Introduction
Lung cancer, which is characterized by high
morbidity and mortality, has become the leading cause of
cancer-related death worldwide. Although the number of novel
anticancer strategies has increased, the 5-year survival rate for
lung cancer remains low, ranging from 9% to 20% (1), indicating the limited effectiveness
of therapy in recent years. The obstacle in improving the lifespan
of lung cancer patients is related to metastasis, which is widely
accepted as the most devastating stage of cancer progression,
following the dissemination of cancer. During the process of
metastasis, tumor cells are required to overcome a series of
rate-limiting barriers. As the primary tumor develops
vascularization, degradation of the extracellular matrix (ECM)
follows, then intravasation, and finally cells arrested in a new
organ form micrometastases after extravasating from the circulating
blood (2). Therein, the subsequent
step that cancer cells extravasated from blood circulation into
secondary organ is the most ineffective step in metastatic
processes and is suggestive of a promising target.
The metastatic processes involve VEGF, MMPs, the
host ECM and several signaling pathways (3). Clinical trials on the inhibition of
VEGF and MMPs have shed light on broad development prospects, but
are inevitably accompanied by side effect due to low selectivity.
Additional evidence suggests that VEGF modulates Livin expression
via mTOR signaling (4), as well as
MMP-2/9 through the ERK1/2 or Notch pathway (5,6).
Moreover, MMPs also mobilize pro-angiogenic factors, for example
VEGF, to promote tumor angiogenesis (7,8), and
Livin downregulate the expression of VEGF and MMP-2 to inhibit cell
growth and invasion through the MAPK signaling (9). Thus, we propose that there are
signaling feedback loops between Livin, VEGF, and MMPs in spite of
the ambiguous mechanism, and synergistic inhibition of VEGF and
MMPs triggered by Livin knock-down can elicit tumor regression in
lung cancer (9).
Livin, known as an inhibitor of apoptosis (IAP), is
selectively overexpressed in specific tumors, and correlates with
survival, prognosis, chemotherapeutic and radiotherapeutic
resistance (10–12). Many studies have discussed the
importance of Livin in the control of invasion of oral squamous
cell carcinoma (13),
laryngohypopharyngeal cancer (14), breast cancer (15), osteosarcoma (16), prostate cancer (17) and digestive system neoplasms
(9,18,19).
We demonstrate that knock-down of Livin induces cell cycle arrest
at the G0/G1 phases and promote apoptotic death through a
caspase-dependent pathway in lung cancer (20,21).
Thus, Livin may be an alternative, efficient and safe target in
lung cancer therapy, based on the fact that Livin is undetectable
in most normally differentiated tissues, with the exception of the
placenta, normal testes, spinal cord and lymph nodes (22).
However, few of the studies have evaluated the
invasive ability in animal models and shown the function of Livin
in lung cancer invasion. Here, we successfully blocked the in
vitro invasion and in vivo metastasis of lung
adenocarcinoma cells in the animal model by silencing Livin, and
demonstrate that Livin affects tumor cell biology through the VEGF
and MMPs signaling pathway.
Materials and methods
Cell culture
The human A549 adenocarcinoma cell line was
purchased from American Type Culture Collection (ATCC) and was
propagated in RPMI-1640 (HyClone Corp., Logan, UT, USA) medium
supplemented with 10% (v/v) fetal bovine serum (FBS) (HyClone
Corp.), 100 U/ml penicillin and 100 g/ml streptomycin, at 37°C in a
humidified atmosphere of 5% CO2 and 95% air.
Construction of lentivectors for specific
silencing of Livin expression and stable transfection
Short hairpin RNA (shRNA) consisting of double
chains of oligonucleotide with complementary sequences were
obtained from Sangon Co. Ltd (Shanghai, China). Based on the Livin
sequence (NM_022161.3), Livn-shDNA sequences were:
5′-GGAGAGAGGTCCAGTCTGA-3′ (sense) and 5′-TCAGACTGGACCTCTCTCCTG-3′
(antisense). The pLKD-CMV-G&PR_U6 plasmid vector (NeuronBiotech
Co. Ltd., Shanghai, China) encoded genes for ampicillin and
puromycin resistance, and an enhanced green fluorescent protein
(eGFP) reporter gene. A293T cells (NeuronBiotech Co. Ltd.) were
co-transfected with the recombinant plasmid and packing plasmid,
envelope plasmid VSVG, and the pseudoviral particles were collected
and the titer was determined. The harvested lentivirus was
transfected into A549 cells using Polybrene reagent (Millipore
Corp., Billerica, USA). The stably transfected cells were picked
out with puromycin, and the green fluorescence intensity was
measured by fluorescence microscopy. The cells transfected with
vectors carrying Livn-shRNA were defined as the KD group (the
experimental group), the cells transfected with vectors as a
negative control were defined as the NC group, and the wild-type
A549 cells were defined as the CO group (the blank control
group).
