Introduction
More than six decades after its synthesis (1), 5-fluorouracil (5-FU) remains a key
agent, particularly for the treatment of adenocarcinomas of the
gastrointestinal tract (2,3). In the United States, preferences for
5-FU administration have gradually shifted from bolus i.v.
injection to infusion via a pump to improve safety profile of the
drug, because like most chemotherapeutic agents, 5-FU also has
numerous toxic effects such as diarrhea, mucositis,
myelosuppression and thrombophlebitis of peripheral veins (4). However, more recently, orally
delivered derivatives and pro-drugs have been developed.
Capecitabine in particular is an oral pro-drug that is ultimately
converted to 5-FU by thymidine phosphorylase and uridine
phosphorylase, both enzymes are highly expressed in solid tumors
allowing for a locally increased concentration of the active drug
at the cancer site (5).
Although these systems of drug administration
reduces most of the side effects of the therapy, it increases the
occurrence of a toxic reaction involving keratinocytes of the
palmar region of hands and foot, also called palmar-plantar
erythrodysaesthesia or hand-foot syndrome (HFS) (6,7). The
symptoms of HFS include numbness, dysaesthesia/paraesthesia,
tingling, erythema, painless swelling or discomfort and, in more
severe cases, blisters, ulceration, desquamation or severe pain on
the hands palms and/or feet soles.
The cellular damage has also been identified due to
direct contact of keratinocytes with 5-FU. Therefore, toxic
reactions are also possible in healthcare workers, during
manipulation stages of antiblastic drugs. Several scientific
studies have shown that there is the possibility of exposure to
antineoplastic drugs such as doxorubicin, epirubicin,
cyclophosphamide and 5-FU in workers (8,9).
5-FU, in particular, has been reported as one of the most
concentrated chemotherapeutic drugs in the working areas (950
ng/cm2) outside the laminar flow hood, where operators
do not use protective gloves (8).
5-FU is often co-administered together with L-folinic acid or
levofolene (LF) for their positive interaction on cancer cell
growth inhibition due to the formation of a stable ternary complex
with 5-FU and tymidilate synthase, the target of 5-FU (10). However, studies are required in
order to understand the possible interaction between the two drugs
also on the induction of detrimental effects.
In the present study, we studied the effects of LF
on the toxicity determined by 5-FU in a model of the human
keratinocyte HaCaT cell line. Moreover, we studied the interaction
between the two drugs on the cell death mechanisms and oxidative
stress in the same in vitro model of keratinocytes.
Materials and methods
Materials
DMEM, FBS (fetal bovine serum) and tissue culture
plastic ware were purchased from Microtech (Naples, Italy).
Dihydroethidium (DHE) and monodansylcadaverine (MDC) were purchased
from Sigma-Aldrich (Milan, Italy). Annexin V-FITC Apoptosis
Detection kit was purchased from eBioscience (San Diego, CA, USA).
5-Fluorouracil (5-FU), doxorubicin (DOXO) and Levofolene (LF) were
a gift of Dr Gaetano Facchini (I.N.T. ‘Pascale’, Naples,
Italy).
Cell culture and cell viability
assay
The human keratinocyte HaCaT cell line was obtained
from American Type Tissue Culture Collection (Rockville, MD) and
grown in DMEM supplemented with 10% heat-inactivated fetal bovine
serum, 20 mM HEPES, 100 U/ml penicillin, 100 mg/ml streptomycin, 1%
L-glutamine and 1% sodium pyruvate. Cells were cultured at 37°C in
a 5% CO2-95% air environment in humidified incubator.
Proliferation of HaCaT cells was performed in the presence of 5-FU
with or without 10−4 M of LF and DOXO. After
trypsinization, cells were plated in 100 μl of medium in 96-well
plates at a density of 6×103 per well. Cells were
treated 24 h later with increasing concentrations of
pharmacological agents ranging from 6–2.9×10−3 μM of
5-FU, with or without 10−4 M of LF,
4–9.75×10−4 μM of DOXO. Cell proliferation was evaluated
by MTT assay as previously described (11).
Flow cytometric analysis of
apoptosis
Annexin V binding was identified by flow cytometry
using Annexin V-FITC staining, following the manufacturer’s
instructions. Apoptotic cell death was also analyzed by propidium
iodide (PI) detection systems (eBioscences, Vienna, Austria).
Briefly, HaCaT cells were seeded in 6-well plates in a number of
15×104 cells per well. After 24 h cells were treated
with concentration inhibiting 50% of cell growth (IC50)
of 5-FU, alone or in combination with 10−4 M of LF, and
DOXO. After 24, 48 and 72 h of treatment cells were trypsinized,
washed twice with PBS 1X and pellet were resuspended in 200 μl
binding buffer 1X. Then we added 5 μl Annexin V-FITC to 195 μl cell
suspension, mixed and incubated for 10 min at room temperature.
Cells were washed in 200 μl binding buffer 1X and were resuspended
in 190 μl binding buffer 1X, then we added 10 μl propidium iodide
(20 μg/ml). The detection of viable cells, early apoptosis cells,
late apoptosis cells and necrotic cells were performed by FACSAria™
(BD Bioscences). For each sample, 2×104 events were
acquired. Analysis was carried out by triplicate determination on
at least three separate experiments.
Flow cytometric analysis of oxidative
stress
ROS generation was analyzed by flow cytometry using
the ROS-sensitive dye dihydroethidium (DHE), a probe for
measurement of super-oxide anion (O2−), as previously
described (12). HaCaT cells were
seeded in 6-well plates in a number of 15×104 cells per
well and were treated 24 h later with IC50 of 5-FU,
alone or in combination with 10−4 M of LF, and DOXO.
