Contents
Introduction
Genetic and epigenetic change of AJAP1
Continuing studies on AJAP1
Relationship between AJAP1 and tumors
Closing remarks
Introduction
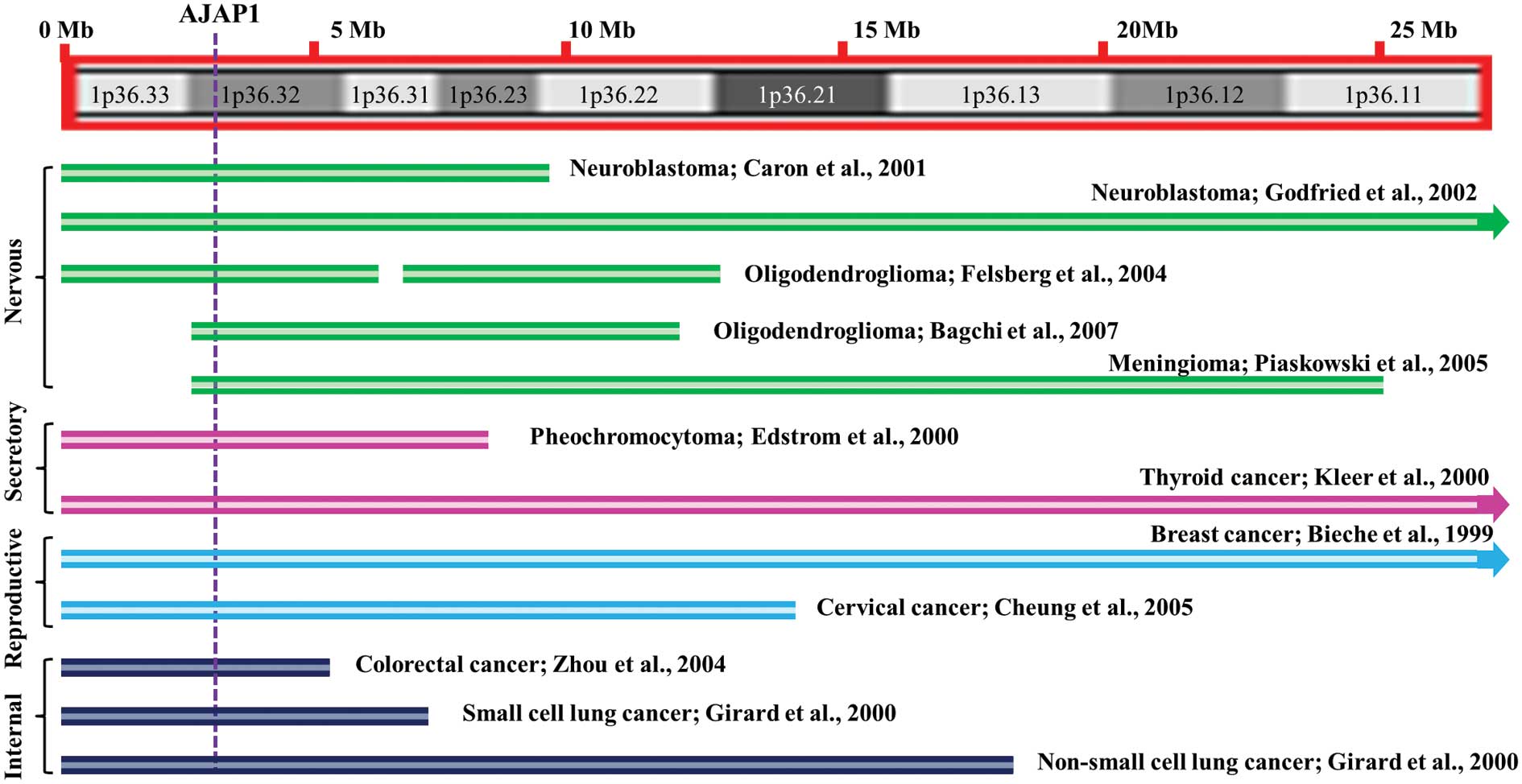
In a broad range of human cancers 1p36 has been a
mutational hotspot, including those of nervous, secretory,
reproductive and internal origin (Fig.
1) (1–11). These studies strongly suggest that
the loss of tumor suppressor activity maps to this genomic region
during tumorigenesis. The large 1p36 deletions led others to
propose that more than one tumor suppressor-related gene may reside
in this region (4,12). Although the quest for the 1p36
tumor suppressor has led to some exciting candidates, such as CHD5,
CAMTA1, KIF1B, CASZ1 and miR-34a which have tumor suppressor-like
capabilities in specific tumor contexts (13–23),
none can provide convincing evidence that their encoded products
offer protection from cancer. Hence, the search for the 1p36 tumor
suppressor continues.
