Introduction
Various lifestyle and environmental factors have
been associated with an increase in the incidence of male-specific
diseases (1), including orchitis,
which is characterised by the inflammation of one or both testicles
due to local or systemic infection, or non-infectious factors, with
infective orchitis being more common (2). The mammalian testis is composed of
spermatogenic tubules that primarily consist of spermatogenic
epithelium and are separated by stromal layers. The spermatogenic
epithelium consists of Sertoli sustentacular cells and
spermatogenic cells, while the interstitial space between the
seminiferous tubules contains capillaries, capillary lymphatic
vessels, nerves and various types of cells, including Leydig cells,
fibroblasts, giant cells, mast cells, lymphocytes and eosinophils.
Leydig cells are primarily responsible for androgen secretion
(3,4).
The development of inflammatory diseases is closely
associated with the activation of various signaling pathways
including the nuclear factor light-chain-enhancer of activated B
cells (NF-κβ) signaling pathway (5,6),
which is closely associated with the development of testicular
inflammation (7). Phosphorylated
transcription factor p65 (p-p65) is an important marker of
inflammation and NF-κβ signaling pathway activation (6).
Protein degradation is important for maintaining
normal physiological functions and homeostasis (8), as the disruption of this system is a
known cause of several diseases. In higher eukaryotes, the majority
of proteins (80-90%) are degraded by the proteasome (9,10).
The normal function of the proteasome is critical for maintaining
relatively stable intracellular activity. The 11S regulatory
complex, which includes proteasome activator complex subunit 1,
proteasome activator complex subunit 2 and proteasome activator
complex subunit 3 (REGγ), has been demonstrated to enhance the
proteasomal degradation of substrate proteins and to alter the type
and substrate selectivity of enzymes (11). One study has suggested that
inflammatory stimulation significantly increases REGγ expression in
mouse colon epithelial cells and human colon cancer epithelial
cells (12). Conversely,
intestinal inflammation was decreased in a REGγ-deficient mouse
model of inflammatory bowel disease (12). However, the association between
REGγ and testicular inflammation remains unclear.
Therefore, the present study aimed to determine the
role of REGγ in testicular inflammation using mouse and cell
models. It was observed that the knockdown of REGγ resulted in a
decreased level of inflammation in the animal and the cell models.
Furthermore, REGγ may regulate the NF-κB signaling pathway via
NF-κB inhibitor (IkB) ε (IkBε). These data indicate that REGγ may
serve as a potential target in the treatment of testis
inflammation.
Materials and methods
Experimental mice
The 6-week-old C57BL/6 wild-type (WT)
(REGγ+/+) and REGγ-deficient (REGγ−/−) male
mice were obtained from the Minhang Laboratory Animal Center of
East China Normal University (Shanghai, China) and housed under
specific pathogen-free conditions at 21±2°C and 55±10% humidity
under a 12 h light/dark cycle, and fed normal chow with ad
libitum access to water. The C57BL/6 REGγ−/− mice
were originally acquired from Dr John J. Monaco (University of
Cincinnati College of Medicine, Cincinnati, OH, USA) (11-12). A total of 36 REGγ+/+
mice and 24 REGγ−/− mice were used for the current
study.
Cell culture and expression
constructs
Primary Leydig cells were collected from mouse
testes. TM3 cells were purchased from the Cell Bank of Type Culture
Collection Chinese Academy of Sciences (Shanghai, China; cat. no.
GNM24). The TM3 cell line is a mouse epithelial Leydig cell line.
Primary Leydig cells and the TM3 cell line were grown in Dulbecco's
modified Eagle's medium/F-12 nutrient mixture (DMEM/F-12)
supplemented with 5% fetal bovine serum, 2.5% horse bovine serum
(all Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
L-glutamine (150 mg/l), NaHCO3 (1.5 g/l), penicillin
(100 U/ml) and streptomycin (100 µg/ml). Cells were cultured
in a 37°C incubator with 5% CO2 and 95% air.
Animal and cell inflammation models
Based on preliminary experiments, 12
REGγ+/+ mice were selected and 6 of them were injected
intraperitoneally with LPS (20 mg/kg body weight) dissolved in
double distilled (dd) water for 6 h. As a control, 6 mice were
injected intraperitoneally with equal volumes of dd water for 6
h.
The optimal conditions for creating an inflammatory
cell model were explored. Concentration (0, 1, 5 or 10 mg/ml) and
time gradients (0, 10, 20, 40 or 60 min) were established to
determine the optimal model conditions. TM3 cells were treated for
10 min with LPS at concentrations of 0, 1, 5 or 10 mg/ml. The
optimal concentration of LPS was then selected to treat the cells
for different times (0, 10, 20, 40 or 60 min) in order to select
the optimal treatment time.
Primary Leydig cells
Upon LPS treatment, mice were sacrificed by
CO2 anesthesia (SMQ-II; Tianhuan Technology, Co., Ltd.,
Shanghai, China; final CO2 concentration, 80-100%). The
displacement rates were between 10-30% of the chamber volume per
minute (cv/min), and death was confirmed by observation of loss of
respiration, heart beat and reflexes. Then, cervical dislocation
was performed as a secondary physical method of sacrifice.
The testes were then collected from the mice in a
sterile hood. Sterile tweezers were used to remove the testicular
capsule. The tissue was digested in 0.25% collagenase at 37°C for
10 min (1 ml per testis). Next, the tissue suspension was filtered
with a 40 µm filter. Following centrifugation at 200 x g and
4°C for 5 min, the cell suspension was placed in culture dishes and
cultured in an incubator for 4 h. The adherent cells that were
attached to the dishes were considered to be primary Leydig cells.
The purity of the Leydig cells, as assessed by immunocytochemical
staining for 3β-hydroxysteroid dehydrogenase.
For immunocytochemical staining, after the cells
adhered to the slip, the glass slips were washed three times with
1X PBS, fixed with 4% paraformaldehyde at room temperature for 20
min and again washed three times with PBS. Then the glass slips
were put in 0.5% Triton X-100 (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at room temperature for 20 min and washed with
PBS. Then, the slips were put in 3% H2O2 at
room temperature for 15 min, washed with PBS and blocked with 5%
bovine serum albumin (BSA; Sigma-Aldrich; Merck KGaA) at room
temperature for 1 h. The glass slips were then treated with
anti-3β-hydroxysteroid dehydrogenase primary antibodies (cat. no.
sc-515120; 1:50; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
overnight at 4°C, washed with PBS, treated with horseradish
peroxidase-conjugated secondary antibodies (cat. no. K5007; 1:200;
Dako; Agilent Technologies, Inc., Santa Clara, CA, USA) at room
temperature for 1 h and again washed with PBS. Next, the slips were
washed with PBS, stained with 0.5 ug/ml DAPI for 3 min at room
temperature and washed with PBS. Then, the glass slips were
observed by fluorescence microscopy at a magnification of ×600. In
each field of vision, the total number of cells and positive cell
were counted, positive cells accounted for >90% of total
cells.
