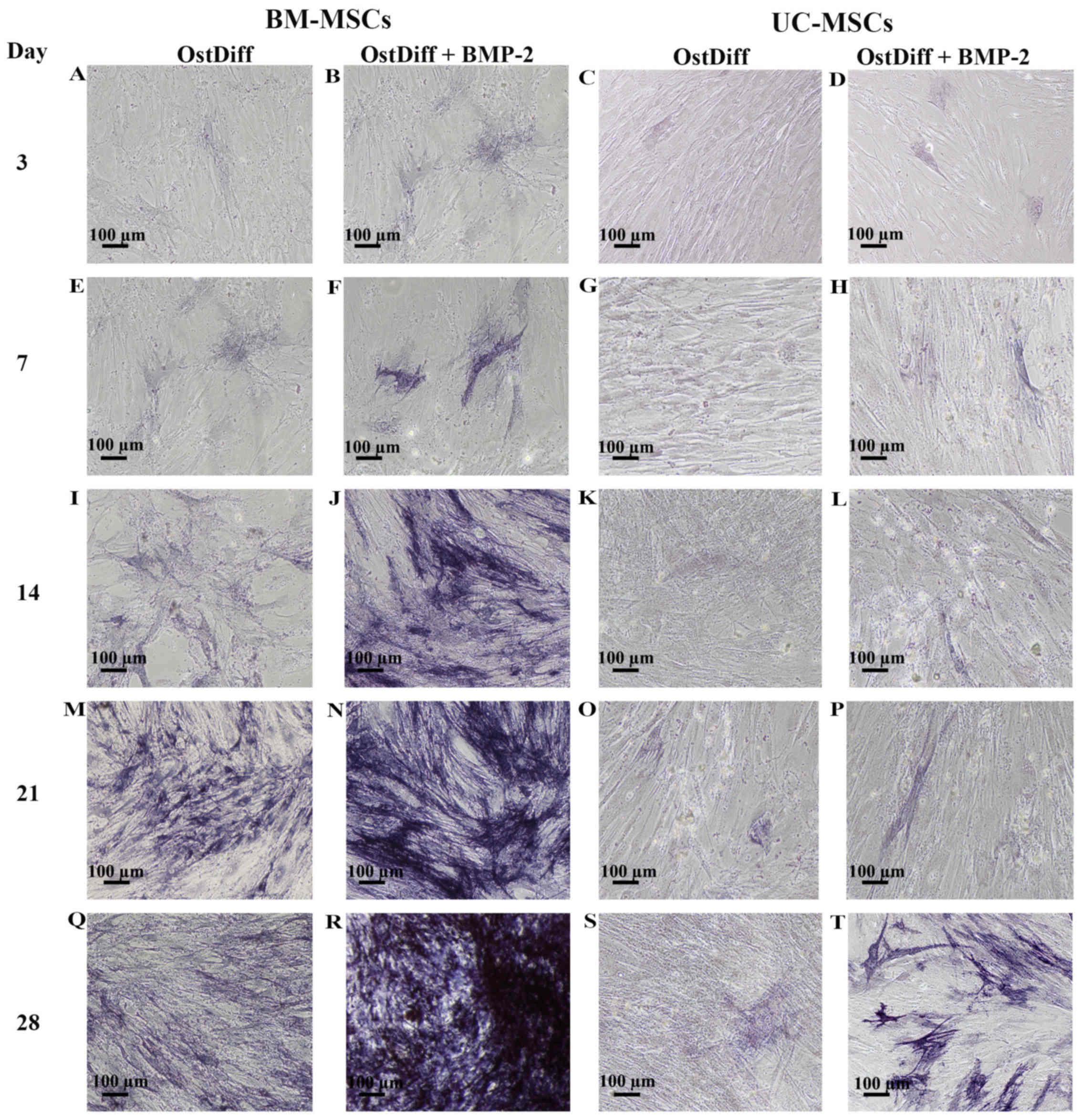
Expression of ALP following treatment
with BMP-2
To determine the effects of the BMP-2 on the
osteogenic differentiation of UC-MSCs in comparison to that of
BM-MSCs, the cells were cultured in three different media, complete
medium, osteogenic differentiation medium and osteogenic
differentiation medium in the presence of BMP-2 for 3, 7, 14, 21
and 28 days. Following induction for 3 days, few ALP-positive cells
were observed in the BM-MSCs cultured with osteogenic
differentiation medium in the absence of BMP-2 (Fig. 5A). Of note, treatment with BMP-2
increased the expression of ALP (Fig.
