Introduction
Stroke is complex neurological diseases in
pathophysiology with high morbidity and is commonly associated with
multiple and long standing impairments (1,2).
Ischemic stroke should be taken seriously in the early stage of
pathological process for study. It is significant to seek for
therapeutic targets for effective protection of cerebral ischemic
impairments (3,4). Extensive research has indicated that
ischemic stroke is followed by highly complex mechanisms such as
the inflammatory (5–7) response, and autophagy (8) and apoptosis (9–11),
leading to ischemic neuronal ischemic and necrotic death, resulting
in cognitive function (12) and
behavior and other impairments (13,14). The mechanism of stroke-induced
cognitive impairment has been extensively studied, while behavioral
disorders caused by cerebral ischemic stroke have been poorly
studied. Neural cell adhesion molecule 2 (Cadm2), which is a
synapse-associated protein, is a cell adhesion factor that has
critical roles in the regulation of neuronal motility and neurite
elongation in developing neurons (15,16). The changes in expression of Cadm2
have been proposed to contribute to nerve growth disorders in
humans (17). However, the role
of Cadm2 in the early stage of stroke remains poorly understood.
Therefore, the extensive identification of a possible basis for
synapse-associated proteins may provide a new strategy for the
treatment of stroke disorders.
Previous studies have revealed that aberrant
regulation of microRNA (miRNA or miR) expression has been linked
with the onset and progression of stroke (18,19). miRNAs are non-coding RNA molecules
that regulate gene expression through binding 3′-untranslated
region (3′-UTR) site in the mRNA to repress translation or induce
mRNA cleavage (20). To data,
miRNAs has been extensively studied and has been shown to serve an
important role in the process of cell growth (21,22), proliferation (23,24), differentiation (25,26), development (27,28) and apoptosis (29,30). It has been reported that miRNAs
are involved in the pathophysiology of ischemic stroke by
regulating their target genes (31). Moreover, miRNAs play important
roles in neurite extension and branching (32,33). However, limited research is
available on the correlation of miRNAs with synapse-associated
proteins, particularly during the early stages of ischemic stroke.
In the present study, the authors used the animal models of middle
cerebral artery occlusion (MCAO) and cell model of neurite
outgrowth to further investigate the role of miRNAs in targeting
synapse-associated proteins, expression involved in behavioral
disorder during the early stage of ischemic stroke. In the present
study, the authors used the animal models of MCAO and cell model of
neurite outgrowth to further investigate the role of miRNAs in
targeting synapse-associated proteins expression involved in
behavioral disorder during the early stage of ischemic stroke.
Materials and methods
Animals
Sprague-Dawley (SD) rats were obtained from the
Department of Animal Experiments, Guangzhou University of Chinese
Medicine. Animals used in all experimental procedures were procured
from the Central Animal House Facility Animal Experimental Center,
Guangzhou Medicine University (Guangzhou, China), under standard
laboratory conditions at a temperature (20–25°C), 12/12 h
light/dark cycle and were provided with a normal pellet diet, water
provided ad libitum. All animals received human care in
accordance with the guideline set by the Care of Experiment Animals
Committee of Guangzhou University of Chinese Medicine. The study
was approved by the Experiment Animals Committee of Guangzhou
University of Chinese Medicine (Guangzhou, China).
PC12 cells culture
PC12 cells were purchased from Shanghai Institute of
Cell Science, Chinese Academy of Sciences (Shanghai, China), and
cultured at 37°C, 5% CO2 in RPMI-1640 medium, 10% fetal
bovine serum (both from Thermo Fisher Scientific, Inc., Waltham,
MA, USA), 1% penicillin-streptomycin (HyClone; GE Healthcare Life
Sciences, Chalfont, UK). The medium was changed every 3 days. For
neurite outgrowth experiments, after placing in reduced serum
medium, PC12 cells were plated on 24- or 6-well cell culture plates
coated with ploy-l-lysine at a density 2×104 or
3×105 cells/well, respectively.
Induction of cerebral ischemia
Male SD rats (200–230 g) were randomized into two
groups (n=10/group): Control group (normal) and model group. Model
group was induced in adult male SD rats (280–300 g) using the
intraluminal filament technique. Rats were anesthetized with an
intraperitoneal injection of 10% chloral hydrate at a dose of 0.33
ml/100 g. A midline neck incision was made; the right common
carotid artery and external carotid artery were isolated. A nylon
filament was inserted into the middle cerebral artery and
maintained for 120 min. Reperfusion was achieved by withdrawing the
suture after 120 min of occlusion. Following the operation, rats
were transferred to a temperature-controlled chamber to maintain
body temperature at 37.5°C. Following 120 min of occlusion, rats
were scored for neurological function. The neurologic scoring
method was based on Longa method which was classified as a
five-point scale (34): Point 0,
no neurologic deficit; point 1, failure to extend the left forepaw
fully; point 2, constant circling to the left; point 3, falling to
the left side; and point 4, no spontaneous walking with a depressed
level of consciousness. After neurological evaluation of MCAO rats,
brains were removed and placed on ice. Seven brains were removed
and fixed in 10% buffered formalin phosphate for 24 to 48 h for
paraffin embedding. Serial coronal frontal cortex and cortex tissue
sections (5 µm) were cut and every tenth section was
systematically assigned to a series of sections, yielding a total
of 10 series. One series of sections was saved for
immunohistochemical analysis. For reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analysis, six ipsilateral cortices and cortex tissue
were rapidly removed and placed on ice. The cortices and cortex
tissue were stored at −80°C for analysis.
microRNA microarray assay
Total RNA was extracted from cortex tissues using
TRIzol reagent according to manufacturer's instructions (Life
Technologies, Carlsbad, CA, USA). The RNA was subsequently
fluorescently labeled with cy3 by ULS notation, and hybridized to
CustomArray™ microarray following the manufacture's instructions
(Guangzhou RiboBio Co., Ltd., Guangzhou, China). In order to
decrease the specific hybridization background, the microarray was
washed and the imaging solution was taken to the semiconductor
microarray. Data were analyzed by MeV (Multi Experiment Viewer
4.9.0; Springer, Boston, MA, USA).
RT-qPCR
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
miR-125a level was analyzed using all-in-one miRNAs RT-qPCR
detection kit (GeneCopoeia, Rockville, MD, USA) according to the
manufacturer's instructions. The reaction conditions of 95°C for 10
sec, 60°C for 20 sec and 72°C for 15 sec, for a total of 40 cycles.
Finally, the relative miR-125a level was normalized to the
endogenous control U6 expression for each sample in triplicate and
was calculated by the 2−ΔΔCt method, as previously
described (35). The primers used
were fluorescent quantitative rno-miR-125a-3p PCR primers,
5′-GACGACAGGTGAGGTTCTTGG-3′ (Invitrogen; Thermo Fisher Scientific,
Inc.).
