Introduction
Hepatitis C virus (HCV) is the leading cause of
serious liver diseases, including fibrosis, cirrhosis and
hepatocellular carcinoma, which affect >185 million people
worldwide and >3 million people are newly infected each year
(1). However, chronic HCV
infection is associated with various extrahepatic manifestations,
and it is estimated that a high proportion of patients with chronic
hepatitis C (CHC) (~90%) may develop at least one extrahepatic
manifestation during the course of the disease (2,3).
Certain extrahepatic manifestations are most likely associated with
the HCV infection, such as mixed cryoglobulinemic syndrome and
B-cell non-Hodgkin lymphoma, while other manifestations are
potentially associated with HCV infection, such as type 2 diabetes
mellitus, membrane proliferative glomerulonephritis, neurological
impairments, sicca syndrome, porphyria cutanea tarda, lichen
planus, autoantibodies, moorens corneal ulcers and cardiovascular
events (3). These linkages are
confirmed following antiviral treatment, which partly ameliorate
the above-mentioned syndromes. However, those extrahepatic
manifestations are, in turn, correlated with poor outcomes,
including accelerated progression of hepatic fibrosis, reduced
sustained virological response rates, and the development of
hepatocellular carcinoma (HCC), resulting in increasing the
morbidity and mortality of CHC (2).
Yet, the mechanism by which the virus evokes the
systemic diseases, including thyroid, cardiovascular, renal, eye
and skin diseases, and lymphomas and diabetes, remain to be
elucidated (4). Certainly, the
mechanisms causing the extrahepatic effects of HCV infection are
likely multifactorial, and include direct or indirect effects
(2,3,5).
Previous studies indicate that HCV directly infects and replicates
in various types of extrahepatic cell, and thus induces
extrahepatic manifestations, although increasing evidence indicates
that it also results from endocrine effects or a heightened immune
reaction with systemic effects (6). Furthermore, successful eradication
of HCV with antiviral drugs was shown to improve certain
extrahepatic effects, which are not always associated with the
severity of liver diseases (7).
Thus, understanding the internal linkages between systemic
manifestations and HCV infection is useful for improving the
treatment of CHC in future.
With the rapid development of microarray techniques,
increasing applications of those high-throughput methods reveal the
pathogenic mechanism of the HCV infection (8–10).
Previous studies demonstrated that the HCV infection affects
signaling pathways associated with diseases, including type II
diabetes signaling, hepatic cholestasis and cancer (11–14). However, the results are not
entirely consistent with one another. The variability may result
from various factors, such as the cell lines, cells with different
statuses, virus genotypes and viral infectious dose. It may also
result from the use of different microarray platforms and different
algorithms (15). The methods of
dealing with these data have recently been developed (16). Gene set enrichment analysis (GSEA)
and signaling pathway impact analysis (SPIA), which are two mature
algorithms and are used proverbially, consider a set of genes as a
group in order to calculate the correlation with different
phenotypes (17). In the present
study, GSEA and SPIA were applied to compare the expression levels
of mRNAs in HCV-infected and -uninfected cells. Using this method,
signaling pathways associated with liver and extrahepatic diseases
were selected, and the important molecules in those pathways were
also demonstrated. These data will contribute to clarifying the
internal driving factors contributing to the systemic diseases of
HCV infection.
Materials and methods
Cells and treatments
Huh7.5 cells and plasmid pFL-J6/JFH/JC1 containing
the full-length chimeric HCV complementary DNA (cDNA) were provided
by Vertex Pharmaceuticals, Inc. (Boston, MA, USA). HCV virus stock
was prepared as previously described (18). Huh7.5 cells were infected with HCV
virus stock (150 IU/cell) or virus mock stock (as a control). After
48 h, the culture supernatants were removed and the cells were
lyzed directly with TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and total intracellular RNAs
were extracted accordingly. mRNA microarray was performed by
Kangchen Biotech Co., Ltd. (Shanghai, China) using Agilent chips
(Agilent Technologies, Santa Clara, CA, USA). Each of the
experiments was independently performed four times.
Data preprocess
All data obtained from the microarray chips were
transferred to the genomic data structure format using the Biobase
package in Bioconductor (version 3.1) (19). The data annotation was applied
using the package, hgug4112a.db (20). Data from four arrays were combined
and quantile normalized prior to analysis (21), then transformed to a
log2 scale (22), and
batch effects were removed using the sva package in Bioconductor
(23).
GSEA
GSEA analysis of signaling pathways and genes was
performed using the category package in version 3.1 of Bioconductor
(24). The goal of GSEA analysis
is to determine whether the members of a gene set 'S' are randomly
distributed throughout the entire reference gene list 'L' or are
primarily found at the top or bottom of the list. Gene sets
represented by ≤5 genes were excluded. The t-statistic mean of the
genes was computed in each Kyoto Encyclopedia of Genes and Genomes
(KEGG) pathway. The information regarding the pathways was obtained
from KEGG.db in Bioconductor (25). Using a permutation test with
10,000 permutations, the cut-off of the significance level was
determined as P<0.05 to indicate the significant signaling
pathways associated with HCV infection.
SPIA
SPIA analysis was performed using the SPIA package
in version 3.1 of Bioconductor (26). SPIA calculated the P-value combing
the different expression gene data with the perturbation of
signaling pathways. In this case, the different expression genes
were confirmed by applying the limma package (27). Genes with P<0.05 by row t-test
were selected. The detailed procedure of SPIA was performed
according to a previous study (28).
Anni 2.1
The association between a gene and the HCV infection
was analyzed using the online tool Anni 2.1 (http://biosemantics.org/index.php/software/anni).
The calculation was performed as previously described (29). The entry of each pathway, such as
map04010 for mitogen-activated protein kinases (MAPK) signaling
pathway was submitted, along with the key word 'HCV' as a pair to
Anni 2.1. Anni 2.1 retrieves data from MEDLINE (https://www.medline.com) to score the association
between HCV and each gene in these signaling pathways. The
association score is based on concepts co-occurrences and a higher
score indicates a greater association between a gene and HCV.
Pathview
The visualization of gene changing in signaling
pathways was performed using the Pathview package in version 3.1 of
Bioconductor (30).