Real-time fluorescent quantitative
PCR
Total RNAs were extracted using TRIzol reagent
(Invitrogen/Life Technologies, Grand Island, NY, USA) from cells or
harvested tissues. cDNA synthesis was performed using reverse
transcription reagents (Thermo Scientific, Waltham, MA, USA).
According to the manufacturer’s instructions, real-time PCR was
performed using ViiA 7 (ABI, Carlsbad, USA) sequence detection
system with SYBR-Green I Mix (CoWin Biotech Co, Ltd. Beijing,
China). The primers for the genes are listed below: Livin sense:
5′-GCTCTGAGGAGTTGCGTCTG-3′, antisense:
5′-CACACTGTGGACAAAGTCTCTT-3′; VEGF sense:
5′-AGGGCAGAATCATCACGAAGT-3′, antisense: 5′-AGGGTCTCGATTGGATGGCA-3′;
KDR sense: 5′-GGCCCAATAATCAGAGTGGCA-3′, antisense:
5′-CCAGTGTCATTTCCGATCACTTT-3′; MMP2 sense:
5′-TACAGGATCATTGGCTACACACC-3′, antisense:
5′-GGTCACATCGCTCCAGACT-3′; MMP9 sense:
5′-TGTACCGCTATGGTTACACTCG-3′, antisense: 5′-GGCAGGGACAGTTGCTTCT-3′;
GAPDH sense: 5′-CCATGGCACCGTCAAGGCTGA-3′, antisense:
5′-GGGCCATCCACAGTCTTCTGG-3′. The data are presented as ratios
relative to GAPDH levels. Data were analyzed using the comparative
Ct method (2−ΔΔCt).
Western blotting
Cells or tissue were harvested and lysed on ice for
30 min in buffer consisting of 50 mM Tris HCl, pH 8.0, 150 mM
sodium chloride, 5 mM ethylenediaminetetraacetic acid (EDTA), 1%
NP-40, 0.02% NaN3, 50 mM NaF and protease inhibitors (1
mM phenylmethanesulfonyl fluoride (PMSF), 1 μg/ml aprotinin). The
extracts were obtained by centrifugation at 12,000 × g and 4°C for
10 min. The concentration of protein was measured using the BCA
protein assay kit (Thermo Scientific). Equal amounts of proteins
were separated on 12% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto polyvinyldifluoride
(PVDF) membranes. The primary antibodies (Abcam, San Francisco, CA,
USA) against GAPDH (ab110305), Livin (ab97516), VEGF (ab46154),
VEGFR-2 (ab39256), MMP-2 (ab86607) and MMP9 (ab119906) were diluted
according to the instructions of antibodies and incubated overnight
at 4°C. Then, the HRP-conjugated secondary antibody (ab6721) was
added at a dilution ratio of 1:1,000, and incubated at room
temperature for 2 h. The membranes were immunoprobed with
corresponding antibody, and visualized using an enhanced
chemiluminescence reagent (Thermo Scientific).
MTT assay
The exponentially growing A549 cells were seeded in
96-well plates at a density of 8×103 cells/well in media
and left overnight to allow cell adherence. Following incubation
for 1, 2 or 3 days at 37°C, cell viability was measured by adding
20 μl of 5 mg/ml 3-(4,5-dimethylthiazol-2-yl)-2,5-dipheny
ltetrazolium bromide (MTT) to each well, and incubated at 37°C for
a further 4 h, after which the media were substituted with 200 μl
DMSO for formazan crystal dissolution. The absorbance was measured
using a micro-plate reader (Thermo Scientific) at 570 nm. The data
are presented as the percentage of surviving transfected cells vs.
control cells (viability of control cells was considered to be
100%).
Colony-formation assay
Lung cancer cells treated with or without
transfection were plated on 6-well plates at a density of 1,000
cells/well. The culture medium was refreshed after 24 h of
incubation. After 7 days of culture, the colonies were stained with
0.5% of methylene blue, fixed with 4% paraformaldehyde for 2 h, and
counted under a microscope (Olympus Corp., Tokyo, Japan).
Transwell invasion assay
Cell invasive ability was assessed using a 24-well
8.0 μm Transwell chamber (Corning Inc., Corning, NY, USA) coated
with Matrigel (1 mg/ml, BD Biosciences, Franklin Lakes, NJ, USA).
After being starved in serum-free medium for 24 h, 1×105
cells in 200 μl serum-free medium were harvested and seeded in the
top chamber, and 600 μl 10% FBS-RPMI-1640 medium was added to the
lower chamber. Twenty-four hours later, the non-migrated cells were
gently removed with cotton swabs and the migrated cells on the
bottom surface of the membrane were fixed with 4% paraformaldehyde
for 30 min. The cells were then stained with hematoxylin and eosin
(H&E), quantified by manual counting and images were captured
under a light microscope. Rates of cellular invasion through
Matrigel were normalized by that of control cells. Five randomly
chosen fields were analyzed for each group.