Adherens junctional associated protein-1 (AJAP1;
also known as Shrew1) was initially discovered as a novel
trans-membrane protein of adherent junctions in epithelial cells
(24). Recent investigations have
suggested the AJAP1 gene as a promising tumor suppressor candidate
gene located at 1p36.32 (13,24).
Genetic and epigenetic change of AJAP1
Significant data supports the role of AJAP1 loss in
various tumors of the central nervous system. A single region
within 1p36.3 consistently deleted in 25% of neuroblastomas was
defined by White and colleagues; AJAP1 was 1 of 6 predicted genes
in this deleted region (25).
Neuroblastomas are neuroendocrine tumors arising from neural crest
derivatives of the sympathetic nervous system (26). Fujita et al identified 23
genes, while mapping the smallest region of a consistently deleted
segment of 1p36 in neuroblastomas; AJAP1 was included (27). By analyzing 430 primary
neuroblastomas, Okawa et al mapped the smallest region of
deletion to a 2-Mb region of 1p36 using microsatellite and single
nucleotide polymorphisms and identified 23 genes in this region;
including AJAP1 (13). Milde et
al found the loss of AJAP1 in a progressively metastasizing
ependymoma (28). Ependymomas are
tumors that arise from the specialized ependyma which primarily
lines the ventricular surface of the central nervous system
(29). Dong et al reported
two contiguous minimally deleted regions on chromosome
1p36.31-p36.32 in oligodendroglial tumors, one of which contained
AJAP1 (30). Oligodendrogliomas
are believed to originate from the myelin-producing
oligodendrocytes of the brain (31). McDonald et al evaluated 177
oligodendroglial tumor samples and found a consensus region of
deletion of approximately 630 kb. This region encompasses only one
single gene - AJAP1 (32).
Historically, brain gliomas have been described as likely arising
from unique glial cells; however, more recent studies of genomic
changes in various glioma types suggest there may be common cells
of origin with common genomic changes, such as loss of AJAP1
expression.
For any tumor-suppressor activity of AJAP1 to be
inhibited in a tumor, both copies of the gene must be abrogated,
either by deletion or a reduction in expression. Since AJAP1
expression is found to be low in the majority of glioma tumors and
cell lines (13,24,28,32,33),
the remaining allele of AJAP1 has been sequenced to identify
mutations, revealing none in the coding region (13,32,33).
As shown for some tumor suppressors, decreased gene dosage
predisposes cells to tumorigenesis. Additionally, as the expression
of AJAP1 is often reduced or absent irrespective of a deletion at
that locus, epigenetic silencing has been investigated.
Bioinformatic analysis revealed the presence of numerous CpG
islands in the promoter of AJAP1, the favored site for
methylation-regulation of gene expression (33). Methylation analysis of the AJAP1
promoter identified hypermethylation in 21% of oligodendrogliomas,
and the degree of methylation inversely correlated with AJAP1
expression. The AJAP1 promoter was also highly methylated in a wide
spectrum of cell lines (24).
Using multiple high-resolution genomic screening
methods, we found AJAP1 frequently lost or epigenetically silenced
in many glioblastomas (GBM) (33).
Glioblastomas which arise from astrocytes are the most common and
most aggressive malignant primary brain glioma in humans which can
be found throughout the brain or spinal cord (34). Expression studies revealed that
loss of AJAP1 gene expression is more commonly due to epigenetic
silencing of the gene than gene deletion. Expression was reduced or
absent in 86–92% of primary GBM tumors (those with no history of
prior lower grade tumors) and all glioma cell lines, whereas the
gene was deleted in only 16% of the tumor samples. Using mutational
and methylation analyses, we showed that AJAP1 expression was not
due to mutation but epigenetically silenced with promoter
methylation in many cases (33).
Continuing studies on AJAP1
AJAP1 was initially identified by Bharti et
al in epithelial cells of the uterus as a candidate gene
increased in invasive endometriosis (35). Confocal microscopic analysis of
transiently-expressed AJAP1 revealed its localization at the
baso-lateral part of the cell membrane, where it co-localized with
endogenous E-cadherin, a protein also present at the baso-lateral
part of the membrane in polarized Madin-Darby canine kidney (MDCK)
epithelial cells (36,37). It has been demonstrated that
adhesion is mediated by the cytoplasmic domain of E-cadherin
linking to the actin cytoskeleton (38) via associated proteins such as α-
and β-catenin (39). Direct
interaction between β-catenin and AJAP1 in an in vitro
pull-down assay suggested that AJAP1 might be directly linked to
the E-cadherin via β-catenin. It can be found at the baso-lateral
part of the plasma membrane where it co-localizes with, and
apparently integrates into E-cadherin-mediated adherens junctions
(40).