Results
To investigate the internal driving factors
contributing to systemic diseases of the HCV infection, Huh7.5
cells were infected with HCV genotype 2a (virus group) or virus
mock stock (control group), and the intracellular total RNAs were
extracted after 48 h. The level of mRNAs was determined using
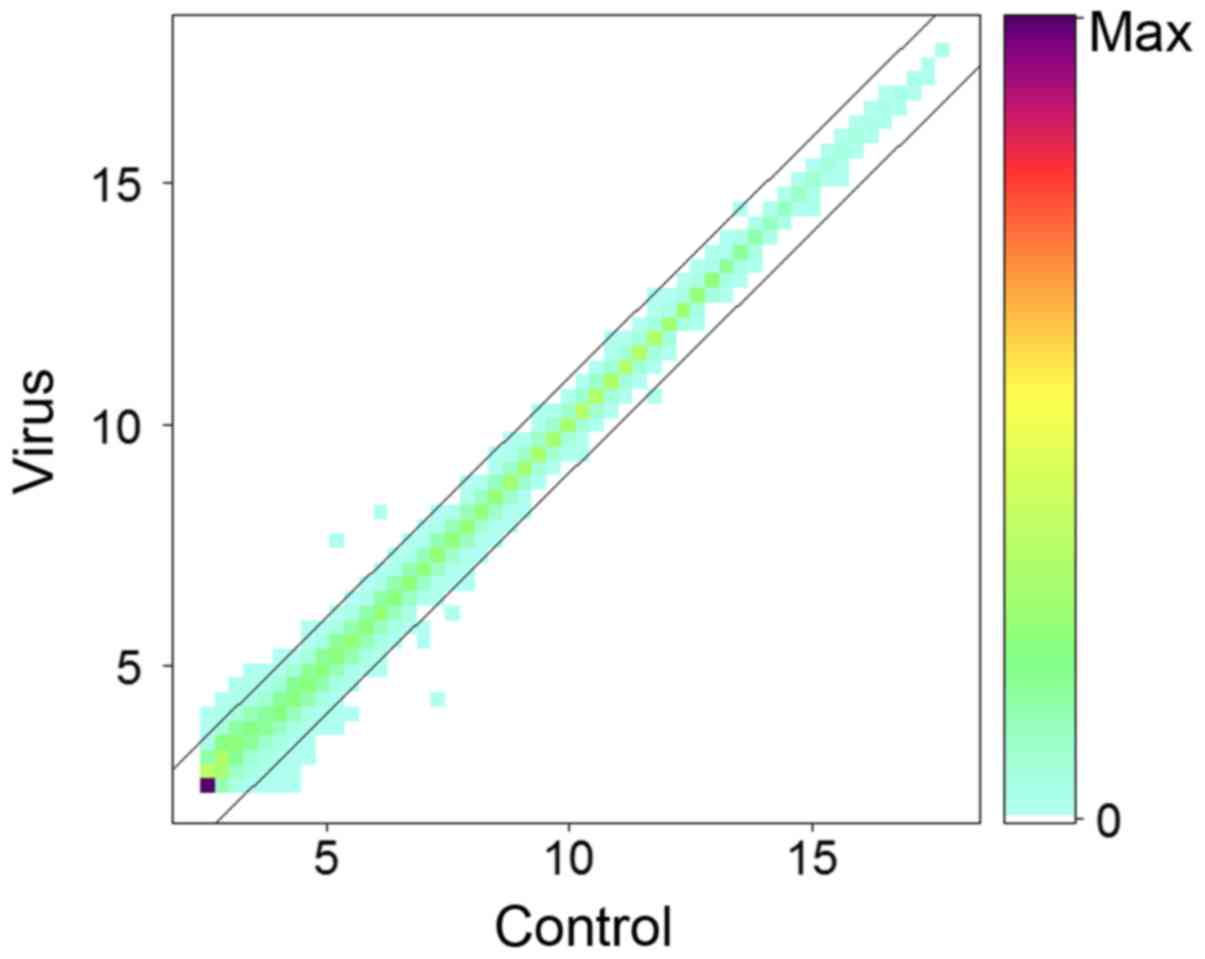
Agilent chips. Following normalization, the numerical average level
of genes in virus and control groups at log2 scale was
plotted (Fig. 1). Results
indicated that the majority of genes remain unchanged >2-fold or
<0.5-fold following infection with HCV, which is consistent with
previous studies (31).
Subsequently, the P-values were calculated using
GSEA or SPIA. In the results obtained with GSEA, 40 pathways were
selected with P<0.05 (Table
I), including signaling pathways with functions of human
diseases, metabolism, organismal systems, cellular processes,
genetic information processing and environmental information
processing. Among them, 35 signaling pathways were inhibited and
five signaling pathways were activated by the HCV infection, and
hepatitis C and hedgehog signaling pathways were previously
confirmed (32). However, in the
result calculated with SPIA, only eight signaling pathways were
selected (Table II), including
gastric acid secretion, the MAPK signaling pathway,
cytokine-cytokine receptor interaction, dilated cardiomyopathy,
amoebiasis, basal cell carcinoma, human T-cell leukemia virus type
I infection and cholinergic synapse. However, apart from the
gastric acid secretion pathway, these signaling pathways were not
included in the results selected with GSEA.
 | Table IPathways selected with gene set
enrichment analysis. |
Table I
Pathways selected with gene set
enrichment analysis.