Gelatin zymography assay of MMP-2
(Gelatinase A) and MMP-9 (Gelatinase B)
Proteins in the culture supernatant collected in
equal amounts of the cell medium were then separated in 10%
SDS-PAGE gel (0.1% w/v of gelatin) under non-reducing conditions at
4°C. After electrophoresis, the gel was washed in 2.5% v/v of
Triton X-100 for 30 min at room temperature for renaturing with
gentle agitation. The gel was subsequently incubated in assay
buffer (50 mmol/l Tris HCl, 5 mmol/l CaCl2, 0.02%
Brij-35, pH 7.6) for 42 h with gentle shaking. The gel was stained
with solution (0.05% Coomassie blue, 30% methanol, 10% acetic acid)
and sequentially destained. Gelatinolytic bands were detected as
transparent zones against the blue background and densitometric
analysis of the bands was performed using ImagePro Plus software
(Media Cybernetics, Silver Spring, MD, USA).
Enzyme-linked immunosorbent assay
(ELISA)
When showing logarithmic growth, the cells were
collected and plated into 6-well culture plates at a density of
2×105 cells/well. Cell culture supernatants in
serum-free medium were homogenized and harvested 72 h later and
centrifuged at 1000 × g for 20 min. The levels of VEGF were
subsequently determined using commercially available ELISA kits,
product no. SEA143Hu (Uscn Life Science, Wuhan, China), according
to the manufacturer’s instructions.
In vivo tumor xenograft study
The protocol for the animal experiment was approved
from the Institutional Animal Ethics Committee, Experimental Animal
Center of Fujian Medical University, China. All surgery was
performed under anesthesia and efforts were taken to minimize
animal suffering. Seventy-five immunodeficient male BALB/C nude
mice, 4–5 weeks old (SLAC Laboratory Animal Co. Ltd., Shanghai,
China) were randomly divided into three groups with 25 per group:
16 for dynamic monitoring and the remaining 9 for the survival
study. The three groups of A549 cells were resuspended in 0.1 ml
PBS and intravenously injected into the lateral tail vein at a
density of 2.5×106 cells/mouse in the corresponding
animal groups. At 5, 6, 7 and 9 weeks after cell inoculation, 4 of
25 mice were randomly chosen, sacrificed by deep anesthesia and
visualization of cancer-cell expressing eGFP dynamics during lung
metastasis in live mice was performed using the imaging system IVIS
lumina II (Caliper Life Sciences, Boston, MA, USA). Mouse body
weight was measured every two to three days throughout their
lifespan. After macroscopic observation, the harvested tissues from
the three groups of nude mice were weighed, processed and conserved
for the follow-up experiments. Tissues were stained with H&E to
further evaluate lung metastasis generation dynamically and
precisely. The mice in the survival study were euthanized when they
became severely cachectic and the day of euthanasia was considered
the end point of follow-up observation.
Immunohistochemical analysis
Tissues from the sacrificed mice were fixed in
formaldehyde overnight, and then dehydrated and coated with wax.
Tissues sections (3-μm) were prepared and then subjected to
de-paraffining and antigen retrieval. Tissues were blocked with
endogenous peroxidase for 10 min followed by protein block for 10
min at room temperature and were incubated with primary antibody
(Abcam) (1/200 in antibody diluent) for 2 h at 20°C, then
ready-made HRP-conjugated antibody (Abcam) for 30 min at 20°C. The
sections were counterstained with diaminobenzidine (DAB) chromogen
substrate and hematoxylin, and the presence of a brown precipitate
indicating immunoreactivity was examined under a microscope.
Assuming that the intensity of immunoreactivity was correlated with
the level of gene expression, semi-quantitative analysis of tissue
samples was conducted using ImagePro Plus software. The images were
captured by microscopy.
Statistical analysis
SPSS 19.0 was used for data analysis. The data are
expressed as means ± SD from independent experiments. Statistical
significance was determined using the Student’s two-tailed t-test
in two groups and one way ANOVA in multiple groups. Kaplan-Meier
survival curves and P-values of group comparisons were conducted
for survival analysis. A P-value of <0.05 was considered
statistically significant.
