Introduction
Pancreatic carcinoma is a neoplasm formed by
abnormal proliferation of pancreatic cells due to the dysregulation
of cellular growth under the effect of multiple tumorigenic
factors. Due to its biological complexity and serious threat to
patients, it is necessary to develop new therapeutic strategies for
pancreatic carcinoma since current approaches have limited
efficacy. For example, gemcitabine is associated with serious side
effects and resistance is observed in various cases (1). Among tumorigenic factors, a
deficiency in cellular apoptosis, the programmed death of cells,
plays a critical role in the onset and development of tumors
(2). Caspase activation is a key
step in apoptosis and can be activated by both intrinsic and
extrinsic pathways to induce a catalytic reaction and mediate
cellular apoptosis (3). It has
been proposed that an increase in the apoptotic threshold through
alteration of molecules contributes to the therapeutic resistance
of pancreatic carcinoma including apoptosis inducers or antitumor
medications (4). Therefore,
enhancing the sensitivity of tumor cells to apoptosis inducers is a
potential strategy for the development of novel therapeutic
strategies.
Tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL), also termed as Apo2 ligand (Apo2L), is a member of
the tumor necrosis factor (TNF) superfamily and the immune
regulator of congenital and acquired immunity. TRAIL can initiate
apoptotic signaling by binding to the death receptor (DR) to induce
apoptosis targeting multiple tumor cells but without obvious
killing of normal cells (5).
However, treating different types of tumors simply dependent on
TRAIL/Apo2L has limited efficacy. Studies have indicated that
normal cells and more than half of passaged tumor cells (even
>60%) demonstrate tolerance to TRAIL (4,6).
The reason for the tolerance is due to the existence of a
deficiency and mutations in the apoptotic signaling pathways of
tumor cells, which include pro-apoptotic factors or anti-apoptotic
factors to increase the apoptotic threshold of drug-resistant tumor
cells to escape apoptotic scavenging. Increasing the sensitivity of
DR to TRAIL in tumor cells and the activity of pro-apoptotic
factors, or removing the inhibition of anti-apoptotic factors can
promote the efficacy of TRAIL to induce the apoptosis of tumor
cells and reverse the tolerance of tumor cells to TRAIL.
Cell-penetrating peptides (CPPs) exhibit high
transportation efficacy, low toxicity and no permanent damage to
the cellular membrane, and therefore show potential value in
reprogramming and gene editing (7). As one member of the CPPs, the basic
amino acid polyarginine (pAr) carries cations under physiological
pH and can bind to the negative glycosaminoglycan or lipids on the
cellular membrane. Because of the small structure and higher
penetrating efficacy, pAr can transport molecules into the
cytoplasm to bind to downstream signals and to display its
biological effects. The features of CPPs demonstrate that the
targeting protein can be transported to the cytoplasm through CPPs
to decrease the clearance rate of drugs in the blood and to ensure
that the drugs directly react with the targeting signal in cells,
which simultaneously activate extracellular and intracellular
signaling to play a role in the effects of drugs. Therefore, pAr
has been used to design antitumor peptides (8,9).
In the present study, we modified the N-terminal of
TRAIL to R6 by mutating 4 loci at domain 114-281aa which is
enriched in arginine to form a CPP-like amino acid sequence [TRAIL
mutant R6 (MuR6-TR)] and investigated the in vivo and
in vitro antitumor effects of MuR6-TR in pancreatic
carcinoma. This study provides evidence for the modification of
TRAIL in order to enhance the sensitivity of pancreatic carcinoma
cells to apoptosis inducers.
Materials and methods
Design and synthesis of the primers
According to the literature (10) and GeneBank (http://www.ncbi.nlm.nih.gov/nuccore/), the soluble
TRAIL sequence 114-281 at the N-terminal was selected. In reference
to the prefered adjusting coding sequence of Escherichia
coli (E. coli) synonym codon, the TRAIL sequence was
inserted with the initiation codon ATG (M) and termination codon
TAA to harvest E. coli preference codon with 513 bp. The
amino acid sequence VRERGP located at 114-119 was mutated into
RRRRRR, i.e., the N-terminal of natural TRAIL was mutated into R6
with 4 mutation loci to obtain the TRAIL mutant R6
(MuR6-TR). The amino acid sequence of MuR6-TR at
114-281 was: MRRRRRRQRVAAHITGTRGRSNTLSSPNSKNEKALGR
KINSWESSRSGHSFLSNLHLRNGELVIHEKGFYYIYSQ
TYFRFQEEIKENTKNDKQMVQYIYKYTSYPDPILLMK
SARNSCWSKDAEYGLYSIYQGGIFELKENDRIFVSVTN EHLIDMDHEASFFGAFLVG.
The existing TRAIL sequence was used as a template
for PCR to achieve local mutation with upstream NdeI and
downstream EcoRI as restriction enzyme cleavage sites. The
primers were upstream MuR6-TR-NdeI (48 bp), GGTCATATGCGT
CGTCGTCGTCGTCGTCAGCGTGTGGCTGCTCACATC and downstream TR-Eco-R
(41 bp) GTTGAATTCTTATTAACCAACAAGGAAAGCACCGAAGAAAG.
Amplification of the MuR6-TR segment with
PCR
The MuR6-TR was amplified with PCR using 50
µl of the total reaction system containing plasmid DNA
(Pmd19/TRAIL) 1 µl, 10× PCR buffer for KOD-Plus-Neo 5
µl, dNTPs (2 mM) 5 µl, 25 mM MgSO4 3
µl, KOD-Plus-Neo 1 µl,
MuR6-TR-NdeI/TR-Eco-R (10 pmol/µl) 1
µl, MuR6-TR-NdeI/TR-Eco-R (10 pmol/µl)
0.5 µl of each and RNase-free water 33 µl. The
reaction condition included an initial denaturation at 94°C for 2
min followed by 25 cycles of denaturation at 94°C for 15 sec and
annealing/extension at 68°C for 30 sec, with a final extension at
68°C for 5 min.
Transformation and identification of
MuR6-TR
The vector and target gene were digested with
NdeI and EcoRI, harvested with OMEGA recovery kit,
eluted with 30 µl deionized water, electrophoresed and
images were captured, respectively. The target segment and vector
were linked together and 10 µl of the linked product was
added to 100 µl Top10 competent cells for transformation.
