Introduction
Epithelial ovarian cancer (EOC) is one of the most
common fatal tumors of the female reproductive tract, and accounts
for 90% of the incidence and the majority of deaths due to this
malignancy (1–3). High mortality rates for ovarian
cancer are mainly attributed to the late stage diagnosis of disease
because almost 60–65% of ovarian cancer cases are first diagnosed
when cancer has already metastasized beyond the confines of the
ovarian tissue. Despite the promise for better outcomes in
recurrent EOC and advanced-stage ovarian cancer through
chemosensitivity or biologic therapies, abdominal recurrence
remains a serious clinical issue due to poor prognosis (4). Therefore, it is important to identify
and develop specific markers to formulate novel therapeutic methods
for advanced and recurrent ovarian cancer.
The receptor of activated C-kinase 1 (RACK1) was
first identified as an anchoring protein for the conventional
protein kinase C (PKC) (5).
Several signal pathways were regulated by RACK1 interaction with
protein kinases C family members, such as PKCα and PKCβII (6–9).
RACK1 has been involved in a series of cellular processes including
regulation of protein translation (10,11),
tissue development (12–14) cellular stress (15) and mammalian circadian clock
(7). Importantly, it was expressed
in some cancer cells and is involved in cancer progression
(16–19). Shi et al found that RACK1
expression was upregulated in non-small cell lung cancer (NSCLC)
samples, silencing RACK1 dramatically inhibited in vivo
tumor growth and metastasis by blocking the sonic hedgehog
signaling pathway (16). It has
been reported that RACK1 downregulates levels of the proapoptotic
protein Fem1b in metastatic, apoptosis-resistant colon cancer
cells, which may promote apoptosis-resistance during progression of
colon cancer (17). Cao et
al found that RACK1 is an independent prognosis-related factor
and promotes breast carcinoma proliferation and invasion/metastasis
in vitro and in vivo (18). These studies implied that RACK1
could promote carcinoma proliferation and invasion/metastasis, and
the down-regulated RACK1 was able to induce cancer cell apoptosis
and inhibited cancer cell proliferation and migration. However, to
our knowledge, the effect of RACK1 expression on tumorigenicity in
ovarian cancer in vivo and in vitro has rarely been
reported (19).
In this study, we investigate the effect of
downregulating the RACK1 expression on cell growth, cell apoptosis,
cell migration and tumor growth in human ovarian cancer in
vivo and in vitro.
Materials and methods
Patients and immunohistochemistry
Paraffin-embedded tissue samples from 50 EOC
patients, diagnosed at the Department of Obstetrics and Gynecology,
The Second Affiliated Hospital of Jilin University (Changchun,
China) from December 2010 to September 2012 were studied.
Clinicopathological information was collected from clinical data of
EOC patients (Table I). Tumors
were staged according to the tumor node metastasis staging system
of the International Union against Cancer (UICC). This study was
approved by the ethics committee of Jilin Hospital and an informed
consent was obtained from all participants.
 | Table I.The correlation of RACK1
overexpression with clinicopathologic features of epithelial
ovarian cancer. |
Table I.
The correlation of RACK1
overexpression with clinicopathologic features of epithelial
ovarian cancer.
| Clinical
factors | Positive | Negative | P-value |
|---|
| Mean age | | | |
| <50
(n=20) | 14 | 6 | 0.944 |
| ≥50 (n=30) | 22 | 8 | |
| Metastasis | | | |
| No (n=26) | 15 | 11 | <0.01 |
| Yes (n=24) | 21 | 3 | |
| Clinical stage | | | |
| I–II (n=35) | 22 | 13 | <0.01 |
| III–IV
(n=15) | 14 | 1 | |
| Tumor size | | | |
| <1.0 cm
(n=28) | 20 | 8 | 0.457 |
| ≥1.0 cm
(n=22) | 16 | 6 | |
| Histological
grade | | | |
| G1 (n=15) | 11 | 4 | 0.896 |
| G2 (n=24) | 18 | 6 | |
| G3 (n=11) | 7 | 4 | |
To detect the expression and localization of RACK1
protein in ovarian tumor tissue, immunohistochemistry was performed
using an SP reagent kit (Tiangen, Beijing, China) according to the
manufacturer’s instructions. The degree of immunoreactivity was
assessed according to Campo et al (20). Immunoreactivity was measured
semiquantitatively using a scale from 0 to 3 score, where score 0
represents no immunostaining, 1 represents fewer than 25% cells
reactive, 2 represents 25–50% of the cells reactive and 3
represents more than 50% of the cells reactive. Values of 0 and 1
were considered to indicate negative staining, and 2 and 3 to
indicate positive staining. Five cases with discordant results were
re-evaluated to obtain agreement.
Cell culture
SKOV3 cells were obtained from the American Type
Culture Collection (ATCC, Manassas, VA, USA) and cultured in
modified RMPI-1640 medium (Sigma, St. Louis, MO, USA) supplemented
with 10% fetal bovine serum (FBS, Gibco, Carlsbad, CA, USA), cells
were maintained in a humidified 37°C incubator with 5%
CO2.
siRNA design and conduct
Templates of small interfering RNA (siRNA) against
RACK1 were designed according to the instructions in the Silencer
siRNA Construction kit (Ambion Inc., Austin, TX, USA) and
synthesized (Invitrogen, Carlsbad, CA, USA). Sequences of the
siRNA1 templates are: sense, 5′-AAGCAAGAAGTTATCAGTACCCCTGTCTC-3′;
and anti-sense, 5′-AAGGTACTGATAACTTCTTGCCCTGTCTC-3′. Sequences for
the scrambled control are: sense, 5′-CACAAGTC
GAAACGTAAATGTCCTGTCTC-3′; and antisense, 5′-ACA
TTTACGTTTCGACTTGTGCCTGTCTC-3′. Sequences of the siRNA2 templates
are: sense, 5′-AACTATGGAATTCCAC AGCGTCCTGTCTC-3′; and antisense,
5′-AAACGCTGTGGA ATTCCATAGCCTGTCTC-3′. The cDNAs corresponding to
the three siRNAs and NC were subcloned into the
replication-deficient, self-inactivating lentiviral expression
vector pGCSIL-RNAi-GFP (Shanghai Gene Chem, Shanghai, China).