Results
A549 cells were stably transfected with
Livin specific shRNA expression vector
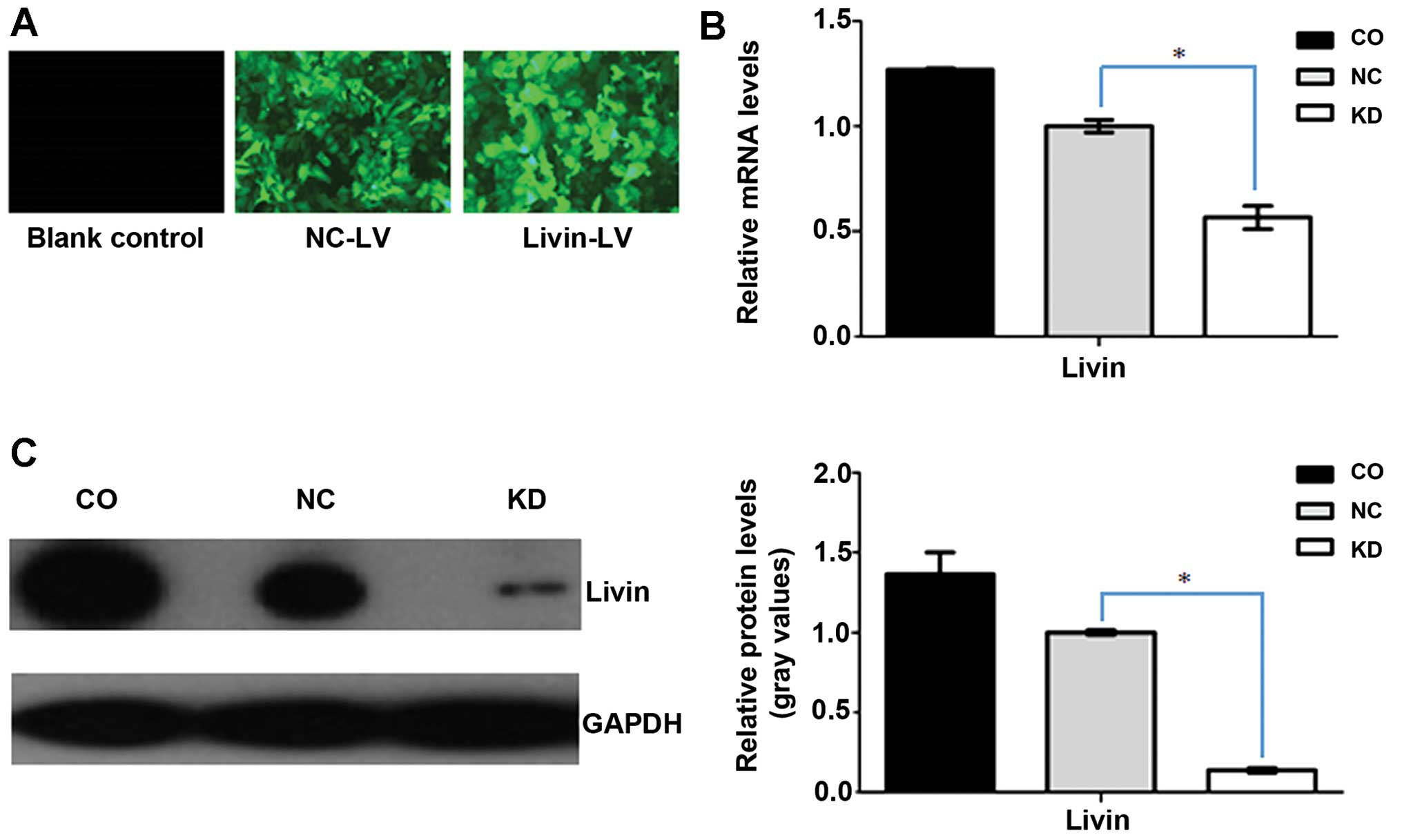
The rate of effective transfection in the three
groups of cells was monitored by fluorescence microscopy. Over 95%
of cells transfected with lentiviral vectors showed green
fluorescence in the KD and NC groups, however, no GFP expression
was observed in the wild-type A549 cells, which indicated
successful transfection (Fig. 1A).
To determine the effect of Livin-shRNA in A549-RNAi cells, the mRNA
levels of Livin were analyzed by real-time PCR assay. The
GAPDH-normalized Livin mRNA expression is shown in Fig. 1B. As, expected, Livin mRNA
expression in A549-RNAi cells was the lowest, clearly showing the
constitutive downregulation of Livin. There were significant
differences between A549-RNAi cells and both NC-LV and
non-transfected A549 cells. The efficient silencing of Livin was
confirmed by western blotting. Densitometric measurements revealed
that the expression of Livin protein decreased sharply in A549-RNAi
cells, and was reduced by 86% compared with wild-type A549 cells
(Fig. 1C).