Transfection of PC12 cells with miR-125a
mimics, inhibitor, miR-125a mimic negative control (mimic NC, m-NC)
and miR-125a inhibitor negative control (inhibitor NC, i-NC)
PC12 cells were plated at 3×105
cells/well in 6-well plates 1 day before transfection. Transfected
Lipofectamine 2000 (Thermo Fisher Scientific, Inc.) 2.5
µl/ml with 50 nM miR-125a mimics or 100 nM inhibitor for 6 h
according to the instructions of Lipofectamine 2000. Following the
terminal transfection for 24 h, the cells were collected with 0.25%
trypsin digestion and lysed with radioimmunoprecipitation assay
buffer (Thermo Fisher Scientific, Inc.) for western blot
analysis.
Immunofluorescence assay
PC12 cells were plated at 2×104
cells/well in 24-well plates 1 day before transfection, then
divided into 4 groups: i), control group; ii), model group
(free-serum incubation 12 h); iii), miR-125a mimic group:
transfected miR-125a mimic followed by free-serum incubation for 12
h; iv), miR-125a inhibitor group: transfected miR-125a inhibitor
followed by free-serum incubation for 12 h. PC12 cells or tissue
sections were incubated with an anti-rabbit Cadm2 antibody (1:200,
Ab64873; Abcam, Cambridge, UK) overnight in 5% BSA (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) at 4°C. After thorough washing,
cells were incubated with a conjugated anti-rabbit secondary
antibody (1:500, Alexa Fluor 488, Ab150077; Abcam) for 30 min at
room temperature. Cells were examined under laser confocal (Carl
Zeiss AG, Jena, Germany) and analyzed by Image Pro Plus software
6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Measurement of neurite outgrowth
PC12 cells were inoculated into the wells at the
initial density of 2×104 cells/well. PC12 cells were
transfected with miR-125a mimics, miR-125a inhibitor, miR-125a
mimics or inhibitor negative control as immunofluorescence assay
described into five groups: i) miR-125a mimics group; ii) miR-125a
mimics negative control group (miR-125a mimics NC group); iii)
miR-125a inhibitor group; iv) miR-125a inhibitor negative control
group (miR-125a inhibitor NC group); v) the control group with
normal cultured. For neurite length assessment, the cells were
observed under a laser confocal microscope; the 20 longest
protrusions in each experimental group were measured and quantified
from six independent fields of vision (three experiments per
condition). The following indicators were used to observe the
effects of miR-125a on cell neurite outgrowth: i) spindle cell
rate, the total number of spindle cells/total number of cells,
observation of cell body shape; ii) cell differentiation rate D1,
the number of cell protrusions/total number of cells; iii) the rate
of cell differentiation D2, the number of cells with neurite
outgrowth >2 cell body lengths/the total number of cells; iv)
average protrusion length of cell, the length of the longest
protrusions/the number of samples. Neurite length was measured and
quantified using the ImageJ 1.4 (National Institutes of Health,
Bethesda, MD, USA) and GraphPad Prism 5 (GraphPad Software, Inc.,
La Jolla, CA, USA) programs.
Western blotting
Briefly, proteins extracted by
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc.) from cultured PC12 cells or tissue followed by 5–10 min
boiling and centrifugation at 12,000 × g for 10–15 min at 4°C to
obtain the supernatant. Protein concentration was determined with a
BCA protein assay kit (Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. Equal amounts of lysate protein
(20 µg/lane) were subjected to 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to nitrocellulose membranes (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) after electrophoresis. At room
temperature, the membranes were blocked with 5% bovine serum
albumin (BSA) in PBST buffer (PBS with 0.01% Tween-20, pH 7.4) and
incubated overnight at 4°C with antibody of Cadm2 (Ab-64873,
dilution, 1:500; Abcam), β-actin (Bs-0061R, dilution, 1:2,000;
Bioss, Beijing, China), in PBST containing 1% BSA. Immunoreactivity
was detected by sequential incubation with horseradish
peroxidase-conjugated secondary antibodies donkey anti-rabbit
immunoglobulin IgG (cat. no. Ab-6721, dilution, 1:10,000; Abcam)
for 2 h at room temperature, and detected by ECL western blot
detection reagent (Bio-Rad Laboratories, Inc.). Densitometry
quantitation was used to analyze with ImageJ 1.4 Software (National
Institutes of Health).
Co-transfection of the luciferase
reporter carrying Cadm2- wild-type (WT) 3′-UTR mutant type (MUT),
Cadm2-MUT-3′-UTR (WT)
Plates (24-well) were plated at a density of
2×104 cells/well. PC12 cells were transfected using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
with Cadm2-WT 3′-UTR, Cadm2-MUT 3′-UTR plasmid and miR-125a mimic,
inhibitor, mimic NC and inhibitor NC for 6 h in culture plates. In
the cloning vector of psiCHECK2-Cadm2, miRNA target sites were
inserted after the Renilla luciferase (hRluc) region,
Firefly luciferase (luc) was taken as internal reference. Cells
treated with mimic NC, and inhibitor NC served as negative
controls. Each value from the wells was collected with the dual
luciferase reporter assay system to quantify luminescent
signal.
Statistical analysis
Data were reported as mean ± standard deviation of
these independent experiments. Statistical analyses were performed
using GraphPad Prism version 5.0 between two groups (GraphPad
Software, Inc.). Data were used with test analysis and one-way
analysis of variance and P<0.05 was considered to indicate a
statistically significant difference.