Lentivirus containing siRNA was produced according to Coleman et
al as previously described (21). The final titer of siRNA1 siRNA2 and
negative control (NC) were 4×108, 5×108 and
1×109 TU/ml, respectively.
Cell viability analysis
MTT assay was used to determine the effect of
siRNA-RACK1 on the proliferation of cells. SKOV3 cells were
incubated at a concentration of 5×103 cell/well in a
96-well plate, and grown at 37°C, 5% CO2 incubator until
cell adherence. The cells on the culture plate were divided into
groups on the basis of parallel lines after an overnight incubation
in fresh RPMI-1640 containing 0.5% FBS, each group had 4 wells in
one line for each group. After the 24-h attachment period, cells
were treated with 5 μl different siRNA and NC. A total of 20
μl MTT (5 mg/ml) was added and the cells were incubated for
another 4 h at the end of the treatment, 200 μl of DMSO was
added to each well after removing the supernatant. After shaking
the plate for 10 min, cell viability was obtained by measuring the
absorbance at 490 nm wavelength by Enzyme-labeling Instrument
(Bio-Tek ELX800, Winooski, VT, USA), this assay was done in
triplicate. The inhibition rate was calculated according to the
following formula (21):
inhibition rate (%) = [1- (average absorbance of experimental
group/average absorbance of blank control group)] × 100%.
Apoptosis analysis
SKOV3 cells were cultured in 6-well plates in
RPMI-1640 with 10% FBS medium and were treated with different siRNA
for 24, 48 and 72 h. The cover slips were washed three times with
phosphate-buffered saline (PBS) and single cell suspensions were
fixed in 1% PBS. Cells were stained with 100 μg/ml acridine
orange (AO) and 100 μg/ml ethidium bromide (EB) for 1 min.
Then cells were observed under fluorescence microscope. At least
200 cells were counted and the percentage of apoptotic cells was
determined. Triplicates were performed in all experiments, and
experiments were performed five times.
Cell migration assay
The migration assay was performed using a 12-well
Boyden Chamber (Neuro Probe, Gaithersburg, MD, USA) with an 8
μm pore size. About 1×105 cells were seeded into
the upper wells of the Boyden Chamber and incubated for 6 h at 37°C
in medium containing 1% FBS. Medium containing 10% FBS was used as
a chemoattractant in the lower wells. Cells that did not migrate
through the pores of the Boyden Chamber were manually removed with
a rubber instrument. Cells that migrated to the lower side of the
membrane were stained with hematoxylin and eosin and photographed
using an inverted microscope.
Invasion analysis
Cells (1×105/well) were seeded in a Cell
Culture Insert (8 μm pore size; Falcon, BD Biosciences, San
Jose, CA, USA), precoated with 25 μl of 20% Matrigel (2–3
mg/ml protein), and then placed in a 24-well plate (Falcon, BD
Biosciences). After cells had been cultured at 37°C for 40 h, they
were fixed and stained with 0.5% crystal violet. The cells on the
top of the cell culture insert were removed by wiping with a cotton
swab, and cell invasion was observed with an immunofluorescence
microscope by counting the cells that had invaded into the lower of
the cell culture insert. All experiments were performed in
triplicate.
Tumor xenograft assay
All animal experiments were performed in accordance
with institutional guidelines, following a protocol approved by the
Ethics Committees of the Disease Model Research Center, The Second
Hospital of Jilin University. About 6 weeks old female BALB mice
were maintained under specific pathogen-free conditions and
provided with food and water ad libitum. All the animals
were fed with a normal pellet diet one week prior to the
experimentation. In vitro cultured SKOV3 cells were injected
s.c. into the right supra scapula region of the mice. Tumor volume
was calculated by formula: volume = length × width2/2.
When tumors grew to an average volume of 75 mm3, mice
were randomly divided into siRNA1, siRNA2, NC group (n=10 in each)
and by administered siRNA1, siRNA2, and NC plus PBS in a total
volume of 15 μl (5 μl virus plus 10 μl PBS)
one time a week for 21 days, respectively. When control mice
started to succumb to their tumors, the mice in all treatment
groups were euthanized. After the mice had been sacrificed, the
tumors were removed and directly embedded in an optimal cutting
temperature (OCT) compound at −80°C.
Real-time quantitative PCR
Total RNA was isolated from SKOV3 cell lines and
ovarian tissues of EOC patients using TRIzol reagent (Invitrogen)
according to the manufacturer’s instructions. RNA was
reverse-transcribed into cDNA using a Primescript™ RT reagent kit
according to the manufacturer’s instructions. Real-time
quantitative polymerase chain reaction (PCR) was performed with the
SYBR-Green fluorescent dye method, and a Rotor Gene 3000 real-time
PCR apparatus. Human primer sequences were: RACK1 forward,
5′-TCTCTTT CCAGCGTGGCCATTAGA-3′ and reverse, 5′-CCTCGAAGC
TGTAGAGATTCCGACAT-3′; β-actin forward, 5′-GATCAT TGCTCCTCCTGAGC-3′
and reverse, 5′-ACTCCTGCTTGC TGATCCAC-3′. The PCR conditions were
as follows: pre-denaturing at 95°C for 2 min, followed by 45 cycles
of denaturation at 95°C for 10 sec, annealing/extension at 60°C for
20 sec. The amplification specificity was checked by melting curve
analysis. The 2−ΔΔCT method was used to calculate the
relative abundance of target gene expression generated by
Rotor-Gene Real-Time Analysis Software 6.1.81. For each cDNA, the
target gene mRNA level was normalized to β-actin mRNA level. The
experiments were performed three times.
