Introduction
MicroRNAs (miRNAs) are a large family of non-coding
RNAs, typically 18–25 nucleotides in length, that are highly
conserved and endogenously expressed in many species. Newly
discovered intrinsic regulators, the miRNAs regulate gene
expression by binding to the 3′-untranslated regions (3′-UTR)
(1–4). Rapidly accumulating studies have
indicated that miRNAs are differentially expressed in various human
cancers, including non-small cell lung cancer, esophageal cancer,
colorectal cancer, bladder cancer and lymphocytic leukemia, and
they function as tumor suppressors and oncogenes (4–8). It
has been reported that miRNAs are involved in various cellular
processes such as differentiation, migration and apoptosis.
Aberrant expression of miRNAs has been reported in
esophageal carcinoma (EC) (9,10),
and may play a role in the development of EC. We screened a series
of miRNAs in EC tissues and in comparison with their normal
counterparts using miRNA microarrays. Notably, miR-1207-5p was
expressed at a lower level in EC tissues. Previous reports have
shown that miR-1207-5p has differential expression in breast cancer
and in human corneal epithelial cells (11,12).
In addition, miR-1207-5p acted as a suppressor in gastric cancer,
inhibited the growth of tumors by targeting human telomerase
reverse transcriptase, and resulted in reduced tumor volume
(13); miR-1207-5p was also shown
to regulate heparin binding epidermal growth factor expression, to
be involved in extracellular matrix accumulation and to be
associated with disease severity in nephropathy (14,15).
However, the functions of miR-1207-5p in EC have not been reported.
We used TargetScan 6.2 (http://www.targetscan.org) and miRBase (http://www.mirbase.org) to predict that the target of
miR-1207-5p is STOML-2, which is a member of the stomatin family,
but differs from other members of the family by the lack of a
hydrophobic membrane anchor at its N-terminus (16–18).
Reports have indicated that STOML-2 was upregulated in several
kinds of tumor tissue and was involved in invasion and metastasis
of cancers including esophageal cancer, gastric cancer, breast
cancer and glioma (19–23). Inhibition of STOML-2 was able to
decrease cell growth and proliferation, and reduced migratory speed
and invasive ability (24,25).
Herein, we report that miR-1207-5p was markedly
downregulated in 49 EC specimens, and STOML-2 was correspondingly
upregulated. Rates of apoptosis were found to be increased in
EC9706 and EC-1 cells transfected with a miR-1207-5p mimic. We
further measured the level of phospho-I κBα (p-IκBα), an important
molecule in the NF-κB signal pathway, which was downregulated in
miR-1207-5p-transfected cells. Taken together, our results
suggested that miR-1207-5p affected cell invasion and apoptosis in
EC cells by targeting STOML-2.
Materials and methods
Patient information and specimens
Tumor center and marginal tissues (n=49) were
collected at the First Affiliated Hospital of Zhengzhou University
and Tumor Hospital of Linzhou City Henan, China, with the consent
of all the patients and approval by the Ethics Committee of
Zhengzhou University. Tissue samples were stored at −80°C until
analysis. Patients who had undergone preoperative adjuvant therapy
were excluded. The clinicopathological characteristics of the 49 EC
patients are listed in Table I.
Post-operative pathological staging is determined for each
individual according to the seventh edition of the UICC/AJCC TNM
staging system for EC.
 | Table IClinicopathological characteristics
of patients with esophageal carcinoma. |
Table I
Clinicopathological characteristics
of patients with esophageal carcinoma.
|
Characteristics | No. of patients
(n=49) |
|---|
| Gender
(male/female) | 33/16 |
| Age (years)
(≥60/<60) | 31/18 |
| Tumor location
(middle/lower) | 38/11 |
| Lymph node
metastasis (negative/positive) | 33/16 |
| Differentiation
(well/moderate/poor) | 15/25/9 |
| TNM stage
(I/II/III) | 13/24/12 |
Cell lines
Human esophageal cancer EC9706 and EC-1 cells were
purchased from the Cell Bank of the Chinese Academy of Medical
Sciences (Beijing, China). The two cell lines were maintained in
RPMI-1640 medium with 100 U/ml penicillin and 100 μg/ml
streptomycin (Solarbio, Beijing, China) supplemented with 10% fetal
bovine serum (Gibco, Carlsbad, CA, USA) and incubated in a
humidified atmosphere of 5% CO2 at 37°C.
Total RNA isolation and quantitative
reverse transcription PCR analysis
Total RNA was extracted from tissues and cells using
the TRIzol reagent (Invitrogen, Carlsbad, CA, USA); cDNA was
synthesized using the RevertAid First Strand cDNA (K1621; Thermo
Fisher Scientific, Waltham, MA, USA) in 20 μl containing 2 μl of
RNA, 1 μl of random hexamer primer, 9 μl of Nuclease-free water, 4
μl of 5× reaction buffer, 1 μl of RNase inhibitor (20 U/μl), 2 μl
of 10 mM dNTPs mix and 1 μl of reverse transcriptase (200 U/μl).
The mixture was incubated for 5 min at 25°C, followed by 60 min at
42°C, and then at 70°C for 5 min. qRT-PCR was performed using SYBR
Green I (DRR041A; Takara, Dalian, China) following the kit manual.
A specific two step Stemaim-it miR qRT-PCR quantitation kit
(LM-0101A; Novland Co. Ltd., Shanghai, China) was used to detect
miR-1207-5p levels. The primers for miR-1207-5p were
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCCCCTCC-3′
(stem-loop RT primer), 5′-TCCGAAGGCAGGGAGGCAG-3′ (forward),
5′-GTGCAGGGTCCGAGGT-3′ (reverse). The primers for U6 were
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAATA-3′ (stem-loop
RT primer), 5′-TCCGATCGTGAAGCGTTC-3′ (forward),
5′-GTGCAGGGTCCGAGGT-3′ (reverse). Reaction solution including 15 μl
of master mix (2X), 1 μl of enzyme mix, 1 μl of miR primer mix (0.1
μM), 2 μl of cDNA and 11 μl of RNase free water in a final volume
of 30 μl, was pretreated for 3 min at 94°C, then incubated for 20
sec at 94°C, followed by 40 sec at 62°C for 40 cycles. qRT-PCR was
carried out on 7500 Fast Real-time PCR system (Applied Biosystems),
the relative expression levels of the miRNA were calculated using
the comparative Ct (2−ΔCt) and were normalized to U6
small nuclear RNA.
Transient transfections
EC9706 and EC-1 were transfected with 50 nM
miR-1207-5p mimic or negative control (NC, GenePharma, Shanghai,
China). In detail, EC9706 and EC-1 were cultured in 6-well plates
at a density of 1.5×105 cells/well; Lipofectamine™ 2000
(Invitrogen) was used according to the manufacturer’s protocol. The
effects of transfection with miR-1207-5p mimic were evaluated in
each experiment by qRT-PCR.
Cell proliferation assay
The cell counting kit-8 assay (CK04-11; CCK-8,
Dojindo, Rockville, MD, USA) was used to detect cell proliferation.