Knockdown of REGγ
Lipofectamine® 2000 reagent (Thermo
Fisher Scientific, Inc.) was used to knock down REGγ and IkBε
expression in TM3 cells using small hairpin (sh)RNA: shREGγ and
shIkBε. TM3 control (shN) and REGγ-knockdown (shR) cells were
generated by retroviral shRNA plasmids specific for REGγ or control
vectors, respectively, from OriGene Technologies, Inc. (Rockville,
MD, USA). The plasmids were provided by Shanghai GenePharma Co.,
Ltd. (Shanghai, China). A total of 1 µg/µl pshRNAIkBε
or pshRNAREGγ1 vectors were incubated with TM3 cells for 12 h to
obtain shN or shR cells, respectively. Then, the expression levels
of REGγ in the shN and shR groups were detected by western blot
analysis.
Western blot analysis
Following treatment, Leydig and TM3 cells were lysed
in 1X loading buffer (5X loading buffer: 250 mM Tris-HCl pH 6.8,
10% SDS, 10% bromphenol blue, 50% glycerin and 5% β-5
Mercaptoethanol) for total protein collection. Protein
concentrations were determined using the bicinchoninic acid method.
The buffer, including all the extracted proteins, was then heated
at 100°C for 30 min. Protein samples (50 µg/lane) were
subsequently separated using SDS-PAGE on 11% gels and transferred
onto nitrocellulose membranes. Upon blocking with 7% BSA dissolve
in PBS at room temperature for 1 h, the membranes were incubated
with primary antibodies (1:1,000) at 4°C for 12 h and then with
IRDye® 800CW-conjugated donkey anti-rabbit secondary
antibodies (cat. no. 925-32213; 1:5,000; LI-COR Biosciences,
Lincoln, NE, USA). β-actin (1:5,000) was used as a loading control.
Finally, the membranes were scanned using an Odyssey imaging system
(LI-COR Biosciences). Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc., Rockville, MD, USA) was used for the
densitometric analysis. The following antibodies were employed:
Anti-IkBε (cat. no. sc-7155; Santa Cruz Biotechnology, Inc.),
anti-IkBα (cat. no. 4812), anti-IkBβ (cat. no. 94101; both Cell
Signaling Technology, Inc., Danvers, MA, USA), anti-β-actin (cat.
no. a2228; Sigma-Aldrich; Merck KGaA) and anti-p-p65 (cat. no.
ab16502; Abcam, Cambridge, UK).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol®
reagent (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol, and reverse transcribed into complementary
DNA using the Mx3005P qPCR system (Agilent Technologies, Inc.)
under the following thermocycling conditions: Denaturation at 94°C
for 5 min, followed by 30 cycles of 94°C for 30 sec, 55-58°C for 30
sec, 72°C for 30 sec and 70°C for 1 min, and a final step at 4°C
for 10 min. Each PCR mixture contained 10 µl SYBR-Green
Premix Ex Taq polymerase (Takara Biotechnology Co., Ltd., Dalian,
China). For each sample, the mRNA expression levels of the genes of
interest were normalized to those of GAPDH. The 2−ΔΔCq
method was used for quantification (13). The primers used for the qPCR were
as follows: Tumor necrosis factor-α (TNF-α) forward (F), 5′-GAC GTG
GAA CTG GCA GAA GAC-3′ and reverse (R), 5′-TCG CAC AAG CAG GAA TGA
GA-3′; interleukin (IL)-6 F, 5′-CCA CGG CCT TCC CTA CTT C-3′ and
IL-6 R, 5′-CTG TTG GGA GTG GTA TCC TCT GT-3′; IL-1β F, 5′-GAT GAT
AAC CTG CTG GTG TGT GA-3′ and R, 5′-GTT GTT CAT CTC GGA GCC TGT
AG-3′; and GAPDH F, 5′-AGG TCG GTG TGA ACG GAT TTG-3′ and R, 5′-GGG
GTC GTT GAT GGC AAC A-3′.
Immunohistochemistry
Tissues were fixed with Bouin's solution
(Sigma-Aldrich; Merck KGaA) overnight at room temperature. The
tissues were transferred to 70% ethanol for 48 h prior to treatment
with a solution of lithium carbonate (1.25%). Then the samples were
dehydrated through a graded series of ethanol and embedded in
paraffin. The inflamed testes were sectioned into 5-µm-thick
slices, and stained with 5% hematoxylin at room temperature for 10
min and 0.5% eosin for at room temperature 3 min to observe
structural changes. Other slices were subjected to
immunohistochemical (IHC) analysis to observe specific expression
patterns of REGγ, IkBε and p-p65 in testicular tissue upon LPS
treatment. IHC was performed using the Histostain-Plus (DAB) kit
(PH0723; Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The slices were stained with
anti-REGγ, anti-IkBε and anti-p-p65 antibodies (dilution, 1:100)
overnight at 4°C and observed under a light microscope (Eclipse
E100; Nikon Corporation, Tokyo, Japan) to evaluate histological
changes. The following antibodies were employed: Anti-IkBε (cat.
no. sc-7155; Santa Cruz Biotechnology, Inc.) and anti-p-p65 (cat.
no. ab16502; Abcam). The slides were observed by a fluorescence
microscope at a magnification of ×200.
Detection of cytokines
Venous blood was collected from the tail vein of
mice, coagulated for 2 h at room temperature and then centrifuged
at 1,800 x g for 15 min at 4°C (Centrifuge 5408R; Eppendorf,
Hamburg, Germany). The upper serum layer was collected to determine
the testosterone levels using the F-TESTO ELISA kit (cat. no.
JL10895; Shanghai Jianglai Biotechnology Co., Ltd., Shanghai,
China), according to the manufacturer's protocol.
Flow cytometry
Leydig and TM3 cells were stained with fluo-rescein
isothiocyanate and propidium iodide, and then analysed with a BD™
LSR II flow cytometer (all BD Biosciences, San Jose, CA, USA) using
FlowJo software (version 10; FlowJo LLC, Ashland, OR, USA).
