Introduction
Gastric cancer is the second most common form of
cancer in the world. Therapeutic surgical techniques are improving
and some chemotherapeutic regimens are available, but the outcomes
of patients with high grade gastric cancer are usually poor
(1).
Activation of PI3K generates second messenger PIP3.
The colocalization of PIP3 with Akt and PDK1 invokes the
phosphorylation of Akt Ser308 (2).
The PI3K/AKT signaling pathway is an important part of
intracellular signal transduction, cell proliferation,
differentiation, apoptosis and migration. The PI3K/AKT signaling
pathway has been implicated in a variety of tumor growth and
metastasis (3). For example,
oncogenic activation of PI3K/Akt molecules enhances cell
proliferation by increasing Cyclin D1 levels (4–6). It
is well known that the aberrant expression of Cyclin D1 and CDK4
proteins is involved in the proliferation of CRC cells (7). Suppression of PI3K/Akt leads to the
blockade of cell proliferation and demonstrates the importance of
these signaling cascades in the control of both cell cycle
progression and cell growth during cancer development (8). Therefore, using the PI3K inhibitors
in cancer therapy is considered to be a very promising solution to
tumor treatment. Recent years have seen an explosion in the number
of phosphoinositide 3-kinase (PI3K) pathway inhibitors under
clinical investigation (9).
Fangchinoline is the main chemical constituent of
Stephania tetrandra S. Moore, which has been shown to
possess a wide range of pharmacological activities (10), including inhibition of histamine
release and antihypertensive activities (11,12),
antiinflammatory effects (13–15),
antiplatelet aggregation activities (16), antihyperglycemic actions (17,18),
neuroprotective effects (19), and
antioxidant and radical scavenging activities (20,21).
Another pharmacological activity is a wide spectrum of antitumor
activity in various cancer cells, the potent antitumor activity of
tetrandrine has been extensively investigated with its proposed
mechanism of inducing G1/S and G2/M arrest
and stimulating apoptotic cell death (22–24).
However, there are not many reports of the antitumor activity of
fangchinoline and its underlying mechanism. Experiments have showed
that fangchinoline inhibits cell proliferation via Akt/Gsk3β/Cyclin
D1 signaling induces apoptosis in breast cancer cell lines and
induces autophagic cell death via p53/sestrin2/AMPK signaling in
human hepatocellular carcinoma cells (25–28).
Here we report that fangchinoline effectively suppressed the
proliferation and invasion of gastric cancer cells SGC7901 and
BGC823 and promoted their early apoptosis. Importantly, we provide
a novel mechanism that fangchinoline targets PI3K, which promotes
tumor cell survival and invasion by suppressing the phosphorylation
of Akt (Ser308). Our evidence suggests that fangchinoline is a
potential anticancer drug as the natural inhibitor of PI3K.
Materials and methods
Cell culture
Human gastric cancer cell lines MKN45, SGC7901 and
HEK293 cells (as the control) were cultured in DMEM (Invitrogen)
supplemented with 10% fetal calf serum (Invitrogen) at 37°C in
incubator with humidified atmosphere of 5% CO2 and 95%
air.
MTT assays
Human cancer cells (1×104/well) were
plated in 0.1 ml of the medium containing 10% FBS in 96-well
plates; 24 h later, the medium was removed and replaced with 0.1 ml
medium containing the indicated concentrations of fangchinoline and
incubated for 24, 36, 48 and 60 h. At the end of the incubation,
the capability of cellular proliferation was measured by the
modified tetrazolium salt-3-(4-5
dimethylthiazol-2-yl)-2-5-diphenyltetrazolium bromide (MTT) assay.
For this, 0.01 ml of MTT solution (5 mg/ml in PBS) was added to
each well. After a 4-h incubation at 37°C, medium was replaced by
0.15 ml DMSO. After 15-min incubation at 37°C, the optical
densities at 490 nm were measured using a Microplate Reader
(Bio-Rad).
Cell-cycle analysis by flow
cytometry
SGC7901 cells were incubated with the indicated
concentrations of fangchinoline for 24 h. After incubation, cells
were collected, washed with PBS and then suspended in a staining
buffer (10 μg/ml propidium iodide, 0.5% Tween-20, 0.1% RNase in
PBS). The cells were analyzed using a FACS Vantage flow cytometer
with the CellQuest acquisition and analysis software program
(Becton-Dickinson Co., San Jose, CA, USA). Gating was set to
exclude cell debris, doublets and clumps.
Cell migration and invasion assay
Migration and invasion assays were performed using
modified boyden chambers with polycarbonate nucleopore membrane.
Precoated filters (6.5 mm in diameter, 8-μm pore size, Matrigel 100
μg/cm2) were rehydrated with 100 μl medium. Then,
1×105 cells in 100 μl serum-free DMEM supplemented with
0.1% bovine serum albumin were placed in the upper part of each
chamber, whereas the lower compartments were filled with 600 μl
DMEM containing 10% serum. After incubation for 18 h at 37°C,
non-invaded cells were removed from the upper surface of the filter
with a cotton swab, and the invaded cells on the lower surface of
the filter were fixed, stained, photographed and counted under
high-power magnification.
Cell apoptosis
Following Annexin V-V-FITC apoptosis detection kit
instructions, the specific steps were: cells were washed twice with
cold PBS, then re-suspended with binding buffer cells at a
concentration of 1×106 cells/ml. Adding 5 μl of Annexin
V-FITC and 10 μl of PI. Cells were incubated in the dark, at room
temperature, for 15 min. Then, 400 μl binding buffer was added to
each tube and the apoptosis rate was measured by flow cytometry
within 1 h.
Hoechst 33258 staining
SGC7901 cells were incubated with the indicated
concentrations of fangchinoline for 24 h. After incubation, cells
were fixed with 4% polyoxymethylene, then washed twice with PBS,
incubated with 10 μg/ml Hoechst 33258 for 5 min at room
temperature, then washed with PBS 3 times. Cells were observed with
fluorescence microscope.