Western blot analysis
Cultured cells were washed twice with PBS and lysed
in radioimmune precipitation assay buffer for 30 min on ice. Cell
lysates were clarified by centrifugation (10,000 × g, 15 min), and
protein concentrations were determined using the Bradford reagent
(Sigma). Lysates were separated on 8 or 15% SDS-PAGE; proteins were
transferred to Immobilon membrane (Millipore, Bedford, MA, USA)
immunoblotted with specific primary antibodies and incubated with
corresponding horseradish peroxidase-conjugated secondary antibody.
The primary antibodies used in the western blots were: antibodies
against Akt, phosphorylated (p)-Akt, MAPK, p-MAPK, anti-RACK1
antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA). All
immunoblots were visualized by enhanced chemiluminescence (Pierce,
Rockford, IL, USA).
Statistical analysis
The data are expressed as mean ± SEM. Statistical
analysis between two samples was performed using Student’s t-test.
Statistical comparison of more than two groups was performed using
one-way ANOVA followed by a Tukey’s post hoc test and considered
significant at *P<0.05 or **P<0.01.
Results
RACK1 is upregulated in EOC and
correlates with clinical features of EOC patients
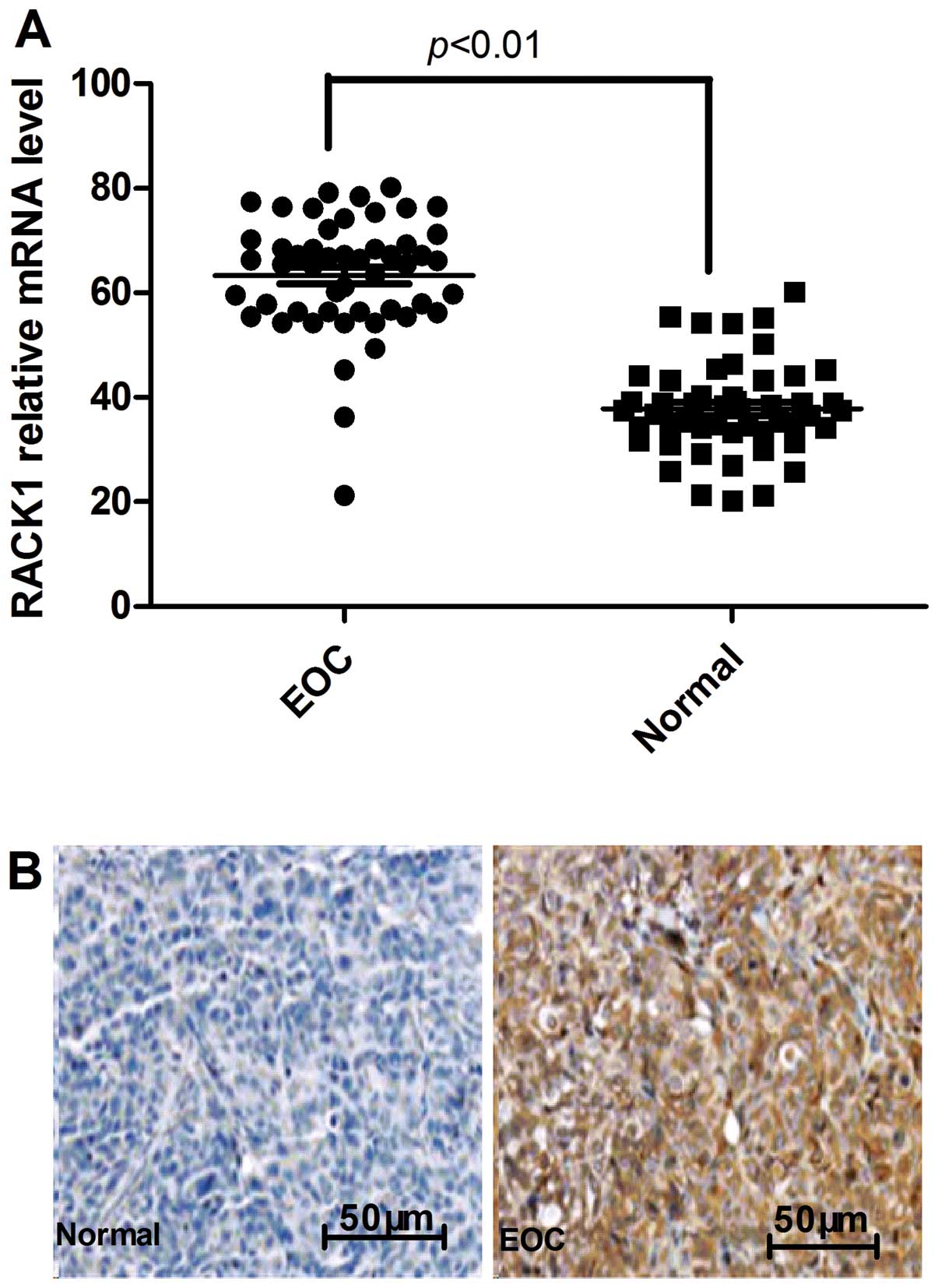
To identify the potential roles of RACK1 in the
development and progression of EOC, we assessed its expression
level by real-time polymerase chain reaction (PCR) in 50 pairs of
matched ovarian tissue samples. Expression levels of RACK1 were
significantly higher in EOC tumors compared with their normal
ovarian counterparts (Fig. 1A).