Cycloheximide treatment
TM3 shN and shR cells were seeded into a six-well
plate at a density of 1×106 cells/well. Cells were then
treated with 50 µg/ml cycloheximide (Sigma-Aldrich; Merck
KGaA) for 0, 20, 40 and 60 min. For this treatment, cells were
culture in DMEM/F-12 supplemented with 5% fetal bovine serum, 2.5%
horse bovine serum, L-glutamine (150 mg/l), NaHCO3 (1.5
g/l), penicillin (100 U/ml) and streptomycin (100 µg/ml) at
37°C with 5% CO2.
Statistical analysis
All data analyses were conducted using the Student's
t-test or one-way analysis of variance followed by a Fisher's Least
Significant Difference post-hoc test for multiple comparisons with
SPSS v12.0 software (SPSS, Inc., Chicago, IL, USA). Values are
presented as the mean ± standard error of the mean. P<0.05 was
considered to indicate a statistically significant difference.
Results
Validation of inflammation models
Testes were collected from C57BL/6 WT mice upon LPS
treatment for hematoxylin and eosin (H&E) staining and western
blot analysis. The results of the western blot analysis indicated
that the p-p65 levels were significantly increased in the
inflammatory group (Fig. 1A).
H&E staining revealed that the testicular structure of the
inflammatory group was damaged compared with the control group
(Fig. 1B). These results
confirmed the successful establishment of a mouse model of
testicular inflammation. The western blot analysis results of
primary Leydig cells demonstrated that the p-p65 expression levels
in the Leydig cells in the inflammation group were increased
compared with those in the control group (Fig. 1C). For the TM3 cell line, the
inflammation was most pronounced in cells treated with LPS at a
concentration of 5 mg/ml (Fig.
1D) for 10 min (Fig. 1E).
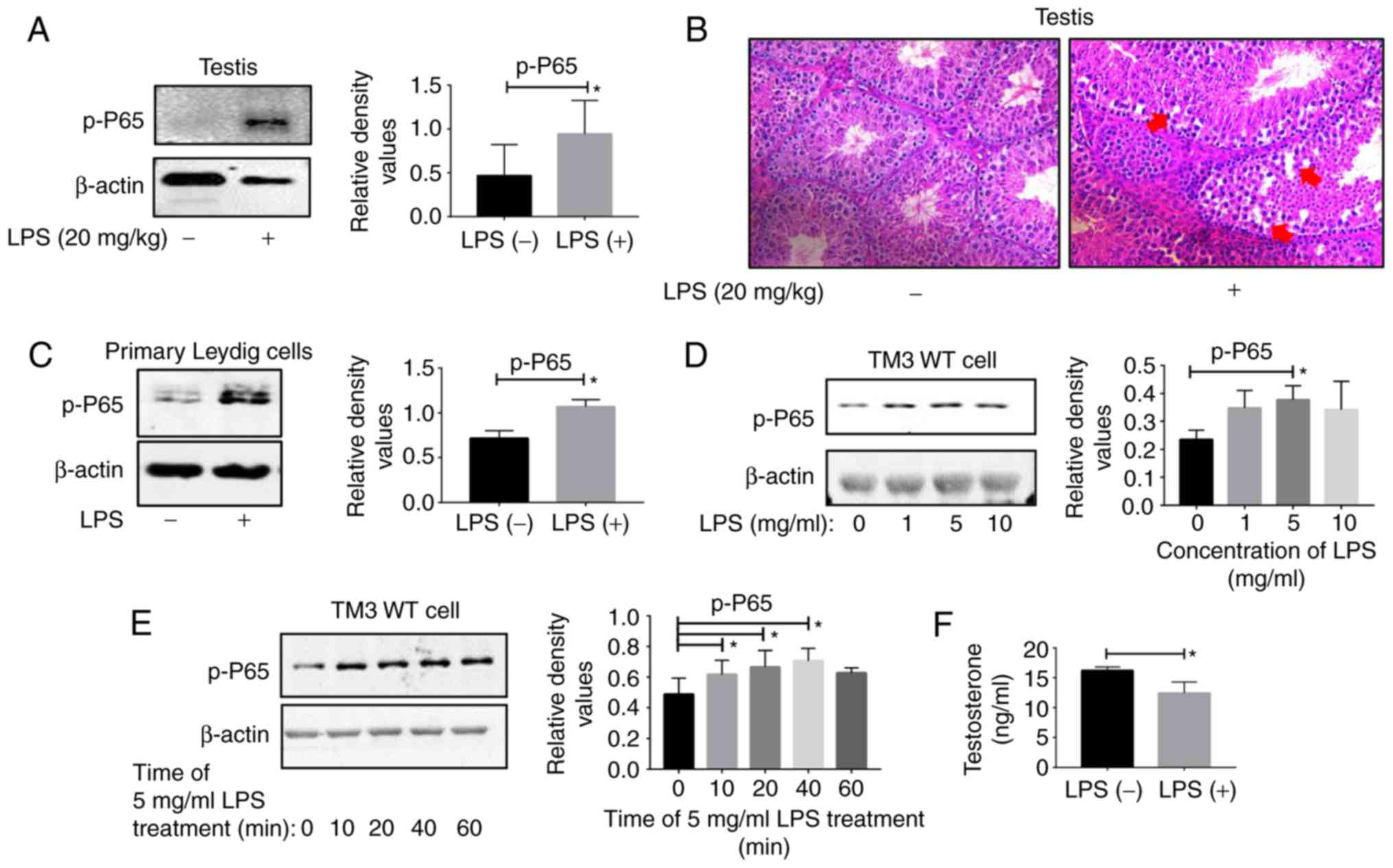 | Figure 1LPS induces testicular inflammation
in mice. Testis tissues were collected from two groups of WT C57
male mice treated with double distilled water and LPS (20 mg/kg)
for 6 h. (A) Proteins collected from testis tissues were subjected
to western blot analysis. (B) The testis tissues of the two groups
were subjected to hematoxylin and eosin staining. The damaged
testis tissues are highlighted by the red arrows. Magnification,
×20. (C) Leydig cells were extracted from the testis tissues of the
two groups, and proteins were collected from Leydig cells for
western blot analysis. (D) TM3 WT cells were treated with different
concentrations of LPS (0,1, 5 and 10 mg/ml) and analyzed by western
blot analysis. (E) TM3 WT cells were treated with LPS at 5 mg/ml
for different times (0, 10, 20, 40 and 60 min) and subjected to
western blot analysis. (F) The serum testosterone levels of the two
groups were determined with an ELISA kit. *P<0.05.