In polarized MCF7 mammary epithelial cells and MDCK
cells, AJAP1 is a transmembrane protein which co-localizes with
endogenous E-cadherin (35).
Additionally, CD147 was shown to interact with AJAP1 and increase
cell invasion (41). However,
AJAP1 does not appear to interact with the N-cadherin complex in
non-polarized epithelial cells (35). These findings suggest a potential
role for AJAP1 in cell-cell and cell-extracellular matrix
interactions that could be involved in cell motility, migration and
invasion.
Apparently conflicting results are associated with
AJAP1 expression. In HeLa cells (derived from a cervical tumor),
loss of AJAP1 expression results in decreased invasiveness
(41); however, its overexpression
in glioma cells also results in decreased invasiveness, McDonald
et al found in U251 cells (glioma cells) that AJAP1
overexpression decreased cell adhesion on extracellular matrix
components and decreased migration in wound healing assays
(32). These studies demonstrate
that the effect of AJAP1 may depend on the specific cell type and
its environment.
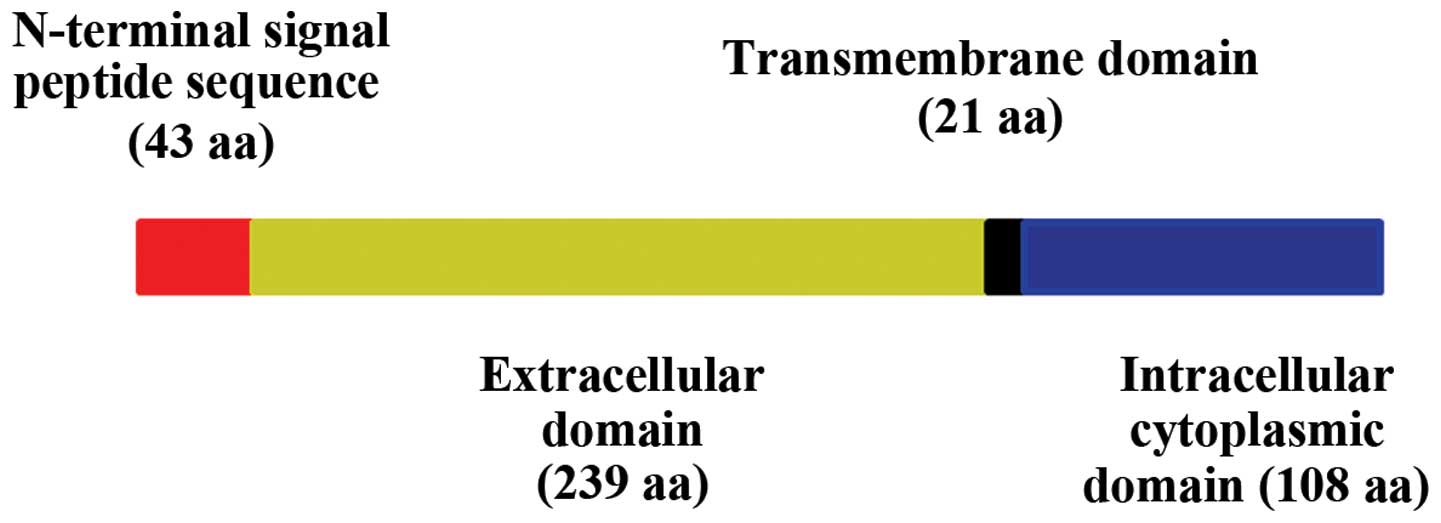
AJAP1 is an integral membrane protein of 411-amino
acid residues (35) and is
comprised of a cleavable N-terminal signal peptide (residues 1–43),
an extracellular domain (residues 44–282), a transmembrane domain
(residues 283–303) and an intracellular cytoplasmic domain
(residues 304–411) (Fig. 2)
(42). The relatively long signal
peptide also has a role in the cellular location of AJAP1 (43,44).
The signal peptide of AJAP1 conforms to the NtraC model describing
the organization of long signal peptides, i.e., an N-terminal
subdomain, a β-turn-rich transition area, and a C-terminal
subdomain. The C-domain, either by itself or with the rest of the
signal peptide, allows the targeting of nascent AJAP1 to the rough
endoplasmic reticulum and subsequent transfer to the plasma
membrane. The N-domain alone directs AJAP1 primarily to
mitochondria. This cryptic mitochondrial targeting signal has been
speculated to be active only under certain physiological
conditions, e.g., apoptosis (43,44).