Then the transformed competent cells were smeared on LB solid
medium containing ampicillin (Amp) at 37°C overnight. The bacterial
colonies were selected and digested with enzyme for identification.
These positive colonies were stored for sequencing.
Expression of MuR6-TR in bacterial
colony
The E. coli BL21(DE3) (in 1,000 µl)
treated with pET3a-MuR6-TR at 37°C overnight was added to 50 ml
LB-Amp+ medium and incubated on a shaking plate (250
rpm) at 37°C for 3 h and then the temperature was decreased to
24°C. Then, IPTG (0.1 M) was added at a 1% ratio for culture
induction overnight. Samples of 0.5 and 0.15 ml collected before
and after induction were centrifuged and the sediments after
removal of the supernatant were re-suspended with 50 µl
H2O, followed by addition of 50 µl 2× loading
buffer for electrophoresis. The remaining bacterial solution was
centrifuged at 12,000 rpm for 5 min to obtain the bacterial
collection which was re-suspended with 8 ml
Na2HPO4 (50 mM) and lysed with ultrasound.
The ultrasound lysis condition was: Φ6 probe, sonication with 200 W
pulses for 2 sec with an interval of 2 sec, and repetition for 10
min. The 1 ml bacterial lysis was centrifuged at 12,000 rpm for 10
min to harvest the supernatant and sediment. Then, the supernatant
and sediment re-suspended with 1 ml H2O (20 µl
for each) were added to 30 µl H2O and 50
µl 2× loading buffer for electrophoresis. The samples for
electrophoresis were heated in boiled water for 10 min and
centrifuged at 12,000 rpm for 10 min. Finally, 10 µl of the
supernatants was used for SDS-PAGE electrophoresis.
Purification of the targeting
protein
The protein was firstly purified with cation SP
Sepharose FF XK16 column (GE Healthcare, Piscataway, NJ, USA),
according to the manual, followed by elution with an anion XK26/20
column filled with Sephadex G-25 medium and anion exchanging buffer
(GE Healthcare). Then the protein was purified using a Q Sepharose
FF XK16 column (GE healthcare), according to the manufacturer's
manual. The penetrating and eluting components were collected,
respectively, and 50 µl 2× loading buffer was added in a 1:1
ratio for electrophoresis. The same volume (50 µl) of
original solution served as the control. The purified MuR6-TR and
TRAIL proteins were subjected to western blotting for
identification.
Activity of MuR6-TR protein in inhibiting
proliferation of tumor cells
TRAIL-insensitive pancreatic carcinoma cell lines
(BxPC-3 and PANC-1) were cultured in a 5% CO2 incubator
at 37°C with different media and density in a 96-well plate as
shown in Table I. The medium was
changed every 2–3 days, and the cells were passaged with medium
containing 0.25% trypsin 0.02% EDTA at a 1:1 ratio. Cells in a
logarithmic growth stage were used for experiments.
 | Table IConditions for cell culture. |
Table I
Conditions for cell culture.
| Cell line | Medium | Density |
|---|
| BxPC-3 | RPMI-1640 + 10% FBS
+ 1.0 mM sodium pyruvate |
4×103/well |
| PANC-1 | Low-glucose DMEM +
10% FBS |
5×103/well |
The MuR6-TR and TRAIL proteins were diluted
with sterile PBS buffer to a final concentration of 5 mg/ml and
sterilized with a filter. The cells were then treated with the
different proteins with initial concentrations of 1 µg/ml
and 200 µg/ml (the concentration of natural TRAIL protein
was adjusted according to preliminary data) followed by a 3-fold
dilution (total 10 dilution concentration). The experiment was
repeated triple times.
The inhibitory effects on the proliferation of cells
were measured with CCK-8 (cat. no. CK04-13; Dojindo), according to
the manufacturer's instructions, analyzed with the equation y = A2
+ (A1−A2)/[1 + (x/x0)p] using OriginPro 9.0 software and
fitted with growth/digmoidal-logistics for the inhibition rate
curve to calculate IC50. The in vitro therapeutic
effect of the drugs was defined as sensitive killing with
IC50 <10 µg/ml, dose-dependent cytotoxicity
and maximal inhibition ratio >80%.
Inhibition of the growth of the implanted
tumors in PANC-1- loaded nude mice by MuR6-TR
All animal procedures were approved by the Animal
Care and Scientific Committee of Sichuan University. Balb/c nude
female mice (SPF, 6–8 weeks, 18–22 g) were provided by Shanghai
Sippr-BK Laboratory Animal Ltd. and bred at 23±2°C, with a humidity
of 40–70% and a 12/12 light/dark cycle with free access to food and
water.
The PANC-1 cells were cultured in DMEM containing
10% FBS in a 5% CO2 incubator at 37°C. The cells at
logarithm growth stage were digested with 0.25% trypsin, rinsed
with PBS and re-suspended with serum-free medium to adjusted the
cell density to 2.5×107 cells/ml (1:1 Matrigel).
Under sterile conditions, each nude mouse was
subcutaneously implanted with a 0.2 ml cell suspension
(5×106 cells/mouse) in the right axillary. When tumors
grew to a size of 150–250 mm3, 32 mice with a healthy
appearance and with a similar tumor size (single, global tumor
without irregular shape or cluster tumors) were divided into 4
groups: i) saline; ii) gemcitabine; iii) natural TRAIL; and iv)
MuR6-TR (n=8 for each group). The mice in the different groups were
injected via tail vein with the different agents according to
Table II consecutively for 5
days at fixed times.
| Table IITreatments of mice in the different
groups. |
Table II
Treatments of mice in the different
groups.
| Group |
Concentration
(mg/ml) | Volume
(ml/kg) | Strategy |
|---|
| Vehicle
(saline) | – | 10 | i.v., q.d. × 5
days |
| Gemcitabine | 5 | 10 | i.v., q.o.d. × 3
times |
| TRAIL | 6 | 10 | i.v., q.d. × 5
days |
| MuR6-TR | 6 | 10 | q.d. × 5 days |
The growth of tumors was measured twice weekly with
the long (Y) and short (X) diameters to calculate the tumor growth
inhibition (TGI, %), and simultaneously the body weight of mice was
determined consecutively for 4 weeks. The TGI (%) was calculated
according to the formula:
TGI(%)=(1−tumor volumetreatment/tumor volumevehicle)×100%,
where volumetreatment is the tumor volume in the
treatment groups and tumor volumevehicle is the tumor
volume in the vehicle group.