Representative data from 3 replicates are presented. WT, wild-type;
LPS, lipopolysaccharide; p, phosphorylated; p65, transcription
factor p65. |
Inflammation affects the function of
Leydig cells
In addition, the ELISA results demonstrated that the
serum testosterone levels in the inflammatory group were decreased
compared with in the control group (Fig. 1F). Leydig cells are the primary
cells that secrete testosterone, which indicated that the function
of these cells was impaired.
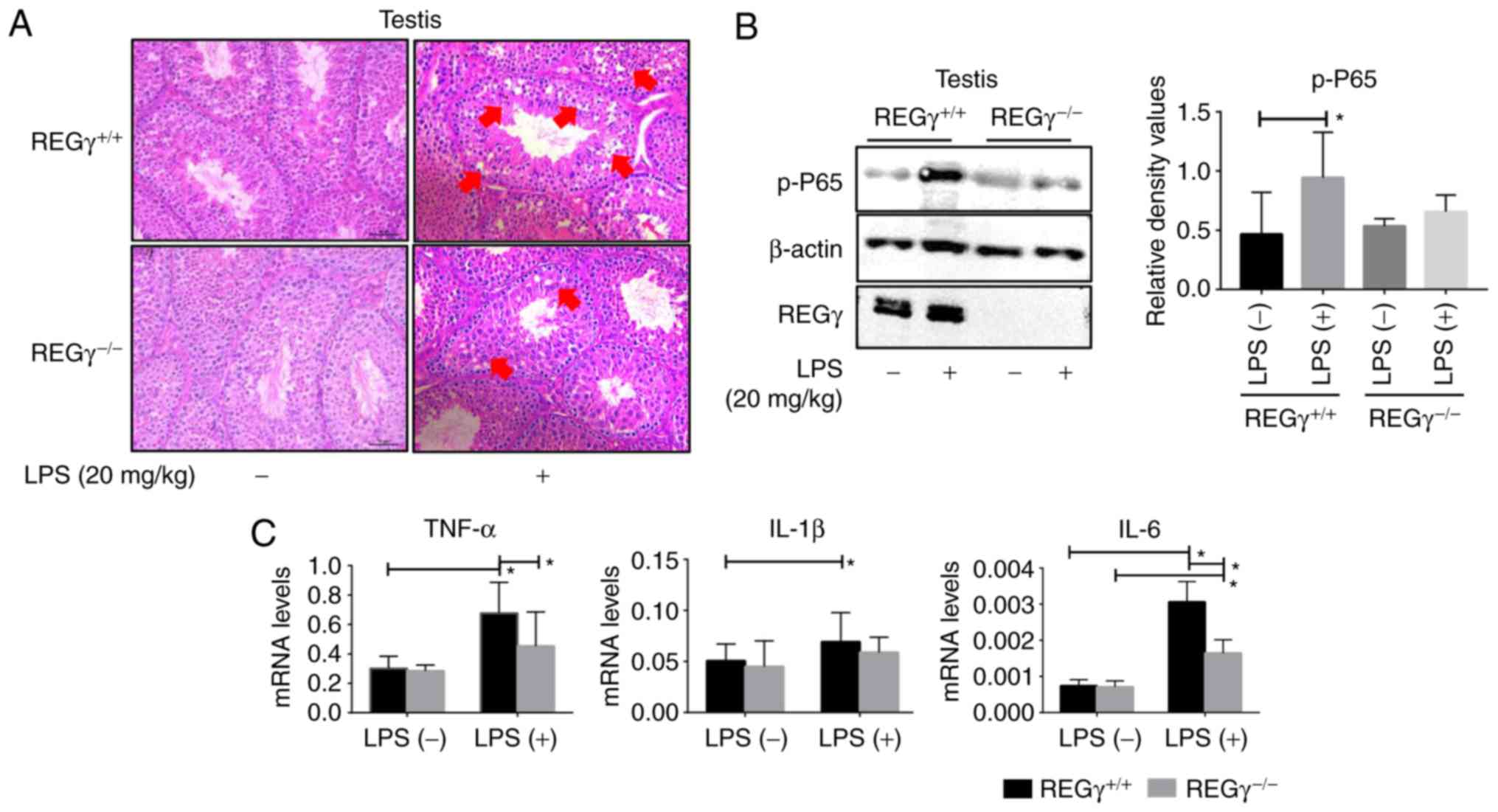
REGγ deficiency decreases LPS-induced
inflammation in vivo
Control and inflammatory groups were established in
6-week-old C57 WT male mice and REGγ−/− mice. Then,
testes from these mice were collected for western blot analysis,
qPCR assays and H&E staining. The western blot analysis results
revealed that the expression level of p-p65 was significantly
decreased in REGγ−/− mice (Fig. 2B) compared with WT mice. These
results were confirmed by H&E staining (Fig. 2A). The results of qPCR analyses
demonstrated that the expression levels of TNF-α, IL-1β and IL-6
were significantly increased in the C57 WT mice testes compared
with the REGγ−/− mice testes following LPS treatment
(Fig. 2C).
REGγ deficiency decreases LPS-induced
inflammation in vitro
Primary Leydig cells were extracted from C57 WT and
REGγ−/− mice testes and divided into LPS- or
LPS+ groups. Then, western blot analysis and qPCR assays
were performed. The western blot analysis results revealed that
p-p65 expression was significantly decreased in REGγ−/−
mice compared with the WT group (Fig.
3A). The qPCR results indicated that the expression levels of
TNF-α were significantly decreased in REGγ−/− mice
compared with the WT group (Fig.
3B).