A total of 1×104 cells/well were placed in a 96-well
plate, and grown for 24, 48, 72 and 96 h. The CCK-8 solution was
added to each well at 10 μl/well, and incubated at 37°C for 2 h.
The absorbance at 450 nm was measured daily over four consecutive
days by a microplate reader (Bio-Rad, Japan). Results were
collected from three separate experiments with five replicate wells
per group.
Cell invasion assays
Cell invasion assays were performed using a
transwell assay (8.0 μm, Corning, NY, USA); the upper chambers were
coated with Matrigel basement membrane matrix (356234; BD
Biosciences, San Jose, CA, USA) in serum-free RPMI-1640 for 3 h at
37°C. EC9706 and EC-1 cells were suspended in serum-free medium at
a density of 2×105 cells/ml at 48 h post-transfection
and placed into the upper chamber (200 μl/well). Medium containing
10% FBS (500 μl) was added to the lower chamber, followed by
incubation at 37°C in 5% CO2 for 30 h. Following this,
the medium and cells in the upper chamber were removed, and the
cells that had migrated to the other side of the membrane were
fixed in 4% paraformaldehyde, stained with 10% crystal purple for
>30 min, and counted in three random fields under an inverted
microscope (Leica Microsystems, Wetzkar, Germany).
Apoptosis assay
Cells were collected at 24 h post-transfection and
stained with Annexin V-FITC apoptosis detection kit (KGA107;
KeyGen, Nanjing, China) according to the instruction manual.
Apoptosis was analyzed using a FACScan® flow cytometer
equipped with CellQuest software (BD Biosciences).
Western blot analysis
Total protein from cultured cells were obtained
using NP-40 lysis buffer (P0013F; Beyotime, Haimen, China) with 1
mM phenylmethanesulfonyl fluoride (ST506; Beyotime). The protein
concentrations were determined using a BCA protein assay kit
(P0012; Beyotime) according to the protocol. Protein lysates were
subjected to SDS-PAGE, and were transferred into polyvinylidene
fluoride (PVDF) membranes. Membranes were blocked with 5% non-fat
milk in TBST for 2 h at room temperature and incubated overnight at
4°C with anti-STOML-2 (1:2,000, Proteintech™, Wuhan, China),
anti-BCL-2 (1:1,500, Proteintech), or anti-pIκBα (pSer32, 1:2,000,
Epitomics, Burlingame, CA, USA). Horseradish peroxidase-conjugated
goat anti-rabbit IgG (1:3,000, Proteintech) was used for detection
of immunoreactive proteins. Signals were detected using ECL kit
(P0018; Beyotime). An antibody against α-tubulin (1:2,000,
Proteintech) served as endogenous control.
3′-UTR luciferase reporter assay
To construct the STOML-2 3′-UTR luciferase reporter
vector, the 3′-UTR of human STOML-2 (NM_013442, bases 1173–1431)
fragment containing the seed sequence of mature miR-1207-5p was
amplified by PCR from human genomic DNA. The primers for STOML-2
3′-UTR were as follows: 5′-AAGCTCGAGTGGAGCTGGGCTTGGCCAGGGAGTCTG-3′
(forward), 5′-GGGTCTAGATGGTTTGCCACTGGTGAGTTTATTACA-3′ (reverse).
The fragment was cloned into the pmirGLO vector (E1330; Promega,
Madison, WI, USA) downstream of the luciferase reporter gene to
construct the recombinant vector, which was named
STOML-2-3′-UTR-WT. The mutant STOML-2 3′-UTR fragment was generated
by overlap extension PCR. The primers were as follows:
5′-AAGCTCGAGTGGAGCTGGGCTTGGCCAGGGAGTCTG-3′ (primer 1 forward),
5′-CTAGCTTGGGACGGTAGATTTTGGTTTTTATTTTTTTATTTG-3′ (primer 1
reverse). 5′-AATCTACCGTCCCAAGCTAGAGCCAGAATCAGG-3′ (primer 2
forward), 5′-GGGTCTAGATGGTTTGCCACTGGTGAGTTTATTACA-3′ (primer 2
reverse). 5′-AAGCTCGAGTGGAGCTGGGCTTGGCCAGGGAGTCTG-3′ (primer 3
forward), 5′-GGGTCTAGATGGTTTGCCACTGGTGAGTTTATTACA-3′ (primer 3
reverse). In detail, with the STOML-2 3′-UTR fragment serving as a
template, primer 1 and primer 2 were used to amplify fragment 1 and
fragment 2; the two fragments and primer 3 were used to generate
the full length mutant fragment. Subsequently, the mutant fragment
was inserted into the pmirGLO vector (E1330; Promega) and named
STOML-2-3′-UTR-MT. For the luciferase reporter assay, human HEK293T
cells were transiently co-transfected with 50 nM of miRNA
(miR-1207-5p mimic or miR-1207-5p NC) and 50 nM of recombinant
vectors (STOML-2-3′-UTR-WT or STOML-2-3′-UTR-MT) using
Lipofectamine 2000. Luciferase activities were analyzed with a
Dual-Luciferase Reporter assay system (E1910; Promega) and a
CentroXS3 LB960 luminometer (Berthold, Germany) at 24 h
post-transfection.
Statistical analysis
Statistical analyses were carried out using SPSS
17.0 software. Numerical results were presented as mean ± standard
deviation. One-way analysis of variance was used to evaluate the
data. Results were considered significant when P<0.05.