After 24, 48 and 72 h of treatment cells were incubated for 1 h at
the end of treatment with 20 ng/ml dihydroethidium stock solution
(2.5 mg/ml). We compared the effects induced by pharmacological
agents treating HaCaT cells with 500 μM of
H2O2, an inducer of superoxide anion
formation, 2000 μM of N-acetylcysteine (NAC), a scavenger compound,
and H2O2 in combination with NAC. At the time
of processing the cells were trypsinized, washed twice with PBS 1X
and the pellet was resuspended in 500 μl of PBS 1X. The dye
accumulation was analyzed by BD FACSAria (BD Bioscences). For each
sample, 2×104 events were acquired. Analysis was carried
out by triplicate determination on at least three separate
experiments.
Flow cytometric analysis of
autophagy
Autophagy was analyzed by flow cytometry using
monodansylcadaverine (MDC) staining. MDC is an auto-fluorescent
agent used as selective marker for autophagic vacuoles (AVOs) and
especially autolysosomes (13).
Briefly, HaCaT cells were seeded in 6-well plates in a number of
15×104 cells per well and were treated 24 h later with
IC50 of 5-FU, alone or in combination with
10−4 M of LF and DOXO. After 24, 48 and 72 h of
treatment cells were incubated with 50 μM of MDC in PBS 1X at 37°C
for 15 min. After incubation, cells were washed twice in PBS 1X,
trypsinized and the pellet was resuspended in 500 μl of PBS 1X.
Samples were immediately analyzed by flow cytometry by BD FACSAria
(BD Bioscences). For each sample, 2×104 events were
acquired. Analysis was carried out by triplicate determination on
at least three separate experiments.
Evaluation of thiobarbituric
acid-reactive species (Tbars) levels
The levels of Tbars were analyzed using a
spectrophotometric assay (14).
For this purpose HaCaT cells were seeded at 1×106 cells
per well into 100 mm dishes. After 24, 48 and 72 h of treatment
with IC50 of 5-FU, alone or in combination with
10−4 M of LF, and DOXO cells were washed twice in
ice-cold PBS 1X, trypsinized and the pellets were collected. After
cell lysis and the evaluation of protein concentration, samples
were incubated with 0.5 ml of 20% acetic acid, pH 3.5, and 0.5 ml
of 0.78% aqueous solution of thiobarbituric acid. After heating at
95°C for 45 min, the samples were centrifuged at 4000 rpm for 5
min. In the supernatant fractions Tbars were quantified by
spectrophotometry at 532 nm (15).
Results were expressed as Tbars μM/μg of serum protein. Analysis
was carried out by triplicate determination on at least three
separate experiments.
Statistical analysis
All data are expressed as mean ± SD. Statistical
analysis was performed by analysis of variance (ANOVA) with
Neumann-Keul’s multiple comparison test or Kolmogorov-Smirnov where
appropriate.
Results
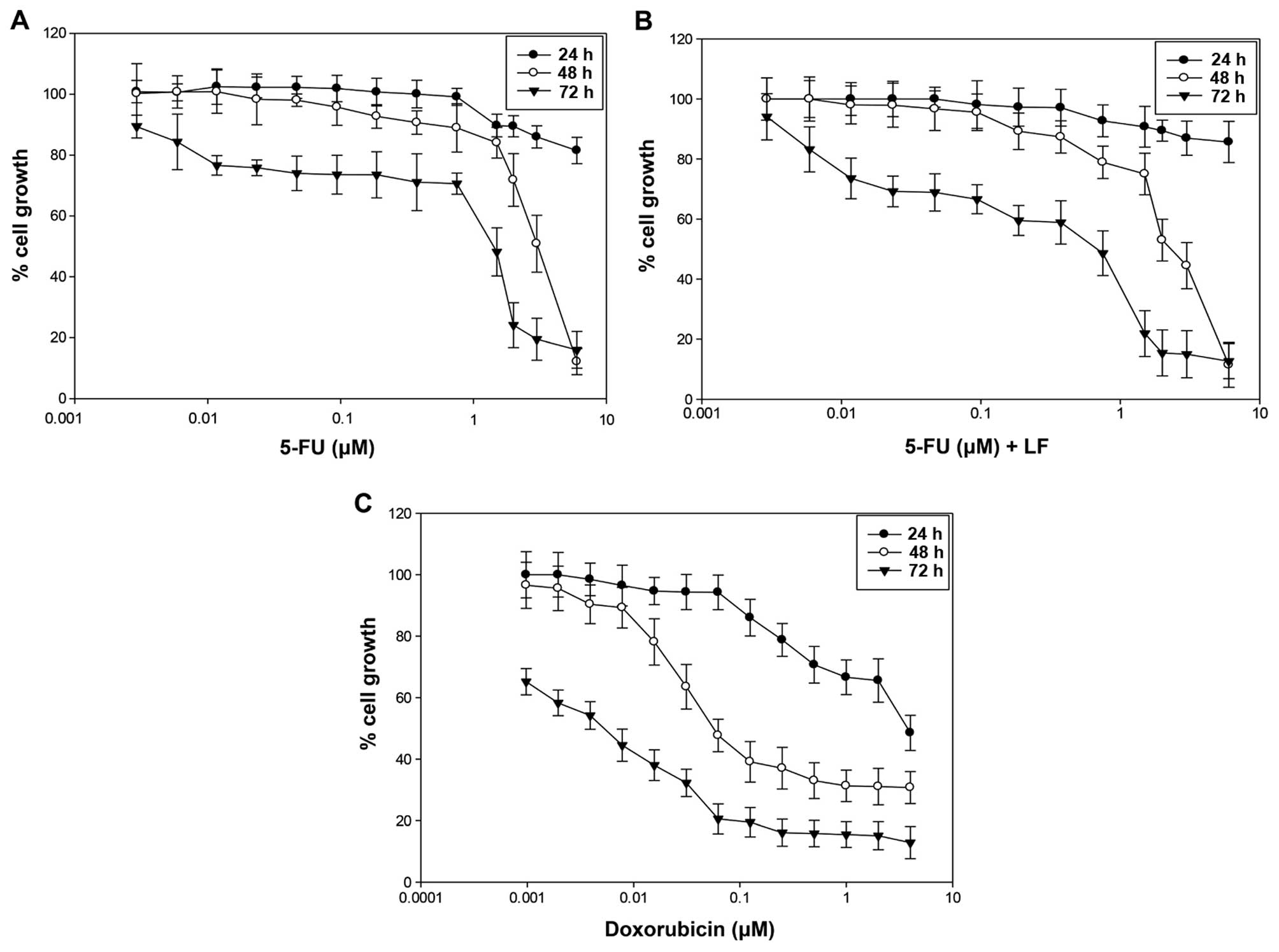
Evaluation of cell growth inhibition of
HaCaT cell line
In this study we evaluated the effects of
5-fluorouracil (5-FU) alone or in combination with levofolene (LF)
and doxorubicin (DOXO) on the proliferation of the human
keratinocyte (HaCaT) cells by MTT assay as reported in Materials
and methods. We found that all the agents induced a time- and
dose-dependent growth inhibition (Fig.