5B). In addition, the UC-MSCs treated with BMP-2 for 3 days
exhibited a higher ALP expression than those in the untreated group
(Fig. 5C and D). Following
sequential culture, the expression of ALP increased over time in
both the BM-MSCs and UC-MSCs cultured in osteogenic differentiation
medium with or without BMP-2. Remarkably, a moderate number of
ALP-positive cells was observed in the BM-MSCs treated with BMP-2
for 14 days (Fig. 5J). The
intensity of ALP staining was significantly increased in the
BM-MSCs treated with BMP-2 for 21 and 28 days as compared to the
BM-MSCs cultured in osteogenic differentiation medium without BMP-2
(Fig. 5M, N, Q and R). In the
UC-MSCs, treatment with BMP-2 increased the expression of ALP
(Fig. 5H, L, P and T) as compared
to the UC-MSCs cultured in osteogenic differentiation medium
without BMP-2 (Fig. 5G, K, O and
S). However, the intensity of ALP staining was significantly
less than that of the BM-MSCs at each time point examined (Fig. 5).
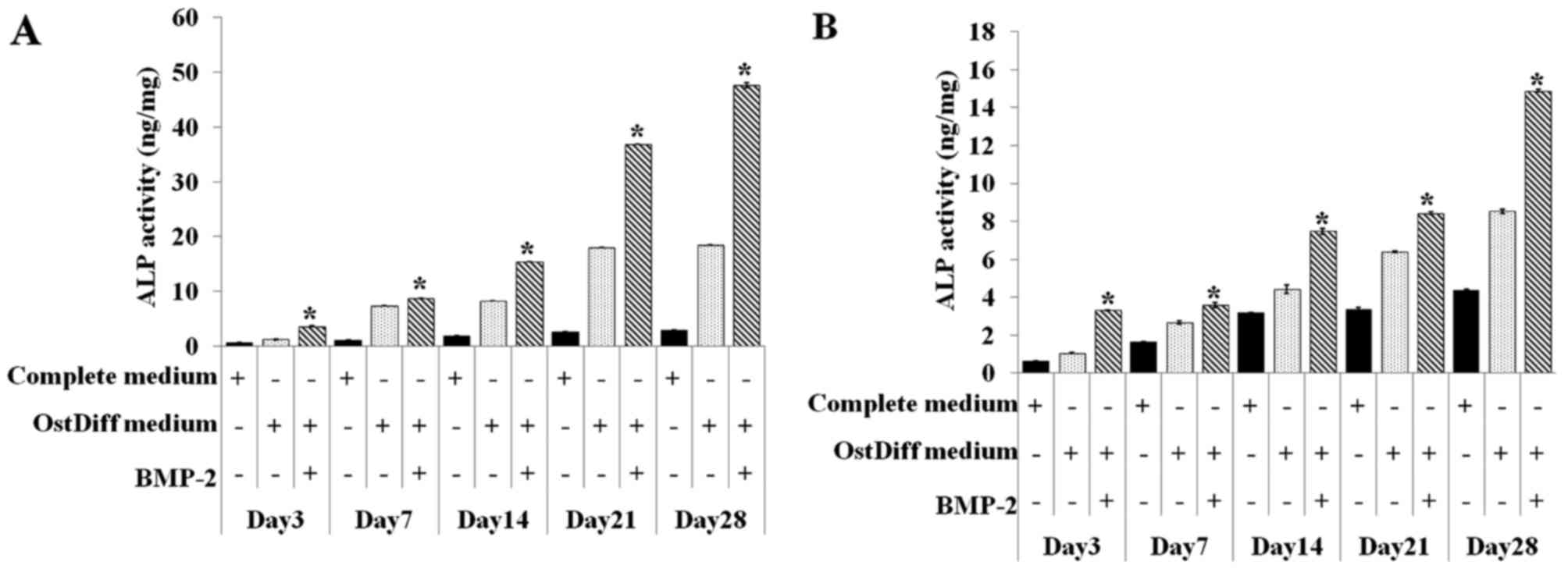
The activity of intracellular ALP in both the
BM-MSCs and UC-MSCs was also quantitative assessed on day 3, 7, 14,
21 and 28 in the three groups, namely the negative control (MSCs
cultured in complete medium), untreated group (MSCs cultured in
osteogenic differentiation medium), BMP-2 treated group (MSCs
cultured in osteogenic differentiation medium supplemented with
BMP-2). Of the three ways of treating MSCs, BMP-2 treatment of both
the BM-MSCs and UC-MSCs was shown to be clearly superior compared
to the other two treatments in regards to increasing ALP activity
(Fig. 6). In addition, the
activity of ALP in the BMP-2-treated groups significantly increased
with time. Similar to ALP staining, although ALP activity increased
over time in the BMP-2-treated groups, ALP activity in the UC-MSCs
was significantly less than that of the BM-MSCs at each time point
examined.
