Introduction
Glutamate is known as an excitatory neurotransmitter
and plays a crucial role in the plasticity of the central nervous
system (CNS) (1,2). When the glutamate concentration is
abnormally high, it functions as a neurotoxin and leads to cell
death (3). Excessive glutamate
levels damage cellulr components, including the mitochondria, and
promote the generation of reactive oxygen species (ROS) and induce
cell death (4). Neuronal cell
loss associated with glutamate neurotoxicity is associated with
certain neurological diseases, such as stroke or trauma, as well as
Alzheimer's, Parkinson's and Huntington's diseases (5-9).
Cells may be protected from oxidative injury by
antioxidant systems. Some intracellular ROS may be counteracted by
superoxide dismutases (SODs), a group of vital antioxidant enzymes,
which include SOD1 and SOD2, as well as by glutathione peroxidase
(GPx-1) (10,11). Metal-containing SODs play a role
in scavenging ROS and facilitate catalytic activity through the use
of copper/zinc (Cu/Zn) or manganese (Mn) (12). SOD1 is a Cu/Zn-containing enzyme
that is located in cytosolic compartments. Low SOD1 levels are
associated with increased ROS levels, which triggers oxidative
damage to cellular components, including to DNA, and the functions
of SOD1 are protect the CNS from damage. The Mn-containing enzyme,
SOD2, is located in the mitochondrial matrix and may protect
mitochondrial DNA from oxidative damage (10,12). GPx-1 is a major enzyme that
protects the cells from lethal oxidative stress by converting
H2O2 to H2O with reduced
glutathione (GSH) as a co-factor (13).
Glutathione is a major cellular oxidant that
protects cells from free radical damage and exists in both reduced
(GSH) and oxidized forms (GSSG). The reduced form is predominant
within the cell and represents >99% of total glutathione
contents (14). Prolonged
oxidative glutamate toxicity causes glutathione depletion in
neurons, and leads to oxidative stress and to the development of
neurodegenerative disorders (15,16). The formation of excessive
oxidative stress also activates mitogen-activated protein kinases
(MAPKs), and results in cell damage and apoptosis. Therefore, it is
important to identify antioxidants that protect neuronal cells from
free radical damage and inhibit cell death.
MAPKs are serine-threonine protein kinases that
regulate a variety of cellular activities, such as cell
proliferation, differentiation, survival and death (17,18). The MAPK family is composed of
extracellular signal-regulated kinases (ERKs), stress-activated
c-Jun N-terminal kinases (JNKs)/stress-activated protein kinases
and p38 kinases (19). Among
these, p38 MAPKs play important roles in mediating apoptotic
pathways (20). Glutamate-induced
oxidative stress may activate p38 MAPKs (21) and activated p38 kinase may induce
apoptosis by regulating caspases (22).
Scrophularia buergeriana (SB) is known as
Hyun-Sam in Korea and is traditionally used to treat fever,
swelling, constipation and age-related memory loss in Northern
China (23). The dried root of SB
possesses compounds, such as phenylpropanoids (24), 7-harpagide-type iridoids (25), E-harpagoside,
8-O-E-p-methoxycinnamoylharpagide (MCA-Hg) (26) and E-p-methoxycinnamic acid
(27), which display
neuroprotective activities. However, food and medicines cannot be
evaluated as whole materials and are instead fractionated into
organic solvents for analysis. In a previous study, the ethanolic
extract of SB (SBE) was found to exhibit cognitive-enhancing and
antioxidant activities. However, the effects of SBE on cells have
not yet been associated with the p38 MAPK pathway and
anti-apoptotic mechanisms, at least to the best of our knowledge.
Therefore, in this study, we investigated whether SBE reduces
oxidative stress and exerts neuroprotective effects against
glutamate-induced neurotoxicity via antioxidant and anti-apoptotic
mechanisms in SH-SY5Y cells.
Materials and methods
Sample preparation
The SBE extraction process was as follows and as
previously described (18): The
pieces of 3-10 cm cut dried roots of SB obtained from Nutrapharmtec
(Seongnam, Korea) were extracted with 70% EtOH. The ratio between
the dried extract and solvent was 1:8 and the extraction proceeded
for 2 h at 90°C before filtering. The extracts were then
concentrated in a vacuum evaporator and the concentrate was
sterilized and cooled. The residue was dried, and a powder was
obtained. The standardized SBE sample was dissolved in dimethyl
sulfoxide (DMSO) prior to use.
Cells and cell culture
The SH-SY5Y human neuroblastoma cells used in this
study were purchased from the American Type Culture Collection
(ATCC, Manassas, VA, USA; CRL-2266) and were authenticated via a
short tandem repeat (STR) profiling service provided by ATCC. The
cells were cultured in Dulbecco's modified Eagle's medium (DMEM; GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (FBS) and 1% antibiotics (both from Life
Technologies; Thermo Fisher Scientific, Waltham, MA, USA)
containing 100 U/ml penicillin and 100 g/ml streptomycin. The cells
were incubated for 24 h at 37°C in a humidified incubator with 5%
CO2.
Cell cytotoxicity assay
The SH-SY5Y cells were seeded in 96-well plates at a
density of 5x104 cells/well and incubated for 24 h. The
following day, the cells were pre-treated with various
concentrations (125, 250 or 500 µg/ml) of SBE for 1 h and then
exposed to 100 mM glutamate (Wako, Osaka, Japan) for 3 h with or
without SBE. Sample stock solution was prepared (125, 250 and 500
mg/ml) and diluted 1/1,000 in the medium to allow the working
concentration (125, 250 and 500 µg/ml) to be processed. Following a
3-h incubation period at 37°C with 5% CO2 conditions, 10
µl cell viability assay reagent (DoGenBio, Seoul, Korea) was added
and the cells were incubated at 37°C with 5% CO2 for a
further 4 h. The absorbance was measured at 450 nm using a
microplate reader (Tecan, Männedorf, Switzerland) and cell
viability was expressed as a percentage relative to that in the
control group (100%) with medium and DMSO (Wako) treatment not
exposed to glutamate.
Measurement of acetylcholinesterase
(AchE) activity
To measure AchE activity, the SH-SY5Y cells were
treated with 5% all-trans retinoic acid (RA; Sigma-Aldrich, St.
Louis, MO, USA) to induce differentiation. The culture medium was
replaced with fresh medium containing RA every 2 days until day 6.
On day 7, the differentiated cells were pre-treated with SBE for 1
h and the medium was exchanged for that containing glutamate with
or without SBE. The cells were further incubated at 37°C with 5%
CO2 conditions for 3 h. The cells were then washed with
ice-cold phosphate buffered saline (PBS)3 times, lysed with 0.1 M
phosphate buffer (pH 7.5), centrifuged at 2,000 x g for 10 min, and
the supernatant was collected. AchE activity was determined using
commercial assay kits (Abnova, Taipei, Taiwan) and was calculated
as the optical density (OD) at 412 nm per mg protein.