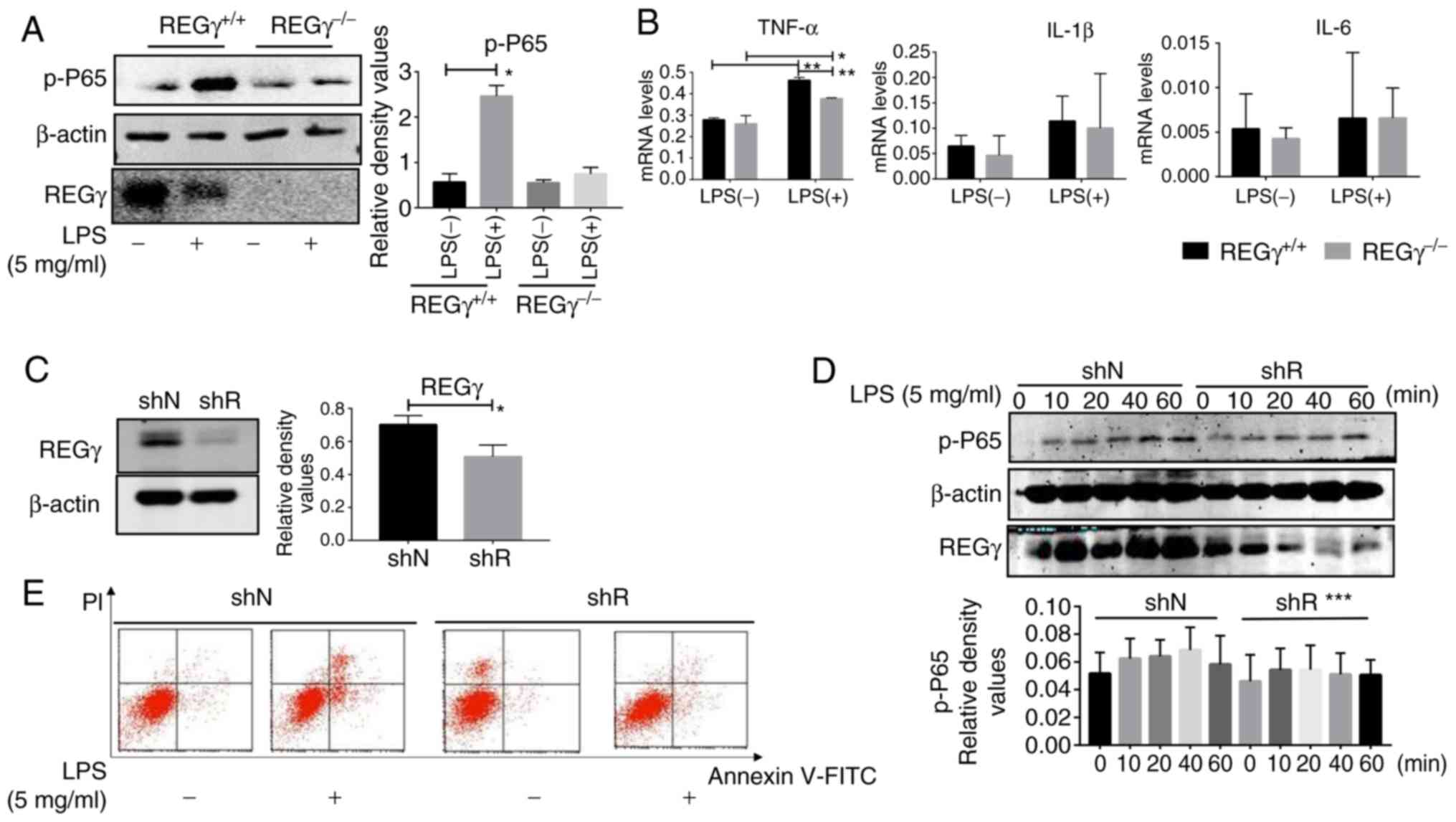 | Figure 3REGγ deficiency decreases LPS-induced
inflammation in vitro. (A) Testis tissues were collected
from REGγ+/+ and REGγ−/− mice treated with
double distilled water and LPS (20 mg/kg) for 6 h. Leydig cells
were extracted from the testis tissues of each group, and proteins
were then collected from Leydig cells for western blot analysis.
(B) mRNA expression of TNF-α, IL-1β and IL-6 in the Leydig cells of
each group was evaluated by quantitative polymerase chain reaction.
(C) shR TM3 cells and shN TM3 cells were subjected to western blot
analysis to evaluate the expression of REGγ. (D) shN and shR cells
were treated with 5 mg/ml LPS for different times (0, 10, 20, 40
and 60 min) prior to being subjected to western blot analysis. (E)
shN and shR cells treated with 5 mg/ml LPS for 10 min were
collected for flow cytometry analysis to evaluate apoptosis.
*P<0.05, **P<0.01 and
***P<0.001. Representative data from 3 replicates are
presented. REGγ, proteasome activator complex subunit 3; LPS,
lipopolysaccharide; p, phosphorylated; p65, transcription factor
p65; TNF-α, tumor necrosis factor-α; IL, interleukin.; shR,
REGγ-knockdown; shN, negative control; FITC, fluorescein
isothiocyanate; PI, propidium iodide. |
Western blot analysis confirmed that REGγ was
successfully knocked down (Fig.
3C). shN and shR cells were treated with LPS (5 mg/ml) for 0,
10, 20, 40 and 60 min. The western blot analysis results revealed
that the expression level of p-p65 in the shN groups was increased
compared with in the shR groups, with increasing incubation time
(Fig. 3D).
REGγ deficiency promotes apoptosis
Flow cytometry was used to detect levels of cell
apoptosis. Briefly, shN and shR cells were treated with 5 mg/ml LPS
for 10 min. The results indicated that the number of apoptotic
cells was significantly increased in the shR cells compared with
the shN cells (Fig. 3E).