Further analysis of the AJAP1 protein sequence revealed a putative
nuclear localization signal in the predicted extracellular domain
and putative glycosylation signals in the cytoplasmic domain
(35).
Direct interaction between β-catenin and AJAP1 in an
in vitro pull-down assay suggested that AJAP1 might be
linked to the E-cadherin-mediated junctional complexes via
β-catenin (35). β-catenin
contains armadillo repeats and is able to bind to other proteins
such as transcription factors (45). The ability of β-catenin to bind to
other proteins is regulated by tyrosine kinases (46) and serine kinases such as GSK-3
(47). Various signals such as the
Wnt signaling pathway can inhibit GSK-3-mediated phosphorylation of
β-catenin, allowing β-catenin to translocate to the cell nucleus,
interact with transcription factors, and regulate gene
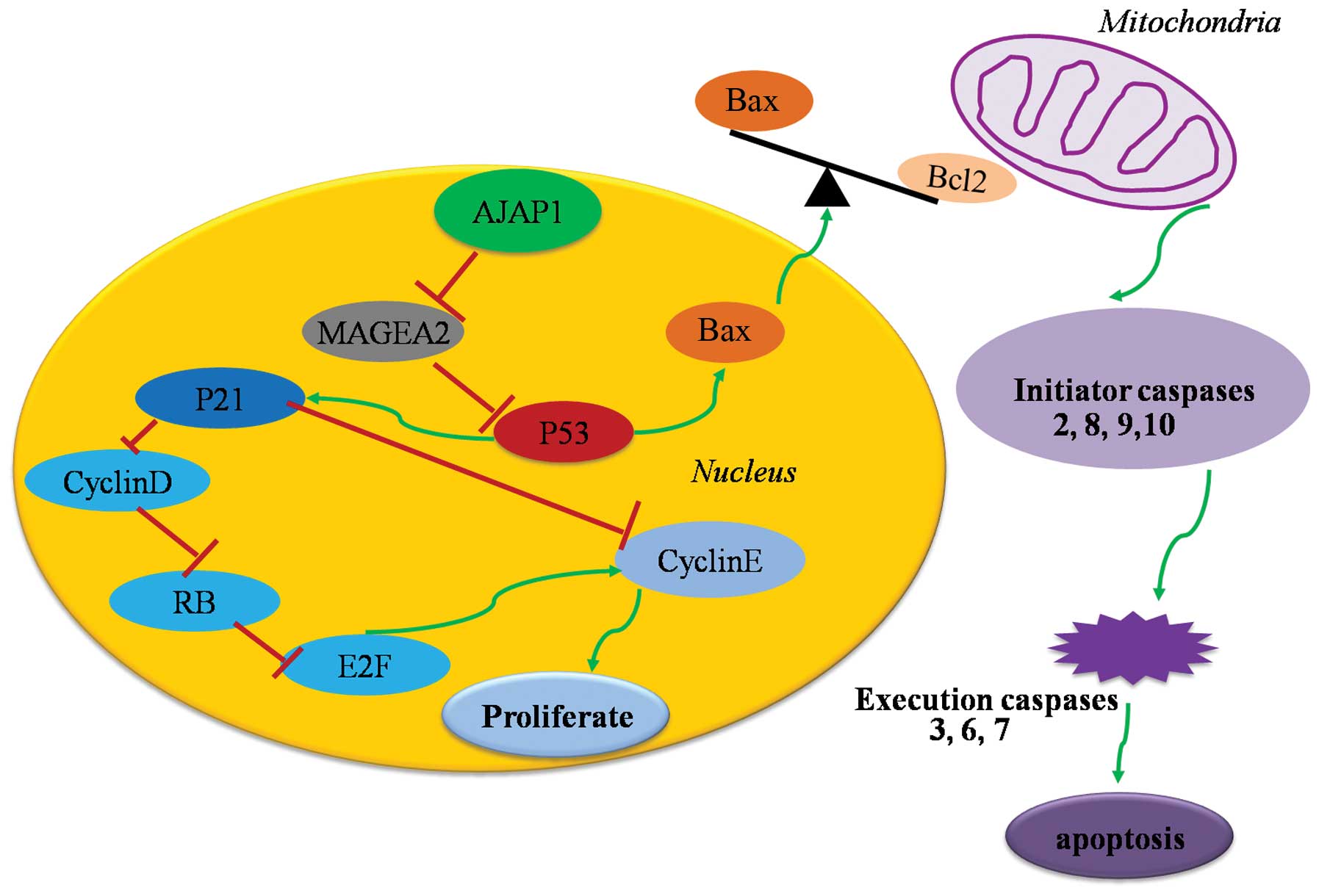
transcription (48). AJAP1 may be
translocated to the nucleus, via its interaction with β-catenin
complexes, where it can regulate gene transcription, therefore
possibly having a potent impact on cell cycling and apoptosis
(Fig. 3). One of our studies
suggested a regulatory effect of AJAP1 on Bax/Bcl-2 ratios with
increased caspase-3/7 activity in the presence of AJAP1, which
indicated that AJAP1 could, at least in part, induce the
mitochondria-related apoptosis pathways (49).