Total glutathione contents
Total intracellular glutathione contents were
determined using a commercial assay kit (Cell Biolabs, San Diego,
CA, USA) according to the manufacturer's protocol. The cells
pre-treated with SBE for 1 h were changed to medium containing
glutamate with or without SBE for 3 h. The cells were then washed
with ice-cold PBS, lysed using assay reagent and centrifuged at
2,000 x g for 10 min. The supernatant was used to measure the
glutathione contents as the sum of GSH and GSSG. The absorbance was
measured at 405 nm using a microplate reader (Tecan) and total
glutathione contents were expressed as a percentage relative to
that in the control group (100%).
4′,6-Diamidino-2-phenylindole (DAPI)
staining
Glutamate-induced nuclear morphological changes were
assessed by DAPI staining (Sigma-Aldrich). Following glutamate
treatment with and without sample, cells were washed with ice-cold
PBS three times and fixed with 4% paraformaldehyde in PBS for 10
min. Fixed cells were washed with PBS and permeabilized with 0.25%
Triton X-100 in PBS for 10 min. After washing again with PBS, the
cells were stained with DAPI staining solution for 10 min at room
temperature. The stained cells were examined under a fluorescence
microscope (x40 magnification; Nikon, Tokyo, Japan) to confirm the
presence of apoptotic cells exhibiting size-reduced nuclei,
chromatin condensation, intense fluorescence and nuclear
fragmentation. The number of apoptotic cells was expressed as a
percentage relative to that in the control group (100%).
Terminal deoxynucleotidyl transferase
deoxyuridine triphosphate (dUTP) nick end labeling (TUNEL)
assay
Glutamate-induced DNA damage was evaluated using a
TUNEL assay commercial kit (BioVision, Milpitas, CA, USA) according
to the manufacturer's instructions. The SH-SY5Y cells were seeded
and incubated for 24 h in 5% CO2 at 37°C. The cells were
then pre-treated with various concentrations (125, 250 or 500
µg/ml) of SBE for 1 h and exposed to 100 mM glutamate for 3 h with
or without SBE. The treated cells were washed with ice-cold PBS and
fixed by the addition of 1% paraformaldehyde in PBS on ice for 15
min. The cells were then treated with 70% ethanol (EtOH) for 30 min
on ice and then washed 3 times with wash buffer. A total of 50 µl
staining solution contained with FICT-dUTP was added to the washed
cells prior to incubation for 60 min at 37°C. After staining, the
cells were washed with rinse buffer 3 times and incubated with
propidium iodide (PI)/RNase solution in the dark for 30 min at room
temperature. The stained cells were analyzed under a fluorescence
microscope (x20 magnification; Nikon).
Protein extraction and western blot
analysis
The treated SH-SY5Y cells were lysed with
radioimmunoprecipitation assay (RIPA) buffer [50 mM Tris-HCl pH
7.4, 150 mM NaCl, 1 mM ethylenediaminetetraacetic acid (EDTA), 1%
Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate
(SDS) and 1 mM PMSE] and 1% protease inhibitor cocktail (Roche,
Basel, Switzerland), followed by centrifugation at 2,000 x g for 15
min at 4°C. The supernatant was collected, and the protein
concentration was assessed using a bicinchoninic acid (BCA) protein
assay kit (Thermo Fisher Scientific). Equal amounts of protein (20
µg/lane) were separated by 10-15% SDS-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene difluoride
membranes (Bio-Rad, Hercules, CA, USA). The membranes were blocked
with commercial blocking buffer (Thermo Fisher Scientific) and
washed 3 times with Tris-buffered saline containing 0.1% Tween-20
(TBST). After washing, the membranes were incubated at 4°C
overnight with the following appropriate antibodies: SOD1 (1:1,000;
cat. no. sc-515404), SOD2 (1:1,000; cat. no. sc-137254) (both from
Santa Cruz Biotechnology, Santa Cruz, CA, USA), GPx-1 (1:1,000;
cat. no. 3206), p-p38 (1:1,000; cat. no. 9211), p38 (1:1,000; cat.
no. 8690), Bax (1:1,000; cat. no. 2772), Bcl-2 (1:1,000; cat. no.
3498), cleaved caspase-3 (1:1,000; cat. no. 9664), cleaved PARP
(1:1,000; cat. no. 5625) and β-actin (1:2,000; cat. no. 8457) (all
from Cell Signaling Technology, Danvers, MA, USA) washed 3 times
with TBST, and incubated with horseradish peroxidase-conjugated
secondary antibodies: goat anti-rabbit (1:1,000; cat. no. SA002),
goat anti-mouse (1:1,000; cat. no. SA001) (both from genDEPOT,
Katy, TX, USA) for 1 h at room temperature. The membranes were
washed 3 times and visualized via enhanced chemiluminescence (ATTO,
Tokyo, Japan). Densitometry was performed using Image-Pro Plus
software (6.0 version; Media Cybernetics, Inc., USA)
Statistical analysis
Data are expressed as the means ± standard error of
the mean (SEM) and were analyzed using SPSS Statistics 22.0
software (SPSS Inc., Chicago, IL, USA). Different treatment groups
were compared using the Student's t-test and one-way analysis of
variance (ANOVA) followed by multiple comparisons correction using
Dunnett's post-hoc test using Origin 7.0 software (OriginLab,
Northampton, MA, USA). Differences were considered statistically
significant or highly significant at values of P<0.05 and
P<0.01, respectively.
Results
Effects of SBE on glutamate-induced cell
death and cytotoxicity in SH-SY5Y cells
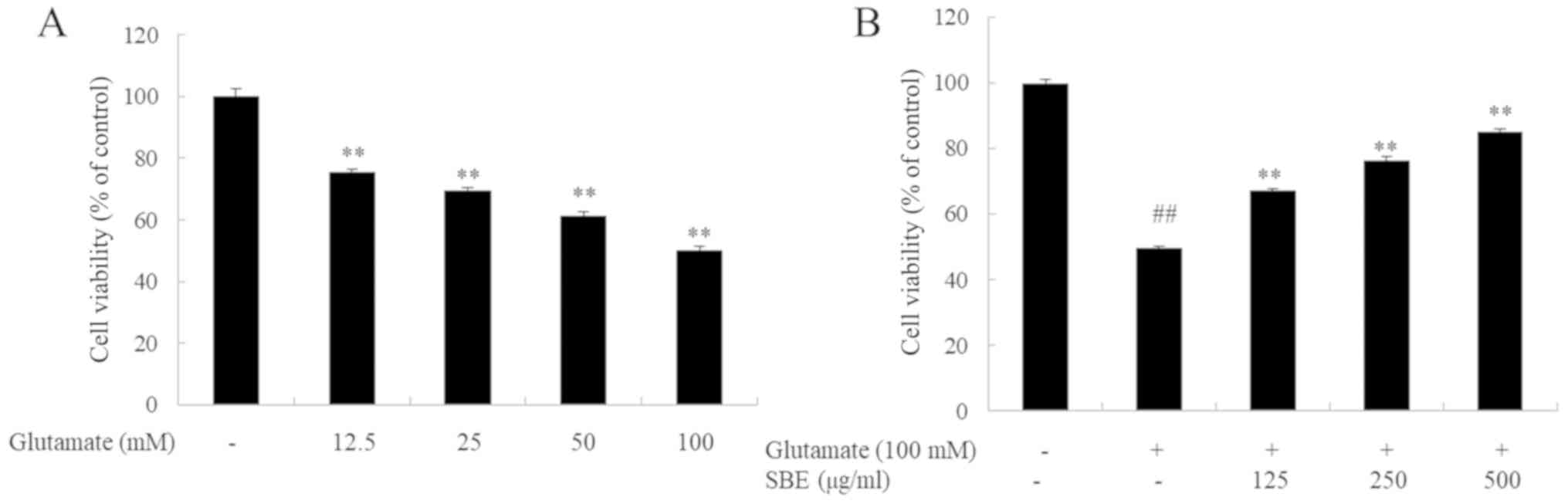
To investigate the protective effects on SBE against
the glutamate-induced death of SH-SY5Y cells, we measured cell
viability. To select the appropriate concentration of glutamate,
the SH-SY5Y cells were exposed to various concentrations of
glutamate for 3 h. As shown in Fig.