Mitochondrial membrane potential
Cells (1×105) were cultured in 6-well
plates for the assay, then collected, centrifuged and re-suspended
in 0.5 ml DMEM medium. The cells were washed twice in staining
buffer and then incubated in 0.5 ml JC-1 staining buffer, at room
temperature, in the dark. Flow cytometry was used to determine the
fluorescence intensity of the red/green ratio
semi-quantitatively.
Reverse transcription and quantitative
real-time PCR
Total cellular RNA from DMSO and fangchinoline
treated SGC7901 cells were extracted after 24 h using TRIzol
(Invitrogen) according to the manufacturer’s protocol. One
microgram of total RNA was reverse transcribed to cDNA in a total
volume of 20 μl system using a RT reaction kit (Promega). Real-time
PCR was performed using an Mx 3000P real-time PCR system (Applied
Biosystems) according to the manufacturer’s instructions and SYBR
Premix Ex Taq (Takara) as a DNA-specific fluorescent dye. PCR was
carried out for 50 cycles of 95°C for 10 sec and 60°C for 30 sec.
Primer sequences for detection of mRNA expression were synthesized
(Table I). All the reactions were
repeated at least three times. Gene expression levels were
calculated relative to the housekeeping β-actin by using Stratagene
Mx 3000P software.
 | Table IPrimer sequences for detection of mRNA
expression. |
Table I
Primer sequences for detection of mRNA
expression.
| Name | Forward primer
(5′→3′) | Reverse primer
(5′→3′) |
|---|
| Gsk3β |
GTATGGTCTGCTGGCTGTGT |
GGGTCGGAAGACCTTAGTCC |
| CDK2 |
GCCATTCTCATCGGGTCCTC |
ATTTGCAGCCCAGGAGGATT |
| Caspase-9 |
GGTGACCCCAGAATTGACCC |
TCGACAACTTTGCTGCTTGC |
| Caspase-3 |
TGTGAGGCGGTTGTAGAAGTT |
GCTGCATCGACATCTGTACC |
| Bcl-2 |
GGTGAACTGGGGGAGGATTG |
GGCAGGCATGTTGACTTCAC |
| Bax |
AGCTGAGCGAGTGTCTCAAG |
GTCCAATGTCCAGCCCATGA |
| MMP2 |
CGCATCTGGGGCTTTAAACAT |
TCAGCACAAACAGGTTGCAG |
| MMP9 |
CGACGTCTTCCAGTACCGAG |
TTGTATCCGGCAAACTGGCT |
| β-actin |
TCGTGCGTGACATTAAGGAG |
ATGCCAGGGTACATGGTGGT |
Western blot analyses
To determine the expression of protein, whole cell
extracts (lysate) were prepared from 1×106 cells in
lysis buffer (20 mM Tris pH 7.4, 250 mM sodium chloride, 0.1%
Triton-X-100, 2 mM EDTA, 10 μg/ml leupeptin, 10 μg/ml aprotinin,
0.5 mM phenylmethylsulfonyl fluoride, 4 mM sodium orthovanadate and
1 mM DTT), and 60 μg of the protein was resolved on 10%
SDS-polyacrylamide gels. After electrophoresis, the proteins were
eletrotransferred to nitrocellulose filters, the membrane
(Amersham) was blocked with 5% non-fat dry milk in TBS-T (20 mM
Tris, pH 7.6, 137 mM NaCl, 0.05% Tween-20) for 1 h at room
temperature, and the proteins were probed with specific
antibodies-Gsk3β, CDK2, MMP2, MMP9 (Bioworld), Akt, phospho-Akt
(Ser308) (Santa Cruz), caspase-3, caspase-9, Bax and Bcl-2
(Neomarker). To assure equal loading, gels were stripped and
reprobed with antibodies against GAPDH (Kangchen Bio-tech Inc.,
Shanghai, China). All PVDF membranes were detected by
chemiluminescence (ECL, Pierce Technology).
Results
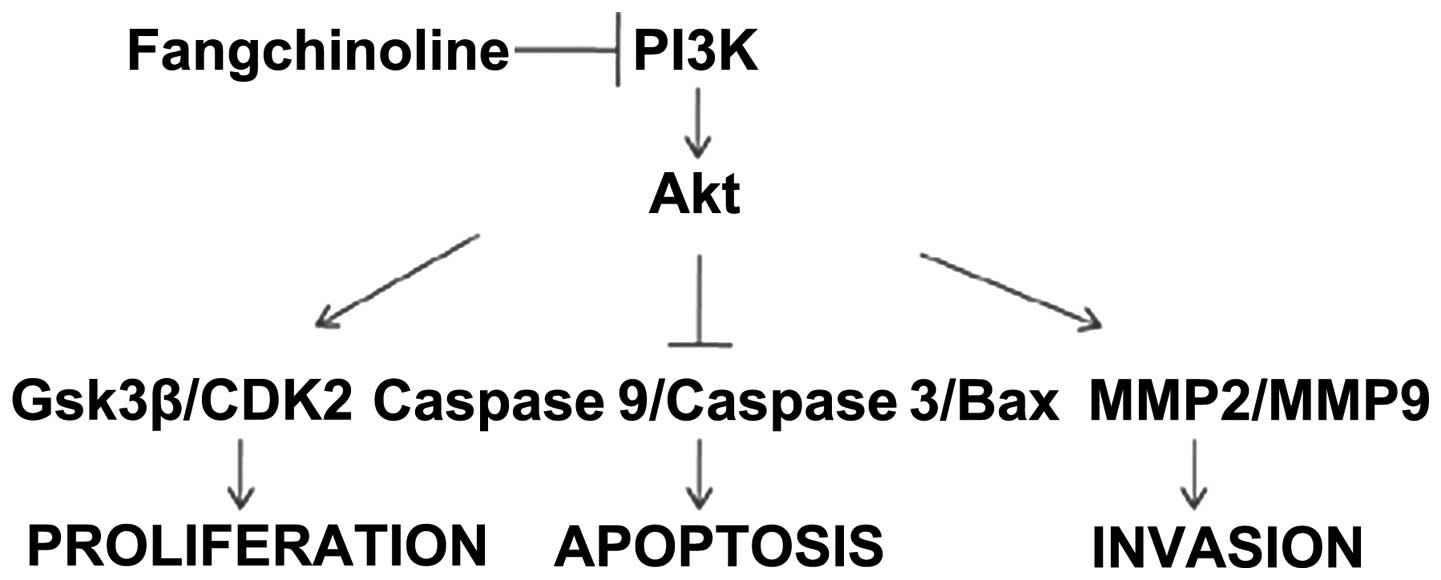
Fangchinoline inhibits the the expression
of PI3K
MKN45, SGC7901 and HEK293 cells were used to detect
the inhibitory effect of fangchinoline on growth of these cells. As
shown in MTT assay, fangchinoline treatment inhibited the
proliferation of SGC7901 cells in a concentration-dependent manner
but have little effect on other cells (Fig. 1B). Since proteins regulating
signaling through the phosphatidylinositol 3-kinase (PI3K)/Akt
pathway is frequently altered in human cancer, including gastric
cancer (29), the expression level
of PI3K in gastric cancer cell lines was examined. Interestingly,
the protein and mRNA levels of PI3K were dramatically higher in
SGC7901 cells than that in MKN45 cells and HEK-293 cells (Fig. 1C and D) indicating PI3K might be
targeted by fangchinoline and be involved in fangchinoline-induced
growth inhibition of gastric cancer cells. Furthermore, we examined
whether fangchinoline inhibited the PI3K in SGC7901 cells, and
found that Fangchinoline at 20 μmol/l markedly inhibited the level
of PI3K (Fig. 1E and F).