Results
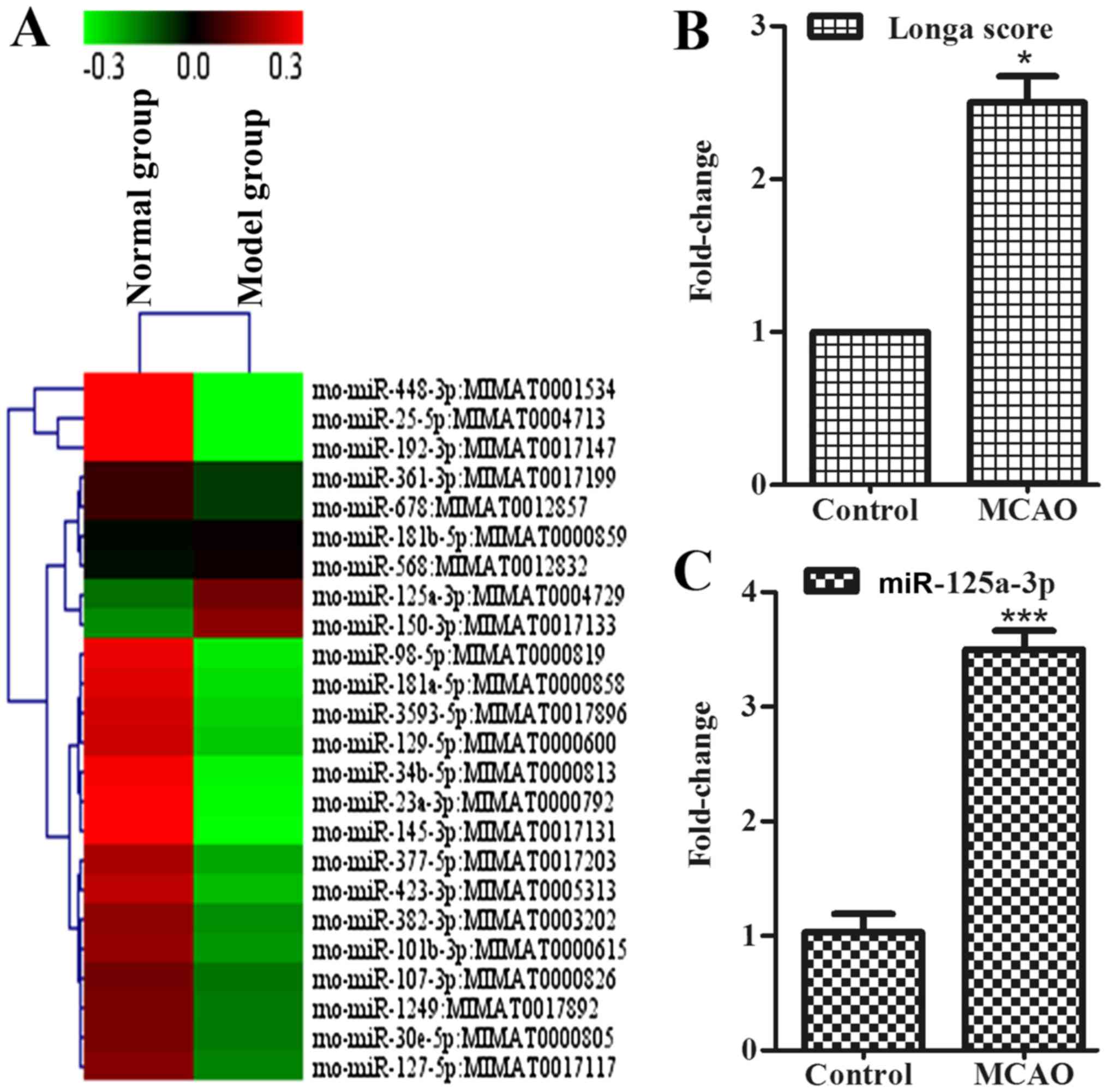
Score of neurofunction
To explore whether the mice with MCAO presented
early symptoms of behavioral disorder, the authors used Longa
method. The present study demonstrated that the MCAO rats showed
these symptoms of behavioral disorder, as assessed by the Longa
score (Fig. 1B).
miR-125a expression is increased in the
cortex of ischemic rats
To determine whether miRNAs exist in the cortex to
connect with neurite outgrowth-associated proteins during the
development of stroke, the authors used a miRNA microarray assay to
detect the association of mRNAs with diseases, such as stroke. Data
were analyzed and illustrated with cluster diagram (Fig. 1A). Results showed that the
expression of miR-125a in the model group was increased in contrast
to the control group, and, considering the acquisition error, the
authors also took miR-125a into the following study. RT-qPCR
indicated that, compared with the control group, the expression of
miR-125a in the cortex of the model group was also higher (Fig. 1C). Moreover, miR-125a may have
contact directly with Cadm2, which has the function of affecting
neurite outgrowth. In vitro, it is of great significance to
the further study of miR-125a.
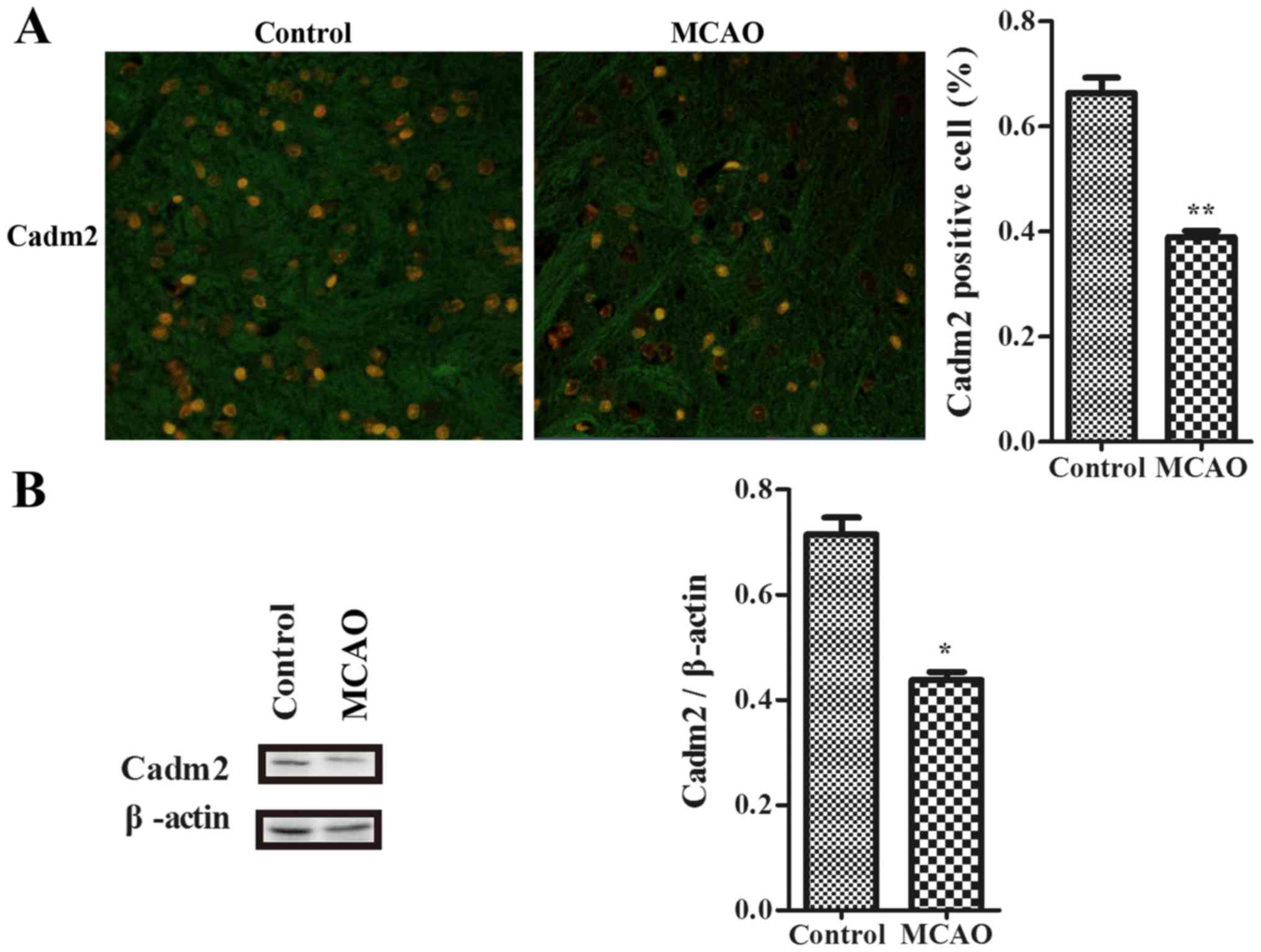
Cadm2 expression is decreased in the
cortex of ischemic rats
To determine whether Cadm2 exists in the cortex to
connect with neurite outgrowth-associated proteins in the
development of stroke, the authors used an immunofluorescence assay
to examine with association with stroke. Data were detected with
laser confocal microscope (Fig.