1A–C). The results are expressed as concentration inhibiting
50% of cell growth (IC50) after 72 h of treatment
(Table I). The IC50 was
reached with 1.4 μM of 5-FU (Fig.
1A), 0.7 μM of 5-FU in combination with 10−4 M of LF
(Fig. 1B) and 0.005 μM of DOXO
(Fig. 1C). These data suggested
that human keratinocyte cells were more sensitive to the treatment
with 5-FU in combination with LF compared to those treated with
5-FU alone, confirming that LF potentiated cytotoxic effects of
5-FU. However, HaCaT cells were more sensitive to the treatment
with doxorubicin as IC50 was reached with a lower dose
of drug after 72 h of treatment.
 | Table IConcentrations inhibiting 50% of cell
growth (IC50) in HaCaT cells after 72 h of treatment
with 5-FU, 5-FU in combination with 10−4 M of LF and
doxorubicin.a |
Table I
Concentrations inhibiting 50% of cell
growth (IC50) in HaCaT cells after 72 h of treatment
with 5-FU, 5-FU in combination with 10−4 M of LF and
doxorubicin.a
| Compounds | IC50 ±
SD |
|---|
| 5-FU | 1.4±0.04 μM |
| 5-FU+LF | 0.7±0.01 μM |
| DOXO | 0.005±0.03 μM |
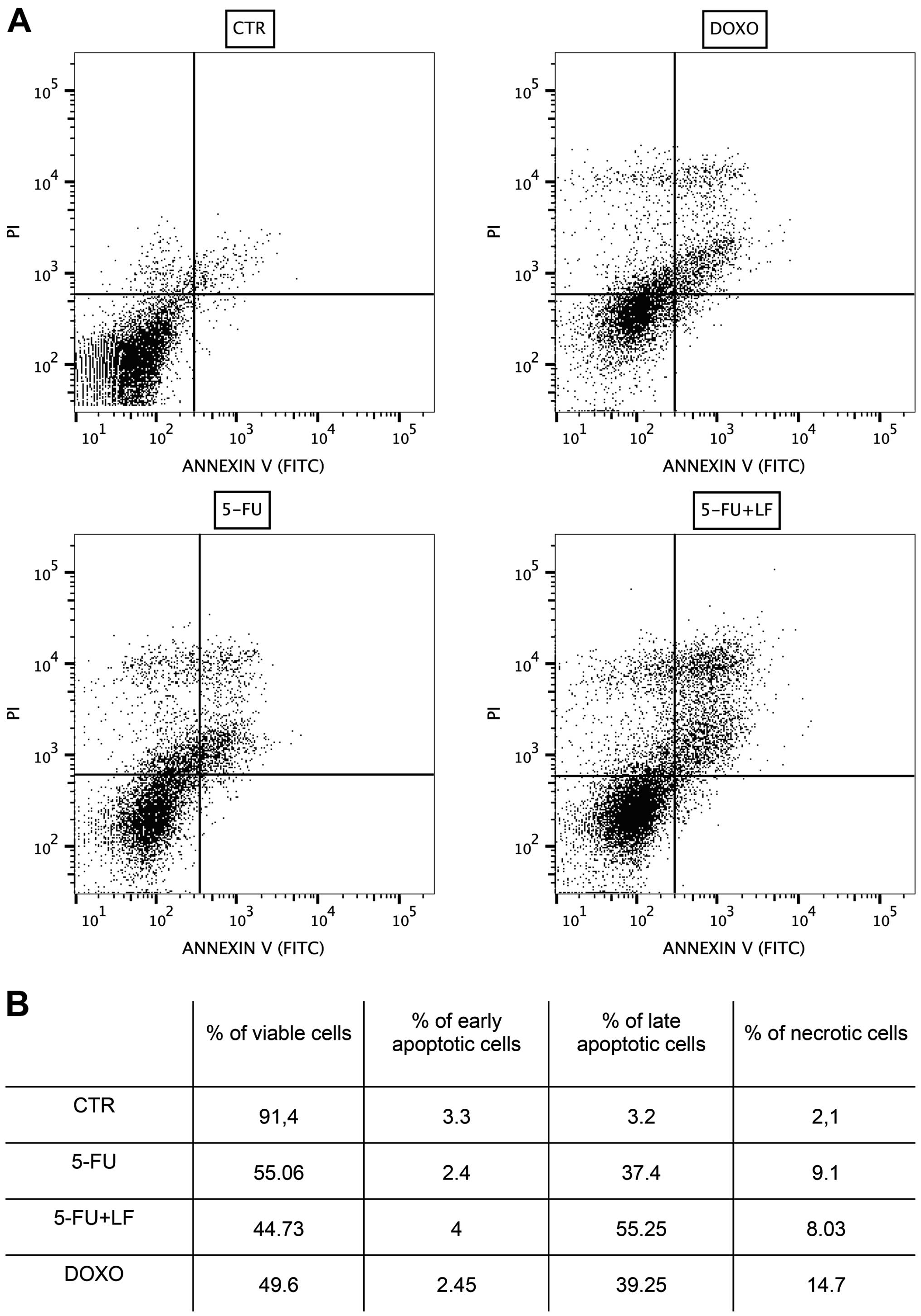
Evaluation of apoptosis by flow
cytometric analysis
The effects of 5-FU, alone or in combination with LF
and DOXO in inducing apoptosis or necrosis in HaCaT cell line were
evaluated after treating cells for 48 h with IC50 of
each compound, as reported in Materials and methods. We have found
that 5-FU induced late apoptosis in ~37% of cells and necrosis in
~9% of cells while the combination with LF induced late apoptosis
in ~55% of cells and necrosis in ~8% of cells. Noteworthy, we found
that DOXO induced late apoptosis in only ~39% of cells and necrosis
in ~15% of cells. The percentage of cells induced in early
apoptosis by each pharmacological treatment was not significant
(Fig. 2). These data suggested
that after 48 h 5-FU damaged human keratinocytes principally
through apoptosis and this effect was potentiated by the
combination with LF while doxorubicin acted also through occurrence
of necrosis.