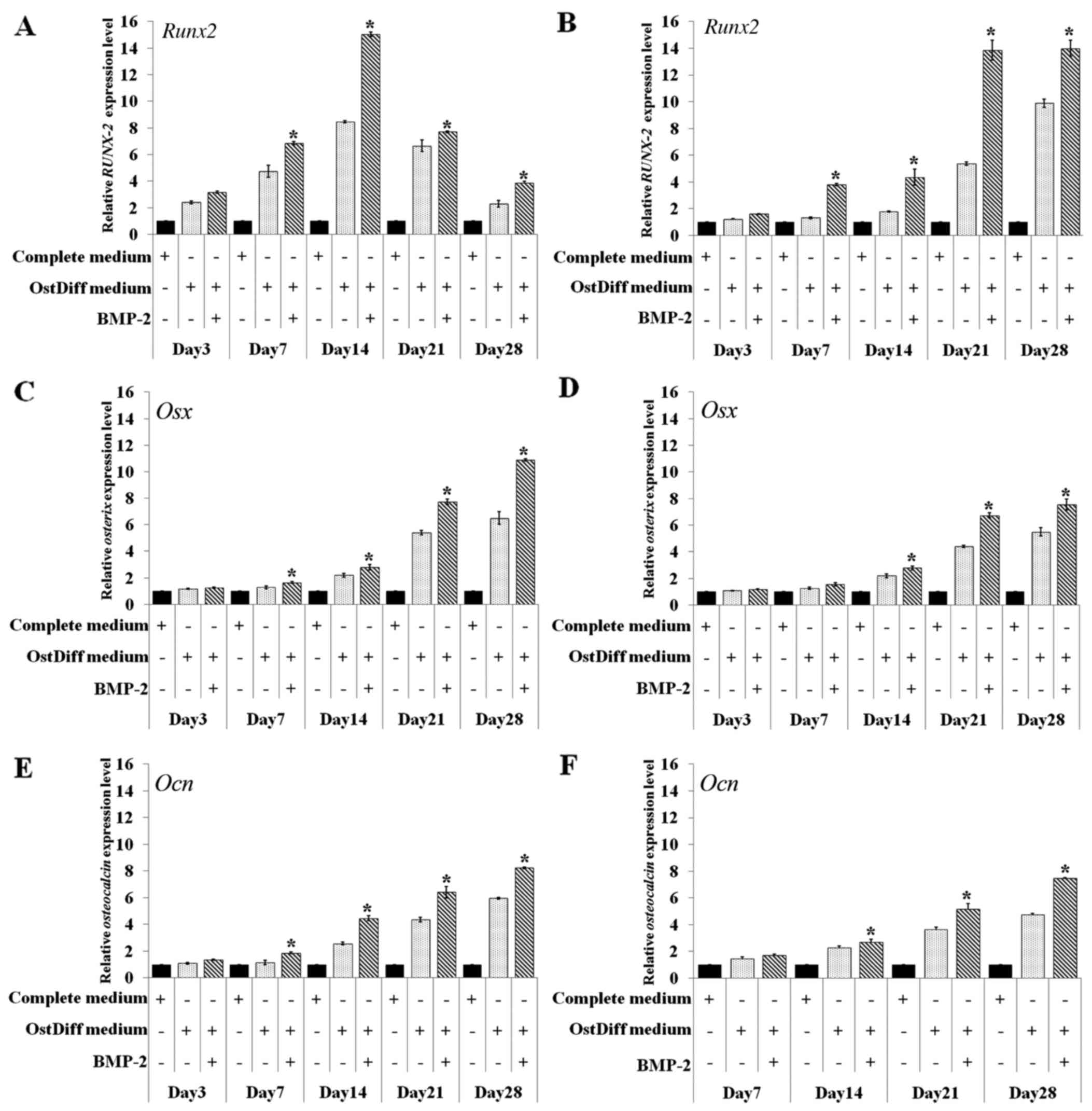
Effect of BMP-2 on the expression levels
of osteogenic lineage genes
In this study, the effect of BMP-2 on the osteogenic
differentiation of BM-MSCs and UC-MSCs was further investigated
through gene expression analysis of Runt-related transcription
factor 2 (Runx2), osterix (Osx) and osteocalcin
(Ocn) following 3, 7, 14, 21 and 28 days of culture. The
results revealed that in the BM-MSCs, BMP-2 significantly
upregulated the expression of the osteogenic lineage genes,
Runx2, Osx and Ocn on days 7, 14, 21 and 28
following osteogenic induction, while there were no significant
differences in the expression levels of these osteogenic lineage
genes during the earlier time points (day 3; Fig. 7A, C and E). The expression of
Runx2 increased over time from day 3 to 14 in the BM-MSC
cultures. The peak in Runx2 mRNA expression was observed on
day 14 in the BM-MSCs cultured in osteogenic differentiation medium
with or without BMP-2. Nevertheless, the BM-MSCs cultured in
osteogenic differentiation with BMP-2 exhibited a significantly
higher expression of Runx2 than those cultured in osteogenic
differentiation medium without BMP-2 (Fig. 7A).
In contrast to the BM-MSCs, Runx2 mRNA
expression increased over time from days 3 to 28 in the UC-MSCs
cultured in osteogenic differentiation medium with or without
BMP-2. Of note, the UC-MSCs treated with BMP-2 exhibited a
significantly higher expression of Runx2 than those in the
untreated group (Fig. 7B). The
effect of BMP-2 on the expression levels of other osteogenic
lineage genes in the cultured UC-MSCs also differed from that of
the BM-MSCs. The mRNA expression of Osx increased over time
from day 3 to 28 in the BM-MSCs and UC-MSCs cultured in osteogenic
differentiation with or without BMP-2 (Fig. 7C and D). However, BMP-2