1, exposure to 12.5-100 mM glutamate dose-dependently decreased
cell viability: Cell viability decreased by 50% following 3 h of
exposure to 100 mM glutamate compared to the group not exposed to
glutamate. Conversely, the viability of the cells pre-treated with
SBE for 1 h and exposed to glutamate increased by 18, 27 and 36%
compared to that of the cells exposed to glutamate only and not
treated with SBE.
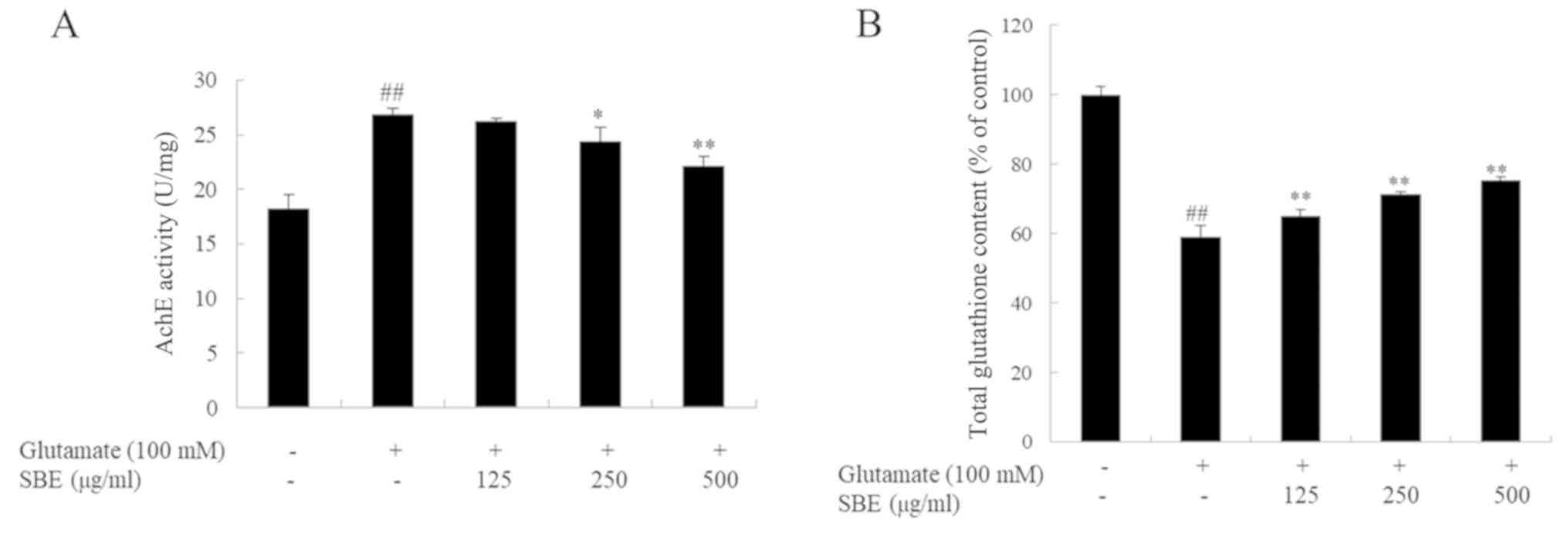
Inhibitory effects of SBE on AchE
activity in glutamate-exposed SH-SY5Y cells
To confirm the neuroprotective effects of SBE, AchE
activity was investigated in the SH-SY5Y cells with
glutamate-induced neurotoxicity. As shown in Fig. 2A, AchE activity in the
glutamate-exposed group was significantly higher than that in the
control group. However, co-treatment with SBE dose-dependently
decreased AchE activity. AchE activity in the groups treated with
250 and 500 µg/ml SBE was reduced by 9.4 and 18.5%, respectively,
compared to that in the group exposed to glutamate only.
Effects of SBE on total glutathione
content in the glutamate-induced apoptosis of SH-SY5Y cells
To evaluate the antioxidant effects of SBE, we
measured the total glutathione content in the glutamate-exposed
SH-SY5Y cells. As expected, and as shown in Fig. 2B, exposure to glutamate induced
oxida-tive stress and markedly decreased the total glutathione
contents in the cells compared to that in the control cells.
However, the total glutathione contents in the SBE-treated cells
were recovered in a dose-dependent manner. The total glutathione
contents in the groups treated with 125, 250 and 500 µg/ml SBE were
increased by 9.3, 17.1 and 21.5%, respectively, compared to those
in the group exposed to glutamate only; these results provide
evidence of the antioxidant effects of SBE.