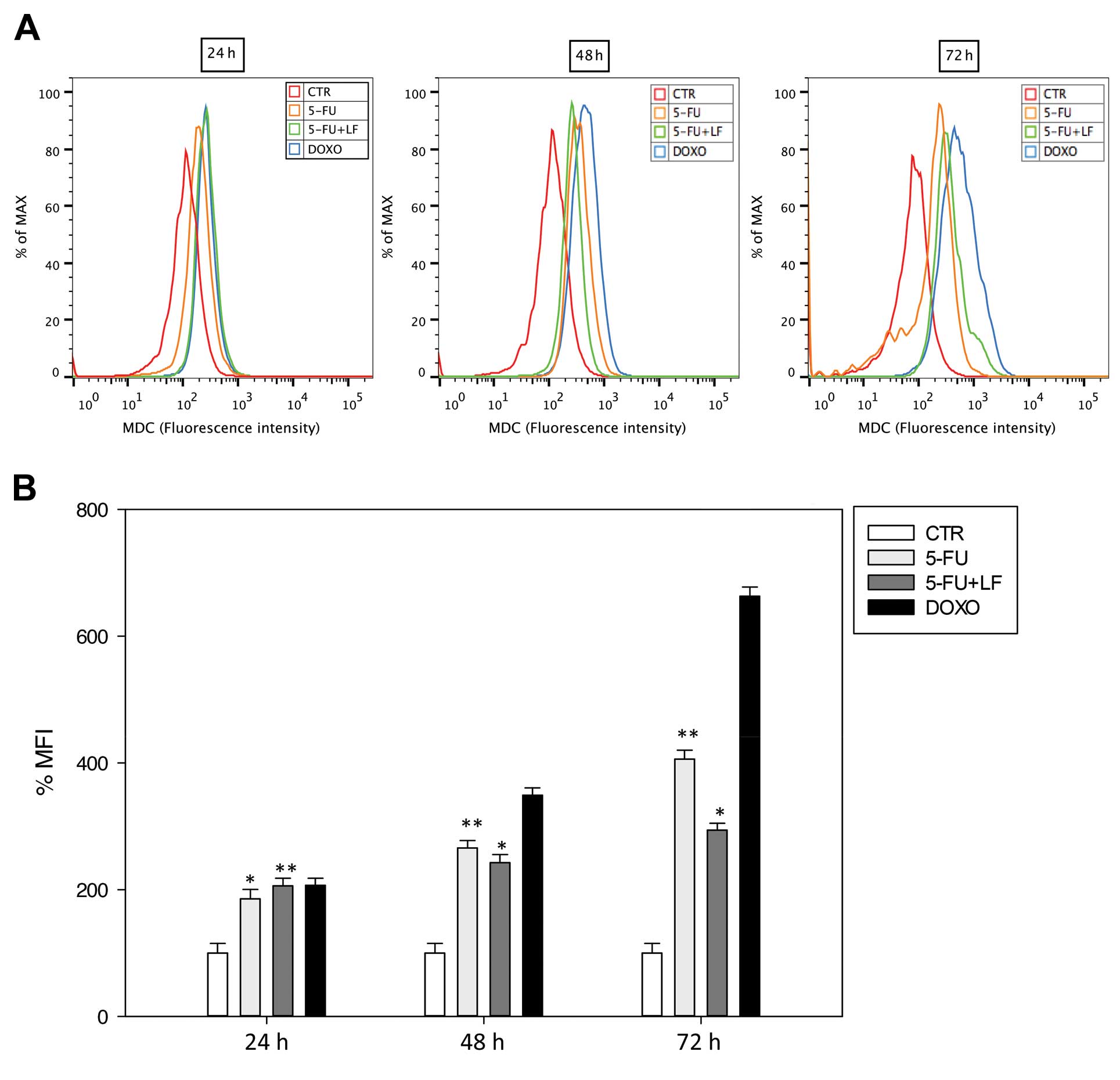
Evaluation of autophagy
In this study we evaluated the effects of the
pharmacological agents in inducing autophagy in HaCaT cell line as
reported in Materials and methods. After 24 h, 5-FU induced an
increase of ~185% of MFI while 5-FU in combination with LF induced
~206% of MFI such as DOXO (Fig.