Results
The levels of miR-1207-5p expression are
downregulated and STOML-2 are upregulated in EC tissues
To determine whether miR-1207-5p was involved in the
tumorigenesis of EC, we investigated miR-1207-5p expression in 49
matched EC specimens and adjacent non-tumor tissues by qRT-PCR. As
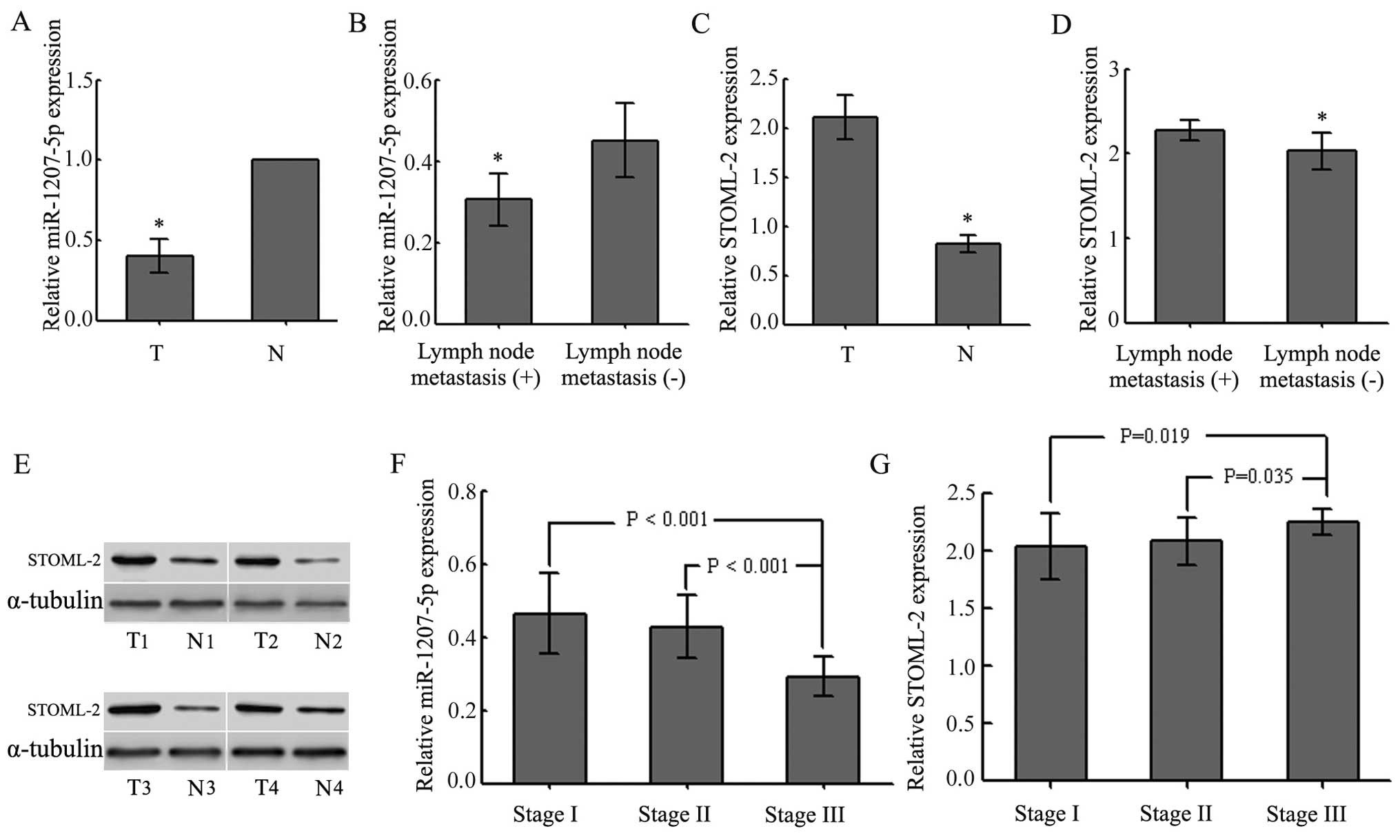
shown in Fig. 1A, the levels of
miR-1207-5p were lower in tumor tissues than in adjacent non-tumor
tissues (P<0.01). We also examined the possible correlation of
miR-1207-5p expression with the occurrence of poor tumor
differentiation and lymph node metastasis (P<0.05; Fig. 1B and Table II) in all patients with EC.
However, there were no significant associations between the levels
of miR-1207-5p expression and gender, age or tumor location
(P>0.05; Table II). Compared
to the counterpart tissues, the levels of STOML-2 in EC tissues
were greatly increased (P<0.05; Fig. 1C and E and Table II). STOML-2 expression levels in
EC tissues were related to lymph node metastases and the
differentiation status (P<0.05; Fig. 1D and Table II), but not to gender, age or
tumor location (P>0.05; Table
II). The expression of miR-1207-5p and STOML-2 exhibited
significant differences at different TNM stages (P<0.05;
Fig. 1F and G and Table II). The results also showed that
miR-1207-5p was negatively correlated with STOML-2. Thus, these
results indicated that downregulation of miR-1207-5p might play an
important role in the progression and development of EC.
| Table IILevels of expression of miRNA-1207-5p
and STOML-2 mRNA in EC samples. |
Table II
Levels of expression of miRNA-1207-5p
and STOML-2 mRNA in EC samples.
| Variables | n | miRNA-1207-5p
(Median ± SD) | P-value | STOML-2 (Median ±
SD) | P-value |
|---|
| Gender | 49 | | | | |
| Male | 33 | 0.4142±0.10604 | 0.320 | 2.0942±0.24529 | 0.571 |
| Female | 16 | 0.3813±0.11099 | | 2.1334±0.17492 | |
| Age | 49 | | | | |
| <60 | 18 | 0.4106±0.12642 | 0.729 | 2.1079±0.25835 | 0.983 |
| ≥60 | 31 | 0.3994±0.09716 | | 2.1065±0.20544 | |
| Tumor location | 49 | | | | |
| Middle | 38 | 0.3892±0.09488 | 0.085 | 2.1442±0.19865 | 0.029a |
| Lower | 11 | 0.4527±0.13741 | | 1.9787±0.26592 | |
| Lymph node