After the last injection of the agents, the animals
were sacrificed with an over-dosage of CO2 and the
tumors were removed to measure the weight. If the animals appeared
moribund or the tumor size was >3,000 mm3 during the
experiment, the animal was sacrificed with an over-dosage of
CO2 to examine the pathological change in organs.
Statistical analysis
All data are expressed as mean ± SEM only when
denoted differently. Comparisons were performed with the Student's
t-test and P<0.05 was considered to indicate a statistically
significant difference. The mortality rate was expressed as a
percentage and compared with the Chi-square test.
Results
Amplification and transformation of the
MuR6-TR targeting gene
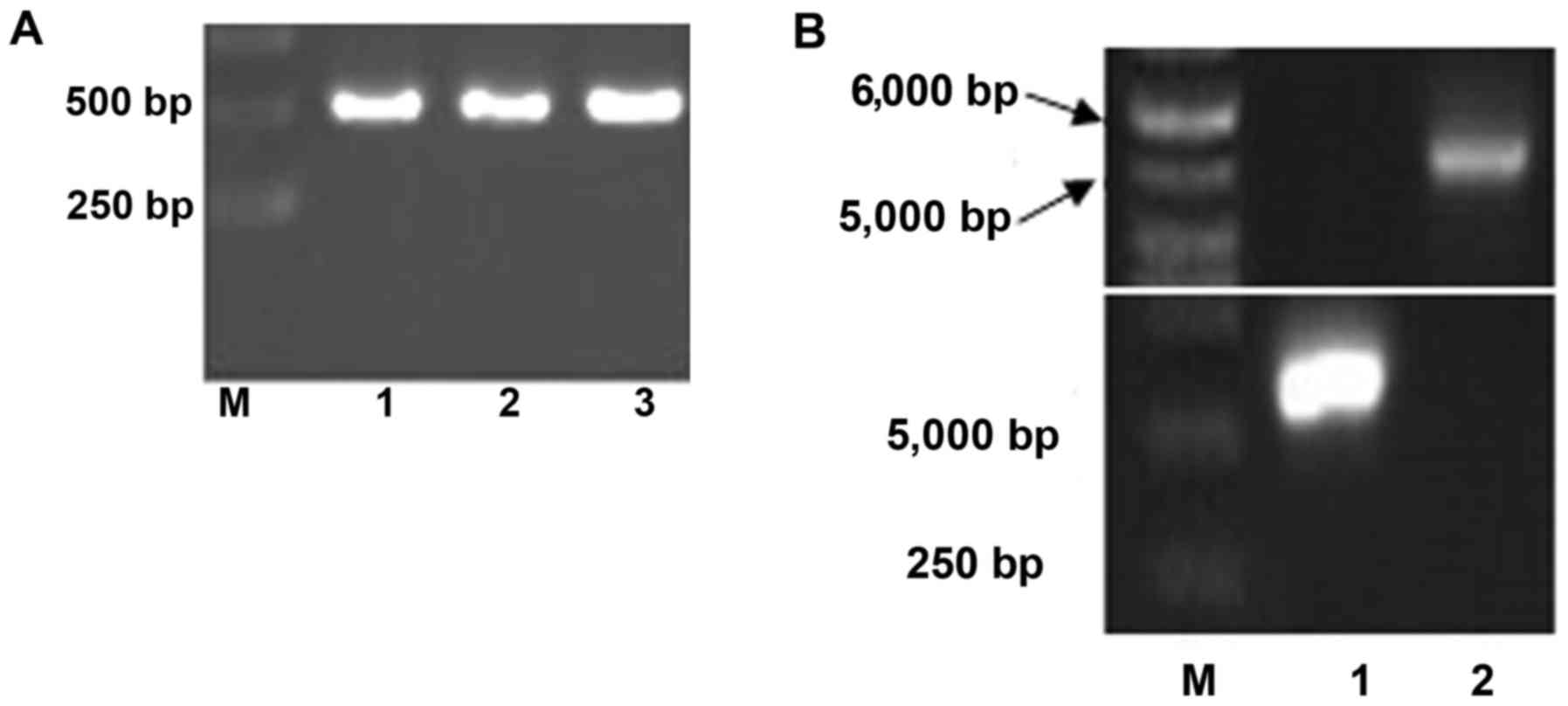
As shown in Fig.
1A, the resulting MuR6-TR segment by one-round PCR with
primers MuR6-TR-NdeI/TR-Eco-R was ~510 bp which was
in agreement with the size of the designed gene product (513
bp).
The segments from MuR6-TR and pET32a digested
by NdeI and EcoRI were 550 bp and 5.4 kb,
respectively, as shown in Fig.
1B. The sequence of the target gene MuR6-TR was
confirmed by Beijing Genomics Institute (Shenzhen, China) and was
in agreement with the designed sequence as shown in Fig. 1C.
After transformation by connection of
MuR6-TR/pET32a, the bacteria grew well with normal density. The
transformed plasmid carrying pET32a/MuR6-TR in the bacteria was
digested with XbaI and EcoRI, resulting in an ~5.4 kb
vector segment and ~550 bp target segment. As shown in Fig. 1D, the target segment of MuR6-TR
was positive in 8 of 10 samples.
Expression and purification of
pET32a/MuR6-TR in bacteria
The expression of pET32a/MuR6-TR in bacteria was
high. The supernatant of lysate contained 80% target protein and
the sediment contained 20% target protein (Fig. 2A).