3). Of note, after 48 and 72 h 5-FU increased the percentage of
MFI in contrast to the combination with LF. 5-FU induced in HaCaT
cells an increase of ~266 and 406% of MFI after 48 and 72 h,
respectively, while the combination with LF induced an increase of
~242 and 294% of MFI after 48 and 72 h, respectively (Fig. 3). On the other hand, DOXO induced a
time-dependent increase of autophagic vacuoles and the maximal
effect was reached after 72 h with an increase of ~663% of MFI
(Fig. 3). These results confirmed
data obtained by apoptosis studies since 5-FU alone induced an
increased cell death that lasted until the end of the treatment. LF
potentiated autophagic effect of 5-FU only at 24 h but protected
keratinocytes from cell death at longer exposure times.
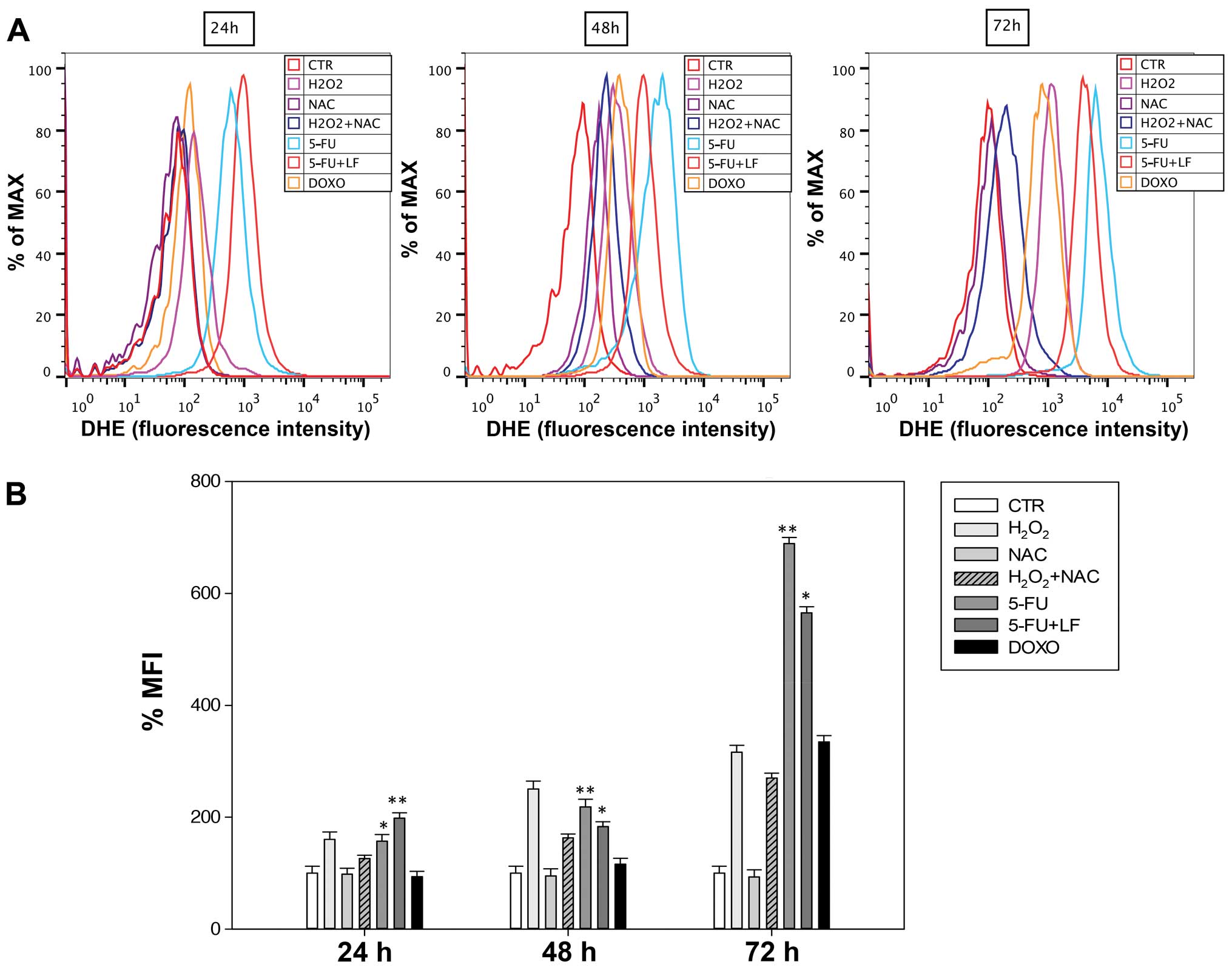
Evaluation of oxidative stress
We evaluated the effects of 5-FU, 5-FU in
combination with LF and DOXO on the accumulation of superoxide
anions (O2−) in HaCaT cells as reported in Materials and
methods. We observed after 24 h that 5-FU induced an increase of
superoxide anions of ~157% of MFI against an increase of ~198% of
MFI induced by the combination with LF (Fig. 4). However, after 48 and 72 h 5-FU
in combination with LF induced an increase of O2− levels
significantly lower compared to that one induced by 5-FU alone.
5-FU induced in HaCaT cells an increase of superoxide anions of
~218 and 689% of MFI while the combination with LF induced an
increase of superoxide anions of ~183 and 565% of MFI after 48 and
72 h, respectively (Fig. 4). On
the other hand, we observed a time-dependent accumulation of ROS
levels in HaCaT cells treated with doxorubicin but it was
significantly lower compared to that one induced by other
pharmacological treatments and the maximal increase of
O2− anions (~334% of MFI) was reached after 72 h
(Fig. 4). NAC had no effect on the
increase of O2− levels and it acted as a scavenger in
combination with H2O2 decreasing the
accumulation of superoxide anions (Fig. 4). Therefore, these data suggested
that LF in combination with 5-FU induced a protective effect on the
formation of ROS in long-time exposure.