metastasis | 49 | | | | |
| Negative | 33 | 0.4509±0.09112 | 0.000b | 2.0286±0.22070 | 0.000b |
| Positive | 16 | 0.3056±0.06491 | | 2.2688±0.12119 | |
|
Differentiation | 49 | | | | |
| Well | 15 | 0.4720±0.11283 | 0.000b | 1.9742±0.22912 | 0.003b |
| Moderate | 25 | 0.4116±0.07238 | | 2.1267±0.20930 | |
| Poor | 9 | 0.2667±0.03841 | | 2.2739±0.10592 | |
| TNM stage | 49 | | | | |
| I | 13 | 0.4631±1.0980 | 0.000b | 2.0344±0.28936 | 0.043a |
| II | 24 | 0.4271±0.08590 | | 2.0787±0.20213 | |
| III | 12 | 0.2917±0.05424 | | 2.2424±0.11626 | |
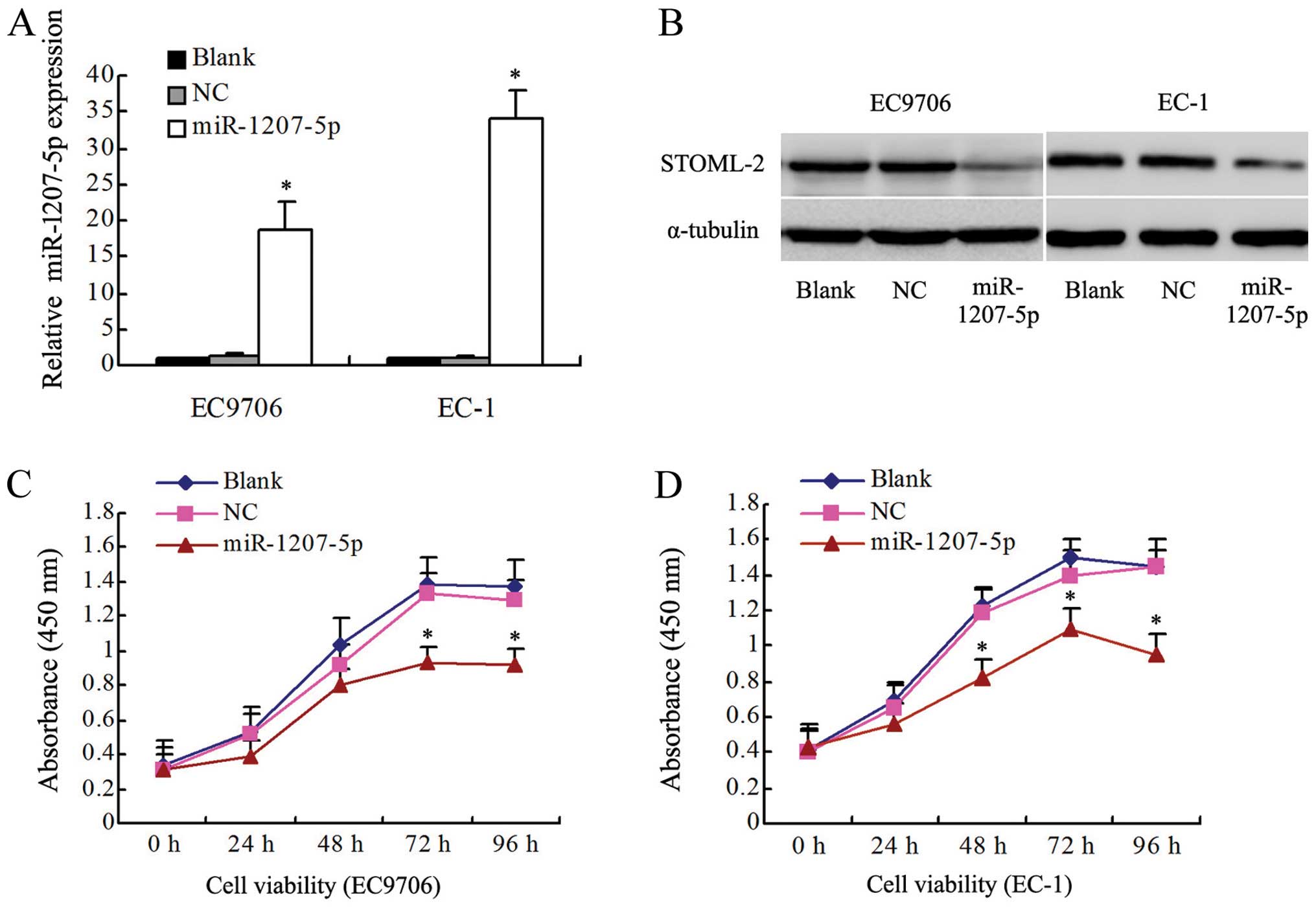
Overexpression of miR-1207-5p inhibits
proliferation in EC9706 and EC-1 cells
To investigate whether miR-1207-5p functions as a
tumor suppressor, the effects of upregulation of miR-1207-5p on the
proliferation of EC cells were determined in vitro. The
levels of miR-1207-5p expression in EC cells were detected by
qRT-PCR. The levels of miR-1207-5p in EC9706 and EC-1 cells
transfected with the miR-1207-5p mimic were higher than in the
control groups, including the non-transfected blank group (blank)
and the negative control group (NC) (P<0.01, Fig. 2A). In contrast, the levels of
STOML-2 were found to be lower in the miR-1207-5p group when
compared with the blank and NC groups (Fig. 2B). As shown in Fig. 2C and D, miR-1207-5p overexpression
significantly decreased the growth rate of EC cells. The absorbance
at 450 nm for the miR-1207-5p group at 48, 72 and 96 h was
significantly decreased (P<0.05) in both EC9706 and EC-1 cells.
However, there were no significant differences between the blank
and NC groups (P>0.05).
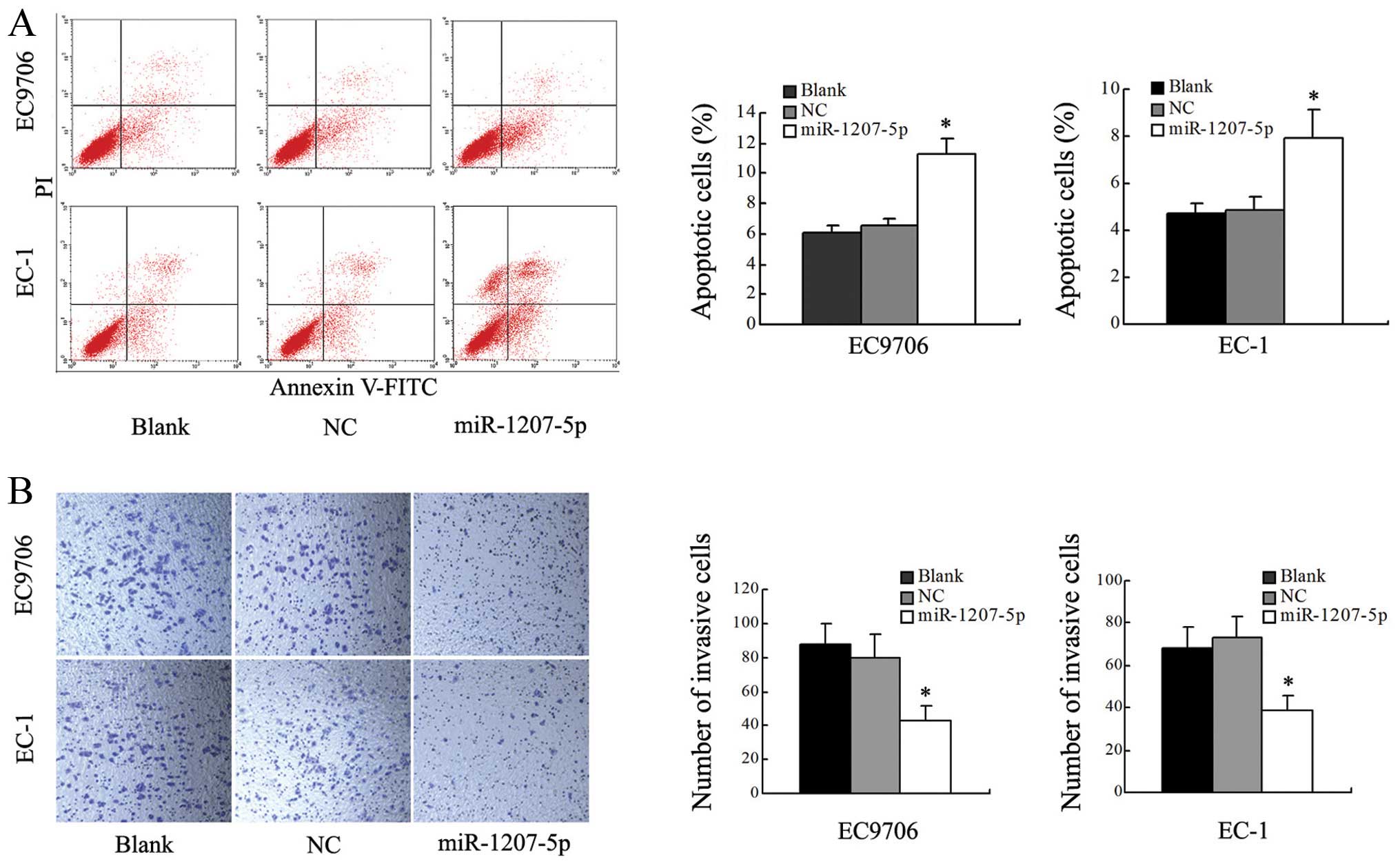
Overexpression of miR-1207-5p induces
apoptosis and restricts cell invasion of EC cells
To determine whether miR-1207-5p was contributing to
apoptosis, we performed flow cytometric analysis of EC cells after
transfection with miR-1207-5p mimic or NC. The results revealed
that the level of apoptosis of cells transfected with the
miR-1207-5p mimic was significantly increased when compared with
the blank and NC groups (P<0.05; Fig. 3A). These results suggested that
upregulation of miR-1207-5p was able to induce apoptosis in EC9706
and EC-1 cells.