Furthermore, elevated levels of RACK1 protein were found in EOC
tumors compared with the paired normal tissues from the same
patients as shown by immunochemical staining (Fig. 1B). Further analysis revealed that
the upregulation of RACK1 protein in EOC samples was positively
correlated with both lymph node metastasis and tumor TNM stage
(P<0.01, Table I). No
correlation occurred between RACK1 protein levels, age, tumor size
and histologic grade. These data suggested that RACK1 expression
correlates with advanced EOC.
siRACK1 transformed into lentiviral
vector efficiency assay
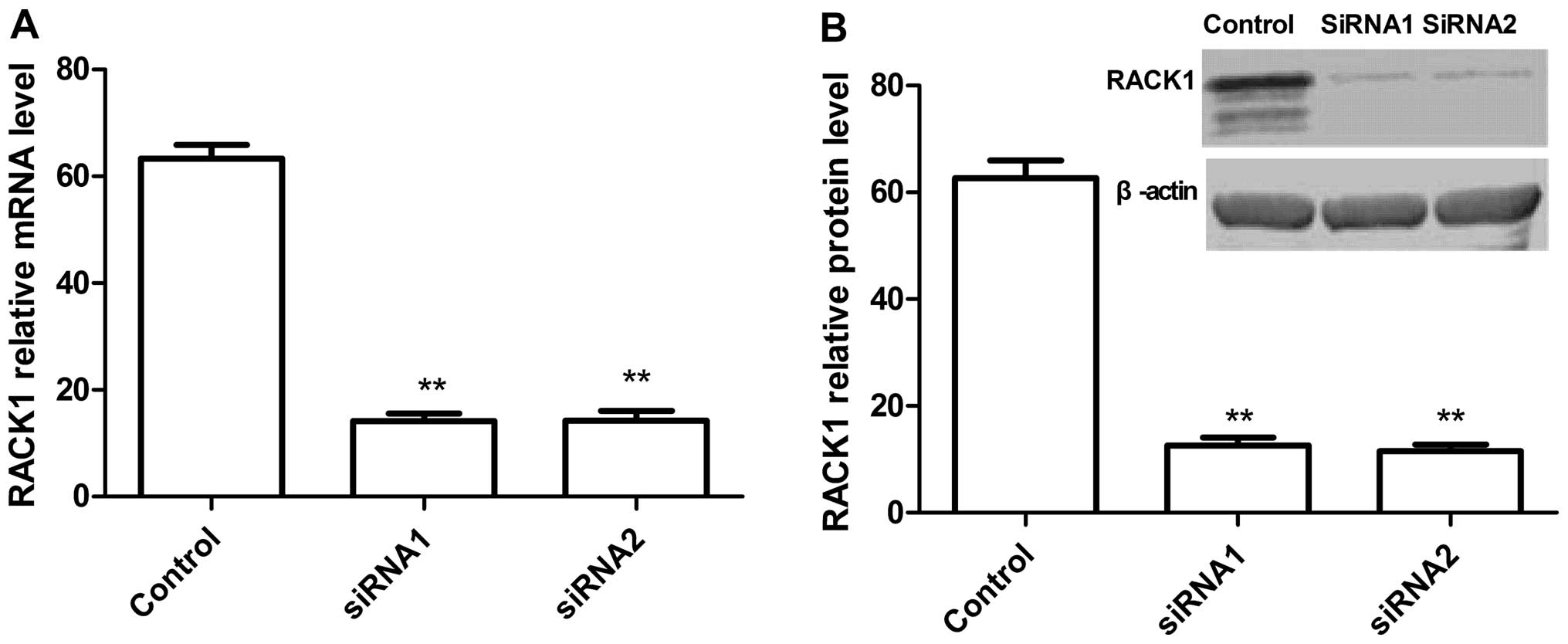
To understand the function of RACK1 in EOC, we used
the RNAi-mediated knockdown lentiviral vector to decrease the basal
level of RACK1 (knockdown of RACK1) in SKOV3 cells by real-time
RT-PCR and western blot analysis. Real-time RT-PCR and western blot
analysis results showed that two independent target sequences
siRNA1 and siRNA2 markedly decreased the expression of RACK1
compared with the control sequence (Fig. 2). These results demonstrated that
siRNA targeting RACK1 significantly silenced RACK1 expression in
vitro.
Silencing RACK1 in ovarian cancer cells
reduces proliferation, migration and invasion in vitro
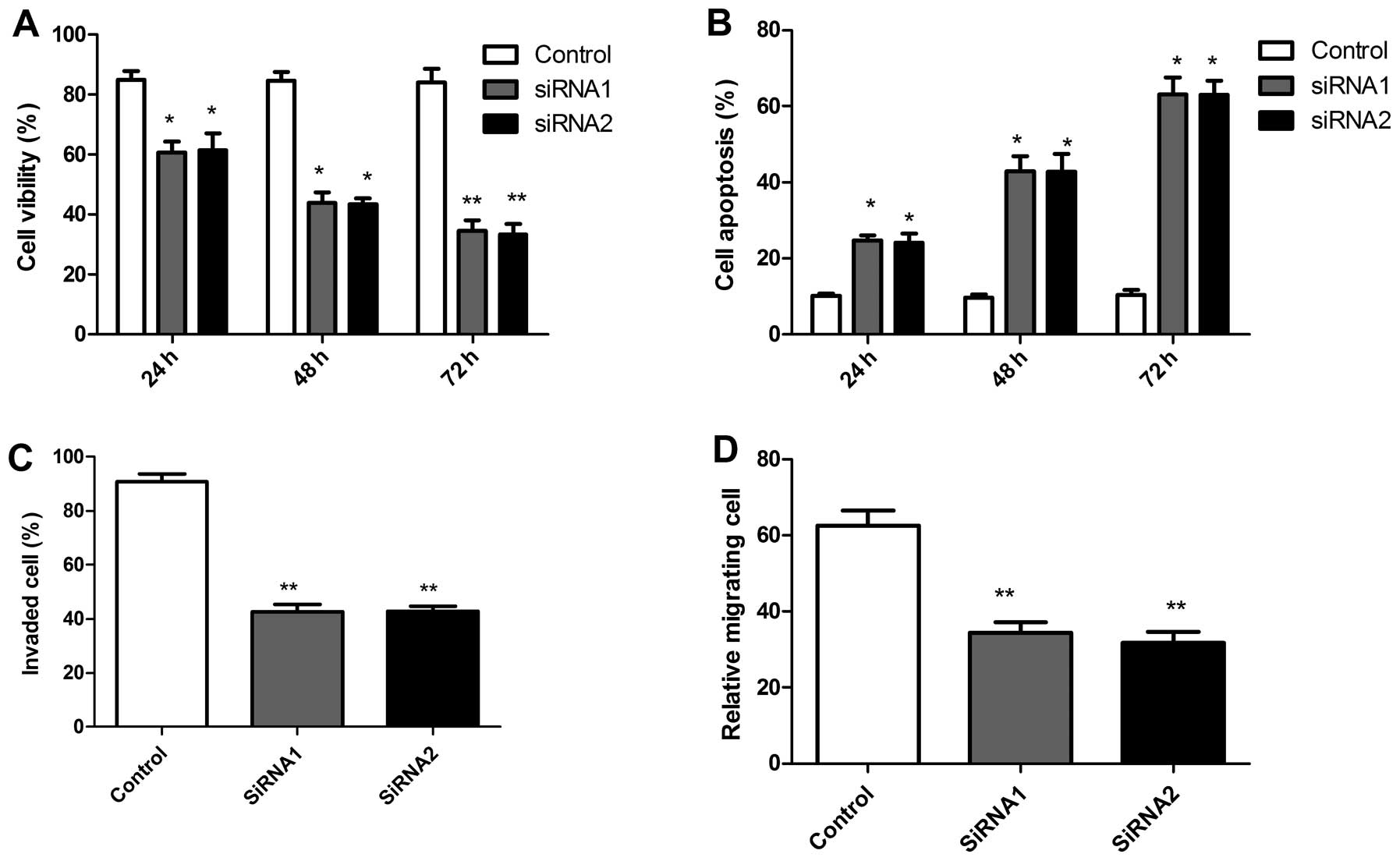
Using these two siRNAs, we examined the effects of
RACK1 silencing on tumor cell growth in vitro. The
anti-proliferative effect of RACK1 silencing on SKOV3 tumor cell
were examined using MTT assays. It was found that it significantly