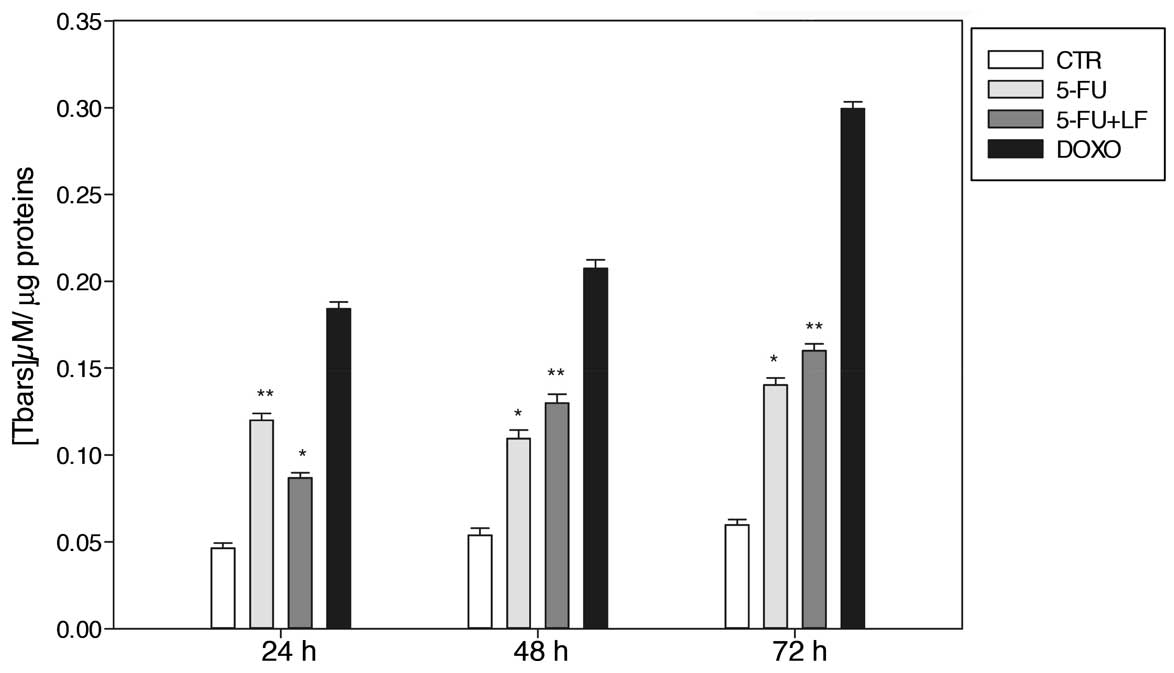
Evaluation of the levels of
thiobarbituric acid-reactive species (Tbars)
In this study, we evaluated the effects of 5-FU,
alone or in combination with LF, and DOXO on the modulation of the
Tbars level in HaCaT cell line as reported in Materials and
methods. After 24 h we found that 5-FU induced the formation of
~0.085 μM Tbars/μg of proteins, while 5-FU in combination with LF
induced a lower formation of the Tbars level which was ~0.087 μM
Tbars/μg of proteins (Fig. 5).
Noteworthy, after 48 and 72 h of treatment the combination 5-FU/LF
induced a greater amount of the Tbars species compared to that one
induced by 5-FU alone. 5-FU in combination with LF induced the
formation of 0.11 μM Tbars/μg of proteins and 0.15 μM Tbars/μg of
proteins after 48 and 72 h, respectively, while 5-FU alone induced
an amount of 0.10 μM Tbars/μg of proteins and 0.16 μM Tbars/μg of
proteins after 48 and 72 h, respectively (Fig. 5). On the other hand, free
doxorubicin caused a time-dependent accumulation of the Tbars level
and the maximal was reached after 72 h with ~0.3 μM Tbars/μg of
proteins (Fig. 5). These data
suggest that the effects of the combination on the increase of
lipid peroxidation were late when compared to those on
intracellular O2− increase suggesting that the decrease
of O2− levels at 48 and 72 h was, at least in part, due
to the formation of reactive species with intracellular complex
molecules such as lipids.
Discussion
5-Fluorouracil (5-FU) has been known to cause
hand-foot syndrome (HFS) since the first description by Lokich and
Moore in 1984 (16). The continued
prolonged exposure to 5-FU, provided by oral administration of
capecitabine, leads to high incidence of HFS.
The clinical manifestation of HFS can be divided
into four grades from slight dysesthesia to desquamation,
blistering, and ulceration. The management of these side effects
can also require the interruption of the therapy for the cancer
disease depending upon the severity of HFS. In fact, the occurrence
of HFS is never life-threatening, but can develop into a
debilitating condition that may severely interfere with the quality
of life (17).
The pathophysiological mechanisms of HFS are an
active area of investigation. The involved factors could be the
following: i) rapid cell division rate of palm and sole
keratinocytes, ii) gravitational forces, iii) peculiar vascular
anatomy of these areas, iv) temperature gradients in the distal
extremities, v) increased levels of thymidine phosphorylase in
keratinocytes. On the basis of these considerations, 5-FU can have
increased cytotoxic effects in these areas if compared with other
body skin sites (6,18). Moreover, the oral 5-FU derivative
capecitabine can be preferentially eliminated by eccrine glands,
resulting in increased excretion in palms and soles that have
higher number of these glands. Histologic features of HFS are
non-specific. They include vacuolar degeneration of the basal cell
layer, mild spongiosis, keratinocyte necrosis, papillary dermal
edema, lymphohistiocytic infiltrates and partial separation of the
epidermis from the dermis (19).