Transwell assays were used to test the effect of
miR-1207-5p on cells invasion. Cells transfected with miR-1207-5p
showed a remarkable decrease in invasive capacity compared with
cells in the blank and NC groups (P<0.01, Fig. 3B). However, there were no
significant differences between the blank and NC groups. In
summary, these results indicated that overexpression of miR-1207-5p
decreased the invasive ability of both EC9706 and EC-1 cells.
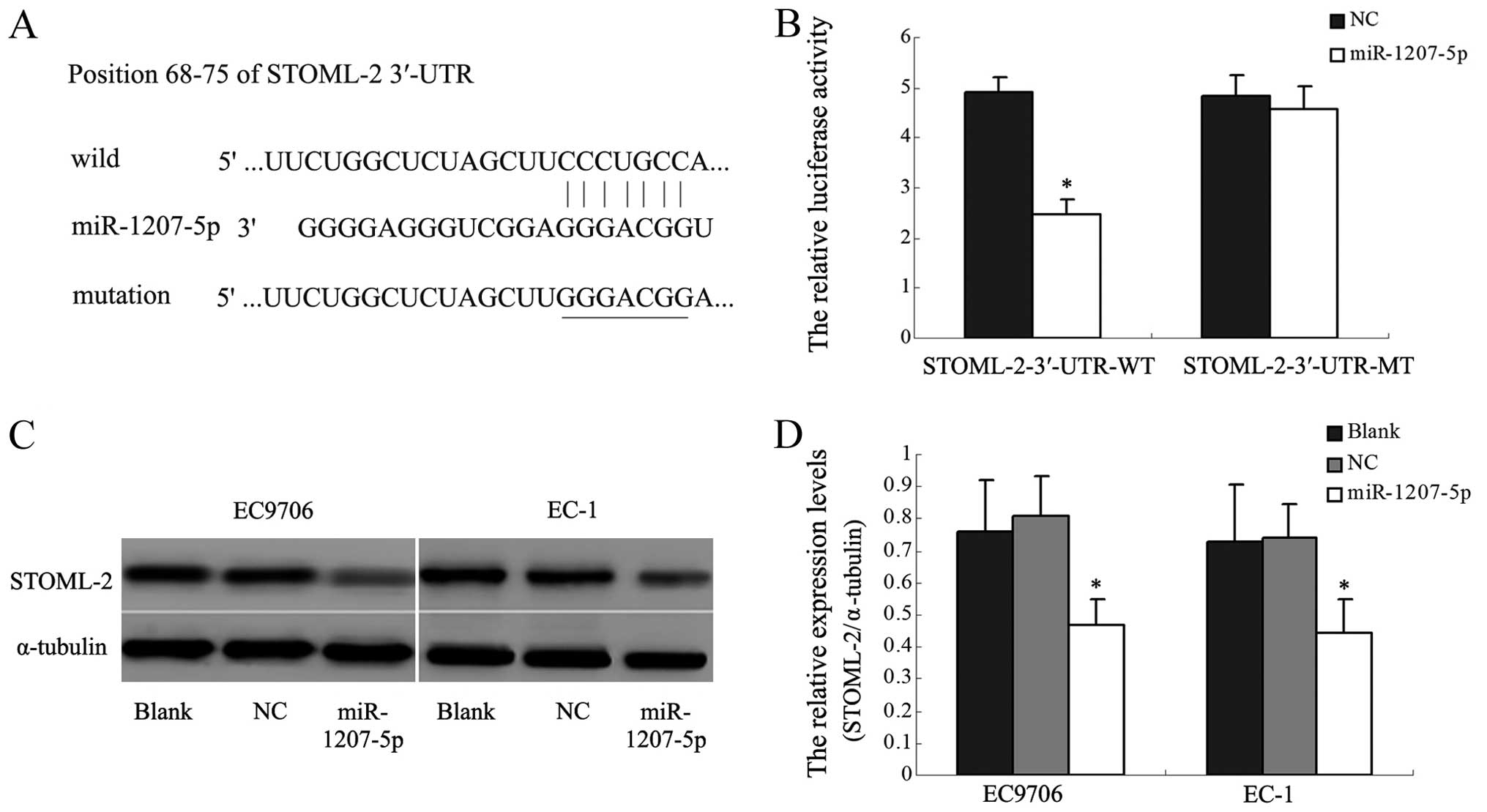
miR-1207-5p directly targets the STOML-2
gene by binding with the 3′-UTR
The TargetScan and miRBase database predicted that
the 3′-UTR of STOML-2 mRNA contained the seed region for
miR-1207-5p. To verify this prediction, we used a dual-luciferase
reporter system. The wild STOML-2 3′-UTR sequence and the mutant
STOML-2 3′-UTR sequence were inserted into the pmirGLO vector to
construct the recombinant vectors STOML-2-3′-UTR-WT and
STOML-2-3′-UTR-MT. The recombinant vectors were co-transfected with
miR-1207-5p mimic or NC into human HEK293T cells. The luciferase
activity of the reporter was decreased in the group co-transfected
with miR-1207-5p mimic and the STOML-2-3′-UTR-WT (P<0.05,
Fig. 4A and B). However, the
luciferase activity of the reporter was unaffected by
co-transfection with miR-1207-5p and the STOML-2-3′-UTR-MT. Western
blot analysis indeed showed that STOML-2 protein expression was
significantly inhibited in EC9706 and EC-1 cells transfected with
the miR-1207-5p mimic, compared with control cells (P<0.05,
Fig. 4C and D). These results
indicated that miR-1207-5p might inhibit STOML-2 expression by
binding directly to a putative seed region in the 3′-UTR of
STOML-2.