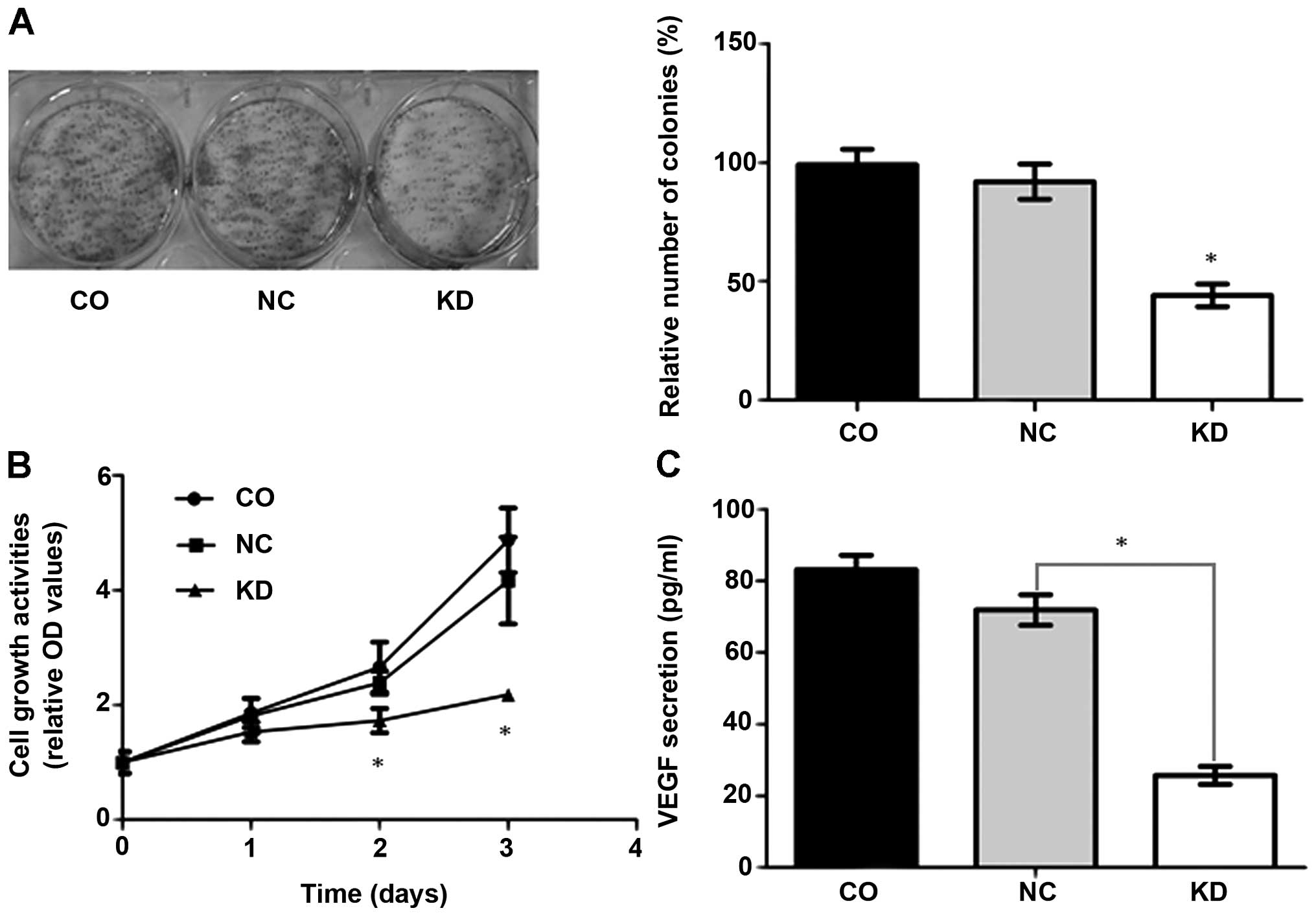
Effect of Livin small hairpin RNA on cell
proliferation and cell growth
The effect of Livin downregulation on transfected
cell proliferation was determined by colony formation assay. As
shown in Fig. 2A, a significantly
lower colony number was observed in A549-RNAi cells than in NC-LV
or blank control cells (P<0.001). The colony number in A549-RNAi
cells was reduced by 52%. To gain further insight into the effect
of Livin shRNA on A549 cell growth, the metabolic activity of these
cells was quantified by MTT assay. Additional comparisons showed
that growth of the stably expressing Livin shRNA cells was slower.
Compared with the vector control or blank control cells,
significant differences were noted in growth rates on the second
day (P<0.005), which were amplified on the third day
(P<0.001). The growth rates of the vector control cells were not
significantly different from the blank control A549 cells (Fig. 2B).
In addition, to detect whether Livin knock-down
affected extracellular VEGF, ELISA was performed to quantify the
amount of VEGFA secreted into the culture media. The inhibitory
effect on VEGFA protein levels was demonstrated by a significant
decrease in VEGFA concentration in the media containing A549-RNAi
cells (25.7±2.5 pg/ml) compared with the vector control group
(71.9±4.3 pg/ml) and the control group (83.1±4.1 pg/ml)
(P<0.001) (Fig. 2C). Real-time
PCR and western blot analysis were carried out to investigate
intracellular VEGF expression. The data, summarized in Fig. 3C and D, indicated a marked decrease
in VEGF protein expression in A549-RNAi cells. The data described
above suggest that knock-down of Livin suppressed VEGFA secretion,
as well as intracellular VEG F expression. In contrast, no
significant differences (P>0.05) in the expression level between
the vector control and blank control were observed.