Relationship between AJAP1 and tumors
Cogdell and colleagues suggested overexpression of
AJAP1 could suppress cell invasion and migration in GBM cell lines
(24). We identified loss of AJAP1
expression as being associated with increased glioma cell
proliferation and migration (33).
Interaction with β-catenin, a critical member of the
Wnt signaling pathway, also suggests a possible role of AJAP1 in
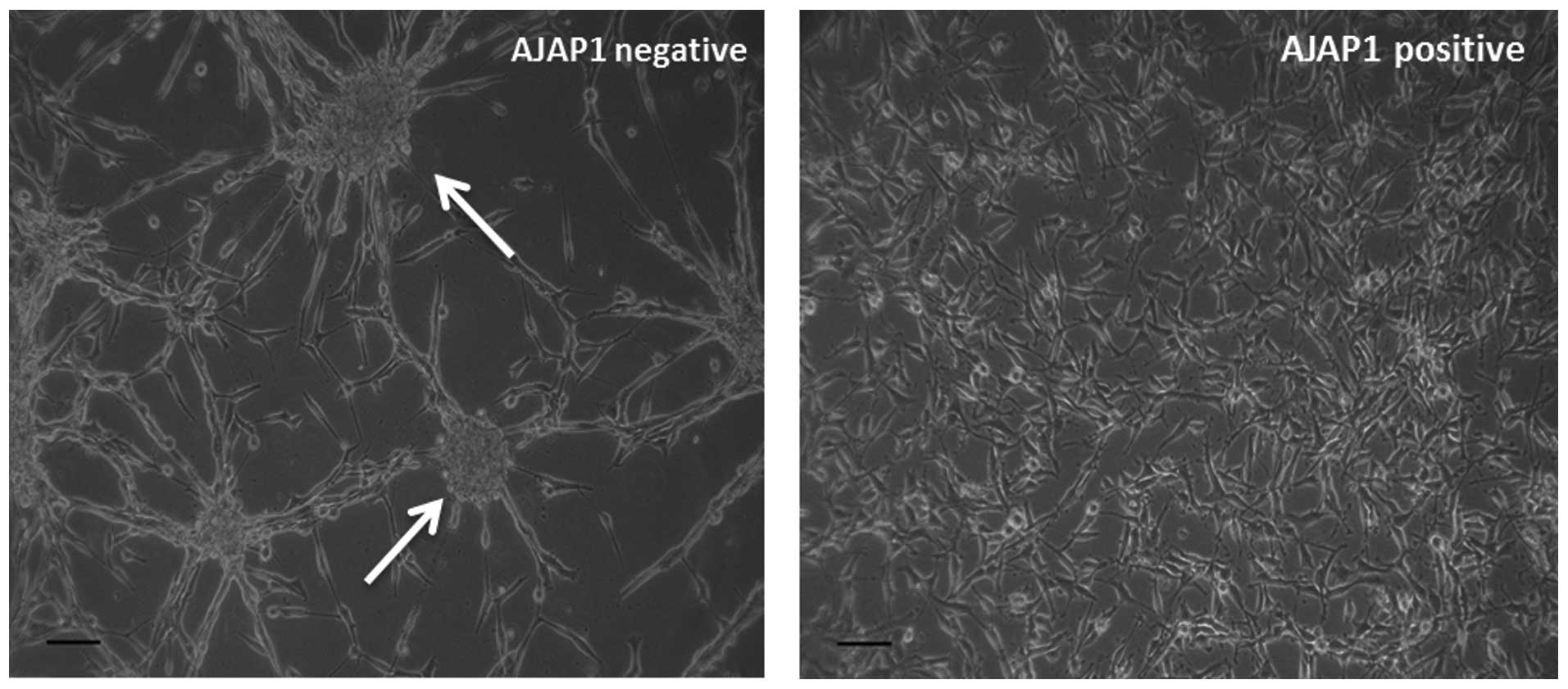
glioma stem cell (GSC) modulation. We observed that in the presence
of AJAP1, U87MG cells (a GBM cell line) do not form tumorspheres
while in the absence of AJAP1, get tumorspheres (Fig. 4). When probed for CD133, a marker
of GSCs, we also noticed a decrease in its expression in AJAP1
transfected cells (data not shown) indicating either a loss of
stem-like properties of the cell or decrease in the GSC
subpopulation.