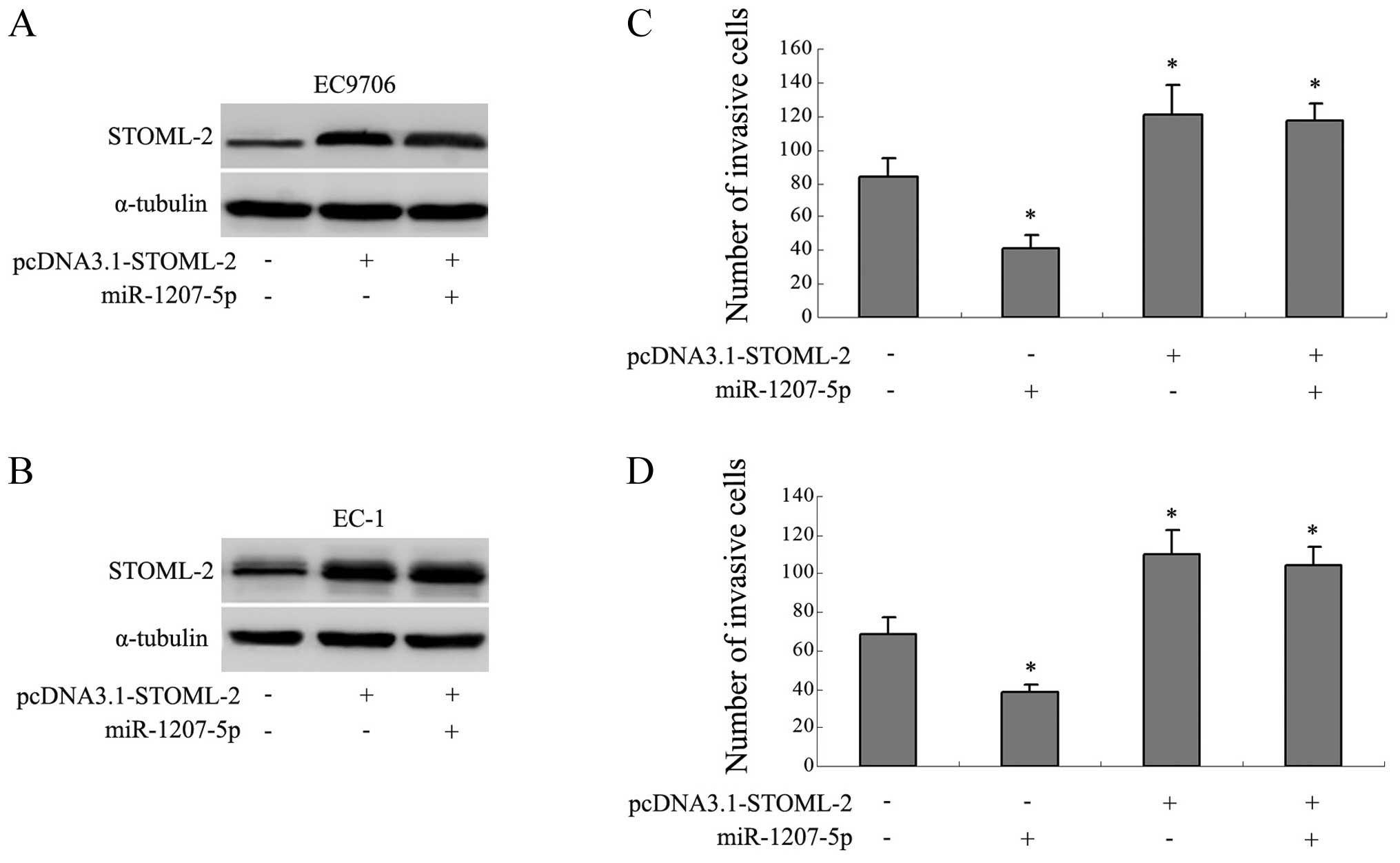
Expression of STOML-2 abrogates the
anti-invasion function of miR-1207-5p
The previous studies suggested that STOML-2 was
relevant to tumor invasion, so we constructed an expression verctor
containing the OFR of STOML-2 instead of the 3′-UTR of STOML-2
(pcDNA3.1-STOML-2) to explore whether STOML-2 was involved in the
miR-1207-5p-related invasion. We found that transfection of
pcDNA3.1-STOML-2 separately, or co-transfection with the
miR-1207-5p mimic, resulted in increased expression of STOML-2
(Fig. 5A and B). Interestingly,
the average numbers of invaded cells were correspondingly increased
in the above groups (P<0.05; Fig.
5C and D). Cells transfected with miR-1207-5p mimic separately
showed a decrease in invasive capacity compared with the control
groups (P<0.05). These results indicated that the overexpression
of STOML-2 abrogated the anti-invasion function of miR-1207-5p.
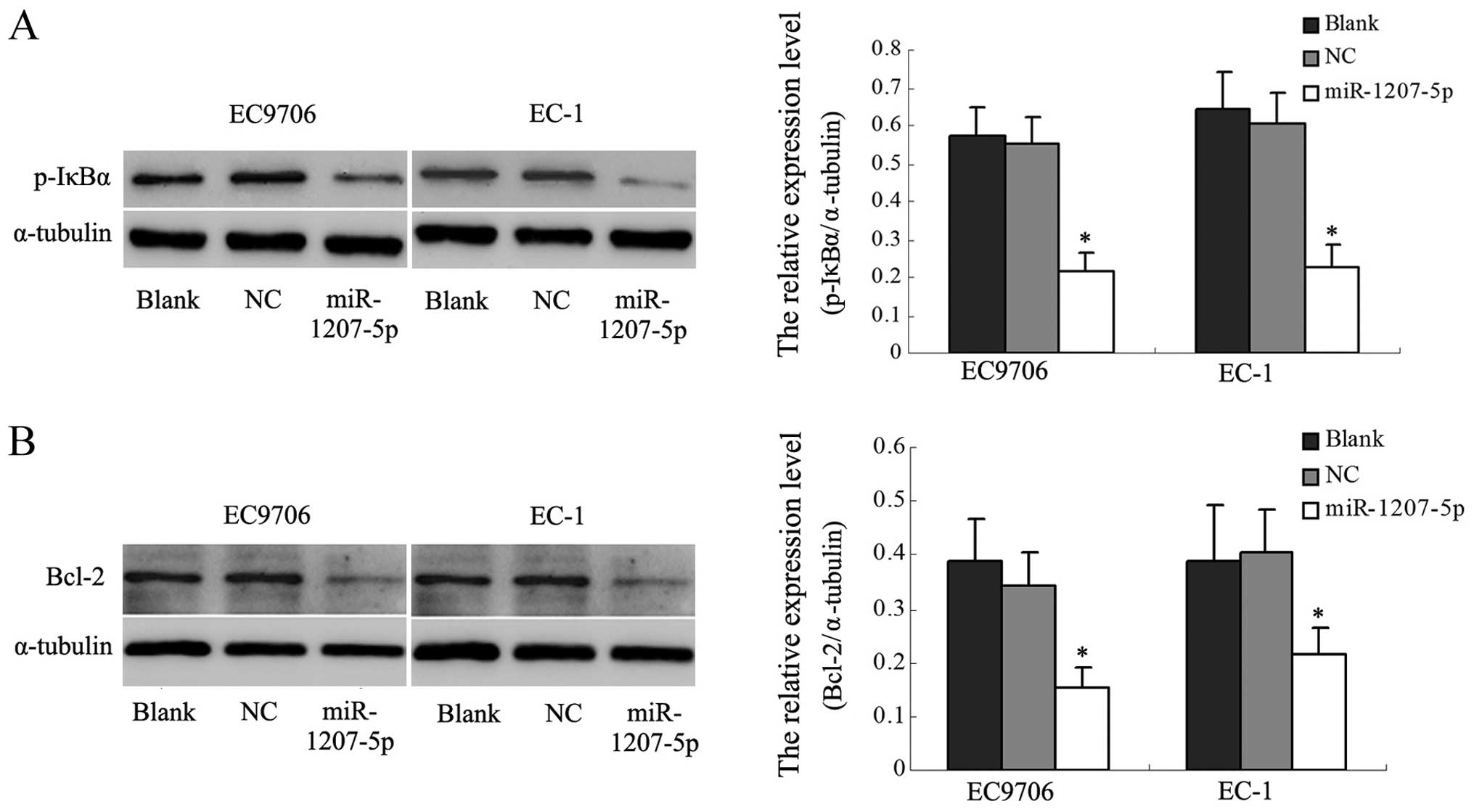
miR-1207-5p might be involved in the
NF-κB signal pathway
In the above studies, we found that upregulation of
miR-1207-5p decreased the expression of STOML-2. Other researchers
have reported that silencing STOML-2 in cells significantly
inhibited the NF-κB activity and decreased the levels of expression
of NF-κB target genes, including BcL-xL. Therefore, we examined
whether miR-1207-5p has effects on the NF-κB pathway by analyzing
the phosphorylation of the main signaling molecule, IκBα, and
Bcl-2, one of the NF-κB target genes. As shown in Fig. 6A, p-IκBα was significantly reduced
in miR-1207-5p-transfected EC cells compared with the control
cells. It was found that Bcl-2 expression was downregulated in
miR-1207-5p-transfected cells (Fig.