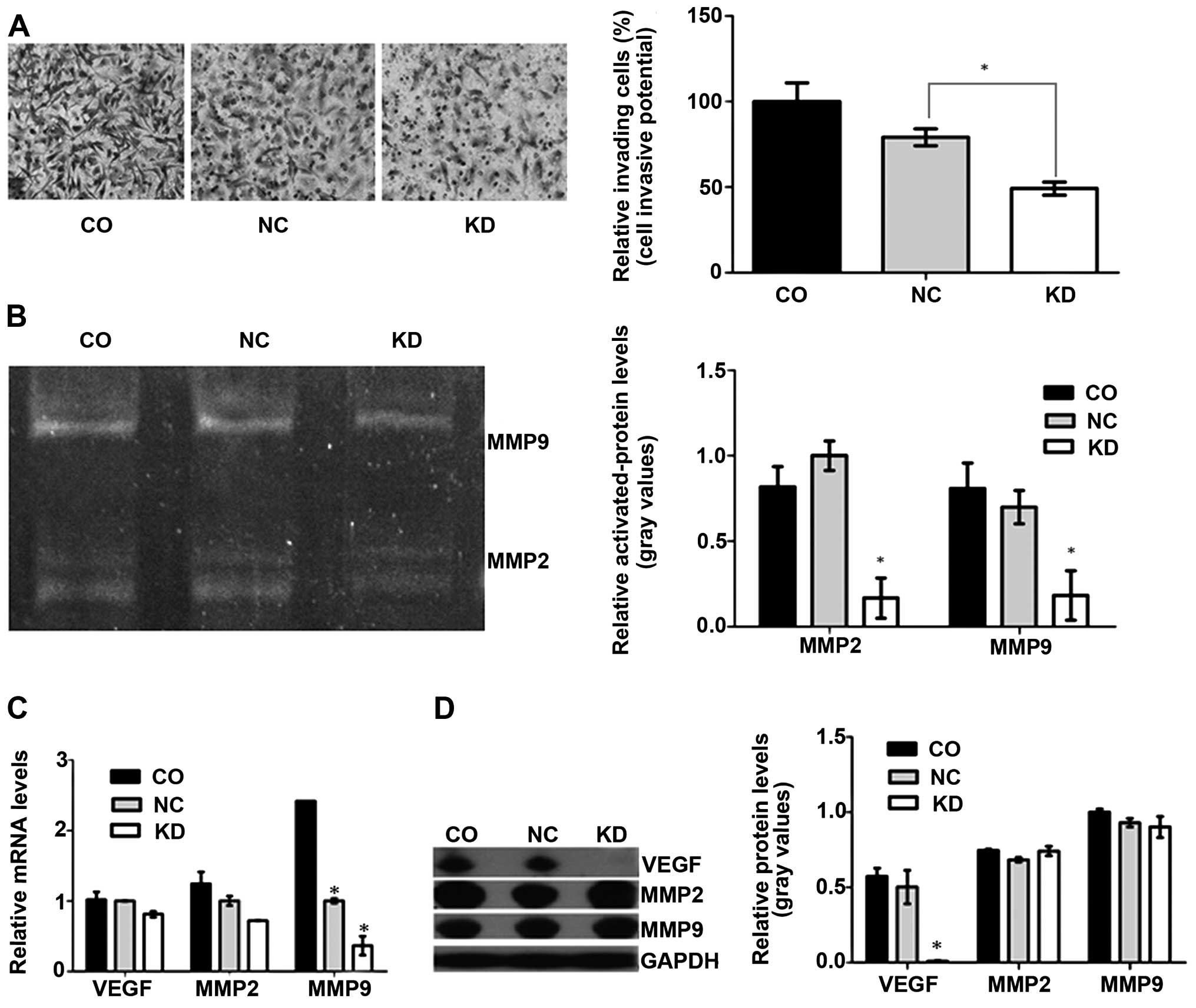
Knock-down of Livin suppresses cell
invasion in vitro through the MMPs signaling pathway
To gain knowledge on the effect of Livin knock-down
on A549 cell invasion, we evaluated the invasive ability of cells
using a Transwell assay. The results indicated that the number of
A549-RNAi cells which passed through the Matrigel-coated membranes
was significantly less than that of the negative control or blank
control A549 cells (P<0.001) (Fig.
3A). We performed gelatin zymography to determine whether Livin
knock-down inhibited cell invasion via downregulation of activated
forms of MMP-2/-9. The marked decrease in invasive ability of the
A549-RNAi cells was further confirmed, and statistically
significant differences in MMP-2 and MMP-9 activities, which
declined by >50%, were observed between the two groups of cells
(Livin-LV, vector-LV) (Fig. 3B).
No differences in activated MM P-2 and MMP-9 between the negative
control and blank control group were observed. To detect total MMP
expression, real-time PCR and western blot analysis were conducted.
Fig. 3C and D show significant
differences in total mRNA level among the three groups, while no
significant differences in total MMP protein level was noted.