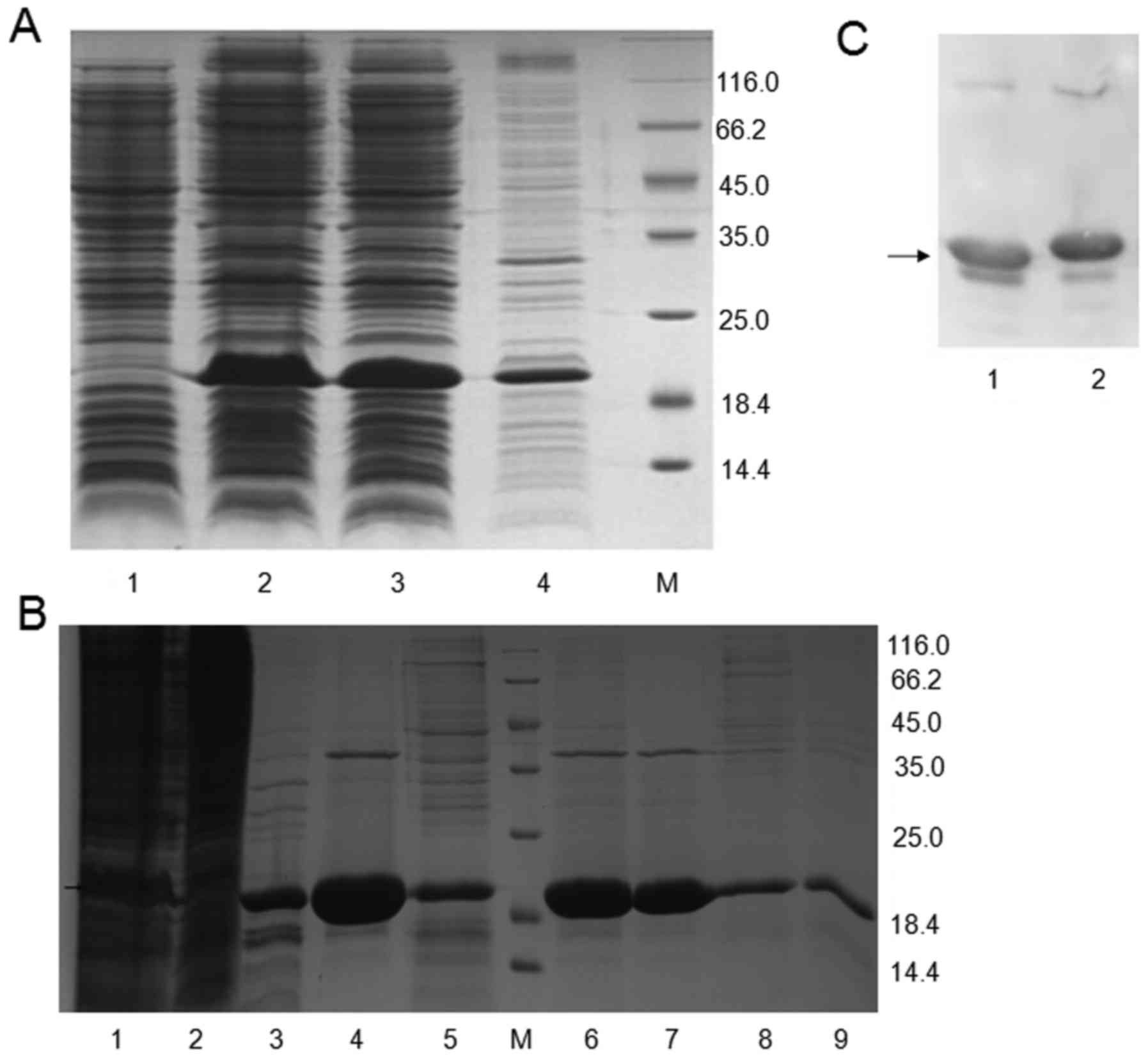 | Figure 2Identification of MuR6-TR
targeting protein. (A) Electrophoresis of pET32a/MuR6-TR protein
from transformed bacteria: lane 1, before induction; lane 2, after
induction; lane 3, supernatant; lane 4, sediment. (B) Proteins in
different eluting solution: lane 1, original MuR6 solution in SP
column; lane 2, penetrating solution in SP column; lane 3, 0.8 M
NaCl eluting solution in SP column; lane 4, 1.5 M NaCl eluting
solution in SP column; lane 5, NaOH eluting solution in SP column;
lane 6, original anion-exchange solution; lane 7, anion-exchange
penetrating solution; lane 8, anion-exchange eluting solution; lane
9, 0.5 M NaOH eluting solution; M, marker. (C) Expression of TRAIL
(lane 1) and MuR6-TR (lane 2) determined by western
blotting. TRAIL, tumor necrosis factor-related apoptosis-inducing
ligand; MuR6-TR, TRAIL mutant R6. |
The concentration of MuR6-TR protein in the
cation-exchange eluting solution, desalination eluting solution and
anion-exchange penetrating solution was measured (Table III). According to Table III, the recovery rate from
cation-exchange to desalination, from desalination to
anion-exchange and from cation-exchange to anion-exchange was
91.33, 87.68 and 80.08%, respectively, which was within the
acceptable range. The solution during exchange was clear and the
purified protein was subjected to electrophoresis as shown in
Fig. 2B. There was a visible
protein band in the cation-exchange eluting solution; in the final
anion-exchange penetrating solution, there was a clear protein band
and few artificial bands.
| Table IIIMeasurement of protein in the
different chromatography solutions. |
Table III
Measurement of protein in the
different chromatography solutions.
| Samples | Volume (ml) | Concentration
(mg/ml) |
|---|
| Cation-exchange
eluting solution | 26 | 7.7267 |
| Desalination
eluting solution | 44 | 4.1701 |
| Anion-exchange
penetrating solution | 69 | 2.3315 |
The expression of TRAIL and MuR6-TR proteins
was confirmed by western blotting (Fig. 2C).