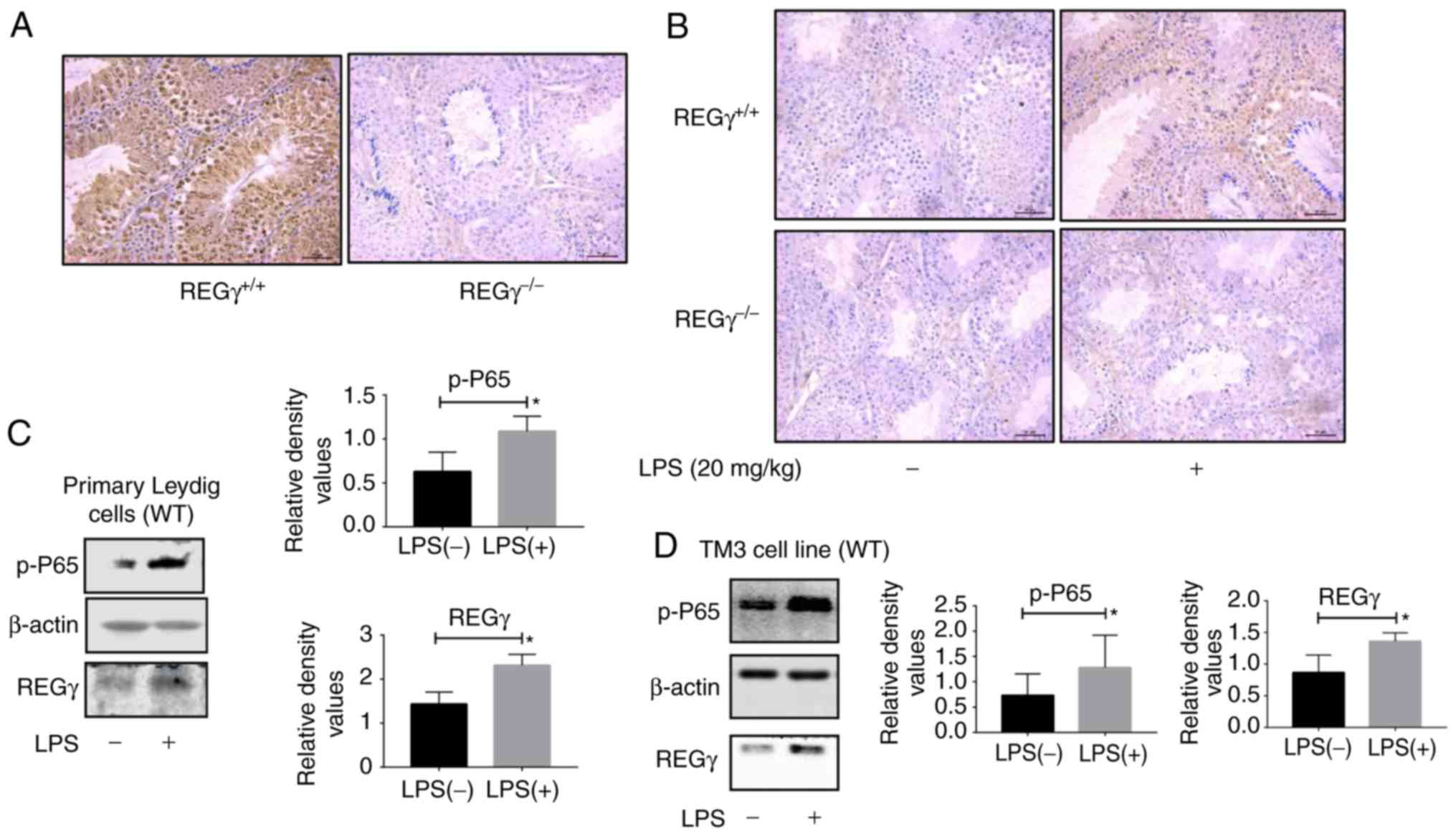
Reciprocal regulation of REGγ and NF-κB
in vivo and in vitro
Inflammation models were established in two groups
of REGγ+/+ and REGγ−/− mice. The testes of
these mice were collected, sliced and stained with an anti-REGγ
antibody. The immunohistochemical staining results revealed no REGγ
expression in REGγ−/− mice testes, whereas REGγ was
highly expressed in REGγ+/+ mouse testis (Fig. 4A). The expression level of p-p65
was markedly decreased in the REGγ−/− mice compared with
the REGγ+/+ mice following LPS treatment (Fig. 4B). LPS treatment increased the
expression level of REGγ in Leydig cells of REGγ+/+ mice
(Fig. 4C). In addition, LPS
treatment upregulated the expression levels of REGγ in TM3 WT cells
compared with the control group (Fig.
4D).
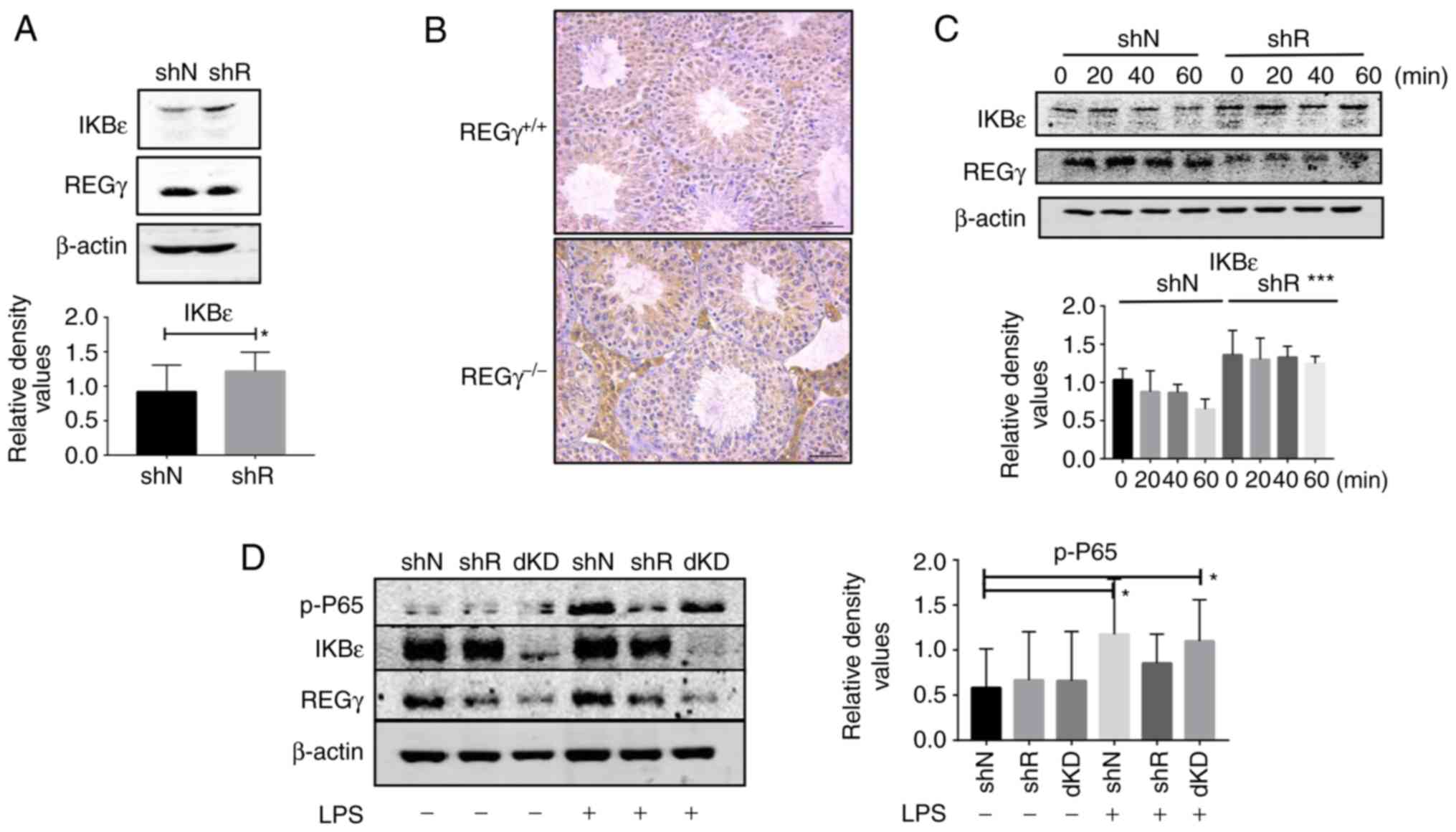
REGγ promotes NF-κB activity by degrading
IkBε
To explore the mechanism behind the association
between REGγ and NF-κβ in Leydig cells, several upstream signaling
pathways were identified from previous studies (5-7,12).