6B). These results suggested that downregulation of STOML-2
protein by miR-1207-5p might be involved in the NF-κB activation
pathway in EC cells.
Discussion
Ectopic miRNAs have been reported in various
cancers, and are involved in various cellular processes such as
differentiation, invasion and apoptosis (26–28).
Previous studies have indicated that miR-1207-5p was downregulated
in several cancers (11,13). Our study showed that the
expressions of mature miR-1207-5p were significantly downregulated
in central EC tissues compared with tissues from the tumor margin,
and that it might be a useful biomarker in EC. In addition, the
levels of miR-1207-5p in patients with EC were associated with
lymph node metastases, the levels of tumor differentiation and
pathological stages. To examine the effects of miR-1207-5p, we
transfected the miR-1207-5p mimic into EC9706 and EC-1 cells.
Upregulation of miR-1207-5p was observed to suppress cell
proliferation and invasion and to promote cell apoptosis. These
results implied that miR-1207-5p might be an inhibitor of EC, and
contributes to the development, progression and metastasis of
EC.
Our study showed that STOML-2 was negatively
regulated by miR-1207-5p at the posttranscriptional level by
binding to the 3′-UTR of STOML-2 mRNA in EC cells. Studies have
revealed that this interaction could decrease the expression of
STOML-2 in EC cells. Previous experiments showed that STOML-2 was
overexpressed in various tumors, such as esophageal carcinoma,
gastric adenocarcinoma, breast cancer and glioma. Levels of STOML-2
were much higher in central tumor tissues than in paired tissues,
and STOML-2-silenced cells showed decreased cell proliferation,
invasive capability and adhesive ability in vitro (25). Accordingly, the level of STOML-2
was associated with tumor metastasis in breast cancer and pulmonary
squamous cell carcinoma (29). Our
study showed that the effect of STOML-2 on EC invasion needs to be
further investigated. Our findings showed that miR-1207-5p might
act as a suppressor of metastasis by targeting STOML-2.
Abnomal activation of NF-κB has been found in many
types of cancer, including esophageal carcinoma (30–32);
however, the activation mechanisms have not been elucidated.
Accumulating publications have shown that miRNAs such as miR-146,
miR-155, miR-21, and miR-301a have pathological relevance to NF-κB
signaling (33–35). The phosphorylation of IκBα is a key
step in NF-κB pathway activation, and Bcl-2 is one of the classical
NF-κB target genes. Bcl-2 is an anti-apoptotic protein and promotes
cell survival (36,37). Previous research has shown that it
was related to the early development of EC (38,39).
We found that upregulation of miR-1207-5p significantly inhibited
STOML-2 expression and IκBα phosphorylation. The expression of
Bcl-2 was downregulated in miR-1207-5p-transfected cells. Studies
have demonstrated that depletion of STOML-2 in glioma cells reduced
NF-κB transcriptional activity (25); this finding was consistent with our
results, and indicated that STOML-2 might be involved in the
regulation of the NF-κB signaling pathway. The precise molecular
mechanism underlying miR-1207-5p/STOML-2 and the NF-κB signaling
pathway needs to be investigated further.
In conclusion, this study showed that miR-1207-5p
was downregulated in EC, and that upregulation of miR-1207-5p
suppressed cell proliferation, invasion and promotes apoptosis.
Based on these results, we propose that miR-1207-5p might act as a
potential therapeutic target in the treatment of esophageal
carcinoma.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (no. 81301726).
References
|
1
|
Lee RC and Ambros V: An extensive class of
small RNAs in Caenorhabditis elegans. Science. 294:862–864. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lau NC, Lim LP, Weinstein EG and Bartel
DP: An abundant class of tiny RNAs with probable regulatory roles
in Caenorhabditis elegans. Science. 294:858–862. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ambros V: microRNAs: Tiny regulators with
great potential. Cell. 107:823–826. 2001. View Article : Google Scholar
|
|
4
|
Hede K: Studies define role of microRNA in
cancer. J Natl Cancer Inst. 97:1114–1115. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cheng T, Hu C, Yang H, Cao L and An J:
Transforming growth factor-β-induced miR-143 expression in
regulation of non-small cell lung cancer cell viability and
invasion capacity in vitro and in vivo. Int J Oncol. 45:1977–1988.
2014.PubMed/NCBI
|
|
6
|
Chiyomaru T, Seki N, Inoguchi S, et al:
Dual regulation of receptor tyrosine kinase genes EGFR and c-Met by
the tumor-suppressive microRNA-23b/27b cluster in bladder cancer.
Int J Oncol. 46:487–496. 2015.
|
|
7
|
Pei J, Robu V, Feder M, Cheung M,
Neumann-Domer E, Talarchek J, Dulaimi E, Millenson MM and Testa JR:
Copy neutral loss of heterozygosity in 20q in chronic lymphocytic
leukemia/small lymphocytic lymphoma. Cancer Genet. 207:98–102.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Y, Li M, Zang W, Ma Y, Wang N, Li P,
Wang T and Zhao G: MiR-429 up-regulation induces apoptosis and
suppresses invasion by targeting Bcl-2 and SP-1 in esophageal
carcinoma. Cell Oncol Dordr. 36:385–394. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang J, Zhang SY, Gao YM, Liu YF, Liu YB,
Zhao ZG and Yang K: MicroRNAs as oncogenes or tumour suppressors in
oesophageal cancer: Potential biomarkers and therapeutic targets.
Cell Prolif. 47:277–286. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Z, Li J, Tian L, et al: MiRNA
expression profile reveals a prognostic signature for esophageal
squamous cell carcinoma. Cancer Lett. 350:34–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Peña-Chilet M, Martínez MT, Pérez-Fidalgo
JA, et al: MicroRNA profile in very young women with breast cancer.