2A). Results indicated that the expression of Cadm2 in the
model group was decreased when contrasted to the control group. By
using western blotting, compared with the control group, the
expression of Cadm2 of the model group was also lower (Fig. 2B).
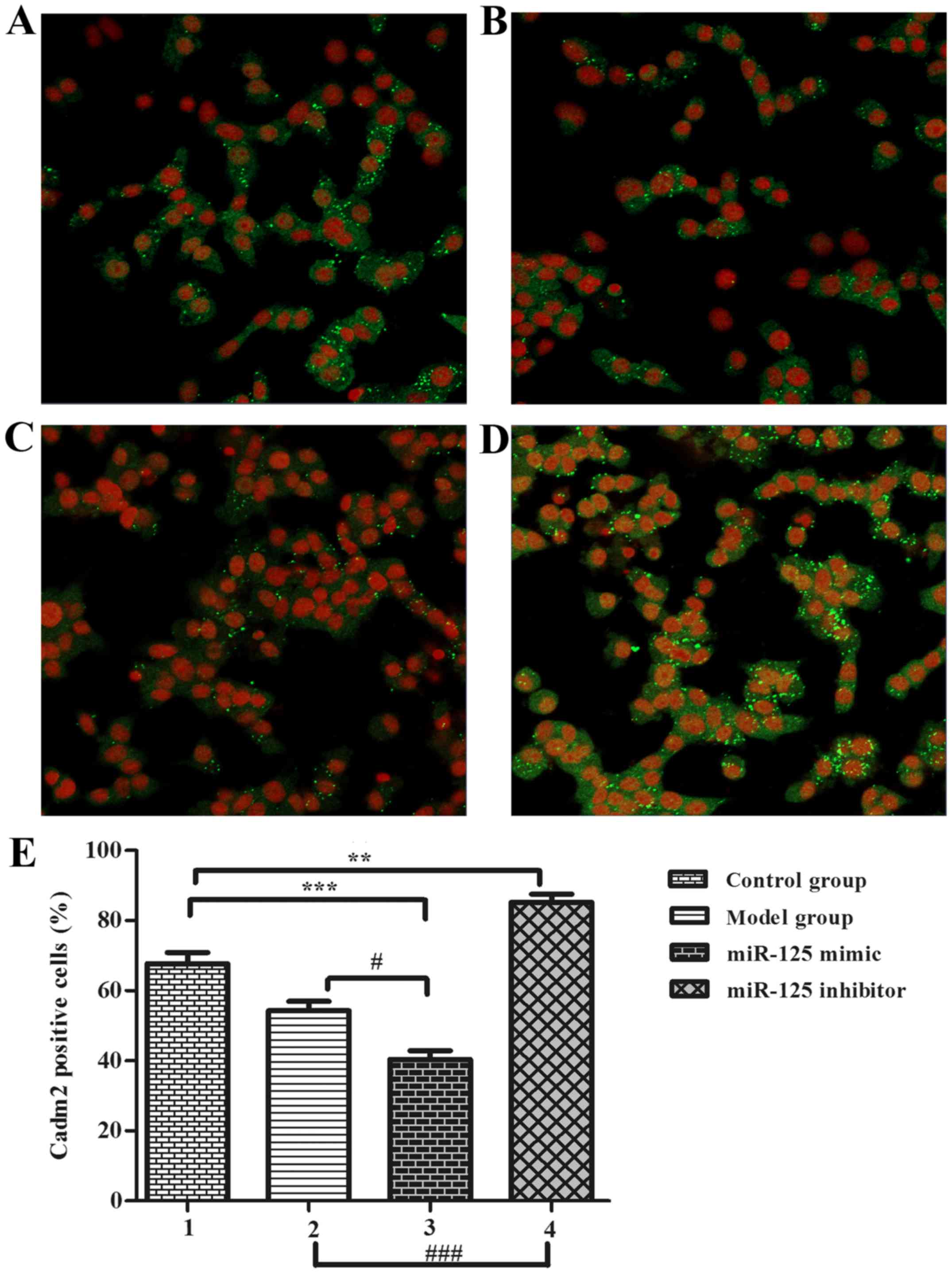
miR-125a inhibits Cadm2 expression
In previous studies, the authors identified that the
neurite outgrowth-associated proteins may be the target protein of
miR-125a. For confirming the target protein using PC12 cells in
vitro, cells were transfected with miR-125a mimics, miR-125a
inhibitor compared with control group and no serum group, PC12
cells were used as a serum-free cell model (12 h) for observing
neurite outgrowth, results were observed in an immunofluorescence
assay (Fig. 3). Results showed
that the ratio of positive cells in the model group (Fig. 3B) was lower than the control group
(Fig. 3A). The mimic group
(Fig. 3C) positive cell number
was lower than the inhibitor group (Fig. 3D) and the control group. These
results indicate that serum-free miR-125a could inhibited neurite
outgrowth in stroke development process (Fig. 3E), thus, PC12 cells could be
suitable for the authors' further study to prove the relationship
between Cadm2 and miR-125a.