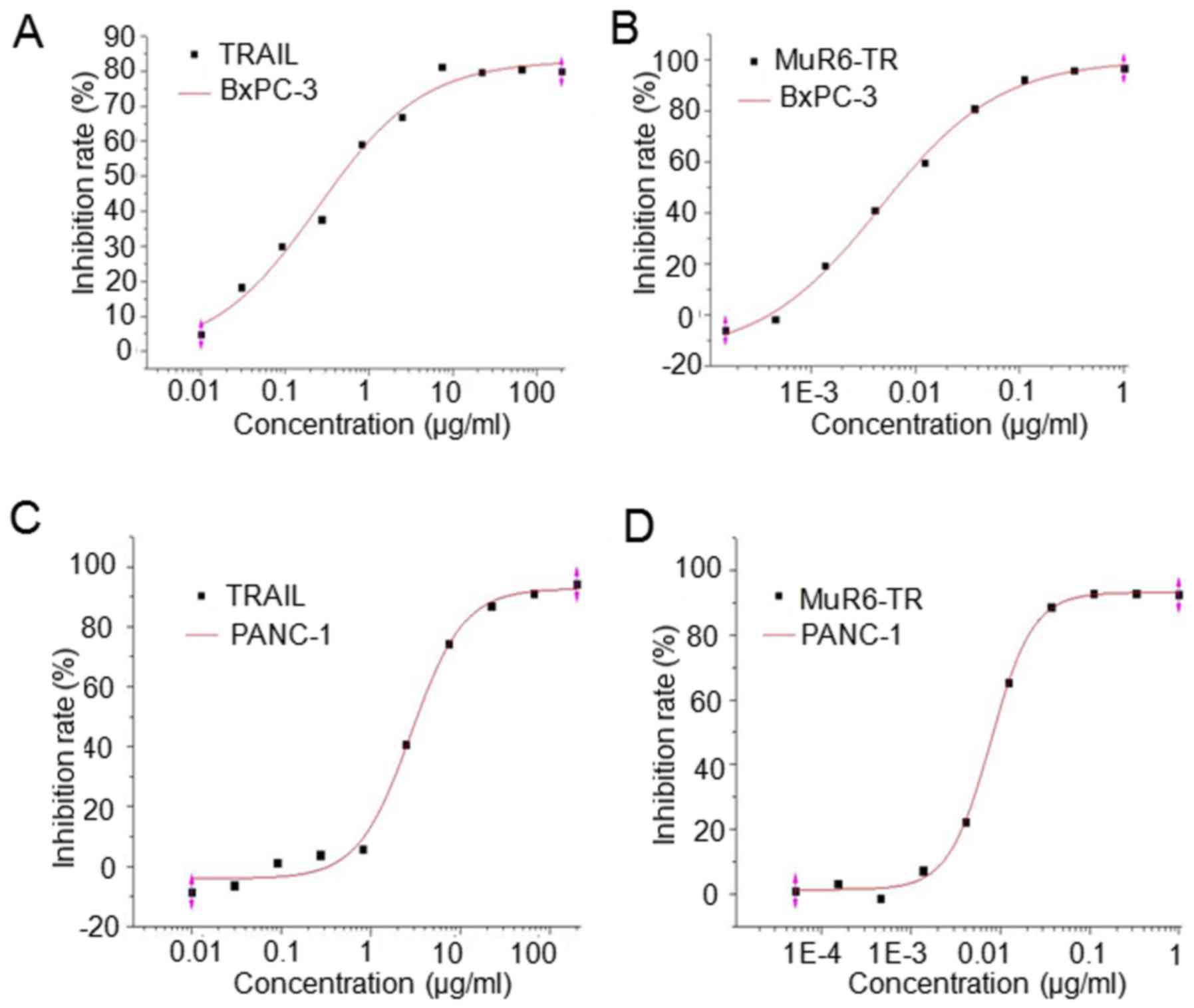
Inhibition of cell growth in culture by
MuR6-TR protein
The antitumor activity of MuR6-TR protein was
measured in pancreatic carcinoma cells BxPc-3 and PANC-1. The
results indicated that the inhibition rate of both cell lines by
MuR6-TR was significantly higher when compared to that by
natural TRAIL when MuR6-TR was at a similar range of
concentrations (P<0.05; Table
IV). The fitting curve showed that the IC50 of
MuR6-TR for both BxPC-3 and PANC-1 cell lines was
significantly lower than that of natural TRAIL (P<0.05; Table V, Fig. 3).
| Table IVProliferation inhibition of BxPC-3
and PANC-1 cells by natural TRAIL and MuR6-TR at different
concentrations. |
Table IV
Proliferation inhibition of BxPC-3
and PANC-1 cells by natural TRAIL and MuR6-TR at different
concentrations.
Natural TRAIL
| MuR6-TR
|
|---|
| Concentration
(µg/ml) | Inhibition rate (%)
| Concentration
(µg/ml) | Inhibition rate (%)
|
|---|
| BxPc-3 | PANC-1 | BxPc-3 | PANC-1 |
|---|
| 200 | 79.890 | 94.191 | 1 | 96.659 | 92.408 |
| 66.667 | 80.561 | 91.061 | 0.333333333 | 95.747 | 92.832 |
| 22.222 | 79.657 | 86.793 | 0.111111111 | 92.254 | 92.872 |
| 7.407 | 81.259 | 74.223 | 0.037037037 | 80.832 | 88.602 |
| 2.469 | 66.873 | 40.823 | 0.012345679 | 59.624 | 65.288 |
| 0.823 | 59.125 | 5.747 | 0.004115226 | 41.020 | 22.361 |
| 0.274 | 37.56 | 3.834 | 0.001371742 | 19.372 | 7.291 |
| 0.091 | 29.993 | 1.147 | 0.000457247 | −1.694 | −1.418 |
| 0.030 | 18.294 | −6.277 | 0.000152416 | −5.981 | 3.178 |
| 0.010 | 4.838 | −8.491 | 0.0000508052 | – | 1.037 |
| Table VIC50 of natural TRAIL and
MuR6-TR in the BxPC-3 and PANC-1 cell lines. |
Table V
IC50 of natural TRAIL and
MuR6-TR in the BxPC-3 and PANC-1 cell lines.