 | Figure 1Fangchinoline inhibits the the
expression of FAKPI3K. (A) The structure of fangchinoline. (B)
MKN45, SGC7901 and HEK293 cells were cultured with indicated
concentrations of fangchinoline for indicated hours in 96-well
plates, then MTT assay was performed, results represent the mean ±
SD of three experiments done in triplicate. (C) Total proteins of
gastric cancer cell lines or HEK-293 cell line were extracted to
detect PI3K level. (D) Total mRNA expression of PI3K in MKN45,
SGC7901 and HEK293 were detected by real-time RT-PCR, results
represent the mean ± SD of three experiments done in triplicate.
(E) SGC7901 cells were treated with DMSO alone or indicated
concentration of fangchinoline for 48 h, proteins were extracted
and subjected to western blot analysis, the membrane was probed
sequentially with PI3K antibody. (F) SGC7901 cells were treated
with DMSO alone or indicated concentrations of fangchinoline for 48
h, cells were harvested, and the mRNA expression of PI3K was
detected by real-time RT-PCR, results represent the mean ± SD of
three experiments done in triplicate. |
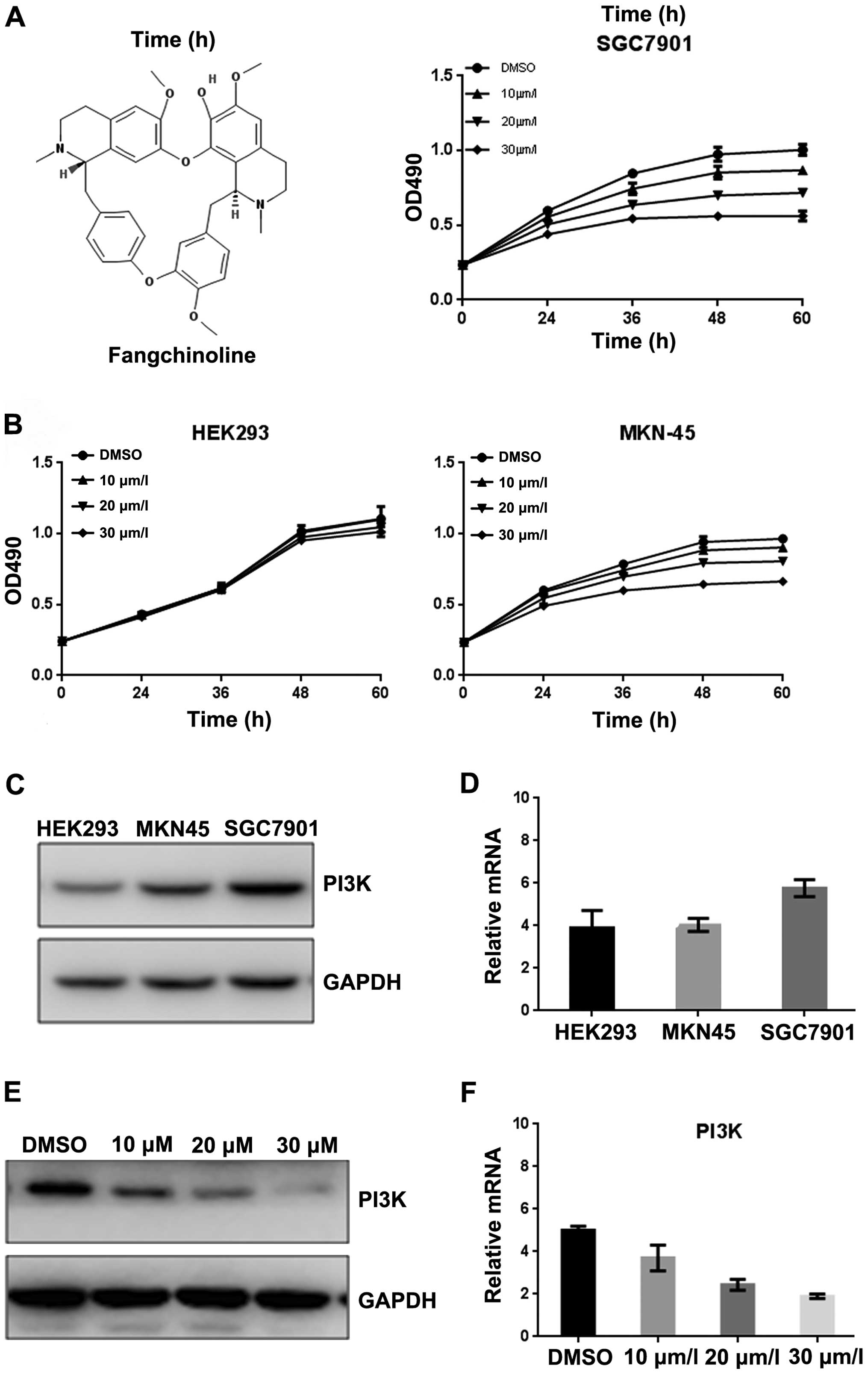
Fangchinoline inhibits the proliferation
of SGC7901 by inhibiting PI3K/Akt pathway
To further investigate the mechanisms of
fangchinoline inhibition of growth of gastric cancer, the SGC7901
cells were exposed to various concentrations of fangchinoline for
24 h, and then cell cycle analysis was performed. Fangchinoline
prominently induced a dose-dependent increase in the percentage of
cells in G1 phase and decrease in S phase compared with
the control (Fig. 2A), indicating
that fangchinoline arrested SGC7901 cells at the G1
phase of the cell cycle. Since Gsk3β and CDK2 are key regulators in
the G1 phase of the cell cycle, we examined the
indicated regulator expression level in fangchinoline-treated
cells. Western blot analysis showed that exposure of SGC7901 to
10/20/30 μmol/l fangchinoline for 48 h dramatically decreased
protein expression of Gsk3β and CDK2 (Fig. 2B), indicating fangchinoline arrests
cells at G1 phase and then suppresses cell growth via
downregulated Gsk3β and CDK2. Furthermore, real-time RT-PCR showed
that expression of Gsk3β and CDK2 in SGC7901 was downregulated at
mRNA level after exposure to fangchinoline (Fig. 2C). Western blot analysis showed
that Aktp-Ser308 was downregulated in a dose-dependent
manner without affecting its total expression (Fig. 2D).