| Name | P-value | Status | Function |
|---|
| Cell cycle | 0.042 | Inhibited | Cellular processes;
cell growth and death |
| Adherens
junction | 0.042 | Inhibited | Cellular processes;
cellular community |
| Hedgehog signaling
pathway | 0.014 | Activated | Environmental
information processing; signal transduction |
| ErbB signaling
pathway | 0.014 | Inhibited | Environmental
information processing; signal transduction |
| Ubiquitin mediated
proteolysis | 0.014 | Inhibited | Genetic information
processing; folding, sorting and degradation |
| Basal transcription
factors | 0.042 | Inhibited | Genetic information
processing; transcription |
| Pathways in
cancer | 0.014 | Inhibited | Human diseases;
cancers |
| Prostate
cancer | 0.014 | Inhibited | Human diseases;
cancers |
| Melanoma | 0.014 | Inhibited | Human diseases;
cancers |
| Non-small cell lung
cancer | 0.014 | Inhibited | Human diseases;
cancers |
| Bladder cancer | 0.014 | Inhibited | Human diseases;
cancers |
| Thyroid cancer | 0.025 | Inhibited | Human diseases;
cancers |
| Chronic myeloid
leukemia | 0.027 | Inhibited | Human diseases;
cancers |
| Small cell lung
cancer | 0.027 | Inhibited | Human diseases;
cancers |
| Pancreatic
cancer | 0.029 | Inhibited | Human diseases;
cancers |
| Colorectal
cancer | 0.042 | Inhibited | Human diseases;
cancers |
| Renal cell
carcinoma | 0.043 | Inhibited | Human diseases;
cancers |
| Glioma | 0.044 | Inhibited | Human diseases;
cancers |
| Endometrial
cancer | 0.045 | Inhibited | Human diseases;
cancers |
| Systemic lupus
erythematosus | 0.014 | Inhibited | Human diseases;
immune diseases |
| Hepatitis C | 0.014 | Inhibited | Human diseases;
infectious diseases |
| Bacterial invasion
of epithelial cells | 0.027 | Inhibited | Human diseases;
infectious diseases |
| Pathogenic
Escherichia coli infection | 0.040 | Inhibited | Human diseases;
infectious diseases |
| Shigellosis | 0.043 | Inhibited | Human diseases;
infectious diseases |
| Histidine
metabolism | 0.027 | Inhibited | Metabolism; amino
acid metabolism |
| Tryptophan
metabolism | 0.029 | Inhibited | Metabolism; amino
acid metabolism |
| Nitrogen
metabolism | 0.029 | Inhibited | Metabolism; energy
metabolism |
| Glycosaminoglycan
biosynthesis - heparan sulfate | 0.043 | Activated | Metabolism; glycan
biosynthesis and metabolism |
| Glycosphingolipid
biosynthesis - globo series | 0.045 | Activated | Metabolism; glycan
biosynthesis and metabolism |
| One carbon pool by
folate | 0.014 | Inhibited | Metabolism;
metabolism of cofactors and vitamins |
| Porphyrin and
chlorophyll metabolism | 0.040 | Activated | Metabolism;
metabolism of cofactors and vitamins |
| Dorso-ventral axis
formation | 0.042 | Inhibited | Organismal systems;
development |
| Vitamin digestion
and absorption | 0.027 | Inhibited | Organismal systems;
digestive system |
| Gastric acid
secretion | 0.045 | Inhibited | Organismal systems;
digestive system |
| Circadian
rhythm-mammal | 0.031 | Inhibited | Organismal systems;
environmental adaptation |
| Collecting duct
acid secretion | 0.030 | Activated | Organismal systems;
excretory system |
| Proximal tubule
bicarbonate reclamation | 0.044 | Inhibited | Organismal systems;
excretory system |
| Nucleotide
oligomerization domain-like receptor signaling pathway | 0.042 | Inhibited | Organismal systems;
immune system |
| Leukocyte
transendothelial migration | 0.043 | Inhibited | Organismal systems;
immune system |
| Neurotrophin
signaling pathway | 0.027 | Inhibited | Organismal systems;
nervous system |
| Table IIPathways selected with signaling
pathway impact analysis. |
Table II
Pathways selected with signaling
pathway impact analysis.
| Name | P-value | Status | Function |
|---|
| Mitogen-activated
protein kinases signaling pathway | 0.011 | Inhibited | Environmental
information processing; signal transduction |
| Cytokine-cytokine
receptor interaction | 0.017 | Inhibited | Environmental
information processing; signaling molecules and interaction |
| Basal cell
carcinoma | 0.025 | Inhibited | Human diseases;
cancers |
| Dilated
cardiomyopathy | 0.021 | Activated | Human diseases;
cardiovascular diseases |
| Amoebiasis | 0.023 | Inhibited | Human diseases;
infectious diseases |
| Human T-cell
leukemia virus type I infection | 0.039 | Inhibited | Human diseases;
infectious diseases |
| Gastric acid