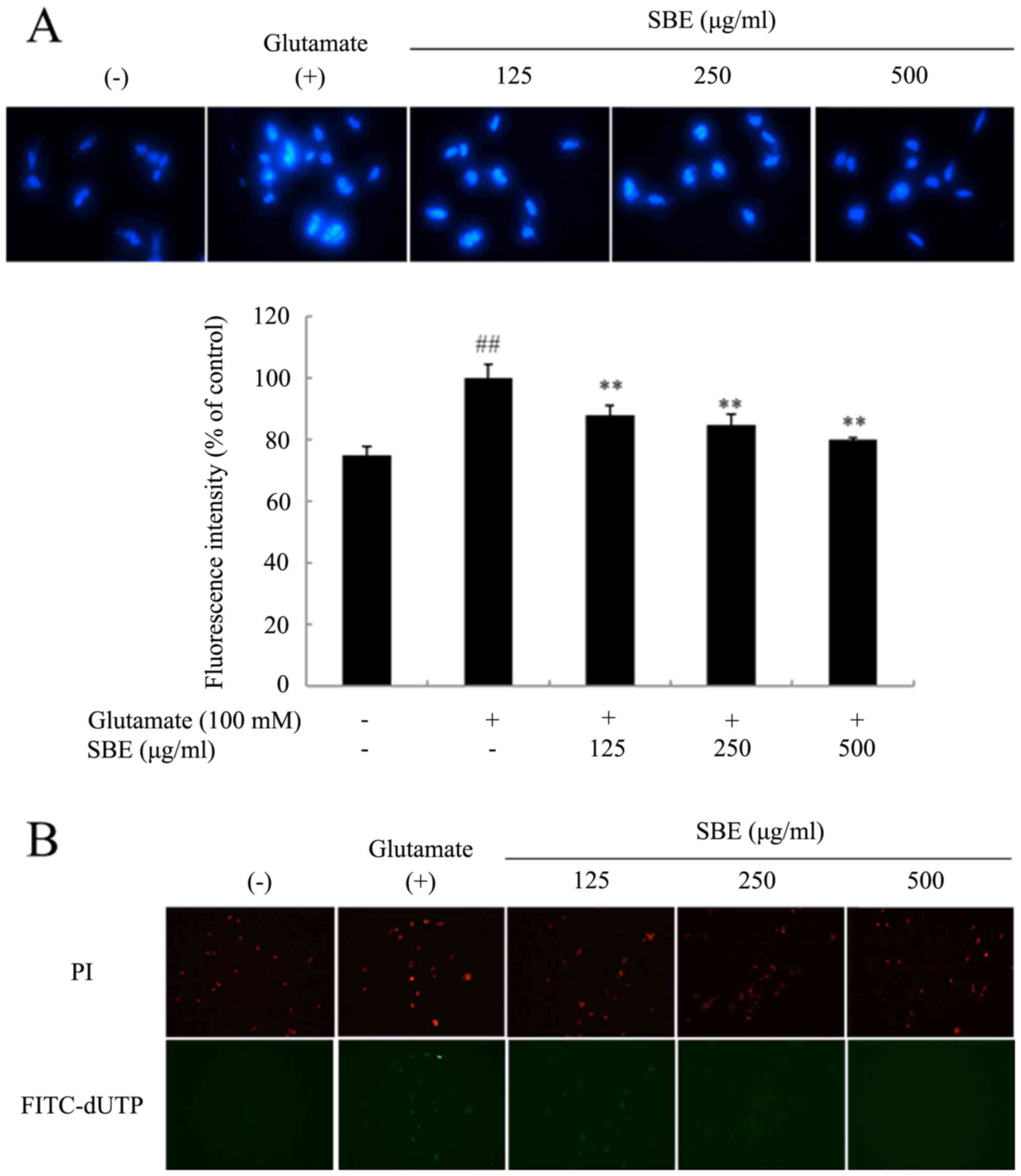
SBE treatment attenuates the
glutamate-induced apoptosis of SH-SY5Y cells
To observe the nuclear morphological changes
following exposure to glutamate, the cells were stained with DAPI.
As shown in Fig. 3A, the control
cells exhibited regular oval shapes, whereas the glutamate-exposed
cells displayed nuclear condensation and DNA fragmentation, and
were unevenly stained. However, the number of DAPI-positive cells
in the SBE-treated groups was significantly lower than that in the
group not treated with SBE, and the glutamate-induced nuclear
morphological changes were attenuated. Furthermore, we examined the
anti-apoptotic effects of SBE on glutamate-induced cell death by
confirming DNA fragmentation by TUNEL assay using PI and
fluorescein isothiocyanate (FITC)-dUTP double staining. PI stains
each cell including normal and apoptotic cells, whereas FITC-dUTP
stains only apoptotic cells with DNA fragmentation. As shown in
Fig. 3B, there were more
FITC-dUTP-stained cells in the glutamate-exposed group than in the
control group. However, the number of stained cells in the
SBE-treated group decreased, indicating that SBE reduced
glutamate-induced DNA fragmentation.
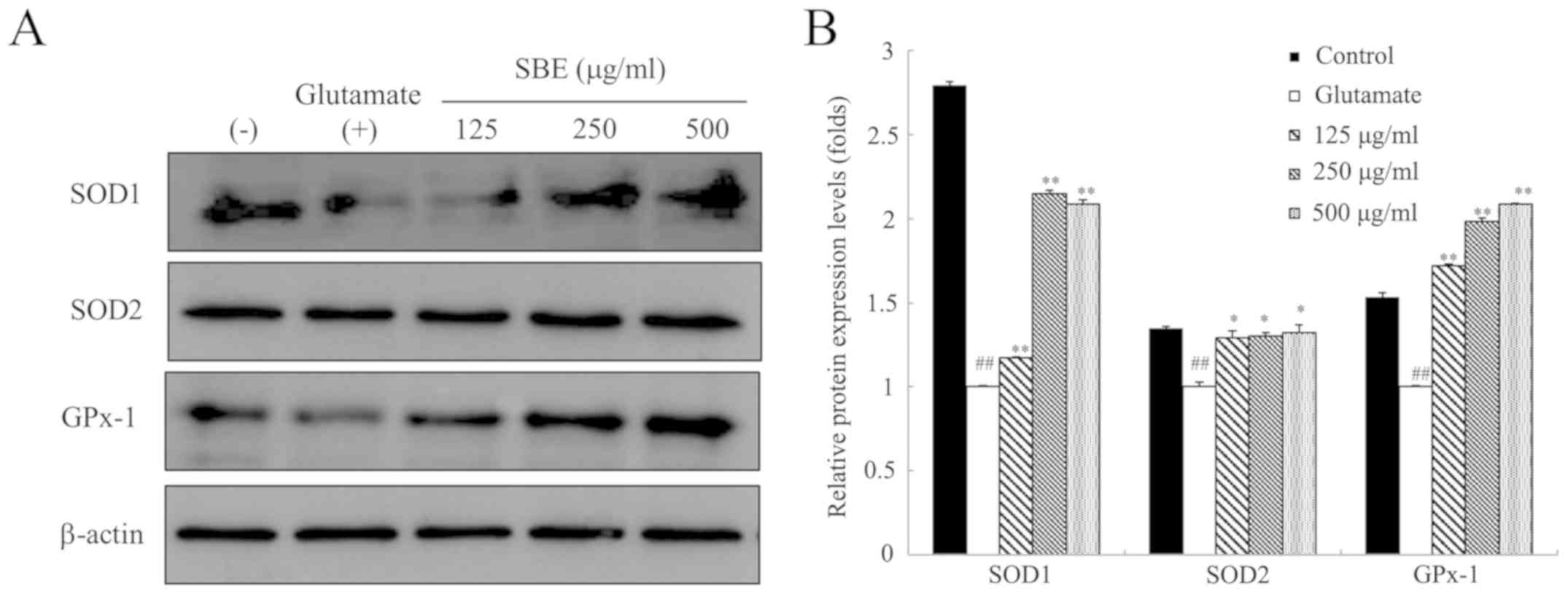
SBE inhibits glutamate-induced oxidative
stress by regulating antioxidant enzyme expression in SH-SY5Y
cells
To examine the antioxidant mechanisms of action of
SBE, we examined the expression levels of antioxidant proteins,
such as SOD1, SOD2, and GPx-1 in the SH-SY5Y cells undergoing
glutamate-induced oxidative stress. As shown in Fig. 4, SOD1, SOD2 and GPx-1 protein
expression decreased following exposure to glutamate compared to
that in the control cells. However, SOD1 protein expression in the
SBE-treated cells dose-dependently increased by 1.17-, 2.14- and
2.08-fold, respectively in the cells treated with 125, 250 and 500
µg/ml SBE, compared to that in the cells exposed to glutamate only
and not treated with SBE. Furthermore, treatment with 125, 250 and
500 µg/ml SBE increased SOD2 protein expression compared to that in
the cells exposed to glutamate only and not treated with SBE. GPx-1
protein expression was significantly increased by SBE treatment
compared to that in the control group. On the whole, these results
demonstrate that SBE reduces glutamate-induced oxidative stress via
the regulation of SOD1, SOD2 and GPx-1 protein expression, and
enhances the expression of antioxidant enzymes.