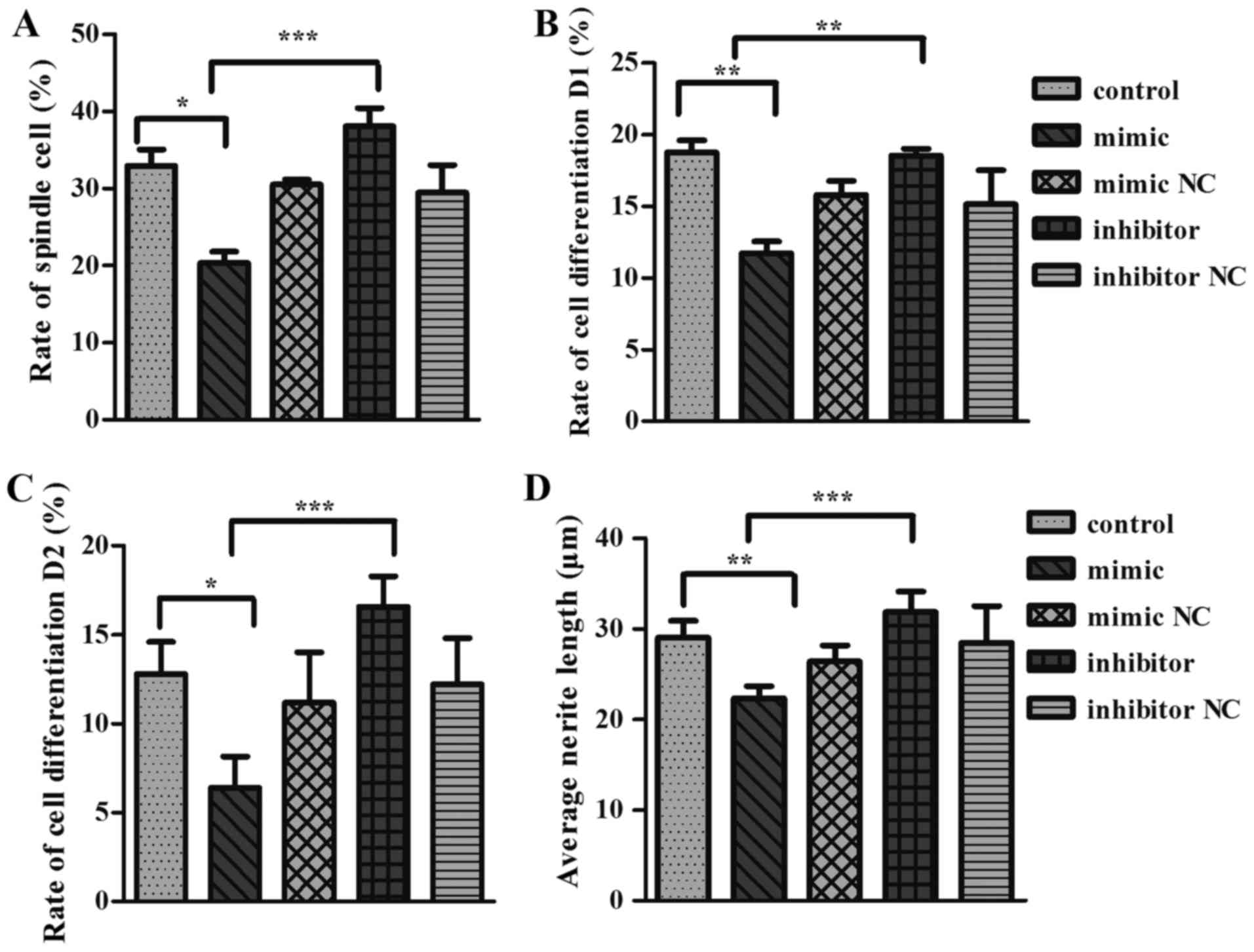
miR-125a inhibits neurite outgrowth in
PC12 cells
To further validate the role of Cadm2 in miR-125a
mediated neurite outgrowth, PC12 cells were transfected with
miR-125a mimics, inhibitor, mimic NC or inhibitor NC and the
neurite length was reported (Fig.
4). The results demonstrated that cells transfected with the
miR-125a mimic had a significantly decreased neurite extension
compared than miR-125a mimic NC and control groups in terms of
spindle cell rate (Fig. 4A), cell
differentiation rate D1 (Fig.
4B), the rate of cell differentiation D2 (Fig. 4C) and average protrusion length of
cell (Fig. 4D). PC12 cells
transfected with miR-125a inhibitor, which suppresses mature
miR-125a expression (Fig. 4),
exhibited a significant increase in neurite outgrowth.
miR-125a can specifically reduce the
protein expression of Cadm2
To investigate whether miR-125a inhibits the
expression of target protein Cadm2, the authors transfected cells
with miR-125a mimic or inhibitor, or miR-125a mimic NC or inhibitor
NC into PC12 cells. It was determined that Cadm2 with the function
of neurite outgrowth was a potential target of miR-125a. Compared
with the control group and mimic NC, the expression of Cadm2
protein was reduced in mimic group by western blotting and the
results were analyzed with ImageJ (Fig. 5B). The expression of Cadm2 protein
was increased in the inhibitor group, contrasted to the control
group inhibitor NC. To identify whether miR-125a targeted the
expression of Cadm2, the authors co-transfected the cells with
miR-125a mimic, inhibitor or miR-125a mimic NC, inhibitor NC with
Cadm2-WT 3′-UTR or Cadm2-MUT 3′-UTR into PC12 cells. Results
indicated that the WT+miR-125a mimic significantly reduced the
luciferase activity, compared with MUT+miR-125a mimic and mimic NC.
The WT+miR-125a inhibitor increased the luciferase activity,
compared with MUT+miR-125a inhibitor and inhibitor NC results
reported that miR-125a specifically regulated the expression of
Cadm2, inhibiting neurite outgrowth (Fig. 5C).