significantly upregulated the gene expression in the cultured
UC-MSCs on days 14, 21 and 28 of culture (Fig. 7D), while the effect of BMP-2 in
upregulating gene expression was observed on days 7, 14, 21 and 28
in the cultured BM-MSCs (Fig.
7C). Similar to Osx, a minimum Ocn mRNA
expression was detected in the BM-MSCs on day 3 and significantly
increased Ocn levels were observed in the BM-MSCs treated
with BMP-2 compared with the untreated controls on days 7, 14, 21
and 28 (Fig. 7E). The mRNA
expression of Ocn was increased to the same extent with time
in the UC-MSCs in both the BMP-2-treated and untreated groups
(Fig. 7F). However, the increased
gene expression of Ocn in the UC-MSCs was significantly
observed in the BMP-2-treated group on days 14, 21 and 28.
Discussion
MSCs are adherent marrow stromal cells which have a
self-renewal ability and the potential to differentiate into
osteoblasts, chondrocytes, myoblasts and adipocytes (19). As multipotent progenitors, MSCs
have been regarded as ideal seed cells for scientific research and
bone tissue engineering (2,20).
However, the promotion of MSC differentiation into osteoblasts by
osteogenic induction factors is one of the most crucial issues in
bone tissue engineering (2,21).
BMP-2 is one of the most potent BMPs for promoting the osteogenic
differentiation of BM-MSCs both in vitro and in vivo
(22,23). A previous study reported that
BMP-2-producing cells, via adenoviral gene transfer, produced
sufficient protein to heal segmental bone defects in a rat model
(24). In addition, the
acceleration of bone regeneration using rat BM-MSCs transduced with
BMP-2 was higher than that of using MSCs alone (25). Several other animal studies
performed with the implantation of autologous BM-MSCs using
different scaffolds have resulted in bone regeneration (9,26).