Of these, the present study focused on the IkB family proteins.
Western blot analysis of IkBα, IkBβ and IkBε were performed in shN
and shR cells. The results demonstrated significant differences in
IkBε expression levels between shN and shR cells (Fig. 5A). The results of the
immunohistochemical staining also revealed that IkBε expression
levels were increased in the testicular tissues of
REGγ−/− mice compared with REGγ+/+ mice
(Fig. 5B). Based on these
results, cycloheximide degradation analyses were conducted. The
results revealed that IkBε degradation was increased in shN cells
compared with in shR cells treated for the same time interval.
These results demonstrated that the degradation of IkBε increased
with increased expression of REGγ (Fig. 5C).
REGγ/IkBε double knockdown (dKD) restores
inflammation levels
shRNA was used to knock down the REGγ and IkBε genes
in the TM3 cell line. shN, shR (IkBε knockdown) and dKD (REGγ and
IkBε knockdown) cells were treated with 5 mg/ml LPS for 10 min. The
western blot analysis results demonstrated that the expression
levels of REGγ and IkBε were significantly decreased in the dKD
cells compared with the shN cells (Fig. 5D). In addition, p-p65 was highly
expressed in dKD cells, which was similar to the expression levels
observed in WT cells (Fig.
5D).
Discussion
The results of the present study demonstrated that
the protease activator REGγ was involved in the development of
Leydig cell inflammation in a mouse model of LPS-induced
inflammation. These results were additionally validated in primary
Leydig cells and theTM3 cell line. Deletion of REGγ increased the
accumulation of IkBε and inhibited the activation of the NF-κB
signaling pathway, thereby inhibiting inflammation. Furthermore,
dKD of REGγ and IkBε produced a successful cell model of
inflammation.
With environmental degradation and unhealthy habits,
male fertility continues to exhibit a downward trend (14). Numerous factors contribute to male
infertility (15-17), including congenital genital
abnormalities, genetic and endocrine diseases, reproductive system
infection and inflammation, and physical and chemical factors.
Inflammation of the male reproductive system is an important
pathogenic factor of male infertility (16).
LPS is the primary constituent of the cell wall of
gram-negative bacteria and a key cause of infection (18). LPS has been widely used to
generate animal models of inflammation to study the effect of
inflammation in vivo. Injection of LPS produces an
inflammatory response and inhibits testicular steroid production
and subsequent spermatogenesis, and may also damage the
blood-testis barrier, thereby decreasing fertility (19). In a previous study, C57 mice were
intraperitoneally injected with LPS to induce an inflammatory
response, in order to observe the histopathological changes in the
cells (19). In the present
study, the serum testosterone levels decreased following LPS
treatment, which indicated that inflammation damaged the normal
functions of the Leydig cells.
Previous studies have demonstrated the associations
between REGγ regulation and various diseases, which have confirmed
the important roles of REGγ in the regulation of biological
processes (20-23). The majority of these studies on
REGγ have focused on cancer, neurological diseases and heart
disease (20-23), while few have investigated the
association between REGγ and inflammation. A previous study
revealed that REGγ was involved in the development of inflammatory
bowel disease in a mouse model of enteritis (12). Therefore, the present study
focused on the role of REGγ in testicular inflammation.
The NF-κβ signaling pathway has been recognised as a
key signaling pathway in the development of inflammatory-associated
diseases (24,25). As IkBε is activated upstream of
NF-κβ, previous studies explored the association between REGγ and
inflammation (26-28). The results of the present study
demonstrated the role of the protein kinase-activating factor REGγ
in testicular inflammation in animal and cell models.
Testicular tissue damage and NF-κβ activation
induced a high expression level of REGγ in Leydig cells, and REGγ
upregulated the activity of the NF-κβ signaling pathway by
degrading IkBε, which led to the secretion of proinflammatory
cytokines and the persistence of testicular Leydig cell and testis
injury, ultimately promoting the development of testicular Leydig
cell inflammation. Therefore, the absence of REGγ may promote the
accumulation of IkBε, thereby inhibiting the activity of the NF-κβ
signaling pathway in testicular Leydig cells in mice.
To the best of our knowledge, the present study
demonstrated the physiological functions of REGγ and IkBε in
testicular inflammation for the first time. REGγ is a specific
regulatory factor of IkBε in testicular Leydig cells, which extends
awareness of the IkBε regulatory pathway. The testicular tissue of
REGγ−/− mice exhibited decreased levels of inflammatory
molecules. However, the roles of other testicular tissue cells in
this REGγ testicular inflammation model were not investigated.
Therefore, the effects of other types of cells in this process
cannot be ruled out. The knockdown of IkBε was performed in the
present study, but the results were not satisfactory as no
significant differences were identified. This is a limitation of
the study, and will be a focus in future studies.
Various anti-inflammatory drugs, including
glucocorti-coids, methotrexate and anti-TNF-α antibodies,
completely or partially inhibited the NF-κβ signaling pathway
(29). However, these
anti-inflammatory drugs do not specifically inhibit NF-κβ, and the
response to these drugs differs among individuals, which highlights
the ongoing challenges in proteasome studies. Notably, the results
of the present study indicated that REGγ regulates NF-κβ activity
by specifically degrading IkBε, thereby providing novel molecular
targets of the atypical proteasomal pathway. It may be possible to
prevent and treat inflammation by attenuating, as opposed to
completely inhibiting the NF-κβ signaling pathway.
Abbreviations:
|
LPS
|
lipopolysaccharide
|
|
ATP
|
adenosine triphosphate
|
|
WT
|
wild-type
|
|
H&E
|
hematoxylin and eosin
|
|
qPCR
|
quantitative polymerase chain
reaction
|
|
dd water
|
double distilled water
|
|
dKD
|
double knockdown
|
Funding
The present study was partially supported by a grant
from the National Natural Science Foundation of China (grant no.