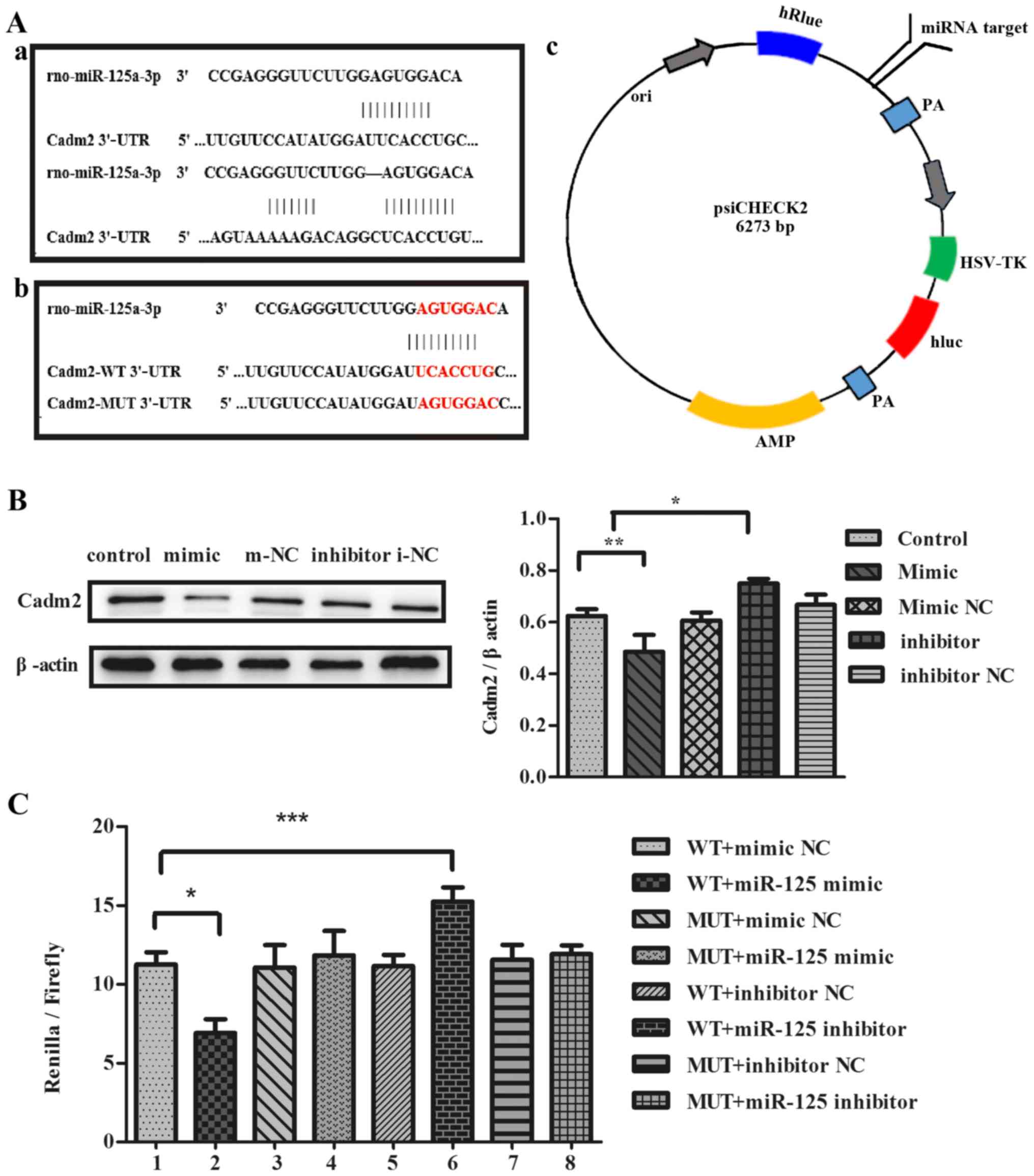 | Figure 5miR-125a regulates the expression of
Cadm2. (A) (a) The target sites of miR-125a in the Cadm2 3′-UTR
were predicted by bioinformatics software, TargetScan and
miRNA.org. (b) The target sites of miR-125a in
Cadm2-WT 3′-UTR and Cadm2-MUT 3′-UTR. (c) The construction profile
of the psiCHECK2-Cadm2 vector is presented in the diagram that
contained the miR-125a-3p target sites in Cadm2 3′-UTR. (B)
Expression of target protein Cadm2 in the PC12 cells were analyzed
with the western blot assay, and the bands were quantified with
ImageJ software. One-way analysis of variance was used to analyze
the variances. By western blotting, the expression of the Cadm2
protein was efficiently decreased by miR-125a mimics, compared with
the control group, and may be increased obviously when treated with
miR-125a inhibitor. (C) The miR-125a mimic, inhibitor, mimic NC or
inhibitor NC were co-transfected with Cadm2-WT 3′-UTR or Cadm2-MUT
3′-UTR into PC12 cells. miR-125a targeted the expression of Cadm2.
One-way analysis of variance test was used to analyze the variances
between groups. *P<0.05, **P<0.01 and
***P<0.001 as indicated. miR, microRNA; UTR,
untranslated region; WT, wild-type; NC, negative control; Cadm2,
cell adhesion molecule 2. |
Discussion
Stoke is the result of ischemic brain and can lead
to neuronal damage (36), and
there is no effective cure for this condition. Ischemic brain
injury to neuronal cells is mediated by multiple mechanisms
(37). One of the processes that
may play a key role in the delayed progression of the neurite
outgrowth is initiated by the expression of miRNAs (38). However, there are few studies on
the roles of miRNAs in the behavioral disorder associated with the
early stage of stroke. The present study used a rat model of MCAO
and PC12 cells (MCAO can mimic many of the salient features of the
early stages of stroke) to further investigate the roles of miRNAs
in neurite outgrowth associated with behavior. The three findings
of our study were: i) the rat model of MCAO displayed significant
behavioral deficits; ii) miR-125a expression was increased
significantly in the rat model of MCAO, but Cadm2 expression was
decreased; iii) Cadm2 (mRNA) is targeted by miR-125a. The results
indicated that the disruption of neurite outgrowth via the miR-125a
targeting of Cadm2 may play a role in the behavioral deficits
associated with stroke and thus miR-125a may be a key target for
drug development for the prevention of stroke.
An important finding of the present study was that
miR-125a expression was increased significantly in the rats with
MCAO. The microarray data and RT-qPCR data identified miR-125a to
be upregulated in the rats with MCAO; these results are consistent
with the hypothesis that some miRNAs can be differentially
expressed in a stroke-affected brain. Previous studies have
indicated that miR-125a is expressed or enriched in multiple
myeloma (39,40), psychiatric disorders (41) and cerebrovascular diseases
(42). miR-125a has been shown to
play a role in cancer development and metastasis, and to be
involved in neurodegenerative diseases (43). However, little is understood about
the roles of miR-125a in neurite outgrowth in stroke-afflicted
rats. In our study, it was found that the upregulation of miR-125a
was concomitant with the significant behavioral deficits displayed
by the rats with MCAO. Therefore, the authors put forward the
hypothesis that miR-125a may be associated with neurite
outgrowth.
Previous studies have confirmed that several mRNAs
are the direct targets of miR-125a, such as Gap43 and Cadm2. Among
these target genes, Cadm2 plays an important role in neuronal
growth, which regulates axon elongation and branching (44,45). In the present study, the authors
observed that expression of Cadm2 was decreased after
overexpression of miR-125a and increased following miR-125a
inhibition, paralleling with the neurite outgrowth. Bioinformatics
analysis predicted that Cadm2 (mRNA) is targeted by miR-125a.