 | Figure 2Fangchinoline inhibits the
proliferation of gastric cancer cells by inhibiting the PI3K/Akt
pathway. (A) SGC7901 cells were pre-incubated with DMSO or
fangchinoline for 48 h, then cells were analyzed using a FACS
vantage flow cytometer with the CellQuest acquisition and analysis
software program, the experiment was repeated three times. (B)
SGC7901 cells were treated with DMSO alone or indicated
concentration of fangchinoline for 48 h, the protein expression of
Gsk3β and CDK2 were detected by western blot analysis. (C) SGC7901
cells were treated with DMSO alone or indicated concentration of
fangchinoline for 48 h, cells were harvested, and the mRNA
expression of Gsk3β and CDK2 were detected by real-time RT-PCR,
results represent the mean ± SD of three experiments done in
triplicate. (D) SGC7901 cells were treated with DMSO alone or
indicated concentration of fangchinoline for 24 h, the protein
expression of Gsk3β, CDK2, Aktp-Ser308, Akt, and PI3K
were detected by western blot analysis. |
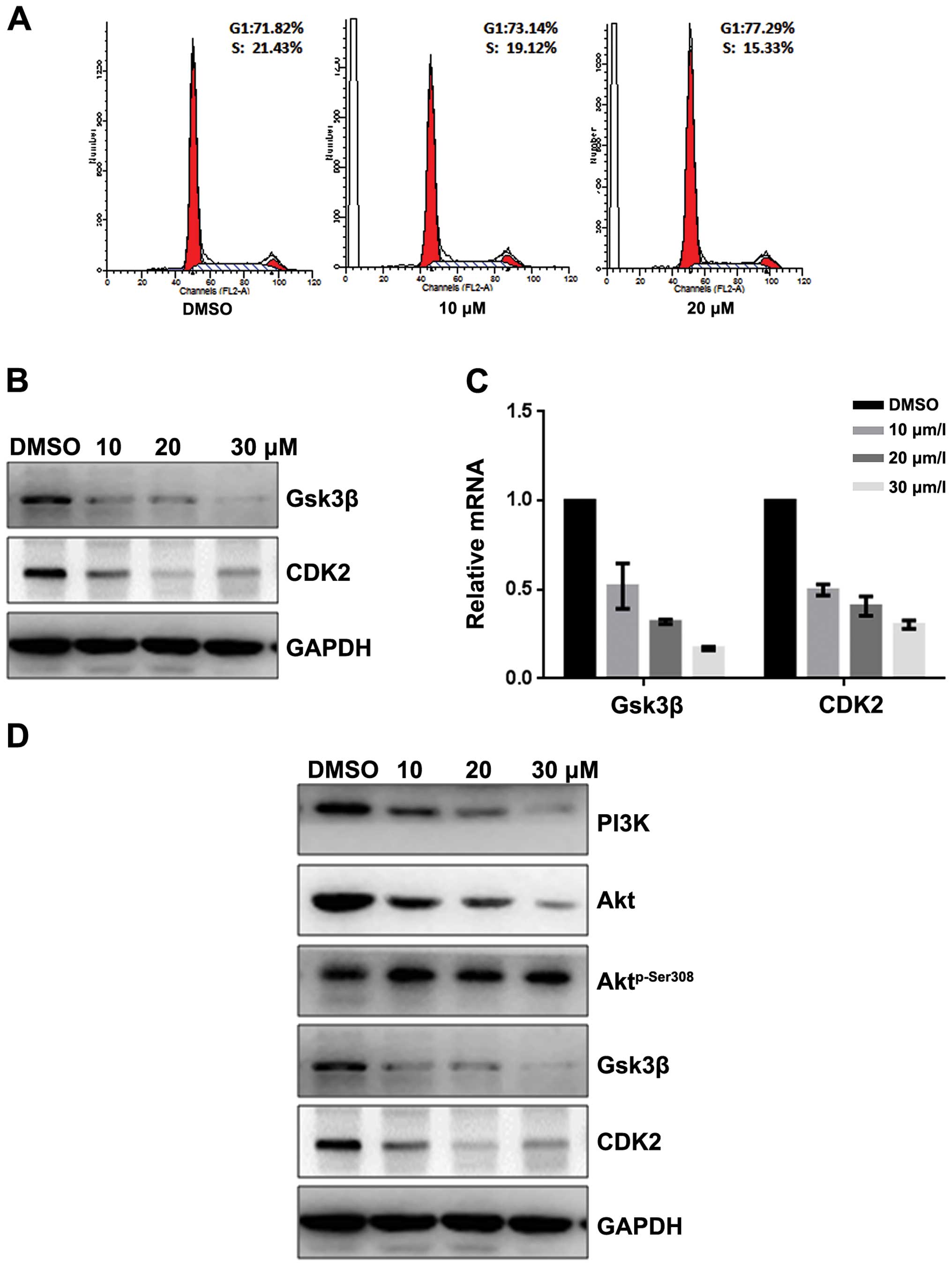
Fangchinoline induces apoptosis of
SGC7901 by inhibiting the PI3K/Akt pathway
To evaluate whether fangchinoline induces apoptosis
of SGC7901 cells, we detected the apoptosis rate by Hoechst 33258
staining and AV-PI. Hoechst 33258 staining was performed to observe
the fangchinoline-induced apoptotic nucleus of SGC7901 cells.