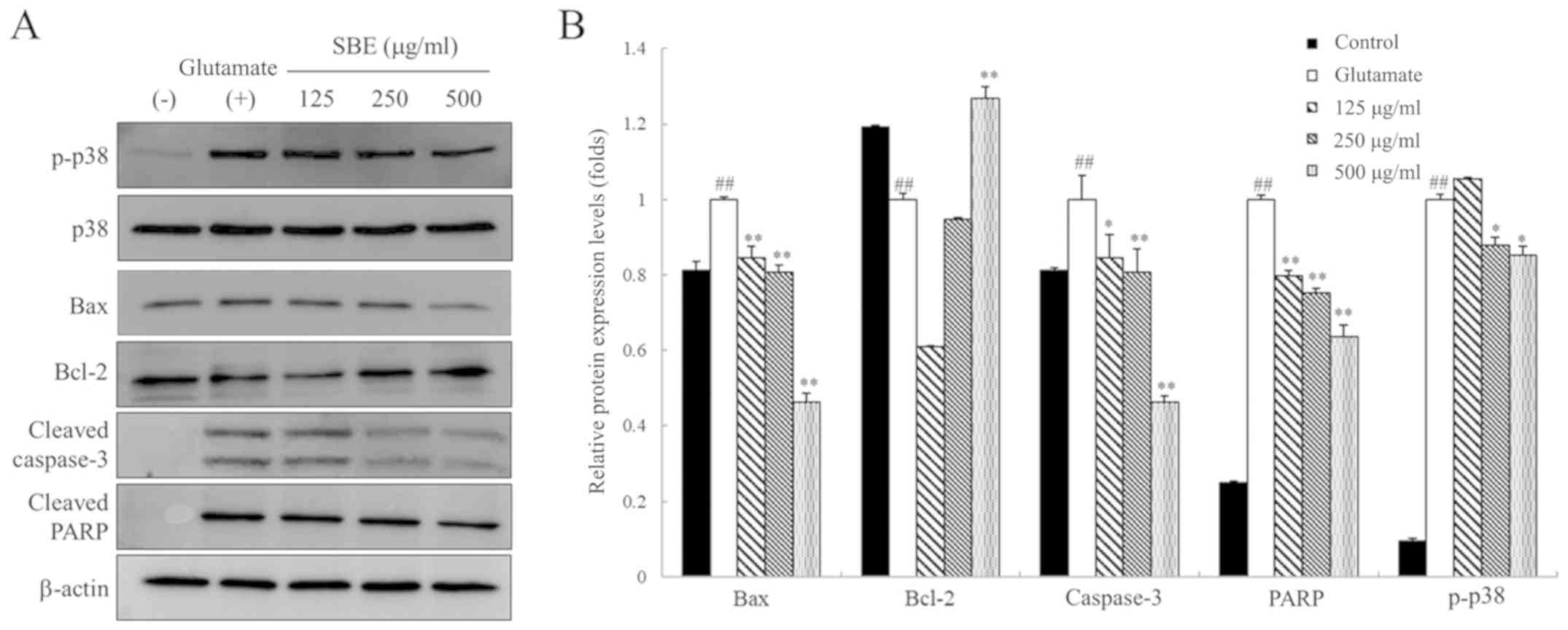
SBE attenuates the glutamate-induced
activation of p38 MAPKs and apoptotic pathways in SH-SY5Y
cells
To examine the anti-apoptotic mechanisms of action
of SBE, we assessed the expression of apoptotic proteins associated
with p38 MAPKs, such as B-cell lymphoma-2 (Bcl-2), Bcl-2-associated
X protein (Bax), cleaved caspase-3 and cleaved poly (adenosine
diphosphate (ADP)-ribose) polymerase (PARP) in glutamate-exposed
SH-SY5Y cells by western blot analysis. As shown in Fig. 5, the levels of phosphorylated
(p)-p38, Bax, cleaved caspase-3 and cleaved PARP were upregulated
in the glutamate-exposed group compared to those in the control
group. By contrast, Bcl-2 expression was downregulated in the
glutamate-exposed cells compared to that in the control cells.
However, treatment with 250 and 500 µg/ml SBE decreased the p-p38
protein expression levels compared to those in the cells exposed to
glutamate only and not treated with SBE. Furthermore, treatment
with SBE decreased Bax protein expression by 11, 12 and 46% in the
cells treated with 125, 250 and 500 µg/ml SBE, and treatment with
500 µg/ml SBE increased Bcl-2 expression compared to that in the
cells exposed to glutamate only and not treated with SBE. SBE
treatment at 125, 250 and 500 µg/ml also dose-dependently decreased
glutamate-induced cleaved caspase-3 protein expression by 19, 50
and 67%, respectively, compared to that in the cells exposed to
glutamate only and not treated with SBE. Cleaved PARP protein
expression was also significantly lower in the SBE-treated group
compared to that in the cells exposed to glutamate only and not
treated with SBE.