Changes in expression of neural Cadm2 have been proposed to
contribute to neurodevelopmental disorders in humans. The role of
Cadm2 in neuronal differentiation remains, however, poorly
understood (17). Therefore, the
authors focused on the miR-125a to test the hypothesis. To confirm
the hypothesis in vitro, a cortex neuronal PC12 cell line
was used, which is an established cell model. PC12 cells were
transfected with miR-125a mimic or inhibitor, and then contrasted
with the control group. The results of immunocytochemistry showed
that mir-125 inhibited the expression of Cadm2 and that Cadm2 level
was decreased in PC12 cells transfected with miR-125a mimics in
western blotting and a luciferase reporter assay. These results
suggested that Cadm2 is targeted by miR-125.
There are several important clinical implications in
these results. First, stroke is one of the primary causes for
morbidity and long-term disability worldwide. There is an urgent
demand for sensitive and specific biomarkers for detection. In the
present study, the authors found that miR-125a was increased
significantly in the cortex of model of MCAO. Thus, the miR-125a
may be potential biomarkers for the early diagnosis of stroke.
Secondly, currently there are no therapies to protect the
progression of behavioral disturbances. The present studies have
shown that miR-125a inhibited the neurite outgrowth in PC12 cells.
Thirdly, the results support a new possible therapeutic target for
stroke based on regulation of the miR-125-neurite outgrowth pathway
on synaptic proteins. Inhibition of miR-125 appears a novel
therapeutic option for behavioral disturbances in stroke.
In conclusion, the observations suggest that the
miR-125a is involved in behavioral disturbances in stroke and that
inhibition of neurite outgrowth is due to the reduction of Cadm2
expression. The study suggested that miR-125a mediates neurite
outgrowth by suppression of Cadm2, thereby inhibiting neurite
outgrowth pathways in PC12 cells. This further indicated that
altering the miR-125a-neurite outgrowth pathway may represent a
targeted and mechanism-based therapeutic strategy against neurite
outgrowth following stroke.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81673770 and
81373525). Guangdong Provincial Natural Science Foundation of China
(grant no. 2014A030313409).
References
|
1
|
Zhai WW, Sun L, Yu ZQ and Chen G:
Hyperbaric oxygen therapy in experimental and clinical stroke. Med
Gas Res. 6:111–118. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boehme AK, Esenwa C and Elkind MS: Stroke
risk factors, genetics, and prevention. Circ Res. 120:472–495.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Isabel C, Calvet D and Mas JL: Stroke
prevention. Presse Med. 45:e457–e471. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Proietti M, Mairesse GH, Goethals P,
Scavee C, Vijgen J, Blankoff I, Vandekerckhove Y and Lip GY;
Belgian Heart Rhythm Week Investigators: Cerebrovascular disease,
associated risk factors and antithrombotic therapy in a population
screening cohort: Insights from the Belgian Heart Rhythm Week
programme. Eur J Prev Cardiol. 24:328–334. 2017. View Article : Google Scholar
|
|
5
|
Perez-Alvarez MJ and Wandosell F: Stroke
and neuroinflamation: role of sexual hormones. Curr Pharm Des.
22:1334–1349. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gokce EC, Kahveci R, Gokce A, Sargon MF,
Kisa U, Aksoy N, Cemil B and Erdogan B: Curcumin attenuates
inflammation, oxidative stress, and ultrastructural damage induced
by spinal cord ischemia-reperfusion injury in rats. J Stroke
Cerebrovasc Dis. 25:1196–1207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gao L, Dong Q, Song Z, Shen F, Shi J and
Li Y: NLRP3 inflammasome: A promising target in ischemic stroke.
Inflamm Res. 66:17–24. 2017. View Article : Google Scholar
|
|
8
|
Wang J, Han D, Sun M and Feng J: Cerebral
ischemic post conditioning induces autophagy inhibition and a HMGB1
secretion attenuation feedback loop to protect against ischemia
reperfusion injury in an oxygen glucose deprivation cellular model.
Mol Med Rep. 14:4162–4172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tiwari HS, Misra UK, Kalita J, Mishra A
and Shukla S: Oxidative stress and glutamate excitotoxicity
contribute to apoptosis in cerebral venous sinus thrombosis.
Neurochem Int. 100:91–96. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang X, Li H and Ding S: Pre-B-cell
colony-enhancing factor protects against apoptotic neuronal death
and mitochondrial damage in ischemia. Sci Rep. 6:324162016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu HJ, Wu C, Niu HJ, Wang K, Mo LJ, Shao
AW, Dixon BJ, Zhang JM, Yang SX and Wang YR: Neuroprotective
Mechanisms of Melatonin in Hemorrhagic Stroke. Cell Mol Neurobiol.
37:1173–1185. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ballard C, Stephens S, Kenny R, Kalaria R,
Tovee M and O' Brien J: Profile of neuropsychological deficits in
older stroke survivors without dementia. Dement Geriatr Cogn
Disord. 16:52–56. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Buesing C, Fisch G, O' Donnell M, Shahidi
I, Thomas L, Mummidisetty CK, Williams KJ, Takahashi H, Rymer WZ
and Jayaraman A: Effects of a wearable exoskeleton stride
management assist system (SMA®) on spatiotemporal gait
characteristics in individuals after stroke: A randomized
controlled trial. J Neuroeng Rehabil. 12:692015. View Article : Google Scholar
|
|
14
|
O' Hare A: Management of developmental
speech and language disorders. Part 2: Acquired conditions. Arch
Dis Child. 101:278–283. 2016. View Article : Google Scholar
|
|
15
|
Sandig M, Rao Y and Siu CH: The homophilic
binding site of the neural cell adhesion molecule NCAM is directly
involved in promoting neurite outgrowth from cultured neural
retinal cells. J Biol Chem. 269:14841–14848. 1994.PubMed/NCBI
|
|
16
|
Kulahin N and Walmod PS: The neural cell
adhesion molecule NCAM2/OCAM/RNCAM, a close relative to NCAM. Adv
Exp Med Biol. 663:403–420. 2010. View Article : Google Scholar
|
|
17
|
Sheng L, Leshchyns'ka I and Sytnyk V:
Neural cell adhesion molecule 2 promotes the formation of filopodia
and neurite branching by inducing submembrane increases in
Ca2+ levels. J Neurosci. 35:1739–1752. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y and Cai Y: Obtaining human ischemic
stroke gene expression biomarkers from animal models: A
cross-species validation study. Sci Rep. 6:296932016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou J and Zhang J: Identification of
miRNA-21 and miRNA-24 in plasma as potential early stage markers of
acute cerebral infarction. Mol Med Rep. 10:971–976. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brooks SA, Martin E, Smeester L, Grace MR,
Boggess K and Fry RC: miRNAs as common regulators of the
transforming growth factor (TGF)-β pathway in the preeclamptic
placenta and cadmium-treated trophoblasts: Links between the
environment, the epigenome and preeclampsia. Food Chem Toxicol.