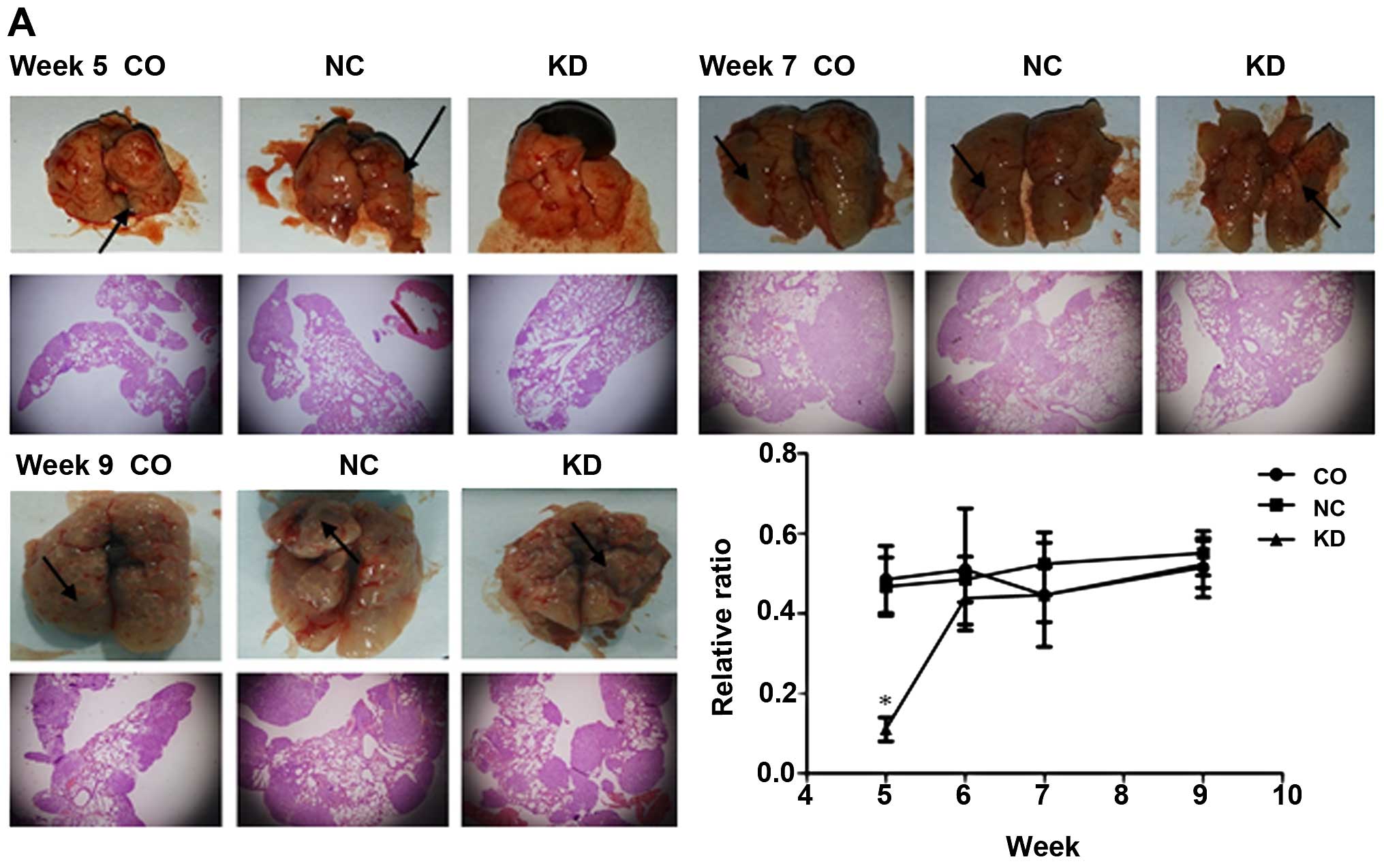
Silencing of Livin preventes the
formation of tumor nodules resulting from impaired metastasis and
neoangiogenesis in vivo
To explore Livin gene function in the development of
lung cancer in vivo, the mice with successfully constructed
lung cancer were sacrificed. Lung metastasis occurred in 100% of
the mice injected with tumor cells. Five weeks after cell
inoculation, scattered tumor nodules which showed a soft texture
and a greyish-white color in the NC and CO groups were occasionally
seen by the naked eye in comparison with the KD group which showed
no visible tumor nodules. The already formed tumor nodules were of
unequal size and unevenly distributed in the lung under the
microscope. The pathological result showed that the relative ratio
of the areas with pulmonary nodules to the total lung tissue area
was the least in the KD group. In the other two groups, the ratio
was not significantly different (P>0.05). At 6, 7 and 9 weeks
after cell inoculation, gross view and microscopic pathology showed
no significant differences between the three groups. Based on tumor
formation in the lungs at the different time points, week seven was
considered as the stationary phase in which the size of tumors in
the experimental group changed slightly, in contrast to the period
from week five to week six (Fig.
4A).
To determine the transcription and translation of
Livin and associated genes in vivo, we used real-time PCR
and western blot analysis. During genomic and proteomic detection
of either GAPDH or Livin, to avoid cross-reaction between
cancer nodules derived from human lung cancer cells and normal lung
tissues of mice, we chose human-specific primers and antibodies.