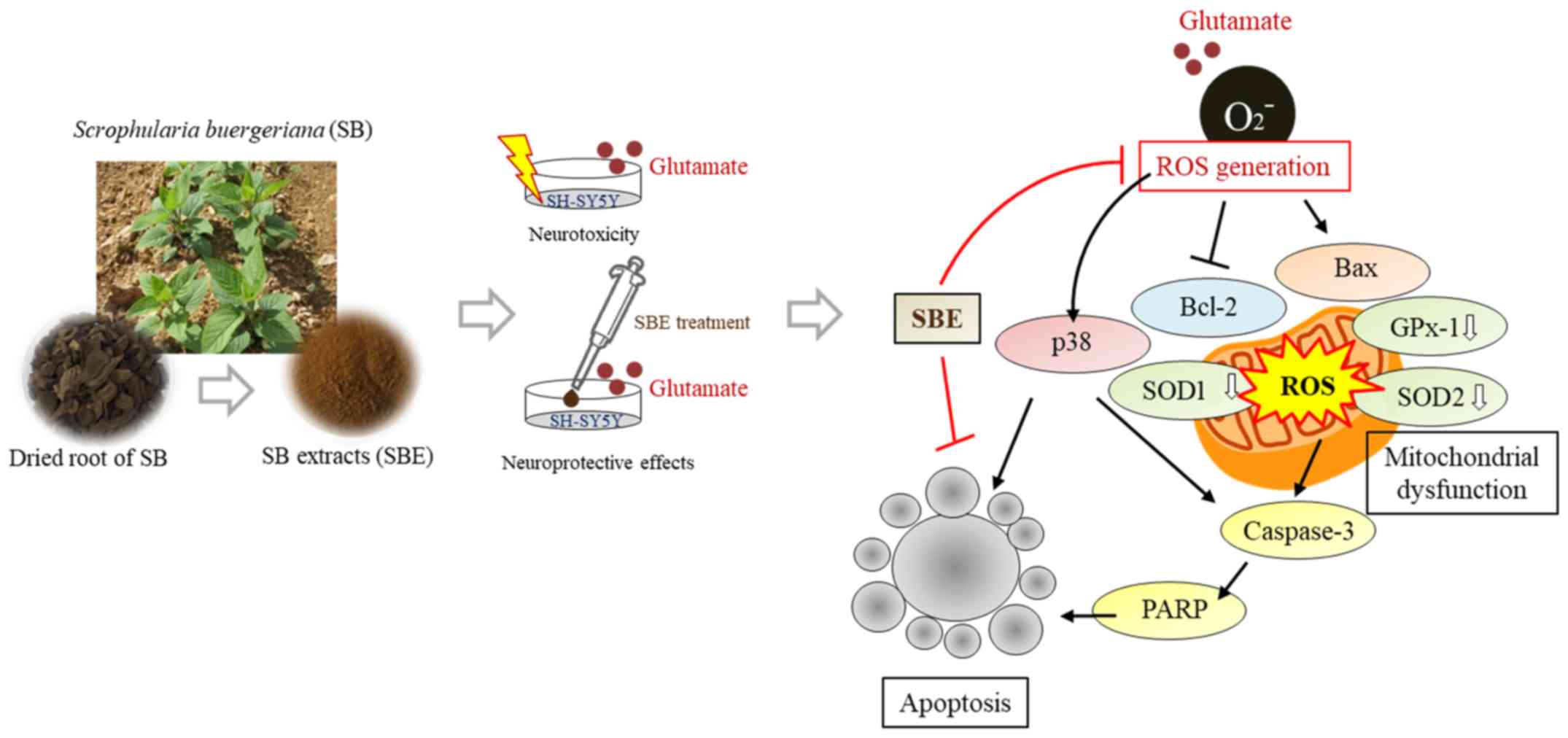
Treatment with SBE significantly decreased p-p38,
Bax, cleaved caspase-3 and cleaved PARP expression, and upregulated
Bcl-2 expression. The relevant mechanisms of action of SBE in this
study are summarized in Fig.
6.
Discussion
Glutamate is an excitatory amino acid
neurotransmitter. It is involved in several physiological processes
and mediates brain function, including cognition, memory and
learning (28,29). However, glutamate induces
oxidative stress and neurotoxicity, and may lead to the development
of various neurodegenerative diseases when present at high
concentrations. Oxidative stress is considered to be a key factor
that leads to neuronal loss and death (30). In glutamate-induced cell death,
glutamate plays a role in the inhibition of glutathione synthesis
and depletion, which induces the excessive production of ROS,
resulting in oxidative stress (31,32). The accumulation of excessive ROS
leads to structural and functional changes in the mitochondria and
activates cell death pathways (32).
SB is a black plant with a strong scent, whose roots
are used in oriental medicine. The roots of SB have been reported
to possess anti-allergic (33),
immune-protective (34) and
antioxidant activities (35), and
compounds isolated from SB roots such as MCA have been reported to
have neuro-protective effects. Therefore, we hypothesized that SBE
may also inhibit the apoptotic process by exerting antioxidant
effects.
SH-SY5Y neuroblastoma cells have been used as an
excitotoxic in vitro model following exposure to high
concentrations of exogenous glutamate. SH-SY5Y cells may be useful
in exploring excitatory amino acid-induced processes as they
express both ionotropic and metabotropic receptors (29,36). Therefore, in this study, we
examined whether treatment with SBE would exert antioxidant effects
and prevent the apoptosis of SH-SY5Y cells induced by
glutamate-induced cytotoxicity.
The present study demonstrated that glutamate
reduced cell viability, whereas SBE treatment significantly
inhibited cell death caused by glutamate as a measurement of cell
viability. Cell viability is used as an indication of cell death.
To examine the neuroprotective effects of SBE, we also assessed
AchE activity. AchE hydrolyzes acetylcholine to choline and acetate
and is an essential enzyme responsible for the inactivation of the
neurotransmitter acetylcholine. It maintains normal function of the
nervous system. However the problems in synaptic integrity, neurite
outgrowth and neurodevelopment are observed, and apoptosis is
induced when AchE concentrations are high (37). Therefore, it is important to
consider low AchE levels as a marker of neurotoxicity. In this
study, we confirmed that glutamate caused cytotoxicity by
increasing AchE expression, although SBE treatment inhibited AchE
activity compared to that in the SH-SY5Y cells exposed to glutamate
only, consistent with the results from a previous study (23). To confirm the antioxidant activity
of SBE, we evaluated the total glutathione contents. Glutathione is
known as a neuronal antioxidant essential for the removal of ROS
and the reduction of oxidative stress. Excessive glutamate
indicates the depletion of glutathione, which is caused by
inhibiting cysteine uptake (38).
In this study, glutamate treatment significantly decrease dthe
glutathione contents. However, SBE treatment upregulated the
glutathione contents compared to those in the cells exposed to
glutamate only and not treated with SBE. Furthermore, we evaluated
the expression of antioxidant enzymes, such as SOD1, SOD2 and
GPx-1, which scavenge free radicals (39). SOD converts superoxide radicals
(O2−) to H2O2, which is converted
to H2O by GPx in the cytosol (40). These results suggest that the SBE
protects against oxidative neuronal damage by exerting antioxidant
effects.
ROS causes DNA damage and nuclear fragmentation, and
leads to abortive apoptosis that can be detected by TUNEL assay and
DAPI staining (41). Therefore,
in this study, we investigated whether SBE inhibits oxidative
stress-induced DNA impairment and cell death. The findings of this
study demonstrated that SBE attenuated glutamate-induced cell death
by inhibiting nuclear condensation, and DNA fragmentation and
degradation.
In this study, we also demonstrated that SBE
significantly inhibited the glutamate-induced activation of
caspase-related proteins. Caspase is known as an apoptotic effector
and is regulated by pro- and antiapoptotic Bcl-2 family proteins.