| Natural TRAIL
| MuR6-TRAIL
|
|---|
| BxPC-3 | PANC-1 | BxPC-3 | PANC-1 |
|---|
| IC50
(µg/ml) | 0.284 | 2.817 |
4.63×10−3a |
7.84×10−3b |
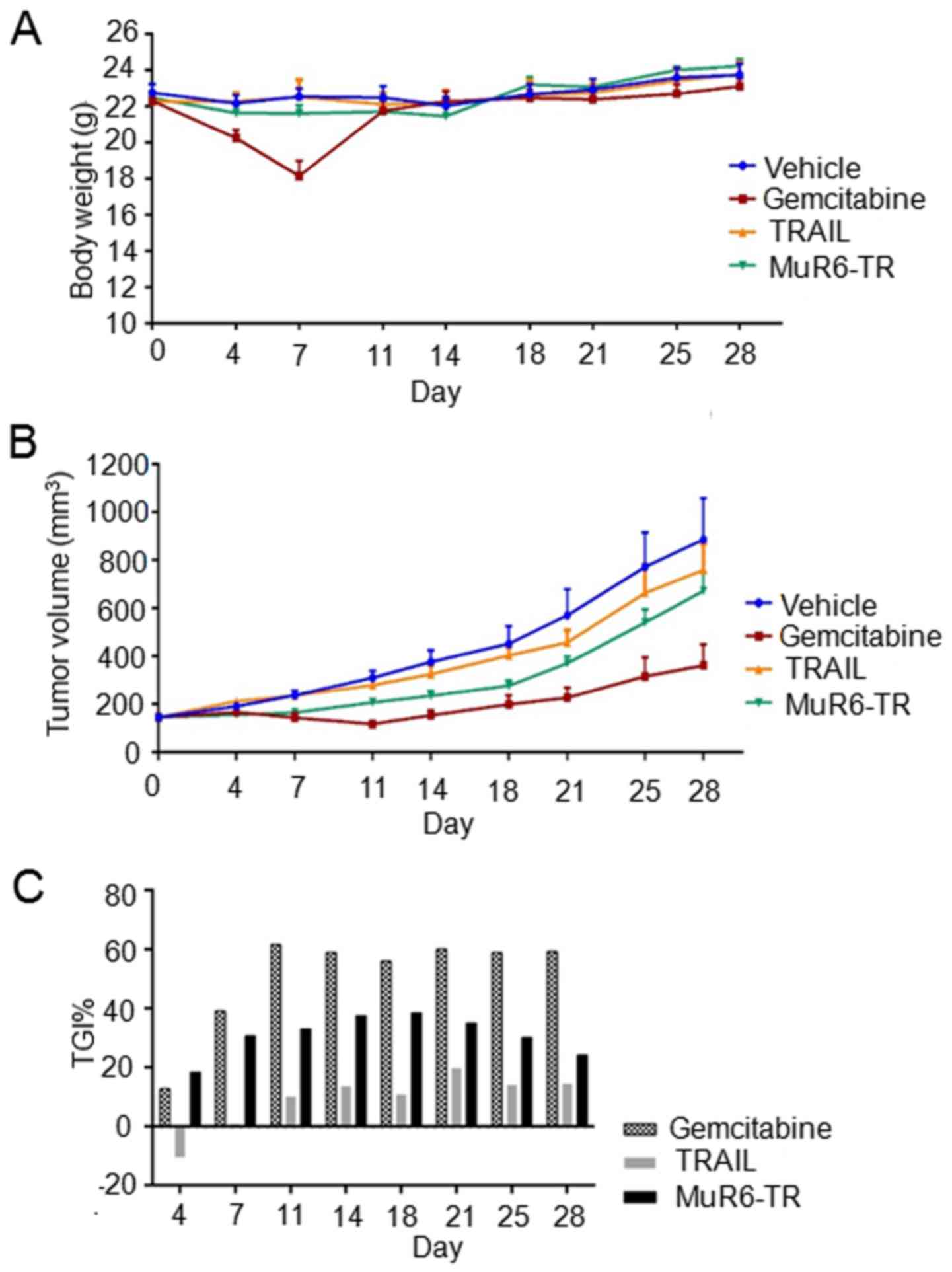
Inhibition of tumor growth in nude mice
by MuR6-TR
The effects of MuR6-TR on the body weight,
tumor growth, tumor inhibition rate and mortality rate of mice are
shown in Fig. 4 and Table VI. As shown in Fig. 4A, the body weight of the mice in
the 4 groups was not significantly increased during the observation
time and there was no significant different among the different
groups (P>0.05); but with exception in the gemcitabine group at
day 7 when there was a significant decrease noted in body weight
(P<0.05).
| Table VIMortality rate of mice in different
groups. |
Table VI
Mortality rate of mice in different
groups.
| Group | Mortality rate (%)
|
|---|
Day
|
|---|
| 0 | 4 | 7 | 11 | 14 | 18 | 21 | 25 | 28 |
|---|
| Vehicle | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gemcitabine | 0 | 0 | 12.5 | 25 | 25 | 25 | 25 | 25 | 25 |
| Natural TRAIL | 0 | 12.5 | 37.5 | 37.5 | 37.5 | 37.5 | 37.5 | 37.5 | 37.5 |
| MuR6-TR | 0 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25a |
The results of tumor growth indicated that the tumor
volumes at the different time-points and in the different groups
were variable. At day 7 after treatment, the tumor volume in the
gemcitabine group was significant smaller than that in the other
groups (P<0.05). Furthermore, the tumor volume in the
MuR6-TR group was significant smaller than that in the
vehicle group and TRAIL group (P<0.05; Fig. 4B). Although the inhibition rate of
tumor growth by MuR6-TR was significantly lower than
gemcitabine, the inhibition rate of tumor growth by MuR6-TR
was significantly higher than that by TRAIL at all observation
time-points (P<0.05; Fig.
4C).
For the mortality rate, the results indicated that
the mortality rate in the MuR6-TR group was significantly
lower than that in the TRAIL group at days 7–28 and similar to that
in the gemcitabine group at days 11–28.
Discussion
In the present study, we mutated the natural TRAIL
protein at 114-119 residues with CPP-like change to form TRAIL
mutant MuR6-TR which was verified by western blotting and
demonstrated the antigen determinant of TRAIL. The MuR6-TR
protein also displayed antitumor effects in pancreatic carcinoma
cell lines and in implanted pancreatic carcinoma tumors in a nude
mouse model. These results indicated that natural TRAIL mutant
MuR6-TR enhanced the sensitivity of tumor cells to apoptosis
inducers and could be a potential targeting therapy for pancreatic
cancer.
Human natural TRAIL is a type II transmembrane
glycoprotein composed of 281 amino acids. The structural domain of
TRAIL C-terminal at residues of 114-281 has several β-motifs to
form hollow tubular structure. Three TRAIL monomers can form
homogenous trimer through the β-motif to display biological
activity (11). The C-terminal of
natural TRAIL can be hydrolyzed by metalloprotease into soluble
functional segments (sTRAIL) while the trimer of either entire
TRAIL or sTRAIL can induce the apoptosis of tumor cells. The defect
of N-terminal amino acid of TRAIL has no effect on the protein
function, which provides the possibility for the directed
reformation of TRAIL.
In a primary study, we modified sTRAIL by forming
CPPs in the N-terminal and found that different CPPs in
classification and molecular weight demonstrated large variation in
the stability, soluble expression and biological activities of
TRAIL (data not shown). After studying the structure of TRAIL, we
found that the arginine (R) dominant VRERGPQR domain in 114-121
residues of C-terminal has a similar structure with the
non-amphipathic pAr of CPPs and is not involved in the specific
spatial conformation of TRAIL. Therefore, we proposed to directly
reform the N-terminal of sTRAIL with CPPs in 114-121 residues of
the C-terminal.
In the study of pAr cell-penetration, R9 is commonly
used because both the linear and branched structures have
cell-penetrating function (12).