98:50–57. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su SH, Wu CH, Chiu YL, Chang SJ, Lo HH,
Liao KH, Tsai CF, Tsai TN, Lin CH, Cheng SM, et al: Dysregulation
of vascular endothelial growth factor receptor-2 by Mmultiple
miRNAs in endothelial colony-forming cells of coronary artery
disease. J Vasc Res. 54:22–32. 2017. View Article : Google Scholar
|
|
23
|
Longqiu Y, Pengcheng L, Xuejie F and Peng
Z: A miRNAs panel promotes the proliferation and invasion of
colorectal cancer cells by targeting GABBR1. Cancer Med.
5:2022–2031. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang W, Cai P, Huo W, Li H, Tang J, Zhu D,
Xie H, Chen P, Hang B, Wang S, et al: Suppressive action of miRNAs
to ARP2/3 complex reduces cell migration and proliferation via RAC
isoforms in Hirschsprung disease. J Cell Mol Med. 20:1266–1275.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Quan L, Wang Y, Liang J, Qiu T, Wang H,
Zhang Y, Zhang Y, Hui Q and Tao K: Screening for genes,
transcription factors and miRNAs associated with the myogenic and
osteogenic differentiation of human adipose tissue-derived stem
cells. Int J Mol Med. 38:1839–1849. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Filip AA, Grenda A, Popek S, Koczkodaj D,
Michalak-Wojnowska M, Budzyński M, Wąsik-Szczepanek E, Zmorzyński
S, Karczmarczyk A and Giannopoulos K: Expression of circulating
miRNAs associated with lymphocyte differentiation and activation in
CLL-another piece in the puzzle. Ann Hematol. 96:33–50. 2017.
View Article : Google Scholar
|
|
27
|
Bao W, Greenwold MJ and Sawyer RH:
Expressed miRNAs target feather related mRNAs involved in cell
signaling, cell adhesion and structure during chicken epidermal
development. Gene. 591:393–402. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martinez-Sanchez A, Rutter GA and
Latreille M: MiRNAs in β-cell development, identity, and disease.
Front Genet. 7:2262017. View Article : Google Scholar
|
|
29
|
Coutinho-Camillo CM, Lourenço SV, de
Araújo Lima L, Kowalski LP and Soares FA: Expression of
apoptosis-regulating miRNAs and target mRNAs in oral squamous cell
carcinoma. Cancer Genet. 208:382–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Venkatadri R, Muni T, Iyer AK, Yakisich JS
and Azad N: Role of apoptosis-related miRNAs in resveratrol-induced
breast cancer cell death. Cell Death Dis. 7:e21042016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Privitera AP, Distefano R, Wefer HA, Ferro
A, Pulvirenti A and Giugno R: OCDB: A database collecting genes,
miRNAs and drugs for obsessive-compulsive disorder. Database
(Oxford). 2015:bav0692015. View Article : Google Scholar
|
|
32
|
Dajas-Bailador F, Bonev B, Garcez P,
Stanley P, Guillemot F and Papalopulu N: microRNA-9 regulates axon
extension and branching by targeting Map1b in mouse cortical
neurons. Nat Neurosci. 15:697–699. 2012. View Article : Google Scholar
|
|
33
|
Lu XC, Zheng JY, Tang LJ, Huang BS, Li K,
Tao Y, Yu W, Zhu RL, Li S and Li LX: MiR-133b Promotes neurite
outgrowth by targeting RhoA expression. Cell Physiol Biochem.
35:246–258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zeng L, Liu J, Wang Y, Wang L, Weng S,
Tang Y, Zheng C, Cheng Q, Chen S and Yang GY: MicroRNA-210 as a
novel blood biomarker in acute cerebral ischemia. Front Biosci
(Elite Ed). 3:1265–1272. 2011.
|
|
36
|
Xu W, Jin W, Zhang X, Chen J and Ren C:
Remote limb preconditioning generates a neuroprotective effect by
modulating the extrinsic apoptotic pathway and TRAIL-receptors
expression. Cell Mol Neurobiol. 37:169–182. 2017. View Article : Google Scholar
|
|
37
|
Chi W, Meng F, Li Y, Li P, Wang G, Cheng
H, Han S and Li J: Impact of microRNA-134 on neural cell survival
against ischemic injury in primary cultured neuronal cells and
mouse brain with ischemic stroke by targeting HSPA12B. Brain Res.
1592:22–33. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mercado AT, Yeh JM, Chin TY, Chen WS,
Chen-Yang YW and Chen CY: The effect of chemically modified
electrospun silica nanofiber on the mRNA and miRNA expression
profile of neural stem cell differentiation. J Biomed Mater Res A.
104:2730–2743. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bi C, Chung TH, Huang G, Zhou J, Yan J,
Ahmann GJ, Fonseca R and Chng WJ: Genome-wide pharmacologic
unmasking identifies tumor suppressive microRNAs in multiple
myeloma. Oncotarget. 6:26508–26518. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Alzrigat M, Parraga AA, Agarwal P,
Zureigat H, Österborg A, Nahi H, Ma A, Jin J, Nilsson K, Öberg F,
et al: EZH2 inhibition in multiple myeloma downregulates myeloma
associated oncogenes and upregulates microRNAs with potential tumor
suppressor functions. Oncotarget. 8:10213–10224. 2017.PubMed/NCBI
|
|
41
|
Camkurt MA, Karababa F, Erdal ME, Bayazıt
H, Kandemir SB, Ay ME, Kandemir H, Ay Öİ, Çiçek E, Selek S, et al:
Investigation of dysregulation of several MicroRNAs in peripheral
blood of schizophrenia patients. Clin Psychopharmacol Neurosci.
14:256–260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dai D, Lu Q, Huang Q, Yang P, Hong B, Xu
Y, Zhao W, Liu J and Li Q: Serum miRNA signature in Moyamoya
disease. PLoS One. 9:e1023822014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dasdag S, Akdag MZ, Erdal ME, Erdal N, Ay
OI, Ay ME, Yilmaz SG, Tasdelen B and Yegin K: Effects of 2.4 GHz
radio-frequency radiation emitted from Wi-Fi equipment on microRNA
expression in brain tissue. Int J Radiat Biol. 91:555–561. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Futerman AH and Banker GA: The economics
of neurite outgrowth - the addition of new membrane to growing
axons. Trends Neurosci. 19:144–149. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Blackmore MG, Moore DL, Smith RP, Goldberg
JL, Bixby JL and Lemmon VP: High content screening of cortical
neurons identifies novel regulators of axon growth. Mol Cell
Neurosci. 44:43–54. 2010. View Article : Google Scholar : PubMed/NCBI
|