Glutamate-induced ROS production affects mitochondrial function by
releasing caspase activators (42). Previous studies have reported that
p38 kinases induce apoptosis and increase caspase-3 expression,
thus leading to PARP activation (43,44). In this study, we confirmed that
exposure to glutamate induced an increase in the levels of p-p38,
Bax, cleaved caspase-3 and cleaved PARP, and induced a decrease in
Bcl-2 levels by promoting the apoptotic pathway (45,46). The relevant mechanisms of action
of SBE in this study are summarized in Fig. 6.
Taken together, the results of the present study
demonstrate that SBE exerts neuroprotective effects against
glutamate-induced toxicity in SH-SY5Y cells by removing ROS and
reducing oxidative stress through the increased expression of
antioxidant enzymes and the inhibition of DNA impairment. SBE also
displayed anti-apoptotic activity by downregulating Bax, cleaved
caspase-3 and cleaved PARP expression via the inhibition of p38
MAPKs. Thus, the findings of this study suggest that SBE may be
used as a functional food for attenuating memory impairment through
its antioxidant and anti-apoptotic activities.
Funding
This study was carried out with the support of the
'Cooperative Research Program for Agriculture Science and
Technology Development (Project no. PJ013215012019)', Rural
Development Administration, Republic of Korea.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
HJL analyzed the experimental data and optimized the
SH-SY5Y cell cultures. DM and BNI performed the data processing and
quality control assessment. DAS contributed to the conception and
design of the study, provided critical comments and revised the
manuscript. AT and SHY designed the project, and contributed to the
analysis of the data and finalization of the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Kawasaki H, Morooka T, Shimohama S, Kimura
J, Hirano T, Gotoh Y and Nishida E: Activation and involvement of
p38 mitogen-activated protein kinase in glutamate-induced apoptosis
in rat cerebellar granule cells. J Biol Chem. 272:18518–18521.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nampoothiri M, Reddy ND, John J1, Kumar N,
Kutty Nampurath G and Rao Chamallamudi M: Insulin blocks
glutamate-induced neurotoxicity in differentiated SH-SY5Y neuronal
cells. Behav Neurol. 2014:6741642014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Çomaklı S, Sevim Ç, Kontadakis G, Doğan E,
Taghizadehg- halehjoughi A, Özkaraca M, Aschner M, Nikolouzakis TK
and Tsatsakis A: Acute glufosinate-based herbicide treatment in
rats leads to increased ocular interleukin-1β and c-Fos protein
levels, as well as intraocular pressure. Toxicol Rep. 6:155–160.
2019. View Article : Google Scholar
|
|
4
|
Atlante A, Calissano P, Bobba A,
Giannattasio S, Marra E and Passarella S: Glutamate neurotoxicity,
oxidative stress and mitochondria. FEBS Lett. 497:1–5. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gubandru M, Margina D, Tsitsimpikou C,
Goutzourelas N, Tsarouhas K, Ilie M, Tsatsakis AM and Kouretas D:
Alzheimer's disease treated patients showed different patterns for
oxidative stress and inflammation markers. Food Chem Toxicol.
61:209–214. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Doble A: The role of excitotoxicity in
neurodegenerative disease: Implications for therapy. Pharmacol
Ther. 81:163–221. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hynd MR, Scott HL and Dodd PR:
Glutamate-mediated excitotoxicity and neurodegeneration in
Alzheimer's disease. Neurochem Int. 45:583–595. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Blandini F, Greenamyre JT and Nappi G: The
role of glutamate in the pathophysiology of Parkinson's disease.
Funct Neurol. 11:3–15. 1996.PubMed/NCBI
|
|
9
|
Zeron MM, Chen N, Moshaver A, Lee AT,
Wellington CL, Hayden MR and Raymond LA: Mutant huntingtin enhances
excitotoxic cell death. Mol Cell Neurosci. 17:41–53. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jia J, Zhang L, Shi X, Wu M, Zhou X, Liu X
and Huo T: SOD2 Mediates amifostine-induced protection against
glutamate in PC12 cells. Oxid Med Cell Longev. 2016:42024372016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jin L, Li D, Alesi GN, Fan J, Kang HB, Lu
Z, Boggon TJ, Jin P, Yi H, Wright ER, et al: Glutamate
dehydrogenase 1 signals through antioxidant glutathione peroxidase
1 to regulate redox homeostasis and tumor growth. Cancer Cell.
27:257–270. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Matsuda S, Nakagawa Y, Tsuji A, Kitagishi
Y, Nakanishi A and Murai T: Implications of PI3K/AKT/PTEN signaling
on superoxide dismutases expression and in the pathogenesis of
Alzheimer's disease. Diseases. 6:282018. View Article : Google Scholar
|
|
13
|
Goutzourelas N, Stagos D, Housmekeridou A,
Karapouliou C, Kerasioti E, Aligiannis N, Skaltsounis AL, Spandidos
DA, Tsatsakis AM and Kouretas D: Grape pomace extract exerts
antioxidant effects through an increase in GCS levels and GST
activity in muscle and endothelial cells. Int J Mol Med.
36:433–441. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Maher P and Schubert D: Requirement
for cGMP in nerve cell death caused by glutathione depletion. J
Cell Biol. 139:1317–1324. 1997. View Article : Google Scholar
|
|
15
|
Kouka P, Chatzieffraimidi GA, Raftis G,
Stagos D, Angelis A, Stathopoulos P, Xynos N, Skaltsounis AL,
Tsatsakis AM and Kouretas D: Antioxidant effects of an olive oil
total polyphenolic fraction from a Greek Olea europaea variety in
different cell cultures. Phytomedicine. 47:135–142. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Margină D, Olaru OT, Ilie M, Grădinaru D,
GuȚu C, Voicu S, Dinischiotu A, Spandidos DA and Tsatsakis AM:
Assessment of the potential health benefits of certain total
extracts from Vitis vinifera, Aesculus hyppocastanum and Curcuma
longa. Exp Ther Med. 10:1681–1688. 2015. View Article : Google Scholar
|
|
17
|
Son Y, Cheong YK, Kim NH, Chung HT, Kang
DG and Pae HO: Mitogen-activated protein kinases and reactive
oxygen species: How can ROS activate MAPK pathways? J Signal
Transduct. 2011:7926392011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi DJ, Cho S, Seo JY, Lee HB and Park
YI: Neuroprotective effects of the Phellinus linteus ethyl acetate
extract against H2O2-induced apoptotic cell
death of SK-N-MC cells. Nutr Res. 36:31–43. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cowan KJ and Storey KB: Mitogen-activated
protein kinases: New signaling pathways functioning in cellular
responses to environmental stress. J Exp Biol. 206:1107–1115. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nakagami H, Morishita R, Yamamoto K,
Yoshimura SI, Taniyama Y, Aoki M, Matsubara H, Kim S, Kaneda Y and
Ogihara T: Phosphorylation of p38 mitogen-activated protein kinase
downstream of bax-caspase-3 pathway leads to cell death induced by
high D-glucose in human endothelial cells. Diabetes. 50:1472–1481.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Son Y, Cheong YK, Kim NH, Chung HT, Kang
DG and Pae HO: Mitogen-activated protein kinases and reactive
oxygen species: How can ROS activate MAPK pathway? J Signal
Transduct. 2011:7926392011. View Article : Google Scholar
|
|
22
|
Cheung EC and Slack RS: Emerging role for
ERK as a key regulator of neuronal apoptosis. Sci STKE.