Although BM-MSCs are dominant seed cell sources for bone
engineering, the limited cell number and the invasive procedure for
harvesting cell restrict their use in clinical fields. Over the
past few years, UC-MSCs are considered as promising alternative
source for MSCs used in the research and application of stem cell
therapy as they are easy to be isolated and expanded in
vitro and can be attained by a less invasive method without
harming the mother or infant (15,27). Although the UC-MSCs share similar
characteristics to the BM-MSCs, MSCs from different sources differ
in their differentiation potential and gene expression profile
(28). There is evidence
suggested that UC-MSCS require a longer period of time for
osteogenic differentiation compared to BM-MSCs (15). Therefore, it is still a challenge
to enhance the osteogenic differentiation potential of UC-MSCs for
clinical application. Although studies have demonstrated the
benefits of BMP-2 in bone tissue regeneration (29,30), the BMP-2-induced osteogenic
differentiation of MSCs derived from the umbilical cord has not
been fully examined.
In this study, MSCs were characterized according to
the criteria of the International Society for Cellular Therapy
(31). The results revealed that
the plastic-adherent MSCs isolated from both bone marrow and
umbilical cord displayed a fibroblast-like morphology. They were
typically negative for CD34, CD45 and were shown to be positive for
CD73, CD90 and CD105. The effect of BMP-2 on osteogenic
differentiation was investigated to clarify the role of BMP-2 in
the function of MSC differentiation. The expression of ALP was
monitored using ALP staining and ALP activity assay. ALP is
produced by osteoblasts and is hypothesized to be involved in the
degradation of inorganic pyrophosphates to provide sufficient local
phosphate or inorganic pyrophosphate for the occurrence of
mineralization (32). Therefore,
ALP activity is commonly used as a marker of osteogenesis to
reflect the degree of osteogenic differentiation. Our results
revealed that BMP-2 treatment enhanced the efficiency of osteogenic
differentiation of both human BM-MSCs and UC-MSCs as evidenced by
the increasing level of ALP staining and ALP activity. Accordingly,
the expression levels of osteogenic marker genes were also
increased by BMP-2 treatment in both human BM-MSCs and UC-MSCs.
Although the molecular mechanisms underlying
BMP-2-mediated osteogenesis remain to be fully understood, various
studies have demonstrated that BMP-2 plays a critical role in the
osteogenic differentiation of MSCs (2,12,23). In addition to the direct
application of recombinant BMP-2 proteins, it has been confirmed
that the adenoviral vector-mediated gene transfer of BMP-2 has the
ability to induce bone formation both in vitro and in
vivo (13). A previous study
using the mouse myoblast cell line, C2C12, demonstrated that BMP-2
inhibited myogenic differentiation, and instead diverted their
differentiation pathway into that of osteoblasts (33). Furthermore, BMP-2 has also been
shown to induce Runx2 expression in mesenchymal progenitors
in a Smad-dependent manner and to regulate the expression of target
genes which are involved in osteoblast differentiation (34).
In this study, we demonstrated that osteogenic
signaling molecules, including Runx2, Osx and
Ocn were upregulated in the BM-MSCs following treatment with
BMP-2. Of note, the peak expression of Runx2 in BM-MSCs
cultured with osteogenic differentiation medium in the presence or
absence of BMP-2 was observed on day 14. The addition of BMP-2
enhanced the expression of Runx2. In contrast to the
BM-MSCs, the highest expression of Runx2 in the UC-MSCS
cultured with osteogenic differentiation medium in the presence or
absence of BMP-2 was observed on day 28 and BMP-2 enhanced the
expression of Runx2 in the UC-MSCs similar to the BM-MSCs.