inhibited the proliferation of SKOV3 tumor cells compared with
control cells (scrambled siRNA) (Fig.
3A). Conversely, RACK1 silencing significantly improved
apoptosis of SKOV3 tumor cells compared with control
time-dependently, as determined using the AO staining assay
(Fig. 3B). Furthermore, the
effects of silencing RACK1 on ovarian tumor cell migration and
invasion were assessed. As shown in Fig. 3C, silencing RACK1 reduced migration
capacity in SKOV3 tumor cells. In addition, cell invasion ability
was significantly decreased, when assessed after 48 h by the
modified Boyden chamber assays (Fig.
3D). Together, these data indicate that RACK1 plays an
important regulatory role in tumor cell growth and migration.
Silencing RACK1 suppresses tumor growth
in vivo
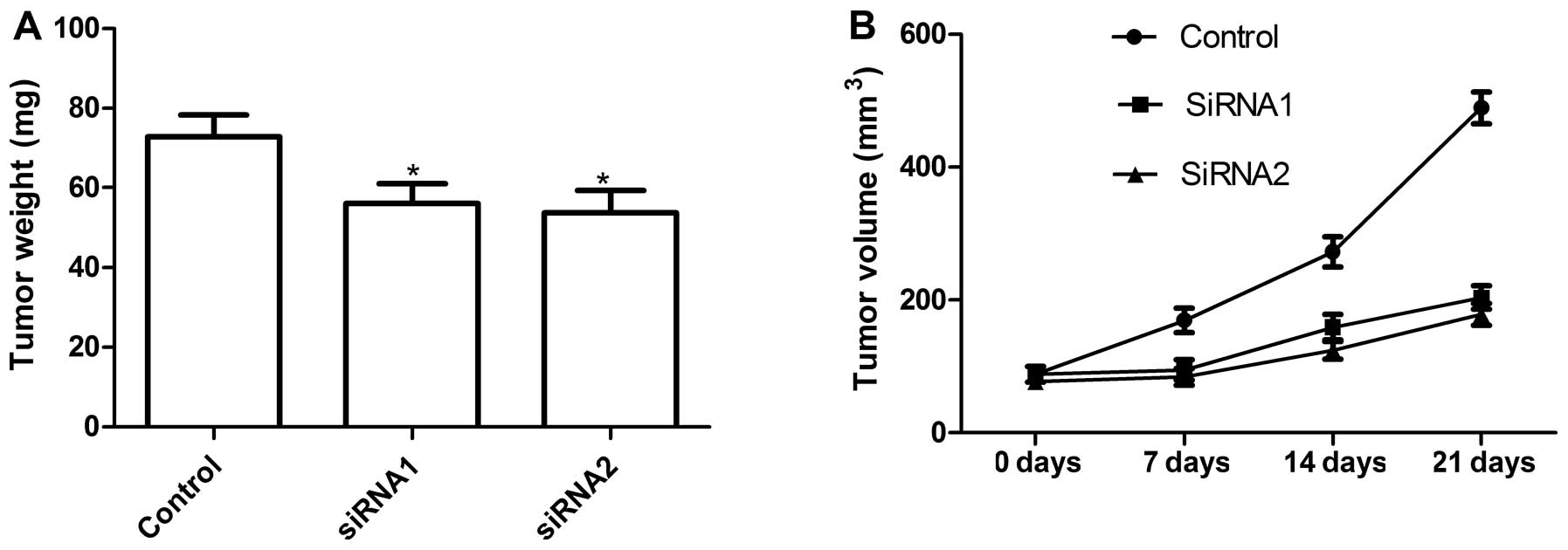
To obtain definitive evidence that RACK1 is required
for ovarian tumor growth in vivo, we conducted a xenograft
assay by injecting the control (si-scrambled) and RACK1-silenced
tumor cells subcutaneously into mice and compared the growth rate
to solid tumors. Mice were sacrificed and taken out tumor tissue 21
days after treatment. Then tumor weight of the animals was
measured. The tumor weight of siRNA group was lower than those of
control group (Fig. 4A). We found
that tumor volume after RACK1 silencing was significantly slower in
SKOV3 tumor cells compared with control cells (Fig. 4B). These results indicate that
suppression of RACK1 expression in ovarian tumor cells markedly
suppresses their tumorigenicity in mice.