secretion | 0.008 | Inhibited | Organismal systems;
digestive system |
| Cholinergic
synapse | 0.042 | Inhibited | Organismal systems;
nervous system |
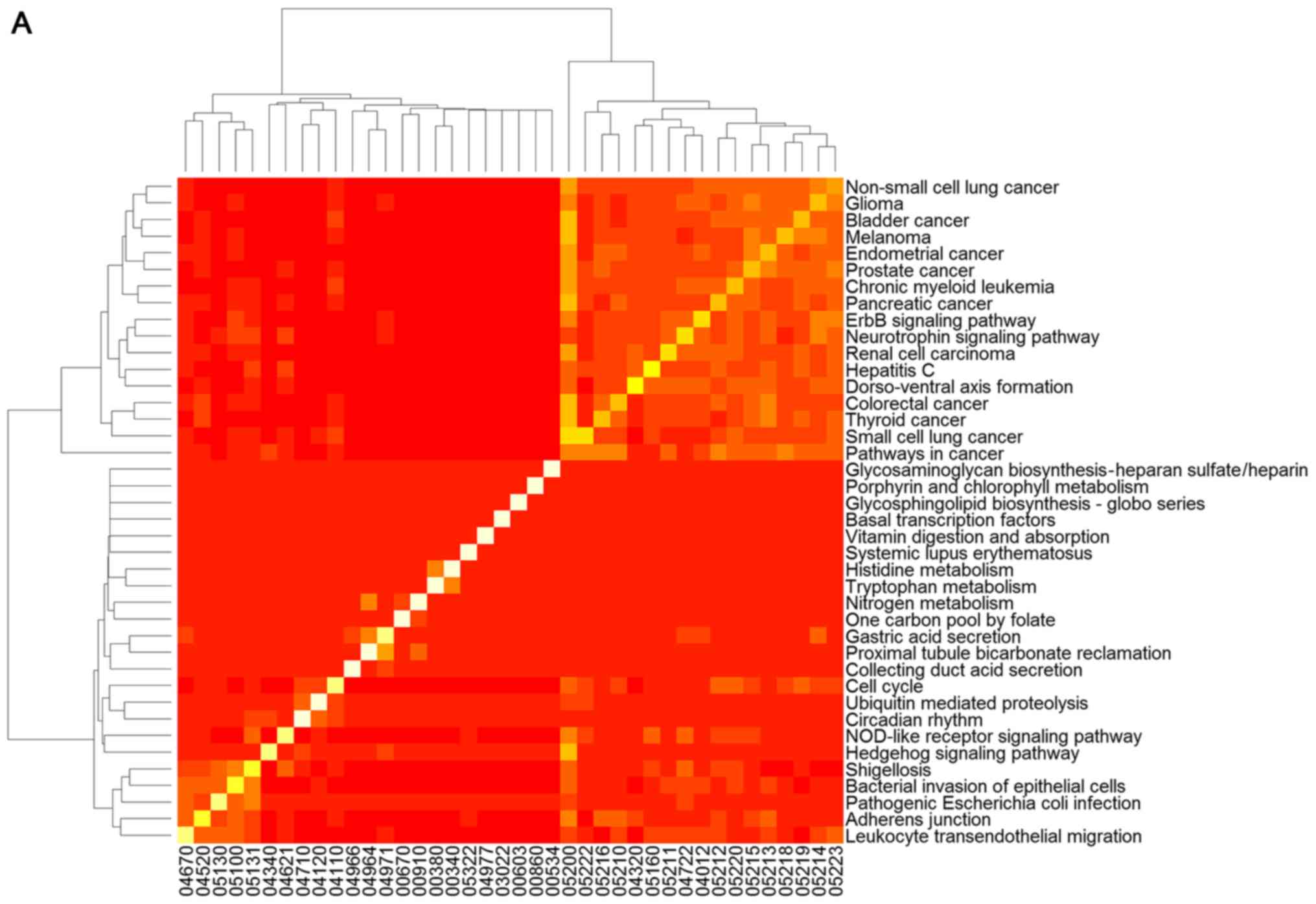
To clarify the association between signaling
pathways, the overlap genes in signaling pathways were further
assessed and heat-maps of the pathways were plotted (Fig. 2). The results clearly demonstrate
that changed genes overlapped more in the two clusters of signaling
pathways selected using GSEA (Fig.
2A). One cluster included the signaling pathway of hepatitis C
and pathways predominantly associated with cancer, including
non-small cell lung cancer, glioma, bladder cancer, melanoma,
endometrial cancer, prostate cancer, chronic myeloid leukemia,
pancreatic cancer, the ErbB and neurotrophin signaling pathways,
renal cell carcinoma, dorso-ventral axis formation, colorectal
cancer, thyroid cancer, small cell lung cancer, and pathways in
cancer (Fig. 2A). The other
cluster was comprised of the hedgehog signaling pathway,
shigellosis, bacterial invasion of epithelial cells, pathogenic
Escherichia coli infection, adherens junction and leukocyte
transendothelial migration, which was predominantly associated with
infection diseases (Fig. 2A).
However, overlap genes between signaling pathways selected using
GSEA were relatively few (Fig.
2B).
To analyze the biological significance of these
selected pathways, the average score of signaling pathways was
calculated to evaluate the biological significance of the pathway
and highlight the gene with the highest score in the signaling
pathway using the online tool Anni 2.1. Six signaling pathways from
SPIA and 11 signaling pathways from GSEA (pathways with P<0.025)
were selected (Table III). The
results demonstrated that the score of the prostate cancer
signaling pathway was the highest, followed by the hepatitis C
pathway, and SOS1 was the highest scoring gene and made the most
contribution to six of the signaling pathways (Table III). However, the average score
of signaling pathways identified by GSEA is higher than that
identified by SPIA (0.0085 vs. 0.0045; P<0.025).
| Table IIIScore of each signaling pathway and
the highest score gene in the pathway. |
Table III
Score of each signaling pathway and
the highest score gene in the pathway.
| Signaling
pathway | Algorithm | Score | Highest scoring
gene |
|---|
| Prostate
cancer | GSEA | 0.0219 | SOS1 |
| Hepatitis C | GSEA | 0.0135 | SOS1 |
| Non-small cell lung
cancer | GSEA | 0.0103 | SOS1 |
| Bladder cancer | GSEA | 0.0102 | RAF1 |
| ErbB signaling
pathway | GSEA | 0.0085 | SOS1 |
| Melanoma | GSEA | 0.0076 | RAF1 |
| Cytokine-cytokine
receptor interaction | GSEA | 0.0073 | IFNA2 |
| Pathways in
cancer | GSEA | 0.0065 | SOS1 |
| Systemic lupus
erythematosus | GSEA | 0.0059 | IFNG |
| Amoebiasis | SPIA | 0.0057 | IFNG |
| MAPK signaling
pathway | SPIA | 0.0050 | SOS1 |
| One carbon pool by
folate | GSEA | 0.0048 | MTHFR |
| Dilated
cardiomyopathy | SPIA | 0.0043 | IGF1 |
| Gastric acid
secretion | SPIA | 0.0038 | CALM3 |
| Ubiquitin mediated
proteolysis | GSEA | 0.0036 | UBE2B |
| Basal cell
carcinoma | SPIA | 0.0036 | TP53 |
| Hedgehog signaling
pathway | GSEA | 0.0036 | BMP7 |
Discussion
To understand the internal driving factors
contributing to the systemic manifestation of the HCV infection, a
comprehensive analysis was performed on microarray data of mRNAs in
HCV-infected and -uninfected Huh7.5 cells using GSEA and SPIA, and
40 signaling pathways that were significantly activated or
inhibited were selected using GSEA, and eight pathways that were
significantly changed were selected using SPIA, although only one
signaling pathway overlapped with GSEA. Indeed, there were few
instances of overlapping signaling pathways in GSEA and SPIA, and
similar differences were previously reported (15,33). The reason may be that GSEA and
SPIA are based upon different theories. GSEA is based upon
functional class scoring (FCS) methods. Compared with traditional
over-representation analysis (ORA) methods, GSEA is accurate and
meaningful as it does not ignore molecular measurements when
identifying significant pathways. SPIA uses pathway topology
(PT)-based methods. Compared with FCS methods, SPIA uses topology
information of signaling pathways when identifying significant
pathways. It is more accurate and meaningful than FCS methods.