Runx2 is a pivotal osteogenic transcription factor. It may
be one of the earliest master transcription factors that directs
the differentiation of MSCs into osteoblasts (35). A previous study reported that
Runx2 is expressed in
Osx−/− mice, while Osx
is not expressed in Runx2−/− mice (36). Based on this finding, Runx2
has been proposed to be an upstream regulator of Osx
expression (37). Runx2
and Osx may control osteogenesis in a manner analogous to
the master transcription factors. A previous study suggested that
Runx2 plays a role in the commitment step for the common
progenitor cells for osteoblasts and chondrocytes, whereas
Osx plays a role in the final differentiation step in
osteogenesis (36). This notion
is further supported by evidence that the expression of Ocn,
the final differentiation marker of osteogenesis, is induced by
Osx overexpression but not by Runx2 overexpression
(38). These data support the
findings of this study. The expression of Osx at each time
point overlapped with the expression of Ocn. Of note, BMP-2
treatment significantly enhanced the expression of Osx and
Ocn in both the BM-MSCs and UC-MSCs. Although BMP-2 enhanced
the osteogenic differentiation capability of UC-MSCs, the effect
was less pronounced compared to the BM-MSCs. This may be due to the
endogenous difference in the osteogenic differentiation capacity
between BM-MSCs and UC-MSCs, as shown by the finding that the
endogenous ALP activity of the UC-MSCs was much lower than that of
the BM-MSCs cultured under the same conditions. It may be possible
that BMP-2 is not the only initial inducer of Runx2
expression during the osteogenic differentiation of UC-MSCs. A
previous study reported that some developmentally important
ligands, such as transforming growth factor (TGF)-β1, fibroblast
growth factors (FGFs) and BMP-9, can stimulate Runx2
expression and enhance ALP expression in bone development (39,40). In addition, the suitable
concentration of BMP-2 is still an issue as regards the enhancement
of the osteogenic capacity of UC-MSCs. Nevertheless, this study
demonstrated that BMP-2 treatment enhanced the osteogenic
differentiation capacity of both BM-MSCs and UC-MSCs, as evidenced
by an increase in ALP expression and osteogenic gene
expression.
In conclusion, the results obtained from this study
indicate that MSCs derived from the umbilical cord provide an
exciting and promising stem cell source for bone repair in skeletal
diseases. Our findings demonstrated that BMP-2 enhanced the
osteogenic differentiation capacity of both BM-MSCs and UC-MSCs.
These MSCs may potentially be used as a therapeutic agent in the
treatment of patients with bone defects. The use of MSCs derived
from bone marrow and umbilical cord may prove to be an innovative
treatment for many musculoskeletal diseases, including fracture
non-unions and a number of metabolic bone diseases; therefore,
their use in clinical applications warrants rigorous
assessment.
Acknowledgments
The authors would like to thank the staff at the
delivery room, Thammasat University Hospital, for facilitating
specimen collection and all the volunteers for kindly donating the
tissues for this study. This study was supported by grants from
Thailand Research Fund and Thammasat University (RSA5980042) and
Center of Excellence in Stem Cell Research, Thammasat
University.
References
|
1
|
Loeser RF: Age-related changes in the
musculoskeletal system and the development of osteoarthritis. Clin
Geriatr Med. 26:371–386. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petite H, Viateau V, Bensaïd W, Meunier A,
de Pollak C, Bourguignon M, Oudina K, Sedel L and Guillemin G:
Tissue-engineered bone regeneration. Nat Biotechnol. 18:959–963.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stock UA and Vacanti JP: Tissue
engineering: current state and prospects. Annu Rev Med. 52:443–451.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang XF, Song Y, Liu YS, Sun YC, Wang YG,
Wang Y and Lyu PJ: Osteogenic differentiation of three-dimensional
bioprinted constructs consisting of human adipose-derived stem
cells in vitro and in vivo. PLoS One. 11:e01572142016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yi H, Ur Rehman F, Zhao C, Liu B and He N:
Recent advances in nano scaffolds for bone repair. Bone Res.