The group of Radlwimmer (50) performed comparative genomic
hybridization and microarray analysis of spheroid cultures from GBM
patients, where AJAP1 as well as EMP3 and PDPN were first described
as novel candidate genes that likely play a role in GBM
pathogenesis and biomarkers associated with GBM outcome. By
analyzing the National Cancer Institute’s Rembrandt dataset, which
contains 343 glioma samples, McDonald and colleagues (32) indicated that low AJAP1 gene
expression was associated with decreased survival. Both genetic
(gene deletion) and epigenetic alterations (promoter methylation)
are likely mechanisms that inactivate the putative tumor suppressor
AJAP1 in gliomas, which contributes to poor prognosis (24). We and others also demonstrated that
the AJAP1 promoter was highly methylated in a wide spectrum of
glioma cell lines, and the loss of expression was associated with
poorer survival in gliomas patients (33). We recently showed that at an early
stage of gliomagenesis AJAP1 is dysregulated and may suppress
invasion through cytoskeleton reorganization associated with actin
and microtubules associations (51). Similar result was found in a cohort
study of cervical cancer (8).
Restoration of AJAP1 gene expression by transfection or
demethylation agents results in decreased tumor cell migration in
GBM cell lines. These investigations show the significant loss of
expression of AJAP1 in GBM and provide evidence of its role in the
highly migratory characteristic of these tumors (33).
Closing remarks
In summary, gene profiling showed AJAP1 on 1p36 is
frequently lost or epigenetically silenced in various kinds of
human cancers. AJAP1 may affect cell motility, migration, invasion
and proliferation by unclear mechanisms in different cell types. In
particular, loss of AJAP1 expression predicts poor clinical outcome
of patients with malignant gliomas such as GBM and it may serve as
a promising tumor suppressor-related target. Although AJAP1 does
not conform to the classical definition of a tumor suppressor,
accumulating evidence from studies on other human cancers may
identify AJAP1 as a tumor suppressor via the alternate dosage
hypothesis.
References
|
1.
|
Caron H, Spieker N, Godfried M, et al:
Chromosome bands 1p35–36 contain two distinct neuroblastoma tumor
suppressor loci, one of which is imprinted. Genes Chromosomes
Cancer. 30:168–174. 2001.
|
|
2.
|
Godfried MB, Veenstra M, Valent A, et al:
Lack of interstitial chromosome 1p deletions in clinically-detected
neuroblastoma. Eur J Cancer. 38:1513–1519. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Felsberg J, Erkwoh A, Sabel MC, et al:
Oligodendroglial tumors: refinement of candidate regions on
chromosome arm 1p and correlation of 1p/19q status with survival.
Brain Pathol. 14:121–130. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Bagchi A, Papazoglu C, Wu Y, et al: CHD5
is a tumor suppressor at human 1p36. Cell. 128:459–475. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Piaskowski S, Rieske P, Szybka M, et al:
GADD45A and EPB41 as tumor suppressor genes in meningioma
pathogenesis. Cancer Genet Cytogenet. 162:63–67. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Edström Elder E, Nord B, Carling T, et al:
Loss of heterozygosity on the short arm of chromosome 1 in
pheochromocytoma and abdominal paraganglioma. World J Surg.
26:965–971. 2002.
|
|
7.
|
Bieche I, Khodja A and Lidereau R:
Deletion mapping of chromosomal region 1p32-pter in primary breast
cancer. Genes Chromosomes Cancer. 24:255–263. 1999. View Article : Google Scholar
|
|
8.
|
Cheung TH, Lo KW, Yim SF, et al:
Clinicopathologic significance of loss of heterozygosity on
chromosome 1 in cervical cancer. Gynecol Oncol. 96:510–515. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Zhou CZ, Qiu GQ, Zhang F, He L and Peng
ZH: Loss of heterozygosity on chromosome 1 in sporadic colorectal
carcinoma. World J Gastroenterol. 10:1431–1435. 2004.PubMed/NCBI
|
|
10.
|
Girard L, Zochbauer-Muller S, Virmani AK,
Gazdar AF and Minna JD: Genome-wide allelotyping of lung cancer
identifies new regions of allelic loss, differences between small
cell lung cancer and non-small cell lung cancer, and loci
clustering. Cancer Res. 60:4894–4906. 2000.PubMed/NCBI
|
|
11.
|
Kleer CG, Bryant BR, Giordano TJ, Sobel M
and Merino MJ: Genetic changes in chromosomes 1p and 17p in thyroid
cancer progression. Endocr Pathol. 11:137–143. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Schlisio S, Kenchappa RS, Vredeveld LC, et
al: The kinesin KIF1beta acts downstream from EglN3 to induce
apoptosis and is a potential 1p36 tumor suppressor. Genes Dev.