In case of direct topic contact with 5-FU, the
histologic alterations involve keratinocytes in the lower third of
the epidermis. The main intracellular identified abnormalities
described are dilatation of endoplasmatic reticulum and Golgi
complex, membrane-limited perinuclear vacuoles and degeneration of
mitochondria (6,18). Concerning the management of
chemotherapy-induced HFS, discontinuation of the drug, or dose
modifications, are the only available recommendations. Some
measures suggested to control HFS symptoms include cold compresses,
application of emollients and to avoid excessive pressure to the
skin and extreme temperatures. Topical corticosteroids or
dimethylsulfoxide have been also used, but with no definitive
results. Oral or topical pyridoxine (vitamin B6) and topic use of
vitamin K have been successfully used in some instances (20). However, the efficacy of these
measures has not been reported in controlled trials and emphasis is
placed on the management of symptoms as they manifest and progress
(6,21). Therefore, the identification of the
causes that lead to the development of the HSF is a key element for
therapy optimization.
It is also important to underline that the
appearance of toxic effects is also possible in exposed workers, as
well as in patients undergoing chemotherapy. In 5 Japanese
hospitals, 5-FU had the highest concentration detected in working
table, on floor and in air-conditioner filters (43±44
ng/m2; 5.2±4.2 ng/m2 and 4600 ng,
respectively) outside the laminar flow hood, where operators do not
use protective gloves (22). It
has also to be considered that 5-FU is used often in combination
with LF that has largely demonstrated to potentiate its anticancer
effects (10,23). To our knowledge, studies on the
exposure of workers and patients to both 5-FU and LF have not yet
been reported and, similarly, the interaction of the two drugs on
normal human keratinocytes has not yet been reported.
In the present study, we show that 5-FU can induce
keratinocyte growth inhibition at concentrations that are close to
those reported in air-conditioner filters of hospitals and that can
be, therefore, potentially detrimental for both patients and
workers that are continuously exposed to this drug for long periods
of time. Moreover, the combination of 5-FU with LF potentiates the
anti-proliferative effects of the anticancer drug on keratinocytes
suggesting that the co-exposure to both agents can increase not
only the therapeutic action of 5-FU but also its detrimental
effects on skin. Of note, the growth inhibition induced by 5-FU
appeared to be induced by the occurrence of apoptosis paralleled by
markers of an increased oxidative stress as both increased
superanion levels and higher membrane lipid peroxidation. These
results suggest an activation of a mitochondria-dependent apoptotic
mechanism since the increase of intracellular ROS is at the basis
of mitochondria membrane potential transition and the consequent
release of cytochrome c (24). In
our experimental model, a time-dependent increased autophagic
vacuole accumulation was observed in keratinocytes treated with
5-FU as a significant increase of the MDC labeling (marker of late
autophagy vacuoles) was recorded. However, the synergism of 5-FU
with LF on apoptotic occurrence was not paralleled by a similar
interactive increase in autophagic vacuoles at 72 h. These results
suggest that the autophagy observed in 5-FU-treated keratinocytes
could be a protective escape mechanism from apoptotic cell death.
Therefore, cells exposed to LF have antagonizing effects on
autophagy thus increasing the apoptotic cell death. In this light,
it was recently reported that the antidepressant agent and
ubiquitin ligase ITCH1 inhibitor desmethylclomipramine had the
ability to block the autophagic flux that was paralleled by a
paradoxical increase of biochemical markers of autophagic vacuoles
(AVOs) (25). Moreover, the
treatment of cancer cells with cytotoxic drugs together with
desmethylclomipramine potentiated apoptosis and caused an
additional accumulation of autophagic vacuoles suggesting that the
block of autophagic flux induced by desmethylclomipramine can favor
apoptosis caused by cytotoxic agents (26).
Differential effects on reactive oxygen species
(ROS) elevation in cells treated with 5-FU alone or the combination
between 5-FU and LF were also observed. 5-FU induced a
time-dependent increase of both O2− and lipid
peroxidation while the combination of 5-FU and LF caused a stronger
intracellular O2− increase only at 24 h while at 48 and
72 h its effect was lower when compared with that of 5-FU alone. On
the other hand, the addition of LF to 5-FU caused a stronger
increase of lipid peroxidation at 48 and 72 h, but its effects were
significantly lower at 24 h. Therefore, it can be hypothesized that
the combination of 5-FU and LF can elicit apoptosis through the
triggering of cell death mechanisms different from oxidative
stress-dependent activation of apoptosis.
Moreover, the late increase of lipid peroxidation
levels induced by the combination compared to 5-FU alone and the
early increase of O2− levels suggests that superanions
are consumed during the oxidative reaction with intracellular
complex molecules such as lipids. We have also previously reported
that the same combination has potent apoptotic effects on
cardiomyocytes (27). In those
experimental conditions, the synergistic effect of the two agents
on apoptosis was paralleled by a strong oxidative stress occurrence
and by mitochondrial membrane potential transition (28). On these bases, it can be suggested
that the change of the experimental model may have relevance on the
mechanisms of interaction between 5-FU and LF.
In conclusion, our results show that LF
pharmacologically interact with 5-FU potentiating the growth
inhibition and apoptosis of human keratinocytes, and that these
effects occur together with antagonistic effects on autophagy
suggesting the interruption of an anti-apoptotic escape mechanism.
These effects are paralleled by a dynamic induction of oxidative
stress in human keratinocytes. These data strongly support the
design of studies aimed to investigate the exposure of patients and
workers to 5-FU and LF.
Acknowledgements
L.M. was supported by a fellowship from Campania
Research in Experimental Medicine (CREME-FSE 2007/13).
References
|
1
|
Duschinsky R, Plevan E and Heidelberg C:
The synthesis of 5-fluoropyrimidines. J Am Chem Soc. 79:4559–4560.