4:160502016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Krampera M, Pizzolo G, Aprili G and
Franchini M: Mesenchymal stem cells for bone, cartilage, tendon and
skeletal muscle repair. Bone. 39:678–683. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Krampera M, Marconi S, Pasini A, Galiè M,
Rigotti G, Mosna F, Tinelli M, Lovato L, Anghileri E, Andreini A,
et al: Induction of neural-like differentiation in human
mesenchymal stem cells derived from bone marrow, fat, spleen and
thymus. Bone. 40:382–390. 2007. View Article : Google Scholar
|
|
8
|
Li X, Ling W, Pennisi A, Wang Y, Khan S,
Heidaran M, Pal A, Zhang X, He S, Zeitlin A, et al: Human
placenta-derived adherent cells prevent bone loss, stimulate bone
formation, and suppress growth of multiple myeloma in bone. Stem
Cells. 29:263–273. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Arinzeh TL, Peter SJ, Archambault MP, van
den Bos C, Gordon S, Kraus K, Smith A and Kadiyala S: Allogeneic
mesenchymal stem cells regenerate bone in a critical-sized canine
segmental defect. J Bone Joint Surg Am. 85-A:1927–1935. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hernigou P, Poignard A, Manicom O, Mathieu
G and Rouard H: The use of percutaneous autologous bone marrow
transplantation in nonunion and avascular necrosis of bone. J Bone
Joint Surg Br. 87:896–902. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tseng SS, Lee MA and Reddi AH: Nonunions
and the potential of stem cells in fracture-healing. J Bone Joint
Surg Am. 90(Suppl 1): 92–98. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
De Biase P and Capanna R: Clinical
applications of BMPs. Injury. 36(Suppl 3): S43–S46. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bais MV, Wigner N, Young M, Toholka R,
Graves DT, Morgan EF, Gerstenfeld LC and Einhorn TA: BMP2 is
essential for post natal osteogenesis but not for recruitment of
osteogenic stem cells. Bone. 45:254–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Osyczka AM and Leboy PS: Bone
morphogenetic protein regulation of early osteoblast genes in human
marrow stromal cells is mediated by extracellular signal-regulated
kinase and phosphatidylinositol 3-kinase signaling. Endocrinology.
146:3428–3437. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Manochantr S, U-pratya Y, Kheolamai P,
Rojphisan S, Chayosumrit M, Tantrawatpan C, Supokawej A and
Issaragrisil S: Immunosuppressive properties of mesenchymal stromal
cells derived from amnion, placenta, Wharton's jelly and umbilical
cord. Intern Med J. 43:430–439. 2013. View Article : Google Scholar
|
|
16
|
Cheng YH, Lin FH, Wang CY, Hsiao CY, Chen
HC, Kuo HY, Tsai TF and Chiou SH: Recovery of oxidative
stress-induced damage in Cisd2-deficient cardiomyocytes by
sustained release of ferulic acid from injectable hydrogel.
Biomaterials. 103:207–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Scarfi S: Use of bone morphogenetic
proteins in mesenchymal stem cell stimulation of cartilage and bone
repair. World J Stem Cells. 8:1–12. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dang PN, Dwivedi N, Phillips LM, Yu X,
Herberg S, Bowerman C, Solorio LD, Murphy WL and Alsberg E:
Controlled dual growth factor delivery from microparticles
incorporated within human bone marrow-derived mesenchymal stem cell
aggregates for enhanced bone tissue engineering via endochondral
ossification. Stem Cells Transl Med. 5:206–217. 2016. View Article : Google Scholar :
|
|
19
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Koç ON and Lazarus HM: Mesenchymal stem
cells: Heading into the clinic. Bone Marrow Transplant. 27:235–239.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Na K, Kim SW, Sun BK, Woo DG, Yang HN,
Chung HM and Park KH: Osteogenic differentiation of rabbit
mesenchymal stem cells in thermo-reversible hydrogel constructs
containing hydroxyapatite and bone morphogenic protein-2 (BMP-2).
Biomaterials. 28:2631–2637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Luu HH, Song WX, Luo X, Manning D, Luo J,
Deng ZL, Sharff KA, Montag AG, Haydon RC and He TC: Distinct roles
of bone morphogenetic proteins in osteogenic differentiation of
mesenchymal stem cells. J Orthop Res. 25:665–677. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Osyczka AM, Damek-Poprawa M, Wojtowicz A
and Akintoye SO: Age and skeletal sites affect BMP-2 responsiveness
of human bone marrow stromal cells. Connect Tissue Res. 50:270–277.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lieberman JR, Daluiski A, Stevenson S, Wu
L, McAllister P, Lee YP, Kabo JM, Finerman GA, Berk AJ and Witte
ON: The effect of regional gene therapy with bone morphogenetic
protein-2-producing bone-marrow cells on the repair of segmental
femoral defects in rats. J Bone Joint Surg Am. 81:905–917. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin Z, Wang JS, Lin L, Zhang J, Liu Y,
Shuai M and Li Q: Effects of BMP2 and VEGF165 on the osteogenic
differentiation of rat bone marrow-derived mesenchymal stem cells.