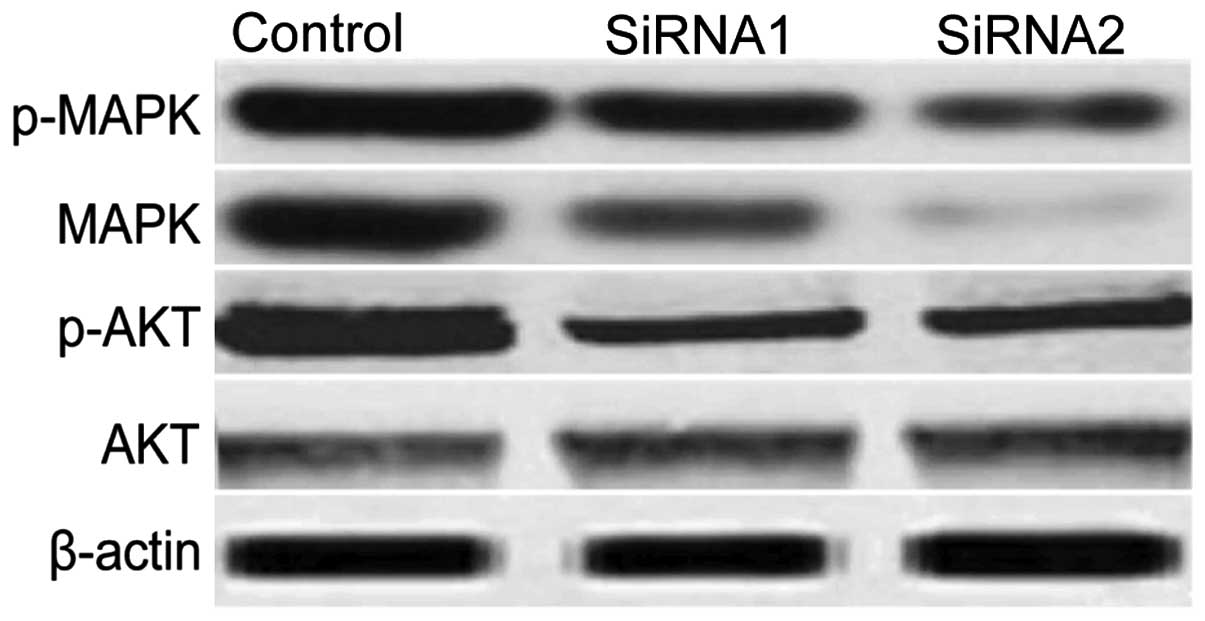
RACK1 silencing inhibites the
phosphorylation of Akt and MAPK in vitro
To clarify the molecular mechanisms involved due to
downregulation of RACK1 inhibition the growth of human tumor
xenografts, in the present study, we focused on the effects of
RACK1 silencing on the activation of MAPK, Akt, which participate
in the main intracellular signaling required for cell proliferation
and survival. As shown in Fig. 5,
compared with control siRNAs, RACK1 silencing led to a significant
suppression of Akt and MAPK activation. These results indicate that
RACK1 silencing inhibits tumor cell growth, to some extent, by
suppressing MAPK and Akt-mediated signaling.
Discussion
RACK1, as a crucial scaffold protein, was originally
cloned as an anchoring protein for PKC; it plays a critical role in
stabilizing the active form of PKC and in permitting its
translocation to different sites within a cell (22,23).
A large number of literature reveals that RACK1 can associate with
various signaling molecules and is involved in the regulation of
various cell functions (24–27).
Increased RACK1 expression has been reported in lung, colon, breast
and hepatocellular carcinomas and human melanomas (28–30).
Zhang et al found that increased RACK1 plays an important
role in IR/IGF-1R-mediated promotion of anchorage-independent
growth of certain ovarian cancer cells (19). However, this study did not focus on
RACK1 expression in EOC patients. To our knowledge, in the present
study, we first found that RACK1 was elevated in most EOC compared
to normal ovarian tissue, and its expression level correlated with
key pathological characteristics including clinical stage and
metastasis. Our findings also show that downregulation of RACK1
results in the inhibition of proliferation migration for SKVO3
cells in vitro, and suppression of solid tumor growth in
vivo. These results provide evidence that RACK1 is required for
tumor growth and that it may be a diagnostic marker.
Previous studies showed that several signal pathways
were regulated by RACK1 interaction with protein kinases C family
members, such as PKC, Src and IGF-1R (19,22,26).
It is widely accepted, that aberrant activation of several PKC
isozymes leads to tumorigenesis; thus, as an important stabilizer
of PKC activity, RACK1 may play a pivotal role in regulating PKC
signaling-related tumor progression (22,23,31).
Berns et al showed that RACK1 is upregulated in vascular
endothelial cells during angiogenesis and may contribute to tumor
growth and spreading (30). Wang
et al found that RACK1 silencing significantly attenuated
tumor-associated angiogenesis by inhibiting the expression of two
critical angiogenic factors (17).
Furthermore, Keily et al showed that the proliferation of
MCF-7 cells was enhanced by overexpression of RACK1, whereas
IGF-1-mediated protection from etoposide-dependent death was
greatly reduced (32). Two other
groups also reported that knockdown of RACK1 suppressed IGF-1R and
androgen receptor-dependent tumor cell growth (33,34).