1957. View Article : Google Scholar
|
|
2
|
Ashraf N, Hoffe S and Kim R: Adjuvant
treatment for gastric cancer: chemotherapy versus radiation.
Oncologist. 18:1013–1021. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rödel C, Hofheinz R and Liersch T: Rectal
cancer: state of the art in 2012. Curr Opin Oncol. 24:441–447.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sorrentino MF, Kim J, Foderaro AE and
Truesdell AG: 5-Fluorouracil induced cardiotoxicity: review of the
literature. Cardiol J. 19:453–458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wan L, Cao D, Zeng J, Yan R, et al:
Modulation of uridine phosphorylase gene expression by tumor
necrosis factor-alpha enhances the antiproliferative activity of
the capecitabine intermediate 5′-deoxy-5-fluorouridine in breast
cancer cells. Mol Pharmacol. 69:1389–1395. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Saif MW: Capecitabine and hand-foot
syndrome. Expert Opin Drug Saf. 10:159–169. 2011. View Article : Google Scholar
|
|
7
|
Walko CM and Lindley C: Capecitabine: a
review. Clin Ther. 27:23–44. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Connor TH, DeBord DG, Pretty JR, et al:
Evaluation of antineo-plastic drug exposure of health care workers
at three university-based US cancer centers. J Occup Environ Med.
52:1019–1027. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pieri M, Castiglia L, Basilicata P, et al:
Biological monitoring of nurses exposed to doxorubicin and
epirubicin by a validated liquid chromatography/fluorescence
detection method. Ann Occup Hyg. 54:368–376. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Avallone A, Di Gennaro E, Bruzzese F, et
al: Synergistic antitumour effect of raltitrexed and 5-fluorouracil
plus folinic acid combination in human cancer cells. Anticancer
Drugs. 18:781–791. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chiosi E, Spina A, Sorrentino A, et al:
Change in TNF-alpha receptor expression is a relevant event in
doxorubicin-induced H9c2 cardiomyocyte cell death. J Interferon
Cytokine Res. 27:589–597. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marra M, Lombardi A, Agostinelli E, et al:
Bovine serum amine oxidase and spm potentiate docetaxel and
interferon-alpha effects in inducing apoptosis on human cancer
cells through the generation of oxidative stress. Biochim Biophys
Acta. 1783:2269–2278. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Biederbick AL, Kern HF and Elsässer H:
Monodansylcadaverine (MDC) is a specific in vivo marker for
autophagic vacuoles. Eur J Cell Biol. 66:3–14. 1995.PubMed/NCBI
|
|
14
|
Caraglia M, Giuberti G, Marra M, et al:
Oxidative stress and ERK1/2 phos-phorylation as predictors of
outcome in hepatocellular carcinoma patientstreated with sorafenib
plus octreotide LAR. Cell Death Dis. 2:e1502011. View Article : Google Scholar
|
|
15
|
Ohkawa H, Ohishi N and Yagi K: Assay for
lipid peroxides in animal tissues by thiobarbituric acid reaction.
Anal Biochem. 95:351–358. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lokich JJ and Moore C:
Chemotherapy-associated palmar-plantar erythrodysesthesia syndrome.
Ann Intern Med. 101:798–799. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Childress J and Lokich J: Cutaneous hand
and foot toxicity associated with cancer chemotherapy. Am J Clin
Oncol. 26:435–436. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Milano G, Etienne-Grimaldi MC, Mari M, et
al: Candidate mechanisms for capecitabine-related hand-foot
syndrome. Br J Clin Pharmacol. 66:88–95. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kirby JS and Miller CJ: Intralesional
chemotherapy for nonmelanoma skin cancer: a practical review. J Am
Acad Dermatol. 63:689–702. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jeung H and Chung HC: Is pyridoxine
helpful in preventing palmar-plantar erythrodysesthesia associated
with capecitabine? Asia Pac J Clin Oncol. 6:141–143. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Eng C: Toxic effects and their management:
daily clinical challenges in the treatment of colorectal cancer.
Nat Rev Clin Oncol. 6:207–218. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yoshida J, Koda S, Nishida S, et al:
Association between occupational exposure levels of antineoplastic
drugs and work environment in five hospitals in Japan. J Oncol
Pharm Pract. 17:29–38. 2011. View Article : Google Scholar
|
|
23
|
Wilson PM, Danenberg PV, Johnston PG, et
al: Standing the test of time: targeting thymidylate biosynthesis
in cancer therapy. Nat Rev Clin Oncol. 11:282–298. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Boccellino M, Giuberti G, Quagliuolo L, et
al: Apoptosis induced by interferon-alpha and antagonized by EGF is
regulated by caspase-3-mediated cleavage of gelsolin in human
epidermoid cancer cells. J Cell Physiol. 201:71–83. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rossi M, Munarriz ER, Bartesaghi S, et al:
Desmethylclomipramine induces the accumulation of autophagy markers
by blocking autophagic flux. J Cell Sci. 122:3330–3339. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rossi M, Rotblat B, Ansell K, et al: High
throughput screening for inhibitors of the HECT ubiquitin E3 ligase
ITCH identifies antidepressant drugs as regulators of autophagy.
Cell Death Dis. 1:e12032014. View Article : Google Scholar
|
|
27
|
Lamberti M, Porto S, Marra M, et al:
5-Fluorouracil induces apoptosis inrat cardiocytes through
intracellular oxidative stress. J Exp Clin Cancer Res. 31:19–23.
2012. View Article : Google Scholar
|
|
28
|
Lamberti M, Porto S, Zappavigna S, et al:
A mechanistic study on the cardiotoxicity of 5-fluorouracil in
vitro and clinical and occupational perspectives. Toxicol Lett.
227:151–156. 2014. View Article : Google Scholar : PubMed/NCBI
|