Condensed chromatin was observed in fangchinoline-treated SGC7901
cells (Fig. 3A). By Annexin V-FITC
staining, the fangchinoline-induced SGC7901 cell apoptosis was
increased compared to that of the control cells (Fig. 3B). The loss of mitochondrial
membrane potential (ΔΨm) is regarded as one of the early events in
the apoptotic pathway, which can trigger the release of cytochrome
c and other apoptosis related molecules after induction by
various stimuli. To detect the change of the mitochondrial membrane
potential, JC-1 was used to stain the cells and then analyzed them
through flow cytometry. Results showed that the number of cells
with loss of ΔΨm increased after treatment with fangchinoline
(Fig. 3C). Then real-time RT-PCR
showed that expression of caspase-3, caspase-9 and Bax in SGC7901
were upregulated at mRNA level and Bcl-2 was downregulated at mRNA
level after exposure to fangchinoline (Fig. 3D). Furthermore the expression of
apoptosis regulators was examined by western blot analysis. The
expression of Bcl-2 and PI3K was obviously decreased and the levels
of caspase-3, caspase-9 and Bax were increased in fangchinoline
treated SGC7901 cells, and Aktp-Ser308 was dramatically
downregulated without changing the expression of Akt (Fig. 3E).
 | Figure 3Fangchinoline induces apoptosis of
gastric cancer cells through inhibiting the PI3K/Akt pathway. (A)
SGC7901 cells were pre-incubated with fangchinoline for 48 h then
cells were stained with Hoechst 33258, and observed with
fluorescence microscope. (B) SGC7901 cells were pre-incubated with
fangchinoline for 48 h then cells were treated with Annexin V-FITC
apoptosis detection kit and analyzed with FCAS. The experiment was
repeated three times. (C) SGC7901 cells were pre-incubated with
fangchinoline for 48 h, and then cells were stained with JC-1 and
analyzed by flow cytometry. Cell percentage of Q4 phase indicating
loss of mitochondrial membrane potential of the three experiments
analyzed. (D) SGC7901 cells were treated with DMSO alone or
indicated concentration of fangchinoline for 48 h, cells were
harvested, and the mRNA expression of caspase-3, caspase-9, Bax and
Bcl-2 were detected by real-time RT-PCR, results represent the mean
± SD of three experiments in triplicate. (E) SGC7901 cells were
treated with DMSO alone or indicated concentration of fangchinoline
for 48 h, the protein expression of PI3K, caspase-3, caspase-9,
Bax, Bcl-2 Akt, and Aktp-Ser308 were detected by western
blot analysis. |
Fangchinoline represses the migratory and
invasive potential of SGC7901 by inhibiting the PI3K/Akt
pathway
Inhibitory effect of fangchinoline on migration and
invasion of MKN45 and SGC7901 cells were analyzed by Τranswell
assay (with or without Μatrigel). Results showed that fangchinoline
significantly decreased invasion and migration potential of gastric
cancer SGC7901 cells (Fig. 4A and
B) in a dose-dependent manner, but weakly decreased invasion
and migration potential of MKN45 cells (Fig. 4C and D). Real-time RT-PCR showed
that expression of MMP2 and MMP9 in SGC7901 was downregulated at
mRNA level after exposure to fangchinoline (Fig. 4E). Western blot analysis showed
that exposure of SGC7901 to fangchinoline (10/20/30 μmol/l) for 48
h dramatically decreased levels of MMP2, MMP9, PI3K and
Aktp-Ser308 but had little effect on Akt (Fig. 4F). These results indicated that
fangchinoline effectively suppressed proliferation and invasion of
SGC7901 by inhibiting the PI3K/Akt pathway (Fig. 5).