Zhang et al further showed that RACK1 acts as an adaptor
protein to facilitate the association of STAT3 with IR/IGF-1R and
subsequent phosphorylation and activation of STAT3 specifically
(19). Shi et al found that
RACK1 activated the sonic hedgehog signaling pathway by interacting
with and activating smoothened to mediate Gli1-dependent
transcription in NSCLC cells (16). These studies show that RACK1 is
simultaneously involved in widespread signaling pathways owing to
its association with PKC, Src family members, and other multiple
pathways related to these proteins appear to be activated in the
process of tumor growth. Thus, RACK1 is implicated in a complicated
signaling network rather than a simple pathway. In the present
study, we focused on the effect of RACK1 on the activation of
central players in growth signaling, such as Akt, MAPK, which are
often constitutively activated in a variety of cancers and
participated in the main intracellular signaling required for cell
proliferation and survival (35,36).
We found that RACK1 silencing significantly inhibited the
phosphorylation of Akt and MAPK to some extent in SKVO3 cells.
Which implied that the suppression of Akt- or MAPK-related pathways
contributes, at least in part, to the inhibition of tumor
growth.
A series of experiments (in vitro and in
vivo) were designed to investigate the characteristics of RACK1
and to evaluate its role in ovarian tumorigenesis. Our results show
that the silencing of RACK1 induced SKOV3 tumor cell apoptosis,
which agrees with the downregulation of RACK1 inducing cell
apoptosis in NSCLC (16). However,
in contrast with a previous report that overexpression of RACK1
induced apoptosis of HT-29 colon carcinoma cells, partly by
inhibiting Src activity (37).
Moreover, recent studies showed that RACK1 knockdown inhibited cell
growth but did not affect cell apoptosis (17,18).
These above studies suggested that RACK1 has opposing effects on
cell apoptosis of different types of tumor cells, and the
discrepancy may arise from differences the cell types used or in
the experimental conditions.
In conclusion, to our knowledge, this is the first
full-scale report concerning the association of RACK1 with ovarian
carcinoma. The present study demonstrates that RACK1 was elevated
in most EOC and its expression level correlated with key
pathological characteristics including clinical stage and
metastasis and it plays a critical role in tumor growth by
affecting cell proliferation and tumor growth in vitro and
in vivo. Based on the multiple functions of RACK1 in tumor
growth, it may be considered a marker for diagnosis and a potential
anticancer therapeutic target.
References
|
1.
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2.
|
Bast RC Jr, Hennessy B and Mills GB: The
biology of ovarian cancer: new opportunities for translation. Nat
Rev Cancer. 9:415–428. 1996. View
Article : Google Scholar
|
|
3.
|
Bowtell DD: The genesis and evolution of
high-grade serous ovarian cancer. Nat Rev Cancer. 10:803–808. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Vaughan S, Coward JI, Bast RC Jr, Berchuck
A, Berek JS, Brenton JD, Coukos G, Crum CC, Drapkin R,
Etemadmoghadam D, Friedlander M, Gabra H, Kaye SB, Lord CJ, Lengyel
E, Levine DA, McNeish IA, Menon U, Mills GB, Nephew KP, Oza AM,
Sood AK, Stronach EA, Walczak H, Bowtell DD and Balkwill FR:
Rethinking ovarian cancer: recommendations for improving outcomes.
Nat Rev Cancer. 11:719–725. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Csukai M and Mochly-Rosen D: Pharmacologic
modulation of protein kinase C isozymes: the role of RACKs and
subcellular localisation. Pharmacol Res. 39:253–259. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Calura E, Cagnin S, Raffaello A, Laveder
P, Lanfranchi G and Romualdi C: Meta-analysis of expression
signatures of muscle atrophy. Gene interaction networks in early
and late stages. BMC Genomics. 9:6302008. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Robles MS, Boyault C, Knutti D,
Padmanabhan K and Weitz CJ: Identification of RACK1 and protein
kinase C as integral components of the mammalian circadian clock.
Science. 327:463–466. 2010. View Article : Google Scholar
|
|
8.
|
Li CL, Toda K, Saibara T, Zhang T, Ono M,
Iwasaki S, Maeda T, Okada T, Hayashi Y, Enzan H, Shizuta Y and
Onishi S: Estrogen deficiency results in enhanced expression of
smoothened of the Hedgehog signaling in the thymus and affects
thymocyte development. Int Immunopharmacol. 2:823–833.
2003.PubMed/NCBI
|
|
9.
|
Bible KC, Suman VJ, Molina JR, Smallridge
RC, Maples WJ, Menefee ME, Rubin J, Sideras K, Morris JC III,
McIver B, Burton JK, Webster KP, Bieber C, Traynor AM, Flynn PJ,
Goh BC, Tang H, Ivy SP and Erlichman C: Efficacy of pazopanib in
progressive, radioiodine-refractory, metastatic differentiated
thyroid cancers. Results of a phase 2 consortium study Lancet
Oncol. 11:962–972. 2010.PubMed/NCBI
|
|
10.
|
Toftgard R: Hedgehog signaling in cancer.
Cell Mol Life Sci. 57:1720–1731. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Gialmanidis IP, Bravou V, Amanetopoulou
SG, Varakis J, Kourea H and Papadaki H: Overexpression of hedgehog
pathway molecules and FOXM1 in non-small-cell lung carcinomas. Lung
Cancer. 66:64–74. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Kadrmas JL, Smith MA and Pronovost SM:
Characterization of RACK1 function in Drosophila
development. Dev Dyn. 236:2207–2215. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Mor I, Sklan EH, Podoly E, Pick M,
Kirschner M, Yogev L, Bar-Sheshet Itach S, Schreiber L, Geyer B,
Mor T, Grisaru D and Soreq H: Acetylcholinesterase-R increases germ
cell apoptosis but enhances sperm motility. J Cell Mol Med.