However, PT-based methods do have limitations, as the knowledge of
signaling pathways is not fully understood and there is only
fragmented knowledge. In addition, the integrity of topology
information of signaling pathways will largely affect results. With
improvement of annotations, PT-based methods are expected to become
more valuable. In view of this, GSEA and SPIA algorithms were
applied to demonstrate the signaling pathways in different
landscapes. These pathways provided important clues for further
clarifying the intrinsic associations between systemic syndromes of
HCV infection.
Cryoglobulinemia, B-cell lymphoma and thyroid
disorders are the major extrahepatic manifestations; however, the
three disorders are not included in the KEGG pathway database,
therefore they are not shown when applying KEGG-based statistics.
The two algorithms, GSEA and SPIA, determine that certain signaling
pathways include genes associated with these diseases. Types II and
III cryoglobulinemia, which are strongly associated with HCV
infection, tend to be associated with immune disease usually
accompanied by abnormal antibodies (34). In the present results, systemic
lupus erythematosus, nucleotide oligomerization domain (NOD)-like
receptor signaling pathway and leukocyte transendothelial migration
were observed. As the biological process by which HCV causes
cryoglobulinemia is not fully understood, the present results may
provide insight into the investigation of the internal driving
factors related with the disease after HCV infection. B-cell
lymphoma is a cancer of the B lymphocytes. In the present results,
15 signaling pathways of cancer were identified, notably the
pathway termed 'pathways in cancer'. A previous study indicated
that NOD-like receptors are key in human B lymphocytes (35). Certainly, NOD-like receptor
signaling pathway was indeed selected out in our result. In
addition to that, the signaling pathway of thyroid cancer was
identified to include certain genes associated with thyroid
disorders. Those results indicate that the present experiments and
the algorithms are valuable.
HCV is the leading cause of HCC and its infection
affects cancer-associated pathways. Following analysis with GSEA,
18 signaling pathways in the functional class of human diseases
were selected, in which 13 signaling pathways are in the subclass
of cancer, one signaling pathway is in the subclass of immune
diseases (systemic lupus erythematosus) and four pathways are in
the subclass of infectious diseases (Table I). In addition, cancer-associated
pathways were selected using SPIA, although far fewer than with
GSEA (Table II). However, the
HCV infection often leads to abnormal metabolism (36). Indeed, certain metabolism pathways
were selected using GSEA (Tables
I and II). There are other
syndromes in HCV-infected patients, such as left ventricular
systolic, diastolic dysfunction and cardiac arrhythmias, although
the intrinsic associations with HCV infection remain unclear
(37). Signaling pathways were
selected with GSEA or SPIA, which were involved with environmental
information processing, organismal systems, and certain signaling
pathways may have direct or indirect associations with HCV
(Tables I and II).
It's important to note that the majority of these
signaling pathways have been reported to be associated with HCV
infection (11–14), which confirmed the two algorithms
are effective in selecting pathways. However, seven of the
signaling pathways provide novel clues to demonstrate the driving
factor for extrahepatic manifestations of HCV infection, including
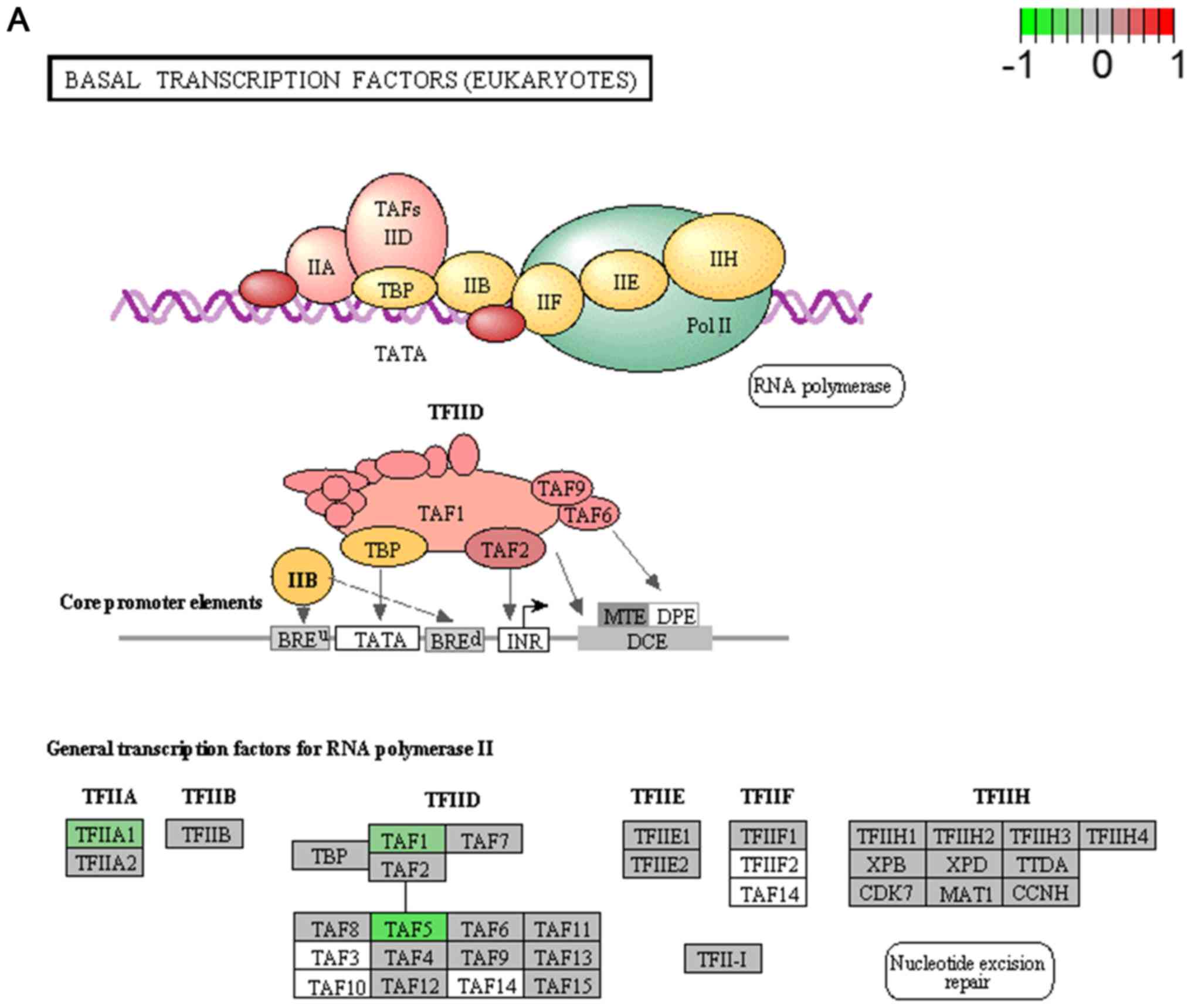
basal transcription factors (Fig.