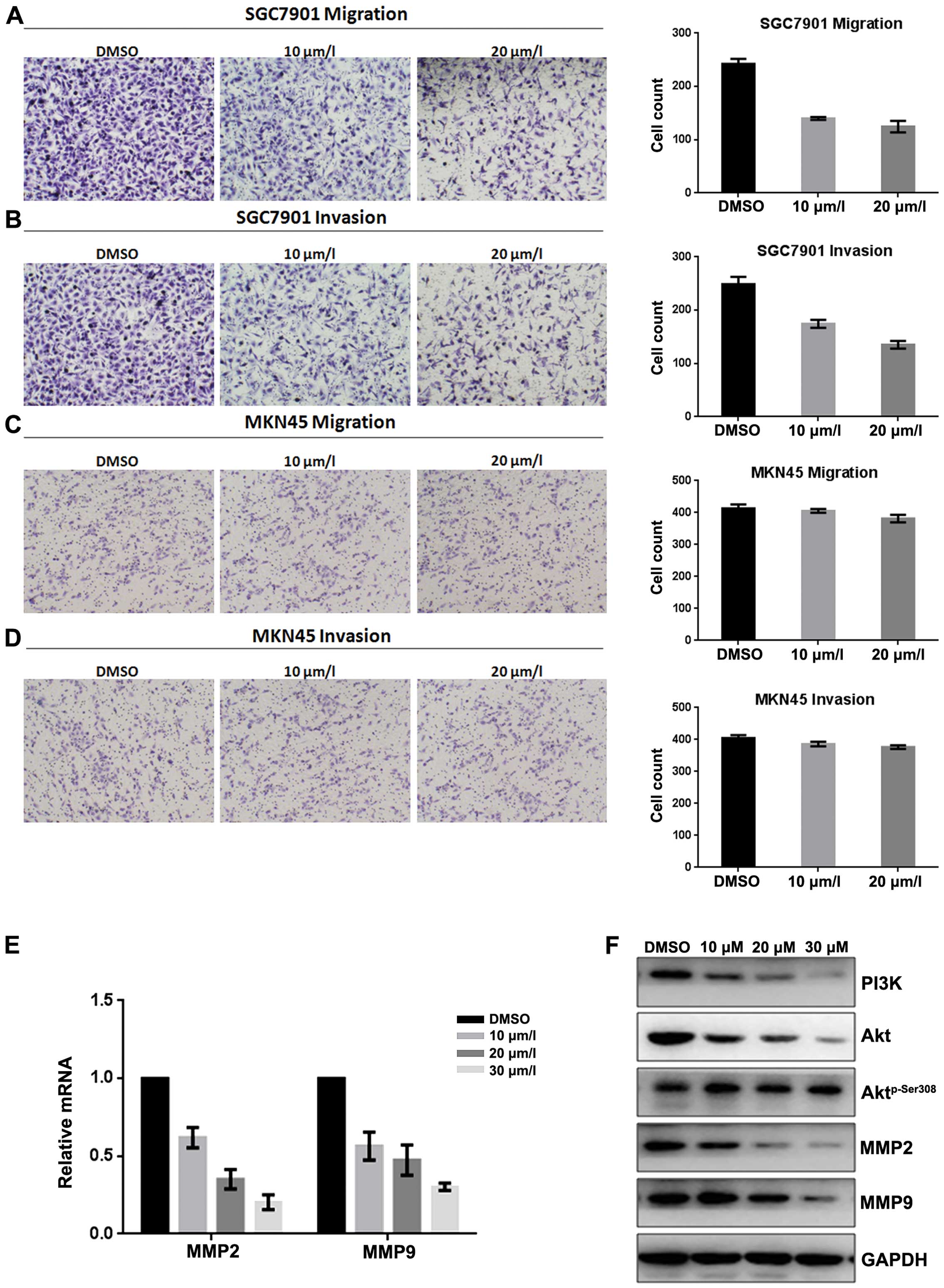 | Figure 4Fangchinoline represses the migratory
and invasive potential of gastric cancer cells by inhibiting the
PI3K/Akt pathway. (A) SGC7901 cells were pre-incubated with
fangchinoline for 48 h, Transwell assay without Μatrigel was
performed. Cells were counted and results represent the mean ± SD
of three experiments. (B) SGC7901 cells were pre-incubated with
fangchinoline for 48 h, Τranswell assay with Μatrigel was
performed. Cells were counted and results represent the mean ± SD
of three experiments. (C) MKN45 cells were pre-incubated with
fangchinoline for 48 h, Τranswell assay without Μatrigel was
performed. Cells were counted and results represent the mean ± SD
of three experiments. (D) MKN45 cells were pre-incubated with
fangchinoline for 24 h, Τranswell assay with Μatrigel was
performed. Cells were counted and results represent the mean ± SD
of three experiments. (E) SGC7901 cells were treated with DMSO
alone or indicated concentrations of fangchinoline for 48 h, cells
were harvested, and the mRNA expression of MMP2 and MMP9 were
detected by real-time RT-PCR, results represent the mean ± SD of
three experiments in triplicate. (F) SGC7901 cells were treated
with DMSO alone or indicated concentration of fangchinoline for 48
h, the protein expression of PI3K, Akt, Aktp-Ser308,
MMP2 and MMP9 were detected by western blot analysis. |
Discussion
Fangchinoline inhibits cell proliferation and
induces apoptosis as an antitumor agent in several cancer cell
lines, such as MDA-MB-231 and HepG2 cells (25–28).
However, the effects of fangchinoline on gastric cancer cells have
not been previously reported. Our data show fangchinoline treatment
inhibited the proliferation, migration, and invasion of SGC7901
cells in a concentration-dependent manner but had little effect on
MKN45 cell lines or the control cell line HEK293. In elucidating
the mechanism, we found high expression of PI3K in SGC7901 cell
lines but only slight expression in MKN45 cells and the control
HEK293 cells. Interestingly, we found fangchinoline could suppress
the PI3K in SGC7901 cells, which implies that fangchinoline targets
PI3K in tumor cells that highly express PI3K and inhibits their
proliferation, migration, and invasion.
PI3K is considered as a key regulator in cancer cell
signaling. It has been reported that the inhibition of PI3K is
important in tumor treatment. LY294002 can effectively change the
microvascular permeability, reducing fluid pressure in the tumor
stroma (30). PI-103 can not only
inhibit PI3K, but it also inhibits mTOR and DNA-dependent protein
kinase, a feature that has been used in a variety of in vivo
efficacy models, and can even have a certain effect on
glioblastomas (31). In some joint
drug tests, ATP-competitive inhibition of PI3K showed good
tolerability and higher activity, which can improve the efficacy of
other anticancer drugs (32). In
our study, PI3K level was markedly decreased at 20 μmol/l
concentration of fangchinoline in SGC7901. Taken together, these
results indicated fangchinoline acted as a novel inhibitor of PI3K
and suppressed SGC7901 cell line proliferation via PI3K.
It has been recognized that control of cell cycle
progression in cancer cells is an effective strategy to inhibit
tumor growth (33,34). The phosphoinositide 3-kinase
(PI3K)/Akt is a fundamental signaling pathway that mediates several
cellular processes, including cell proliferation, growth, survival,
and motility (35). Our data
showed that fangchinoline arrested SGC7901 cells during the
G1 phase by decreasing the protein levels of Gsk3β,
CDK2, which act as key regulators of the G1-S
check-point. We also found that fangchinoline promotes SGC7901
apoptosis by decreasing Bcl-2 level and increasing caspase-3,
caspase-9 expression. At the same time, Gsk3β and caspase-3 are the
downstream proteins of PI3K/Akt pathway (36–38).
All of these observations are consistent with the finding that
fangchinoline SGC7901 growth adjustment occurs in the PI3K/Akt
pathways.