A previous study indicated that the cell-penetrating efficacy of
the peptide having <5 or >15 R is obviously decreased
(13). With the success in
mutating VRERGPQR with 5 amino acid into RRRRRRRRR8 (R8) in 114-121
residues of TRAIL (data not shown), we further reduced the mutation
by mutating 4 residues of sTRAIL N-terminal (R6) in the present
study and this mutant was confirmed by PCR. This soluble R6
mutation increased the stability while minimizing the change of the
primary structure of TRAIL to exhibit biological activities.
The expression of exogenous protein by E.
coli is affected by many factors, such as IPTG concentration,
induction time and stability of exogenous protein in the host. The
commonly used concentration of IPTG is 0.1–1.0 mmol/l while a
higher concentration of IPTG can increase the cost and inhibit
bacterial growth (14).
Therefore, minimal IPTG should be used under the prerequisite that
there is no effect on the expression of protein. Our primary study
indicated that 0.1 mM IPTG could achieve ideal expression of
protein while a higher concentration did not increase the
expression but decreased the bacterial sum. Therefore, we set the
IPTG concentration to 0.1 mM. In addition, the induction time is
also critical because earlier induction can inhibit the growth of
bacteria while later induction results in aging bacteria and is bad
for expression. We found that induction after 3 h is the best time
for ideal bacterial production and protein expression. The
fermentation of E. coli at a high density requires reduction
of the production of acetic acid which can inhibit the growth and
expression of bacteria, and the glucose can be replaced by glycerol
which produces less acetic acid (15). Under our current condition, the
final total expression of MuR6-TR was high (80%) which is helpful
for the next purification.
In bacteria, there are nucleic acid, polysaccharose
and other proteins which are existent in complex forms with the
expressed targeting proteins. Therefore, extraction and
purification of the targeting protein is critical. The MuR6-TR
protein has an isoelectric point at 9.96 but no disulfide bond, is
lowly adhesive to regular purifying gels under low-salt condition
and resistant to high-salt (11).
While high ion intensity under high-salt condition can deviate the
practical pI from theoretical pI, which is helpful to explore ideal
condition for ion exchange and eluting to obtain targeting proteins
with higher concentration and purity. For the safety of animals in
subsequent experiments, it is necessary to remove the pyrogen which
is a metabolic endotoxin produced by microbes. The first step of
cation-exchange is mainly to elute the targeting protein from the
added samples containing many unspecific proteins which are mostly
negative-charged. Therefore, it is necessary to perform the second
step of anion-exchange for purification to obtain targeting protein
with high purity. The western blot result indicated that the
purified protein after two-step elution in the present study had
the antigenic determinant and was suitable for the next experiment
in animals.
TRAIL mainly exhibits its effect by binding to
superficial DR4 and DR5 on the cell membrane. In pancreatic tumor
tissues, there is universal expression of TRAIL receptor which
plays an important role in modulating the apoptosis of pancreatic
carcinoma and there is differential expression of the TRAIL
receptor. For example, DR4 and DR5 are expressed much higher in
pancreatic tumor tissues than pancreatic normal tissues. The
expression of DR5 in pancreatic carcinoma tissue is related to the
differentiation degree and malignant degree of tumor tissues while
the expression of DR4 and decoy receptor 1 and 2 (DcR1, DcR2) in
pancreatic carcinoma tissue is not related with the differentiation
degree and clinical stage (16,17). According to a review by Di Pietro
and Zauli (6), Apo2L/TRAIL was
sensitive to 61 of 92 studied primary or passaged tumor cell lines,
having a sensitivity rate of 66.3% and a resistance rate of 33.7%.
Other studies indicated that approximately 50% tumor cell lines are
resistant to TRAIL (18,19). In the present study, both
TRAIL-insensitive pancreatic carcinoma cell lines BxPC-3 and PANC-1
showed a higher growth inhibition rate and lower IC50
with MuR6-TR than natural TRAIL, suggesting that the mutant
protein with N-terminal CCPs had an in vitro advantage in
inhibiting the growth of pancreatic carcinoma.
As a platform to mimic the intrinsic environment of
the body, animal experiments can primarily explore the effect and
safety of novel drugs in the body. According to previous studies,
the TRAIL receptor agonists (including recombinant soluble TRAIL)
and the monoclonal antibodies against TRAIL-R1 and TRAIL-R2 which
are specific for TRAIL-induced apoptosis have been evaluated in the
early stage of clinical experiments in hematologic and solid tumors
including pancreatic carcinoma (20,21). However, most pancreatic carcinoma
cell lines showed low sensitivity to apoptosis induced by TRAIL
although they express basic signaling molecules of the TRAIL system
(22). According to the in
vitro experiment, we selected the pancreatic carcinoma cell
line PANC-1 which is resistant to natural TRAIL but sensitive to
MuR6-TR. The present study indicated that MuR6-TR showed an
antitumor effect in inhibiting the growth of implanted pancreatic
tumors in nude mice. Although lower than gemcitabine, MuR6-TR
demonstrated a more effective effect than natural TRAIL protein in
inhibiting the growth of tumors. There are several explanations for
the lower effect of MuR6-TR than gemcitabine. Firstly, the
mutant MuR6-TR had a higher therapeutic effect but a shorter time
of bioactivity in the body, which is supported by the finding that
MuR6-TR had a similar effect with gemcitabine during the
first 4–7 days. We will conduct continuous administration of the
drug and to change the frequency in future experiments to observe
the antitumor effect. Secondly, the optimal dosage for the mutant
protein requires further investigation. Notably, the
IC50 of MuR6-TR was much lower than TRAIL in the
in vitro experiment and the lower dosage MuR6-TR
demonstrated a similar effect with gemcitabine in vivo at
4–7 days. Therefore, we can increase the dosage to enhance the
antitumor activity after determining the safe dosage of the drug.
Thirdly, the in vitro sensitivity of cell lines to mutant
protein may not represent the in vivo sensitivity. It is
possible to combine current first-line clinical medicines to
investigate the potential antitumor effects.