3A), pathogenic Escherichia coli infection (Fig. 3B), shigellosis (Fig. 3C), gastric acid secretion
(Fig. 3D), dorso-ventral axis
formation (Fig. 3E), amoebiasis
(Fig. 3F) and cholinergic synapse
(Fig. 3G). Notably, the pathway
of gastric acid secretion (Fig.
3D) was selected by the two algorithms in the present study. In
this signaling pathway, genes are clearly influenced by the HCV
infection (Fig. 3D). Gastric acid
secretion is associated with liver cirrhosis caused by the
hepatitis virus infection (38),
and is also correlated with acute pancreatitis (39,40). Previous studies demonstrated that
acute pancreatitis is associated with viral hepatitis (41–43), in particular, a case of acute
pancreatitis was described with associated HCV infection (43). However, the direct associations
between HCV infection and gastric acid secretion or acute
pancreatitis require further experiments in order to be clarified.
Basal transcriptional factors (Fig.
3A), also termed general transcription factors (GTFs), are a
class of protein transcription factors, and the factors TFIIA,
TFIIB, TFIID, TFIIE, TFIIF and TFIIH are required for RNA
polymerase II to bind to a promoter (44). Certainly, HCV replication indeed
influences these factors, it was reported that HCV protein, NS5A
interacts with TFIID (45). In
the present study, various factors of TFIIA and TFIID were
inhibited by HCV infection. Shigellosis (Fig. 3C), amoebiasis (Fig. 3F) and pathogenic Escherichia
coli infection (Fig. 3B) are
three different types of infectious disease. No direct connection
was reported between these infections and the HCV infection.
However, certain viral infections may be a risk factor to other
infections; for example, the HIV infection may be a risk factor for
acquiring shigellosis (46–48). Cholinergic synapse (Fig. 3G) is predominantly about the
biological process of acetylcholine (ACh). ACh participates in
numerous functions, such as learning, memory, attention and motor
control. Clinically, the HCV infection causes nervous system
dysfunction, which is primarily caused by upregulation of the host
immune response with production of autoantibodies, immune complexes
and cryoglobulins (49); the
present results may demonstrate a novel insight into this
pathogenesis. However, to the best of our knowledge, there is no
evidence to indicate that dorso-ventral axis formation (Fig. 3E) is associated with the HCV
infection.
Using Anni 2.1, single genes were selected from the
selected signaling pathways for further analysis (Table III). Those top genes selected
with the Anni tool demonstrate their significance in the signaling
pathways and associations with the HCV infection. However, the
level of these does not necessarily change in the pathways, as the
algorithm of the Anni tool did not consider the fold-changes of
genes. Furthermore, those genes in Table III did not present obvious
changes in the present experiments. However, the change in the
protein levels cannot be excluded. SOS1, a guanine
nucleotide exchange factor for Ras/Rac, was the highest scoring
gene and contributed to six signaling pathways (Table III). Although no direct
association has been reported between SOS1 and the HCV infection,
it provides novel evidence to further illustrate the outcome that
HCV infection causes HCC, as a previous study demonstrated that
SOS1 is closely associated with cancer (50). The next highest scoring gene was
Raf1, which is an enzyme in the extracellular
signal-regulated kinases 1/2 signaling pathway that performs a
regulatory role (51). It was
reported that constitutive expression of the HCV core results in a
high basal activity of Raf1 (52) and knockdown of Raf1
inhibits HCV replication (53),
indicating that Raf1 may be important in the HCV life cycle.
The third was IFNA2, which is a gene encoded by prototypic
human interferon α (IFNα). It is associated with antivirus
activity, as the recombinant IFNα2 has been approved for the
treatment of HCV and its pegylated forms (PEG-IFNα2) are the most
widely administered anti-viral agents (54). Furthermore, IFNγ is a human IFN
and contributes to auto-immune activities, and methylene
tetrahydrofolate reductase, insulin-like growth factor and
TP53 exhibit direct or indirect associations with HCC
following infection with HCV (53–57). These data may provide further
evidence to investigate the associations between the HCV infection
and syndromes exhibited by patients. In addition, illustration of
the underlying mechanisms of those molecules may provide potential
targets for interrupting the internal driving factors contributing
to the extrahepatic manifestations of the HCV infection.
In conclusion, the present study demonstrated seven
novel pathways for further clarifying the direct intrinsic factors
contributing to systemic manifestations of the HCV infection,
including basal transcription factors, pathogenic Escherichia
coli infection, shigellosis, gastric acid secretion,
dorso-ventral axis formation, amoebiasis and cholinergic synapse,
and identified 10 genes, SOS1, RAF1, IFNA2,
IFNG, MTHFR, IGF1, CALM3, UBE2B,
TP53 and BMP7, whose expression may be key in those
pathways.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81321004 and
81322050), the National Mega-Project for 'R&D for Innovative
Drugs', Ministry of Science and Technology of China (grant no.
2012ZX09301-002-001), the Ministry of Education of China (grant no.
NCET-12-0072). The authors would also like to thank Kangchen
Biotech Co., Ltd. (Shanghai, China) for performing the mRNA
microarray detection using Agilent chip technology.
References
|
1
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273 e1261. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Negro F, Forton D, Craxi A, Sulkowski MS,
Feld JJ and Manns MP: Extrahepatic morbidity and mortality of
chronic hepatitis C. Gastroenterology. 149:1345–1360. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tang L, Marcell L and Kottilil S: Systemic
manifestations of hepatitis C infection. Infect Agent Cancer.