Exp Ther Med. 7:625–629. 2014.PubMed/NCBI
|
|
26
|
Viateau V1, Guillemin G, Bousson V, Oudina
K, Hannouche D, Sedel L, Logeart-Avramoglou D and Petite H:
Long-bone critical-size defects treated with tissue-engineered
grafts: A study on sheep. J Orthop Res. 25:741–749. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu LL, Liu YJ, Yang SG, Zhao QJ, Wang X,
Gong W, Han ZB, Xu ZS, Lu YX, Liu D, et al: Isolation and
characterization of human umbilical cord mesenchymal stem cells
with hematopoiesis-supportive function and other potentials.
Haematologica. 91:1017–1026. 2006.PubMed/NCBI
|
|
28
|
Wagner W, Wein F, Seckinger A, Frankhauser
M, Wirkner U, Krause U, Blake J, Schwager C, Eckstein V, Ansorge W,
et al: Comparative characteristics of mesenchymal stem cells from
human bone marrow, adipose tissue, and umbilical cord blood. Exp
Hematol. 33:1402–1416. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kratchmarova I, Blagoev B, Haack-Sorensen
M, Kassem M and Mann M: Mechanism of divergent growth factor
effects in mesenchymal stem cell differentiation. Science.
308:1472–1477. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hsiao HY, Yang SR, Brey EM, Chu IM and
Cheng MH: Hydrogel delivery of mesenchymal stem cell-expressing
bone morphogenetic protein-2 enhances bone defect repair. Plast
Reconstr Surg Glob Open. 4:e8382016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop Dj and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The International Society for Cellular
Therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mikami Y, Tsuda H, Akiyama Y, Honda M,
Shimizu N, Suzuki N and Komiyama K: Alkaline phosphatase determines
polyphosphate-induced mineralization in a cell-type independent
manner. J Bone Miner Metab. 34:627–637. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Katagiri T, Yamaguchi A, Komaki M, Abe E,
Takahashi N, Ikeda T, Rosen V, Wozney JM, Fujisawa-Sehara A and
Suda T: Bone morphogenetic protein-2 converts the differentiation
pathway of C2C12 myoblasts into the osteoblast lineage. J Cell
Biol. 127:1755–1766. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nishimura R, Hata K, Ikeda F, Ichida F,
Shimoyama A, Matsubara T, Wada M, Amano K and Yoneda T: Signal
transduction and transcriptional regulation during mesenchymal cell
differentiation. J Bone Miner Metab. 26:203–212. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ryoo HM, Lee MH and Kim YJ: Critical
molecular switches involved in BMP-2-induced osteogenic
differentiation of mesenchymal cells. Gene. 366:51–57. 2006.
View Article : Google Scholar
|
|
36
|
Nakashima K, Zhou X, Kunkel G, Zhang Z,
Deng JM, Behringer RR and de Crombrugghe B: The novel zinc
finger-containing transcription factor osterix is required for
osteoblast differentiation and bone formation. Cell. 108:17–29.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tsao YT, Huang YJ, Wu HH, Liu YA, Liu YS
and Lee OK: Osteocalcin mediates biomineralization during
osteogenic maturation in human mesenchymal stromal cells. Int J Mol
Sci. 18:1592017. View Article : Google Scholar :
|
|
38
|
Lee KS, Kim HJ, Li QL, Chi XZ, Ueta C,
Komori T, Wozney JM, Kim EG, Choi JY, Ryoo HM and Bae SC: Runx2 is
a common target of transforming growth factor beta1 and bone
morphogenetic protein 2, and cooperation between Runx2 and Smad5
induces osteoblast-specific gene expression in the pluripotent
mesenchymal precursor cell line C2C12. Mol Cell Biol. 20:8783–8792.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim HJ, Kim JH, Bae SC, Choi JY, Kim HJ
and Ryoo HM: The protein kinase C pathway plays a central role in
the fibroblast growth factor-stimulated expression and
transactivation activity of Runx2. J Biol Chem. 278:319–326. 2003.
View Article : Google Scholar
|
|
40
|
Kim YJ, Lee MH, Wozney JM, Cho JY and Ryoo
HM: Bone morphogenetic protein-2-induced alkaline phosphatase
expression is stimulated by Dlx5 and repressed by Msx2. J Biol
Chem. 279:50773–50780. 2004. View Article : Google Scholar : PubMed/NCBI
|