22:884–893. 2008. View Article : Google Scholar
|
|
13.
|
Okawa ER, Gotoh T, Manne J, et al:
Expression and sequence analysis of candidates for the 1p36.31
tumor suppressor gene deleted in neuroblastomas. Oncogene.
27:803–810. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Robbins CM, Tembe WA, Baker A, et al: Copy
number and targeted mutational analysis reveals novel somatic
events in metastatic prostate tumors. Genome Res. 21:47–55. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Sjoblom T, Jones S, Wood LD, et al: The
consensus coding sequences of human breast and colorectal cancers.
Science. 314:268–274. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Ichimura K, Vogazianou AP, Liu L, et al:
1p36 is a preferential target of chromosome 1 deletions in
astrocytic tumours and homozygously deleted in a subset of
glioblastomas. Oncogene. 27:2097–2108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Tanas MR, Sboner A, Oliveira AM, et al:
Identification of a disease-defining gene fusion in epithelioid
hemangioendothelioma. Sci Transl Med. 3:92–98. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Henrich KO, Fischer M, Mertens D, et al:
Reduced expression of CAMTA1 correlates with adverse outcome in
neuroblastoma patients. Clin Cancer Res. 12:131–138. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Zhang H, Zhai Y, Hu Z, et al: Genome-wide
association study identifies 1p36.22 as a new susceptibility locus
for hepatocellular carcinoma in chronic hepatitis B virus carriers.
Nat Genet. 42:755–758. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Caren H, Fransson S, Ejeskar K, Kogner P
and Martinsson T: Genetic and epigenetic changes in the common 1p36
deletion in neuroblastoma tumours. Br J Cancer. 97:1416–1424. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Liu Z, Yang X, Li Z, et al: CASZ1, a
candidate tumor-suppressor gene, suppresses neuroblastoma tumor
growth through reprogramming gene expression. Cell Death Differ.
18:1174–1183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
He L, He X, Lim LP, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar
|
|
23.
|
Guessous F, Zhang Y, Kofman A, et al:
microRNA-34a is tumor suppressive in brain tumors and glioma stem
cells. Cell Cycle. 9:1031–1036. 2010. View Article : Google Scholar
|
|
24.
|
Cogdell D, Chung W, Liu Y, et al:
Tumor-associated methylation of the putative tumor suppressor AJAP1
gene and association between decreased AJAP1 expression and shorter
survival in patients with glioma. Chin J Cancer. 30:247–253. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
White PS, Thompson PM, Gotoh T, et al:
Definition and characterization of a region of 1p36.3 consistently
deleted in neuroblastoma. Oncogene. 24:2684–2694. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Cheung NK and Dyer MA: Neuroblastoma:
developmental biology, cancer genomics and immunotherapy. Cancer.
13:397–411. 2013.PubMed/NCBI
|
|
27.
|
Fujita T, Igarashi J, Okawa ER, et al:
CHD5, a tumor suppressor gene deleted from 1p36.31 in
neuroblastomas. J Natl Cancer Inst. 100:940–949. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Milde T, Pfister S, Korshunov A, et al:
Stepwise accumulation of distinct genomic aberrations in a patient
with progressively metastasizing ependymoma. Genes Chromosomes
Cancer. 48:229–238. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Nagasawa DT, Trang A, Choy W, et al:
Genetic expression profiles of adult and pediatric ependymomas:
molecular pathways, prognostic indicators, and therapeutic targets.
Clin Neurol Neurosurg. 115:388–399. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Dong Z, Pang JS, Ng MH, Poon WS, Zhou L
and Ng HK: Identification of two contiguous minimally deleted
regions on chromosome 1p36.31-p36.32 in oligodendroglial tumours.
Br J Cancer. 91:1105–1111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Alentorn A, Sanson M and Idbaih A:
Oligodendrogliomas: new insights from the genetics and
perspectives. Curr Opin Oncol. 24:687–693. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
McDonald JM, Dunlap S, Cogdell D, et al:
The SHREW1 gene, frequently deleted in oligodendrogliomas,
functions to inhibit cell adhesion and migration. Cancer Biol Ther.