11:292016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sherman AC and Sherman KE: Extrahepatic
manifestations of hepatitis C infection: Navigating CHASM. Curr
HIV/AIDS Rep. 12:353–361. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gill K, Ghazinian H, Manch R and Gish R:
Hepatitis C virus as a systemic disease: Reaching beyond the liver.
Hepatol Int. 10:415–423. 2016. View Article : Google Scholar :
|
|
6
|
Zignego AL, Gragnani L, Piluso A,
Sebastiani M, Giuggioli D, Fallahi P, Antonelli A and Ferri C:
Virus-driven autoimmunity and lymphoproliferation: The example of
HCV infection. Expert Rev Clin Immunol. 11:15–31. 2015. View Article : Google Scholar
|
|
7
|
Nuno Solinis R, Arratibel Ugarte P, Rojo A
and Sanchez Gonzalez Y: Value of treating all stages of chronic
hepatitis C: A comprehensive review of clinical and economic
evidence. Infect Dis Ther. 5:491–508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee CM, Hung CH, Lu SN and Changchien CS:
Hepatitis C virus genotypes: Clinical relevance and therapeutic
implications. Chang Gung Med J. 31:16–25. 2008.PubMed/NCBI
|
|
9
|
Waldron PR and Holodniy M: MicroRNA and
hepatitis C virus - challenges in investigation and translation: A
review of the literature. Diagn Microbiol Infect Dis. 80:1–12.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rahman W, Tu T, Budzinska M, Huang P,
Belov L, Chrisp JS, Christopherson RI, Warner FJ, Bowden DS,
Thompson AJ, et al: Analysis of post-liver transplant hepatitis C
virus recurrence using serial cluster of differentiation antibody
microarrays. Transplantation. 99:e120–e126. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Woodhouse SD, Narayan R, Latham S, Lee S,
Antrobus R, Gangadharan B, Luo S, Schroth GP, Klenerman P and
Zitzmann N: Transcriptome sequencing, microarray, and proteomic
analyses reveal cellular and metabolic impact of hepatitis C virus
infection in vitro. Hepatology. 52:443–453. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blackham S, Baillie A, Al-Hababi F,
Remlinger K, You S, Hamatake R and McGarvey MJ: Gene expression
profiling indicates the roles of host oxidative stress, apoptosis,
lipid metabolism, and intracellular transport genes in the
replication of hepatitis C virus. J Virol. 84:5404–5414. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Papic N, Maxwell CI, Delker DA, Liu S,
Heale BS and Hagedorn CH: RNA-sequencing analysis of 5′ capped RNAs
identifies many new differentially expressed genes in acute
hepatitis C virus infection. Viruses. 4:581–612. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Walters KA, Syder AJ, Lederer SL, Diamond
DL, Paeper B, Rice CM and Katze MG: Genomic analysis reveals a
potential role for cell cycle perturbation in HCV-mediated
apoptosis of cultured hepatocytes. PLoS Pathog. 5:e10002692009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cai B and Jiang X: Revealing biological
pathways implicated in lung cancer from TCGA gene expression data
using gene set enrichment analysis. Cancer Inform. 13:113–121.
2014.PubMed/NCBI
|
|
16
|
Hira ZM and Gillies DF: A Review of
feature selection and feature extraction methods applied on
microarray data. Adv Bioinformatics. 2015:1983632015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Braun R: Systems analysis of
high-throughput data. Adv Exp Med Biol. 844:153–187. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Peng ZG, Zhao ZY, Li YP, Wang YP, Hao LH,
Fan B, Li YH, Wang YM, Shan YQ, Han YX, et al: Host apolipoprotein
B messenger RNA-editing enzyme catalytic polypeptide-like 3G is an
innate defensive factor and drug target against hepatitis C virus.
Hepatology. 53:1080–1089. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gentleman RC, Carey VJ, Bates DM, Bolstad
B, Dettling M, Dudoit S, Ellis B, Gautier L, Ge Y, Gentry J, et al:
Bioconductor: Open software development for computational biology
and bioinformatics. Genome Biol. 5:R802004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Carlson M: Agilent 'Human Genome, Whole'
annotation data (chip hgug4112a). R package version 3.2.3. 2016
|
|
21
|
Bolstad BM, Irizarry RA, Astrand M and
Speed TP: A comparison of normalization methods for high density
oligonucleotide array data based on variance and bias.
Bioinformatics. 19:185–193. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Robyr R, Boulvain M, Lewi L, Huber A,
Hecher K, Deprest J and Ville Y: Cervical length as a prognostic
factor for preterm delivery in twin-to-twin transfusion syndrome
treated by fetoscopic laser coagulation of chorionic plate
anastomoses. Ultrasound Obstet Gynecol. 25:37–41. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Johnson WE, Li C and Rabinovic A:
Adjusting batch effects in microarray expression data using
empirical Bayes methods. Biostatistics. 8:118–127. 2007. View Article : Google Scholar
|
|
24
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES, et al: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Carlson M, Falcon S, Pages H and Li N:
KEGG.db: A set of annotation maps for KEGG. KEGG.db. 2012.
|
|
26
|
Draghici ALTaPKaS: SPIA: Signaling Pathway
Impact Analysis (SPIA) using combined evidence of pathway
over-representation and unusual signaling perturbations. (R package
version 2.28.0). 2013
|
|
27
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tarca AL, Draghici S, Khatri P, Hassan SS,
Mittal P, Kim J, Kim CJ, Kusanovic JP and Romero R: A novel
signaling pathway impact analysis. Bioinformatics. 25:75–82. 2009.
View Article : Google Scholar :
|
|
29
|
Jelier R, Schuemie MJ, Veldhoven A,
Dorssers LC, Jenster G and Kors JA: Anni 2.0: A multipurpose
text-mining tool for the life sciences. Genome Biol. 9:R962008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luo W and Brouwer C: Pathview: An
R/Bioconductor package for pathway-based data integration and
visualization. Bioinformatics. 29:1830–1831. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Muffak-Granero K, Bueno P, Olmedo C,
Comino AM, Hassan L, Garcia-Alcalde F, Serradilla M, García-Navarro
A, Mansilla A, Villar JM, et al: Study of gene expression profile
in liver transplant recipients with hepatitis C virus. Transplant
Proc. 40:2971–2974. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Choi SS, Bradrick S, Qiang G, Mostafavi A,
Chaturvedi G, Weinman SA, Diehl AM and Jhaveri R: Up-regulation of
Hedgehog pathway is associated with cellular permissiveness for
hepatitis C virus replication. Hepatology. 54:1580–1590. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Haynes WA, Higdon R, Stanberry L, Collins
D and Kolker E: Differential expression analysis for pathways. PLOS
Comput Biol. 9:e10029672013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cacoub P and Comarmond C: New insights
into HCV-related rheumatologic disorders: A review. J Adv Res.