2004:PE452004.PubMed/NCBI
|
|
23
|
Jeong EJ, Ma CJ, Lee KY, Kim SH, Sung SH
and Kim YC: KD-501, a standardized extract of Scrophularia
buergeriana has both cognitive-enhancing and antioxidant activities
in mice given scopolamine. J Ethnopharmacol. 121:98–105. 2009.
View Article : Google Scholar
|
|
24
|
Kim SR, Sung SH, Jang YP, Markelonis GJ,
Oh TH and Kim YC: E-p-methoxycinnamic acid protects cultured
neuronal cells against neurotoxicity induced by glutamate. Br J
Pharmacol. 135:1281–1291. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim SR, Koo KA, Sung SH, Ma CJ, Yoon JS
and Kim YC: Iri from Scrophularia buergeriana attenuate
glutamate-induced neurotoxicity in rat cortical cultures. J
Neurosci Res. 74:948–55. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jeong EJ, Lee KY, Kim SH, Sung SH and Kim
YC: Cognitive-enhancing and antioxidant activities of iri
glycosides from Scrophularia buergeriana in scopolamine-treated
mice. Eur J Pharmacol. 588:78–84. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim SR, Kang SY, Lee KY, Kim SH,
Markelonis GJ, Oh TH and Kim YC: Anti-amnestic activity of
E-p-methoxycinnamic acid from Scrophularia buergeriana. Brain Res
Cogn Brain Res. 17:454–461. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu Y, Li J, Liu P, Chen X, Guo DH, Li QS
and Rahman K: Protection of SH-SY5Y cells from glutamine-induced
apoptosis by 3,6′-disinapoyl sucrose, a bioactive compound isolated
from radix polygala. J Biomed Biotechnol. 2012:1–5. 2012.
|
|
29
|
Al Mamun A, Hashimoto M, Katakura M,
Hossain S and Shido O: Neuroprotective effect of thymoquinone
against glutamate-induced toxicity in SH-SY5Y cells. Curr Top
Nutraceutical Res. 13:143–152. 2015.
|
|
30
|
Song JH, Kang KS and Choi YK: Protective
effect of casuarinin against glutamate-induced apoptosis in HT22
cells through inhibition of oxidative stress-mediated MAPK
phosphorylation. Bioorg Med Chem Lett. 27:5109–5113. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bak DH, Kim HD, Kim YO, Park CG, Han SY
and Kim JJ: Neuroprotective effects of 20(S)-protopanaxadiol
against glutamate-induced mitochondrial dysfunction in PC12 cells.
Int J Mol Med. 37:378–386. 2016. View Article : Google Scholar :
|
|
32
|
Kumari S, Mehta SL and Li PA: Glutamate
induces mitochondrial dynamic imbalance and autophagy activation:
Preventive effects of selenium. PLoS One. 7:e393822012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim JK, Kim YH, Lee HH, Lim SS and Park
KW: Effect of Scrophularia buergeriana extract on the degranulation
of mast cells and ear swelling induced by dinitrofluorobenzene in
mice. Inflammation. 35:183–191. 2012. View Article : Google Scholar
|
|
34
|
Ki SJ, Park JS, Myung NY, Moon PD, Choi
IY, An HJ, Kim NH, Na HJ, Kim DH, Min-cheol K, et al: Scrophularia
buergeriana regulates cytokine production in vitro. Immunopharmacol
Immunotoxicol. 31:246–252. 2009. View Article : Google Scholar
|
|
35
|
Jeong J, Wahyudi LD, Keum YS, Yang H and
Kim JH: E-p-Me thoxycinnamoyl-α-l-rhamnopyranosyl ester, a
phenylpropanoid isolated from Scrophularia buergeriana, increases
nuclear factor erythroid-derived 2-related factor 2 stability by
inhibiting ubiq-uitination in human keratinocytes. Molecules.
23:7682018. View Article : Google Scholar
|
|
36
|
Naarala J, Nykvist P, Tuomala M and
Savolainen K: Excitatory amino acid-induced slow biphasic responses
of free intracellular calcium in human neuroblastoma cells. FEBS
Lett. 330:222–226. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kalafatakis K, Gkanti V, Mackenzie-Gray
Scott CA, Zarros A, Baillie GS and Tsakiris S: Acetylcholinesterase
activity as a neurotoxicity marker within the context of
experimentally-simulated hyperprolinaemia: An in vitro approach. J
Nat Sci Biol Med. 6(Suppl 1): S98–S101. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Engin AB, Engin ED, Golokhvast K,
Spandidos DA and Tsatsakis AM: Glutamate mediated effects of
caffeine and interferon γ on mercury-induced toxicity. Int J Mol
Med. 39:1215–1223. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chan PH: Role of oxidants in ischemic
brain damage. Stroke. 27:1124–1129. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dröge W: Free radicals in the
physiological control of cell function. Physiol Rev. 82:47–95.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fu YB, Ahmed Z, Yang H and Horbach C:
TUNEL assay and DAPI staining revealed few alterations of cellular
morphology in naturally and artificially aged seeds of cultivated
flax. Plants Basel. 7. pp. 342018, View Article : Google Scholar
|
|
42
|
Chen S, Sun M, Zhao X, Yang Z, Liu W, Cao
J, Qiao Y, Luo X and Wen A: Neuroprotection of hydroxysafflor
yellow A in experimental cerebrai ischemia/reperfusion injury via
metabolic inhibition of phenylalanine and mitochondrial biogenesis.
Mol Med Rep. Feb 15–2019.Epub ahead of print. View Article : Google Scholar
|
|
43
|
Juo P, Kuo CJ, Reynolds SE, Konz RF,
Raingeaud J, Davis RJ, Biemann HP and Blenis J: Fas activation of
the p38 mitogen-activated protein kinase signalling pathway
requires ICE/CED-3 family proteases. Mol Cell Biol. 17:24–35. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Thornberry NA and Lazebnik Y: Caspases:
Enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Teng H, Huang Q and Chen L: Inhibition of
cell proliferation and triggering of apoptosis by agrimonolide
through MAP kinase (ERK and p38) pathways in human gastric cancer
AGS cells. Food Funct. 7:4605–4613. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ya H, Wang X, Chen X, Jing Z, Ai X, Liang
Y, Yu Y, Yi Z, Meng X, Meng X, et al: Establishment and evaluation
of a simulated high-altitude hypoxic brain injury model in SD rats.
Mol Med Rep. 19:2758–2766. 2019.
|