BMC Cancer. 14:5292014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mun J, Tam C, Chan G, Kim JH, Evans D and
Fleiszig S: MicroRNA-762 is upregulated in human corneal epithelial
cells in response to tear fluid and Pseudomonas aeruginosa antigens
and negatively regulates the expression of host defense genes
encoding RNase7 and ST2. PLoS One. 8:e578502013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen L, Lü MH, Zhang D, et al: miR-1207-5p
and miR-1266 suppress gastric cancer growth and invasion by
targeting telomerase reverse transcriptase. Cell Death Dis.
5:e10342014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Papagregoriou G, Erguler K, Dweep H,
Voskarides K, Koupepidou P, Athanasiou Y, Pierides A, Gretz N,
Felekkis KN and Deltas C: A miR-1207-5p binding site polymorphism
abolishes regulation of HBEGF and is associated with disease
severity in CFHR5 nephropathy. PLoS One. 7:e310212012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Alvarez ML, Khosroheidari M, Eddy E and
Kiefer J: Role of microRNA 1207-5P and its host gene, the long
non-coding RNA Pvt1, as mediators of extracellular matrix
accumulation in the kidney: Implications for diabetic nephropathy.
PLoS One. 8:e774682013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y and Morrow JS: Identification and
characterization of human SLP-2, a novel homologue of stomatin
(band 7.2b) present in erythrocytes and other tissues. J Biol Chem.
275:8062–8071. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Owczarek CM, Treutlein HR, Portbury KJ,
Gulluyan LM, Kola I and Hertzog PJ: A novel member of the
STOMATIN/EPB72/mec-2 family, stomatin-like 2 (STOML2), is
ubiquitously expressed and localizes to HSA chromosome 9p13.1.
Cytogenet Cell Genet. 92:196–203. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lapatsina L, Brand J, Poole K, Daumke O
and Lewin GR: Stomatin-domain proteins. Eur J Cell Biol.
91:240–245. 2012. View Article : Google Scholar
|
|
19
|
Liu Z, Yang Y, Zhang Y, Ye X, Wang L and
Xu G: Stomatin-like protein 2 is associated with the
clinicopathological features of human papillary thyroid cancer and
is regulated by TGF-β in thyroid cancer cells. Oncol Rep.
31:153–160. 2014.
|
|
20
|
Cao W, Zhang B, Liu Y, et al: High-level
SLP-2 expression and HER-2/neu protein expression are associated
with decreased breast cancer patient survival. Am J Clin Pathol.
128:430–436. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cui Z, Zhang L, Hua Z, Cao W, Feng W and
Liu Z: Stomatin-like protein 2 is overexpressed and related to cell
growth in human endometrial adenocarcinoma. Oncol Rep. 17:829–833.
2007.PubMed/NCBI
|
|
22
|
Zhang L, Ding F, Cao W and Liu Z, Liu W,
Yu Z, Wu Y, Li W, Li Y and Liu Z: Stomatin-like protein 2 is
overexpressed in cancer and involved in regulating cell growth and
cell adhesion in human esophageal squamous cell carcinoma. Clin
Cancer Res. 12:1639–1646. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cao W, Zhang B, Li J, Liu Y, Liu Z and Sun
B: SLP-2 overexpression could serve as a prognostic factor in node
positive and HER2 negative breast cancer. Pathology. 43:713–718.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cao W, Zhang B, Ding F, Zhang W, Sun B and
Liu Z: Expression of SLP-2 was associated with invasion of
esophageal squamous cell carcinoma. PLoS One. 8:e638902013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Song L, Liu L, Wu Z, Lin C, Dai T, Yu C,
Wang X, Wu J, Li M and Li J: Knockdown of stomatin-like protein 2
(STOML2) reduces the invasive ability of glioma cells through
inhibition of the NF-κB/MMP-9 pathway. J Pathol. 226:534–543. 2012.
View Article : Google Scholar
|
|
26
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Adams BD, Kasinski AL and Slack FJ:
Aberrant regulation and function of microRNAs in cancer. Curr Biol.
24:R762–R776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bouyssou JM, Manier S, Huynh D, Issa S,
Roccaro AM and Ghobrial IM: Regulation of microRNAs in cancer
metastasis. Biochim Biophys Acta. 1845:255–265. 2014.PubMed/NCBI
|
|
29
|
Chang D, Ma K, Gong M, Cui Y, Liu ZH, Zhou
XG, Zhou CN and Wang TY: SLP-2 overexpression is associated with
tumour distant metastasis and poor prognosis in pulmonary squamous
cell carcinoma. Biomarkers. 15:104–110. 2010. View Article : Google Scholar
|
|
30
|
Gasparini C, Celeghini C, Monasta L and
Zauli G: NF-kappaB pathways in hematological malignancies. Cell Mol
Life Sci. 71:2083–2102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tong L, Yuan Y and Wu S: Therapeutic
microRNAs targeting the NF-kappa B signaling circuits of cancers.
Adv Drug Deliv Rev. 81:1–15. 2015. View Article : Google Scholar
|
|
32
|
Abdel-Latif MM, O’Riordan J, Windle HJ,
Carton E, Ravi N, Kelleher D and Reynolds JV: NF-kappaB activation
in esophageal adenocarcinoma: Relationship to Barrett’s metaplasia,
survival, and response to neoadjuvant chemoradiotherapy. Ann Surg.
239:491–500. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gong H, Song L, Lin C, Liu A, Lin X, Wu J,
Li M and Li J: Downregulation of miR-138 sustains NF-κB activation
and promotes lipid raft formation in esophageal squamous cell
carcinoma. Clin Cancer Res. 19:1083–1093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bhaumik D, Scott GK, Schokrpur S, Patil
CK, Campisi J and Benz CC: Expression of microRNA-146 suppresses
NF-kappaB activity with reduction of metastatic potential in breast
cancer cells. Oncogene. 27:5643–5647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kumar V, Palermo R, Talora C, et al: Notch
and NF-κB signaling pathways regulate miR-223/FBXW7 axis in T-cell
acute lymphoblastic leukemia. Leukemia. 28:2324–2335. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hardwick JM, Chen YB and Jonas EA:
Multipolar functions of BCL-2 proteins link energetics to
apoptosis. Trends Cell Biol. 22:318–328. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wood WG, Igbavboa U, Muller WE and Eckert
GP: Statins, Bcl-2, and apoptosis: Cell death or cell protection?
Mol Neurobiol. 48:308–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sarbia M, Bittinger F, Porschen R, Verreet
P, Dutkowski P, Willers R and Gabbert HE: bcl-2 expression and
prognosis in squamous-cell carcinomas of the esophagus. Int J
Cancer. 69:324–328. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Torzewski M, Sarbia M, Heep H, Dutkowski
P, Willers R and Gabbert HE: Expression of Bcl-X(L), an
antiapoptotic member of the Bcl-2 family, in esophageal squamous
cell carcinoma. Clin Cancer Res. 4:577–583. 1998.PubMed/NCBI
|