In addition to the effect on cell proliferation, we
demonstrated inhibition of fangchinoline on migration and invasion
of gastric cancer cells. One of the key steps in cancer invasion
and metastasis is the degradation of the extracellular matrix. MMP2
has been demonstrated to play important roles in this process
(39,40). Matrix metalloproteinases (MMPs) can
affect tumor invasion and metastasis through the PI3K/Akt pathway
by inducing the expression of MMP2, which plays an important role
in tumor cell migration (41). Our
results showed that fangchinoline significantly suppressed the
migratory and invasive ability of SGC7901 in parallel with
downregulation of MMP9 and MMP2. Therefore, it is reasonable to
speculate that fangchinoline inhibits cell invasion and metastasis
by the PI3K/Akt/MMP2/MMP9 pathway.
In conclusion, fangchinoline was identified as
capable of inhibiting PI3K and its downstream signaling pathways
and suppressing PI3K-mediated SGC7901 behavior including growth,
migration, and invasion. Further testing in experimental models
in vivo is warranted. The results presented in our current
study add to the scope of the exploration and application of PI3K
inhibitors and may offer a novel therapeutic strategy for advanced
metastatic gastric cancer.
References
|
1
|
Corso S, Ghiso E, Cepero V, Sierra JR,
Migliore C, Bertotti A, Trusolino L, Comoglio PM and Giordano S:
Activation of HER family members in gastric carcinoma cells
mediates resistance to MET inhibition. Mol Cancer. 9:1212010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Solit DB, Basso AD, Olshen AB, Scher HI
and Rosen N: Inhibition of heat shock protein 90 function
down-regulates Akt kinase and sensitizes tumors to Taxol. Cancer
Res. 63:2139–2144. 2003.PubMed/NCBI
|
|
3
|
Kazlauskas A and Cooper JA:
Phosphorylation of the PDGF receptor beta subunit creates a tight
binding site for phosphatidylinositol 3 kinase. EMBO J.
9:3279–3286. 1990.PubMed/NCBI
|
|
4
|
Chiang EP, Tsai SY, Kuo YH, Pai MH, Chiu
HL, Rodriguez RL and Tang FY: Caffeic acid derivatives inhibit the
growth of colon cancer: Involvement of the PI3-K/Akt and AMPK
signaling pathways. PLoS One. 9:e996312014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gustin JP, Karakas B, Weiss MB, Abukhdeir
AM, Lauring J, Garay JP, Cosgrove D, Tamaki A, Konishi H, Konishi
Y, et al: Knockin of mutant PIK3CA activates multiple oncogenic
pathways. Proc Natl Acad Sci USA. 106:2835–2840. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kang S, Bader AG and Vogt PK:
Phosphatidylinositol 3-kinase mutations identified in human cancer
are oncogenic. Proc Natl Acad Sci USA. 102:802–807. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang QS, Papanikolaou A, Sabourin CL and
Rosenberg DW: Altered expression of cyclin D1 and cyclin-dependent
kinase 4 in azoxymethane-induced mouse colon tumorigenesis.
Carcinogenesis. 19:2001–2006. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Halilovic E, She QB, Ye Q, Pagliarini R,
Sellers WR, Solit DB and Rosen N: PIK3CA mutation uncouples tumor
growth and cyclin D1 regulation from MEK/ERK and mutant KRAS
signaling. Cancer Res. 70:6804–6814. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dienstmann R, Rodon J, Serra V and
Tabernero J: Picking the point of inhibition: A comparative review
of PI3K/AKT/mTOR pathway inhibitors. Mol Cancer Ther. 13:1021–1031.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Chen J, Wang L, Huang Y, Leng Y
and Wang G: Fangchinoline induces G0/G1 arrest by modulating the
expression of CDKN1A and CCND2 in K562 human chronic myelogenous
leukemia cells. Exp Ther Med. 5:1105–1112. 2013.PubMed/NCBI
|
|
11
|
Nakamura K, Tsuchiya S, Sugimoto Y,
Sugimura Y and Yamada Y: Histamine release inhibition activity of
bisbenzylisoquinoline alkaloids. Planta Med. 58:505–508. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim HS, Zhang YH, Oh KW and Ahn HY:
Vasodilating and hypotensive effects of fangchinoline and
tetrandrine on the rat aorta and the stroke-prone spontaneously
hypertensive rat. J Ethnopharmacol. 58:117–123. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hristova M and Istatkova R:
Complement-mediated antiinflammatory effect of
bisbenzylisoquinoline alkaloid fangchinoline. Phytomedicine.
6:357–362. 1999. View Article : Google Scholar
|
|
14
|
Choi HS, Kim HS, Min KR, Kim Y, Lim HK,
Chang YK and Chung MW: Anti-inflammatory effects of fangchinoline
and tetrandrine. J Ethnopharmacol. 69:173–179. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen YC, Chou CJ, Chiou WF and Chen CF:
Anti-inflammatory effects of the partially purified extract of
radix Stephaniae tetrandrae: Comparative studies of its active
principles tetrandrine and fangchinoline on human polymorphonuclear
leukocyte functions. Mol Pharmacol. 60:1083–1090. 2001.PubMed/NCBI
|
|
16
|
Kim HS, Zhang YH and Yun YP: Effects of
tetrandrine and fangchinoline on experimental thrombosis in mice
and human platelet aggregation. Planta Med. 65:135–138. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsutsumi T, Kobayashi S, Liu YY and
Kontani H: Antihyperglycemic effect of fangchinoline isolated from
Stephania tetrandra Radix in streptozotocin-diabetic mice. Biol
Pharm Bull. 26:313–317. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma W, Nomura M, Takahashi-Nishioka T and
Kobayashi S: Combined effects of fangchinoline from Stephania
tetrandra Radix and formononetin and calycosin from Astragalus
membranaceus Radix on hyperglycemia and hypoinsulinemia in
streptozotocin-diabetic mice. Biol Pharm Bull. 30:2079–2083. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin TY, Lu CW, Tien LT, Chuang SH, Wang
YR, Chang WH and Wang SJ: Fangchinoline inhibits glutamate release
from rat cerebral cortex nerve terminals (synaptosomes). Neurochem
Int. 54:506–512. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gülçin I, Elias R, Gepdiremen A, Chea A
and Topal F: Antioxidant activity of bisbenzylisoquinoline
alkaloids from Stephania rotunda: Cepharanthine and fangchinoline.