81671446).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL and YX designed the experiments. TX, HC, SS, GH
and BH performed the experiments and TH analyzed the data. TX wrote
the manuscript.
Ethics approval and consent to
participate
All animal procedures were reviewed and approved by
the Animal Care Committee of East China Normal University, which
followed the Guide for the Care and Use of Laboratory Animals by
the National Research Council.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Krysiak-Baltyn K, Brunak S and Jensen TS:
Environmental and life style factors about male reproductive
disorders. Kgs Lyngby: Technical University of Denmark (DTU);
2012
|
|
2
|
Silva CA, Cocuzza M, Carvalho JF and Bonfá
E: Diagnosis and classification of autoimmune orchitis. Autoimmun
Rev. 13:431–434. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hong CY, Park JH, Ahn RS, Im SY, Choi HS,
Soh J, Mellon SH and Lee K: Molecular mechanism of suppression of
testicular steroidogenesis by proinflammatory cytokine tumor
necrosis factor alpha. Mol Cell Biol. 24:2593–2604. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dobashi M, Fujisawa M, Yamazaki T, Okuda
Y, Kanzaki M, Tatsumi N, Tsuji T, Okada H and Kamidono S:
Inhibition of steroidogenesis in Leydig cells by exogenous nitric
oxide occurs independently of steroidogenic acute regulatory
protein (star) mRNA. Arch Androl. 47:203–209. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barnes PJ and Karin M: Nuclear factor-κB-A
pivotal transcription factor in chronic inflammatory diseases. N
Engl J Med. 336:1066–1071. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tak PP and Firestein GS: NF-kappaB: A key
role in inflammatory diseases. J Clin Invest. 107:7–11. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guazzone VA, Jacobo P, Theas MS and Lustig
L: Cytokines and chemokines in testicular inflammation: A brief
review. Microsc Res Tech. 72:620–628. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goldberg AL: Protein degradation and
protection against misfolded or damaged proteins. Nature.
426:895–899. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang Q and Figueiredo-Pereira ME:
Ubiquitin/proteasome pathway impairment in neurodegeneration:
Therapeutic implications. Apoptosis. 15:1292–1311. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ferrell K, Wilkinson CR, Dubiel W and
Gordon C: Regulatory subunit interactions of the 26S proteasome, a
complex problem. Trends Biochem Sci. 25:83–88. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Barton LF, Runnels HA, Schell TD, Cho Y,
Gibbons R, Tevethia SS, Deepe GS Jr and Monaco JJ: Immune defects
in 28-kDa proteasome activator gamma-deficient mice. J Immunol.
172:3948–3954. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu J, Zhou L, Ji L, Chen F, Fortmann K,
Zhang K, Liu Q, Li K, Wang W, Wang H, et al: The REGγ-proteasome
forms a regulatory circuit with IκBε and NFκB in experimental
colitis. Nat Commun. 22:107612016. View Article : Google Scholar
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
14
|
Kumar N and Singh AK: Trends of male
factor infertility, an important cause of infertility: A review of
literature. J Hum Reprod Sci. 8:191–196. 2015. View Article : Google Scholar
|
|
15
|
Oliva A, Spira A and Multigner L:
Contribution of environmental factors to the risk of male
infertility. Hum Reprod. 16:1768–1776. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sikka SC: Relative impact of oxidative
stress on male reproductive function. Curr Med Chem. 8:851–862.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Du Plessis SS, Cabler S, McAlister DA,
Sabanegh E and Agarwal A: The effect of obesity on sperm disorders
and male infertility. Nat Rev Urol. 7:153–161. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schwandner R, Dziarski R, Wesche H, Rothe
M and Kirschning CJ: Peptidoglycan- and lipoteichoic acid-induced
cell activation is mediated by toll-like receptor 2. J Biol Chem.
274:17406–17409. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie T, Hu G, Dong B, Yan Y, Liu M, Yao X,
Zheng J and Xu Y: Roscovitine protects murine Leydig cells from
lipopolysaccharide-induced inflammation. Exp Ther Med.
13:2169–2176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ali A, Wang Z, Fu J, Ji L, Liu J, Li L,
Wang H, Chen J, Caulin C, Myers JN, et al: Differential regulation
of the REGγ-proteasome pathway by p53/TGF-β signalling and mutant
p53 in cancer cells. Nat Commun. 4:26672013. View Article : Google Scholar
|
|
21
|
Li L, Zhao D, Wei H, Yao L, Dang Y, Amjad
A, Xu J, Liu J, Guo L, Li D, et al: REGγ deficiency promotes
premature aging via the casein kinase 1 pathway. Proc Natl Acad Sci
USA. 110:11005–11010. 2013. View Article : Google Scholar
|
|
22
|
Dong S, Jia C, Zhang S, Fan G, Li Y, Shan
P, Sun L, Xiao W, Li L, Zheng Y, et al: The REGγ proteasome
regulates hepatic lipid metabolism through inhibition of autophagy.
Cell Metab. 18:380–391. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu S, Lai L, Zuo Q, Dai F, Wu L, Wang Y,
Zhou Q, Liu J, Liu J, Li L, et al: PKA turnover by the
REGγ-proteasome modulates FoxO1 cellular activity and VEGF-induced
angiogenesis. J Mol Cell Cardiol. 72:28–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fairchild KD, Singh IS, Patel S, Drysdale
BE, Viscardi RM, Hester L, Lazusky HM and Hasday JD: Hypothermia
prolongs activation of NF-kappaB and augments generation of
inflammatory cytokines. Am J Physiol Cell Physiol. 287:C422–C431.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Killeen MJ, Linder M, Pontoniere P and
Crea R: NF-κβ signaling and chronic inflammatory diseases:
Exploring the potential of natural products to drive new
therapeutic opportunities. Drug Discov Today. 19:373–378. 2014.
View Article : Google Scholar
|
|
26
|
O'Dea E and Hoffmann A: The regulatory
logic of the NF-kappaB signaling system. Cold Spring Harb Perspect
Biol. 2:a0002162010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
O'Dea EL, Kearns JD and Hoffmann A: UV as
an amplifier rather than inducer of NF-kappaB activity. Mol Cell.
30:632–642. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hoffmann A, Levchenko A, Scott ML and
Baltimore D: The IkappaB-NF-kappaB signaling module: Temporal
control and selective gene activation. Science. 298:1241–1245.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barnes PJ: Anti-inflammatory actions of
glucocorticoids: Molecular mechanisms. Clin Sci (Lond). 94:557–572.
1998. View Article : Google Scholar
|