During the selection of the drug dosage and
administration frequency, we considered the tolerance of the mice,
the convenience for patients in future clinical experiments and the
quantity of MuR6-TR entering cells in order to prolong its
half-life period in the body. These considerations approached the
scheme of q.d. × 5 days which will be further improved in future
experiments. Moreover, 2 of 8 mice (25%) died during the early
stage of the experiment (day 1–4 after administration of MuR6-TR);
however, the mice did not die immediately after the administration
of MuR6-TR. Thus, it was not due to an acute allergic reaction. We
tried to ascertain the reasons of the death by dissection of the
two dead mice. We identified a large area of white necrotic foci in
the livers of the dead mice. Thus, we diagnosed liver toxicity. We
will add a toxicity test in future reseach. In addition, a higher
requirement for the purification protocol in the future is needed.
On the other hand, the mortality rate in the gemcitabine group was
the same at 25%, suggesting that the dosage of gemcitabine should
be adjusted in future experiments.
In summary, the present study mutated TRAIL with
CPPs at the N-terminal and demonstrated that the mutant
MuR6-TR had improved antitumor effects both in vitro
and in vivo compared to the natural TRAIL. The mechanism was
not explored in this study. However, since the structure of mutant
MuR6-TR was similar to natural TRAIL, it is likely to also
display its antitumor effects by binding to the TRAIL receptors on
the tumor cell membrane. There was another TRAIL mutant membrane
penetrating peptide alike (TMPPA) which showed significantly
stronger affinity to the cancer cell membrane compared with natural
TRAIL (data not shown). Thus, we believe that TMPPA enhances the
affinity to the cancer cell membrane. Then, it massively aggregates
on the cancer cell membrane and increases the signal transduction
finally increasing the antitumor effects. Nevertheless, the
therapeutic effect of MuR6-TR and the detailed mechanism
warrant further research.
Acknowledgments
This study was supported by grants from the National
Natural Scientific Foundation of China (nos. 81301962 and
81372444).
References
|
1
|
Singh D, Upadhyay G, Srivastava RK and
Shankar S: Recent advances in pancreatic cancer: Biology,
treatment, and prevention. Biochim Biophys Acta. 1856:13–27.
2015.PubMed/NCBI
|
|
2
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boatright KM, Renatus M, Scott FL,
Sperandio S, Shin H, Pedersen IM, Ricci JE, Edris WA, Sutherlin DP,
Green DR, et al: A unified model for apical caspase activation. Mol
Cell. 11:529–541. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arlt A, Müerköster SS and Schäfer H:
Targeting apoptosis pathways in pancreatic cancer. Cancer Lett.
332:346–358. 2013. View Article : Google Scholar
|
|
5
|
Ashkenazi A: Targeting death and decoy
receptors of the tumour-necrosis factor superfamily. Nat Rev
Cancer. 2:420–430. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Di Pietro R and Zauli G: Emerging
non-apoptotic functions of tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL)/Apo2L. J Cell Physiol.
201:331–340. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu H, Zeng F, Zhang M, Huang F, Wang J,
Guo J, Liu C and Wang H: Emerging landscape of cell penetrating
peptide in reprogramming and gene editing. J Control Release.
226:124–137. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Joseph SC, Blackman BA, Kelly ML, Phillips
M, Beaury MW, Martinez I, Parronchi CJ, Bitsaktsis C, Blake AD and
Sabatino D: Synthesis, characterization, and biological activity of
poly(arginine)-derived cancer-targeting peptides in HepG2 liver
cancer cells. J Pept Sci. 20:736–745. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang W, Zhang N, Zhao T, Liu M, Zhang T
and Li D: Inhibition of tumor growth by polyarginine-fused mutant
cytosine deaminase. Appl Biochem Biotechnol. 175:1633–1643. 2015.
View Article : Google Scholar
|
|
10
|
Wiley SR, Schooley K, Smolak PJ, Din WS,
Huang CP, Nicholl JK, Sutherland GR, Smith TD, Rauch C, Smith CA,
et al: Identification and characterization of a new member of the
TNF family that induces apoptosis. Immunity. 3:673–682. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cha SS, Song YL and Oh BH: Specificity of
molecular recognition learned from the crystal structures of TRAIL
and the TRAIL:sDR5 complex. Vitam Horm. 67:1–17. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alhakamy NA, Dhar P and Berkland CJ:
Charge Type, Charge Spacing, and Hydrophobicity of Arginine-Rich
Cell-Penetrating Peptides Dictate Gene Transfection. Mol Pharm.
13:1047–1057. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fujita T, Furuhata M, Hattori Y, Kawakami
H, Toma K and Maitani Y: High gene delivery in tumor by
intratumoral injection of tetraarginine-PEG lipid-coated
protamine/DNA. J Control Release. 129:124–127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Papaneophytou CP and Kontopidis GA:
Optimization of TNF-α overexpression in Escherichia coli using
response surface methodology: Purification of the protein and
oligomerization studies. Protein Expr Purif. 86:35–44. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eiteman MA and Altman E: Overcoming
acetate in Escherichia coli recombinant protein fermentations.
Trends Biotechnol. 24:530–536. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ozawa F, Friess H, Kleeff J, Xu ZW,
Zimmermann A, Sheikh MS and Büchler MW: Effects and expression of
TRAIL and its apoptosis-promoting receptors in human pancreatic
cancer. Cancer Lett. 163:71–81. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stadel D, Mohr A, Ref C, MacFarlane M,
Zhou S, Humphreys R, Bachem M, Cohen G, Möller P, Zwacka RM, et al:
TRAIL-induced apoptosis is preferentially mediated via TRAIL
receptor 1 in pancreatic carcinoma cells and profoundly enhanced by
XIAP inhibitors. Clin Cancer Res. 16:5734–5749. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stuckey DW and Shah K: TRAIL on trial:
Preclinical advances in cancer therapy. Trends Mol Med. 19:685–694.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maksimovic-Ivanic D, Stosic-Grujicic S,
Nicoletti F and Mijatovic S: Resistance to TRAIL and how to
surmount it. Immunol Res. 52:157–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Humphreys RC and Halpern W: Trail
receptors: Targets for cancer therapy. Adv Exp Med Biol.
615:127–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ashkenazi A, Holland P and Eckhardt SG:
Ligand-based targeting of apoptosis in cancer: The potential of
recombinant human apoptosis ligand 2/Tumor necrosis factor-related
apoptosis-inducing ligand (rhApo2L/TRAIL). J Clin Oncol.
26:3621–3630. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vogler M, Dürr K, Jovanovic M, Debatin KM
and Fulda S: Regulation of TRAIL-induced apoptosis by XIAP in
pancreatic carcinoma cells. Oncogene. 26:248–257. 2007. View Article : Google Scholar
|