8:89–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Petterson T, Jendholm J, Månsson A,
Bjartell A, Riesbeck K and Cardell LO: Effects of NOD-like
receptors in human B lymphocytes and crosstalk between NOD1/NOD2
and Toll-like receptors. J Leukoc Biol. 89:177–187. 2011.
View Article : Google Scholar
|
|
36
|
Modaresi Esfeh J and Ansari-Gilani K:
Steatosis and hepatitis C. Gastroenterol Rep (Oxf). 4:24–29.
2016.
|
|
37
|
Demir M and Demir C: Effect of hepatitis C
virus infection on the left ventricular systolic and diastolic
functions. South Med J. 104:543–546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nam YJ, Kim SJ, Shin WC, Lee JH, Choi WC,
Kim KY and Han TH: Gastric pH and Helicobacter pylori infection in
patients with liver cirrhosis. Korean J Hepatol. 10:216–222.
2004.In Korean. PubMed/NCBI
|
|
39
|
Saunders J, Cargill J and Wormsley K:
Gastric secretion of acid in patients with pancreatic disease.
Digestion. 17:365–369. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Carrière F, Grandval P, Renou C, Palomba
A, Priéri F, Giallo J, Henniges F, Sander-Struckmeier S and Laugier
R: Quantitative study of digestive enzyme secretion and
gastrointestinal lipolysis in chronic pancreatitis. Clin
Gastroenterol Hepatol. 3:28–38. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Masini M, Campani D, Boggi U, Menicagli M,
Funel N, Pollera M, Lupi R, Del Guerra S, Bugliani M, Torri S, et
al: Hepatitis C virus infection and human pancreatic beta-cell
dysfunction. Diabetes Care. 28:940–941. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Katakura Y, Yotsuyanagi H, Hashizume K,
Okuse C, Okuse N, Nishikawa K, Suzuki M, Iino S and Itoh F:
Pancreatic involvement in chronic viral hepatitis. World J
Gastroenterol. 11:3508–3513. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Haffar S, Bazerbachi F, Garg S, Lake JR
and Freeman ML: Frequency and prognosis of acute pancreatitis
associated with acute hepatitis E: A systematic review.
Pancreatology. 15:321–326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vaghefi M: Nucleoside Triphosphates and
Their Analogs: Chemistry, Biotechnology, and Biological
Applications. CRC press; London: 2016
|
|
45
|
Lan KH, Sheu ML, Hwang SJ, Yen SH, Chen
SY, Wu JC, Wang YJ, Kato N, Omata M, Chang FY, et al: HCV NS5A
interacts with p53 and inhibits p53-mediated apoptosis. Oncogene.
21:4801–4811. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Niyogi S: Increasing antimicrobial
resistance - an emerging problem in the treatment of shigellosis.
Clin Microbiol Infect. 13:1141–1143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Baer JT, Vugia DJ, Reingold AL, Aragon T,
Angulo FJ and Bradford WZ: HIV infection as a risk factor for
shigellosis. Emerg Infect Dis. 5:820–823. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Aragón TJ, Vugia DJ, Shallow S, Samuel MC,
Reingold A, Angulo FJ and Bradford WZ: Casecontrol study of
shigellosis in San Francisco: the role of sexual transmission and
HIV infection. Clin Infect Dis. 44:327–334. 2007. View Article : Google Scholar
|
|
49
|
Monaco S, Ferrari S, Gajofatto A, Zanusso
G and Mariotto S: HCV-related nervous system disorders. Clin Dev
Immunol. 2012:2361482012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nimnual A and Bar-Sagi D: The two hats of
SOS. Sci STKE. 2002:pe362002.PubMed/NCBI
|
|
51
|
Cutler RE Jr, Stephens RM, Saracino MR and
Morrison DK: Autoregulation of the Raf-1 serine/threonine kinase.
Proc Natl Acad Sci USA. 95:9214–9219. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Giambartolomei S, Covone F, Levrero M and
Balsano C: Sustained activation of the Raf/MEK/Erk pathway in
response to EGF in stable cell lines expressing the hepatitis C
virus (HCV) core protein. Oncogene. 20:2606–2610. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang Q, Gong R, Qu J, Zhou Y, Liu W, Chen
M, Liu Y, Zhu Y and Wu J: Activation of the Ras/Raf/MEK pathway
facilitates hepatitis C virus replication via attenuation of the
interferon-JAK-STAT pathway. J Virol. 86:1544–1554. 2012.
View Article : Google Scholar :
|
|
54
|
Paul F, Pellegrini S and Uze G: IFNA2: The
prototypic human alpha interferon. Gene. 567:132–137. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Petta S, Bellia C, Mazzola A, Cabibi D,
Cammà C, Caruso A, Di Marco V, Craxì A and Ciaccio M:
Methylenetetrahydrofolate reductase homozygosis and low-density
lipoproteins in patients with genotype 1 chronic hepatitis C. J
Viral Hepat. 19:465–472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Adamek A1, Kasprzak A, Mikoś H,
Przybyszewska W, Seraszek-Jaros A, Czajka A, Sterzyńska K and
Mozer-Lisewska I: The insulin-like growth factor-1 and expression
of its binding protein3 in chronic hepatitis C and hepatocellular
carcinoma. Oncol Rep. 30:1337–1345. 2013.PubMed/NCBI
|
|
57
|
Long J, Wang Y, Li M, Tong WM, Jia JD and
Huang J: Correlation of TP53 mutations with HCV positivity in
hepatocarcinogenesis: Identification of a novel TP53 microindel in
hepatocellular carcinoma with HCV infection. Oncol Rep. 30:119–124.
2013. View Article : Google Scholar : PubMed/NCBI
|