J Enzyme Inhib Med Chem. 25:44–53. 2010. View Article : Google Scholar
|
|
21
|
Sekiya N, Hikiami H, Yokoyama K, Kouta K,
Sakakibara I, Shimada Y and Terasawa K: Inhibitory effects of
Stephania tetrandra S. Moore on free radical-induced lysis of rat
red blood cells. Biol Pharm Bull. 28:667–670. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang YH, Fang LH and Ku BS: Fangchinoline
inhibits rat aortic vascular smooth muscle cell proliferation and
cell cycle progression through inhibition of ERK1/2 activation and
c-fos expression. Biochem Pharmacol. 66:1853–1860. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Meng LH, Zhang H, Hayward L, Takemura H,
Shao RG and Pommier Y: Tetrandrine induces early G1 arrest in human
colon carcinoma cells by down-regulating the activity and inducing
the degradation of G1-S-specific cyclin-dependent kinases and by
inducing p53 and p21Cip1. Cancer Res. 64:9086–9092. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun X, Xu R, Deng Y, Cheng H, Ma J, Ji J
and Zhou Y: Effects of tetrandrine on apoptosis and
radiosensitivity of nasopharyngeal carcinoma cell line CNE. Acta
Biochim Biophys Sin (Shanghai). 39:869–878. 2007. View Article : Google Scholar
|
|
25
|
Wang N, Pan W, Zhu M, Zhang M, Hao X,
Liang G and Feng Y: Fangchinoline induces autophagic cell death via
p53/sestrin2/AMPK signalling in human hepatocellular carcinoma
cells. Br J Pharmacol. 164(2b): 731–742. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang CD, Yuan CF, Bu YQ, Wu XM, Wan JY,
Zhang L, Hu N, Liu XJ, Zu Y, Liu GL, et al: Fangchinoline inhibits
cell proliferation via Akt/GSK-3beta/cyclin D1 signaling and
induces apoptosis in MDA-MB-231 breast cancer cells. Asian Pac J
Cancer Prev. 15:769–773. 2014. View Article : Google Scholar
|
|
27
|
Xing Z, Zhang Y, Zhang X, Yang Y, Ma Y and
Pang D: Fangchinoline induces G1 arrest in breast cancer cells
through cell-cycle regulation. Phytother Res. 27:1790–1794. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xing ZB, Yao L, Zhang GQ, Zhang XY, Zhang
YX and Pang D: Fangchinoline inhibits breast adenocarcinoma
proliferation by inducing apoptosis. Chem Pharm Bull (Tokyo).
59:1476–1480. 2011. View Article : Google Scholar
|
|
29
|
Sun HW, Tong SL, He J, Wang Q, Zou L, Ma
SJ, Tan HY, Luo JF and Wu HX: RhoA and RhoC -siRNA inhibit the
proliferation and invasiveness activity of human gastric carcinoma
by Rho/PI3K/Akt pathway. World J Gastroenterol. 13:3517–3522. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schnell O, Krebs B, Wagner E, Romagna A,
Beer AJ, Grau SJ, Thon N, Goetz C, Kretzschmar HA, Tonn JC, et al:
Expression of integrin alphavbeta3 in gliomas correlates with tumor
grade and is not restricted to tumor vasculature. Brain Pathol.
18:378–386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Raynaud FI, Eccles S, Clarke PA, Hayes A,
Nutley B, Alix S, Henley A, Di-Stefano F, Ahmad Z, Guillard S, et
al: Pharmacologic characterization of a potent inhibitor of class I
phosphatidylinositide 3-kinases. Cancer Res. 67:5840–5850. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Leung E, Kim JE, Rewcastle GW, Finlay GJ
and Baguley BC: Comparison of the effects of the PI3K/mTOR
inhibitors NVP-BEZ235 and GSK2126458 on tamoxifen-resistant breast
cancer cells. Cancer Biol Ther. 11:938–946. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pavletich NP: Mechanisms of
cyclin-dependent kinase regulation: Structures of Cdks, their
cyclin activators, and Cip and INK4 inhibitors. J Mol Biol.
287:821–828. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Graña X and Reddy EP: Cell cycle control
in mammalian cells: Role of cyclins, cyclin dependent kinases
(CDKs), growth suppressor genes and cyclin-dependent kinase
inhibitors (CKIs). Oncogene. 11:211–219. 1995.PubMed/NCBI
|
|
35
|
Prasad R, Vaid M and Katiyar SK: Grape
proanthocyanidin inhibit pancreatic cancer cell growth in vitro and
in vivo through induction of apoptosis and by targeting the
PI3K/Akt pathway. PLoS One. 7:e430642012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gan B, Yoo Y and Guan JL: Association of
focal adhesion kinase with tuberous sclerosis complex 2 in the
regulation of s6 kinase activation and cell growth. J Biol Chem.
281:37321–37329. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schlaepfer DD and Hunter T: Signal
transduction from the extracellular matrix - a role for the focal
adhesion protein-tyrosine kinase FAK. Cell Struct Funct.
21:445–450. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao J and Guan JL: Signal transduction by
focal adhesion kinase in cancer. Cancer Metastasis Rev. 28:35–49.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hara T, Miyazaki H, Lee A, Tran CP and
Reiter RE: Androgen receptor and invasion in prostate cancer.
Cancer Res. 68:1128–1135. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Libra M, Scalisi A, Vella N, Clementi S,
Sorio R, Stivala F, Spandidos DA and Mazzarino C: Uterine cervical
carcinoma: Role of matrix metalloproteinases (Review). Int J Oncol.
34:897–903. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kanaki T, Bujo H, Mori S, Yanjuan Z,
Takahashi K, Yokote K, Morisaki N and Saito Y: Functional analysis
of aortic endothelial cells expressing mutant PDGF receptors with
respect to expression of matrix metalloproteinase-3. Biochem
Biophys Res Commun. 294:231–237. 2002. View Article : Google Scholar : PubMed/NCBI
|