5:300–304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Lin N, Di C, Bortoff K, et al: Deletion or
epigenetic silencing of AJAP1 on 1p36 in glioblastoma. Mol Cancer
Res. 10:208–217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Ohgaki H and Kleihues P: The definition of
primary and secondary glioblastoma. Clin Cancer Res. 19:764–772.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Bharti S, Handrow-Metzmacher H,
Zickenheiner S, Zeitvogel A, Baumann R and Starzinski-Powitz A:
Novel membrane protein shrew-1 targets to cadherin-mediated
junctions in polarized epithelial cells. Mol Biol Cell. 15:397–406.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Le Bivic A, Sambuy Y, Mostov K and
Rodriguez-Boulan E: Vectorial targeting of an endogenous apical
membrane sialoglyco-protein and uvomorulin in MDCK cells. J Cell
Biol. 110:1533–1539. 1990.PubMed/NCBI
|
|
37.
|
Shore EM and Nelson WJ: Biosynthesis of
the cell adhesion molecule uvomorulin (E-cadherin) in Madin-Darby
canine kidney epithelial cells. J Biol Chem. 266:19672–19680.
1991.PubMed/NCBI
|
|
38.
|
Sako Y, Nagafuchi A, Tsukita S, Takeichi M
and Kusumi A: Cytoplasmic regulation of the movement of E-cadherin
on the free cell surface as studied by optical tweezers and single
particle tracking: corralling and tethering by the membrane
skeleton. J Cell Biol. 140:1227–1240. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Aberle H, Butz S, Stappert J, Weissig H,
Kemler R and Hoschuetzky H: Assembly of the cadherin-catenin
complex in vitro with recombinant proteins. J Cell Sci.
107:3655–3663. 1994.PubMed/NCBI
|
|
40.
|
Jakob V, Schreiner A, Tikkanen R and
Starzinski-Powitz A: Targeting of transmembrane protein shrew-1 to
adherens junctions is controlled by cytoplasmic sorting motifs. Mol
Biol Cell. 17:3397–3408. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Schreiner A, Ruonala M, Jakob V, et al:
Junction protein shrew-1 influences cell invasion and interacts
with invasion-promoting protein CD147. Mol Biol Cell. 18:1272–1281.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Resch E, Quaiser S, Quaiser T, Schneider
G, Starzinski-Powitz A and Schreiner A: Synergism of shrew-1’s
signal peptide and transmembrane segment required for plasma
membrane localization. Traffic. 9:1344–1353. 2008.PubMed/NCBI
|
|
43.
|
Hiss JA, Resch E, Schreiner A, Meissner M,
Starzinski-Powitz A and Schneider G: Domain organization of long
signal peptides of single-pass integral membrane proteins reveals
multiple functional capacity. PloS One. 3:e27672008. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Resch E, Hiss JA, Schreiner A, Schneider G
and Starzinski-Powitz A: Long signal peptides of RGMa and DCBLD2
are dissectible into subdomains according to the NtraC model. Mol
Biosyst. 7:942–951. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Gottardi CJ and Peifer M: Terminal regions
of beta-catenin come into view. Structure. 16:336–338. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Lilien J and Balsamo J: The regulation of
cadherin-mediated adhesion by tyrosine
phosphorylation/dephosphorylation of beta-catenin. Curr Opin Cell
Biol. 17:459–465. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Castellone MD, Teramoto H, Williams BO,
Druey KM and Gutkind JS: Prostaglandin E2 promotes colon cancer
cell growth through a Gs-axin-beta-catenin signaling axis. Science.
310:1504–1510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Liu X, Rubin JS and Kimmel AR: Rapid,
Wnt-induced changes in GSK3beta associations that regulate
beta-catenin stabilization are mediated by Galpha proteins. Curr
Biol. 15:1989–1997. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Zeng L, Kang C, Di C, et al: The adherens
junction-associated protein 1 is a negative transcriptional
regulator of MAGEA2, which potentiates temozolomide-induced
apoptosis in GBM. Int J Oncol. 44:1243–1251. 2014.
|
|
50.
|
Ernst A, Hofmann S, Ahmadi R, et al:
Genomic and expression profiling of glioblastoma stem cell-like
spheroid cultures identifies novel tumor-relevant genes associated
with survival. Clin Cancer Res. 15:6541–6550. 2009. View Article : Google Scholar
|
|
51.
|
Han L, Zhang KL, Zhang JX, et al: AJAP1 is
dysregulated at an early stage of gliomagenesis and suppresses
invasion through cytoskeleton re-organization. CNS Neurosci Ther.
20:429–437. 2014. View Article : Google Scholar : PubMed/NCBI
|