12:479–495. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Battaini F, Pascale A, Paoletti R and
Govoni S: The role of anchoring protein RACK1 in PKC activation in
the ageing rat brain. Trends Neurosci. 20:410–415. 1997. View Article : Google Scholar
|
|
15.
|
Qiu Y, Mao T, Zhang Y, Shao M, You J, Ding
Q, Chen Y, Wu D, Xie D, Lin X, Gao X, Kaufman RJ, Li W and Liu Y: A
crucial role for RACK1 in the regulation of glucose-stimulated IRE1
activation in pancreatic cells. Sci Signal. 3:ra72010.PubMed/NCBI
|
|
16.
|
Shi S, Deng YZ, Zhao JS, Ji XD, Shi J,
Feng YX, Li G, Li JJ, Zhu D, Koeffler HP, Zhao Y and Xie D: Sonic
Hedgehog signaling pathway cancer tumorigenicity through activating
RACK1 promotes non-small-cell lung. J Biol Chem. 287:7845–7858.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Wang F, Osawa T, Tsuchida R, Yuasa Y and
Shibuya M: Downregulation of receptor for activated C-kinase 1
(RACK1) suppresses tumor growth by inhibiting tumor cell
proliferation and tumor-associated angiogenesis. Cancer Sci.
102:2007–2013. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Cao XX, Xu JD, Xu JW, Liu XL, Cheng YY,
Wang WJ, Li QQ, Chen Q, Xu ZD and Liu XP: RACK1 promotes breast
carcinoma proliferation and invasion/metastasis in vitro and in
vivo. Breast Cancer Res Treat. 123:375–386. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Zhang W, Zong CS, Hermanto U,
Lopez-Bergami P, Ronai Z and Wang LH: RACK1 recruits STAT3
specifically to insulin and insulin-like growth factor 1 receptors
for activation, which is important for regulating anchorage
independent growth. Mol Cell Biol. 26:413–424. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Campo E, Merino MJ, Liotta L, Neumann R
and Stetler-Stevenson W: Distribution of 72-kd type IV collagenase
in nonneoplastic and neoplastic thyroid tissue. Hum Pathol.
23:1395–1401. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Coleman JE, Huentelman MJ, Kasparov S,
Metcalfe BL, Paton JF, Katovich MJ, Semple-Rowland SL and Raizada
MK: Efficient large-scale production and concentration of
HIV-1-based lentiviral vectors for use in vivo. Physiol Genomics.
12:221–228. 2003.PubMed/NCBI
|
|
22.
|
Ron D, Chen CH, Caldwell J, Jamieson L,
Orr E and Mochly-Rosen D: Cloning of an intracellular receptor for
protein kinase C: a homolog of the beta subunit of G proteins. Proc
Natl Acad Sci USA. 91:839–843. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Ron D and Mochly-Rosen D: Agonists and
antagonists of protein kinase C function, derived from its binding
proteins. J Biol Chem. 269:21395–21398. 1994.PubMed/NCBI
|
|
24.
|
Liliental J and Chang DD: Rack1: a
receptor for activated protein kinase C, interacts with integrin
beta subunit. J Biol Chem. 273:2379–2383. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Wang F, Yamauchi M, Muramatsu M, Osawa T,
Tsuchida R and Shibuya M: RACK1 regulates VEGF/Flt1-mediated cell
migration via activation of a PI3K/Akt pathway. J Biol Chem.
286:9097–9106. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Wang F, Yamauchi M, Muramatsu M, Osawa T,
Tsuchida R and Shibuya M: The interaction of Src and RACK1 is
enhanced by activation of protein kinase C and tyrosine
phosphorylation of RACK1. J Biol Chem. 276:20346–20356. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Besson A, Wilson TL and Yong VW: The
anchoring protein RACK1 links protein kinase C epsilon to integrin
beta chains. Requirements for adhesion and motility. J Biol Chem.
277:22073–22084. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Bourd-Boittin K, LePabic H and Bonnier D:
RACK1, a new ADAM12 interacting protein. Contribution to liver
fibrogenesis J Biol Chem. 283:26000–26009. 2008.PubMed/NCBI
|
|
29.
|
Egidy G, Jule S and Bosse P: Transcription
analysis in the MeLiM swine model identifies RACK1 as a potential
marker of malignancy for human melanocytic proliferation. Mol
Cancer. 7:342008. View Article : Google Scholar
|
|
30.
|
Berns H, Humar R, Hengerer B, Kiefer FN
and Battegay EJ: RACK1 is upregulated in angiogenesis and human
carcinomas. FASEB J. 14:2549–2558. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Schechtman D and Mochly-Rosen D: Adaptor
proteins in protein kinase C mediated signal transduction.
Oncogene. 20:6339–6347. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Kiely PA, Sant A and O’Connor R: RACK1 is
an insulin-like growth factor 1 (IGF-1) receptor-interacting
protein that can regulate IGF-1-mediated Akt activation and
protection from cell death. J Biol Chem. 277:22581–22589. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Kraus S, Gioeli D, Vomastek T, Gordon V
and Weber MJ: Receptor for activated C kinase 1 (RACK1) and Src
regulate the tyrosine phosphorylation and function of the androgen
receptor. Cancer Res. 66:11047–11054. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Hermanto U, Zong CS, Li W and Wang LH:
RACK1, an insulin-like growth factor I (IGF-I) receptor-interacting
protein, modulates IGF-I-dependent integrin signaling and promotes
cell spreading and contact with extracellular matrix. Mol Cell
Biol. 22:2345–2365. 2002. View Article : Google Scholar
|
|
35.
|
Falasca M: PI3K/Akt signalling pathway
specific inhibitors: a novel strategy to sensitize cancer cells to
anti-cancer drugs. Curr Pharm Des. 16:1410–1416. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Katz M, Amit I and Yarden Y: Regulation of
MAPKs by growth factors and receptor tyrosine kinases. Biochim
Biophys Acta. 1773:1161–1176. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Mamidipudi V, Dhillon NK, Parman T, Miller
LD, Lee KC and Cartwright CA: RACK1 inhibits colonic cell growth by
regulating Src activity at cell cycle checkpoints. Oncogene.
26:2914–2924. 2007. View Article : Google Scholar : PubMed/NCBI
|