Five weeks after cell inoculation, the human specific GAPDH and
Livin mRNA, as well as their protein content in the mixed tissue in
the KD group showed significant differences compared with the NC
and CO group (P<0.011). These differences were consistent with
the relative ratio of the areas with tumor nodules which could
represent the volume of tumor nodules formed in the lungs. Analysis
of the human specific GAPDH-normalized Livin mRNA and protein
levels in the KD group showed no significant differences compared
with those in the NC and CO group (Fig. 4B). Taking the distinct areas of
formed pulmonary nodules into consideration, we further analyzed
the models to confirm the differences between the three groups at 7
weeks after cell inoculation, at which time the pulmonary nodules
were assumed to be stably formed. As expected, no differences were
observed (P>0.05) (Fig.
4C).
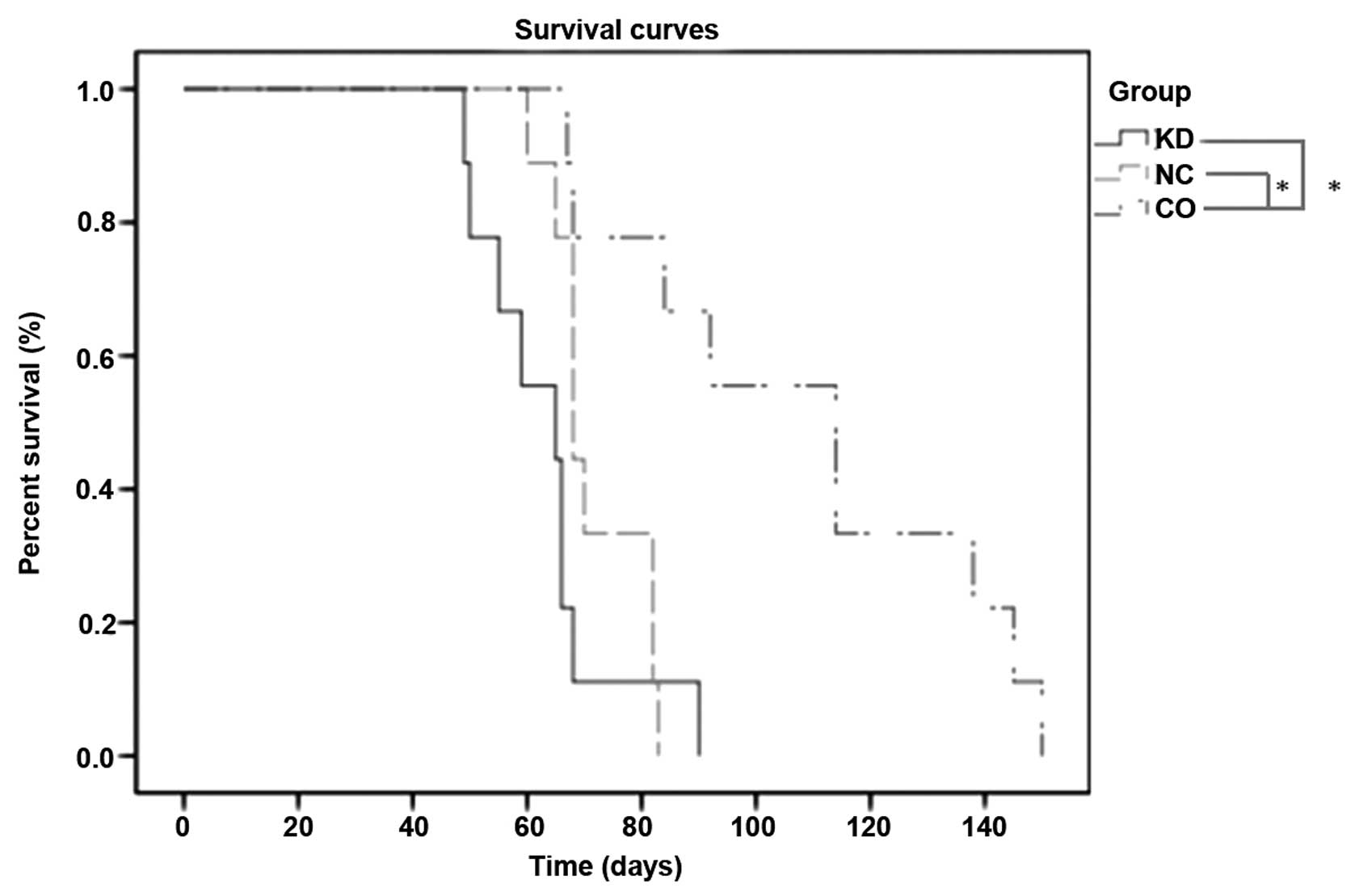
To further evaluate the anti-metastatic activity of
Livin, we conducted a survival study. Unexpectedly, mice inoculated
with wild-type A549 cells had a significantly longer mean survival
time (114 days) compared with 65 days for mice inoculated with
A549-RNAi cells and 68 days for mice inoculated with NC-LV cells
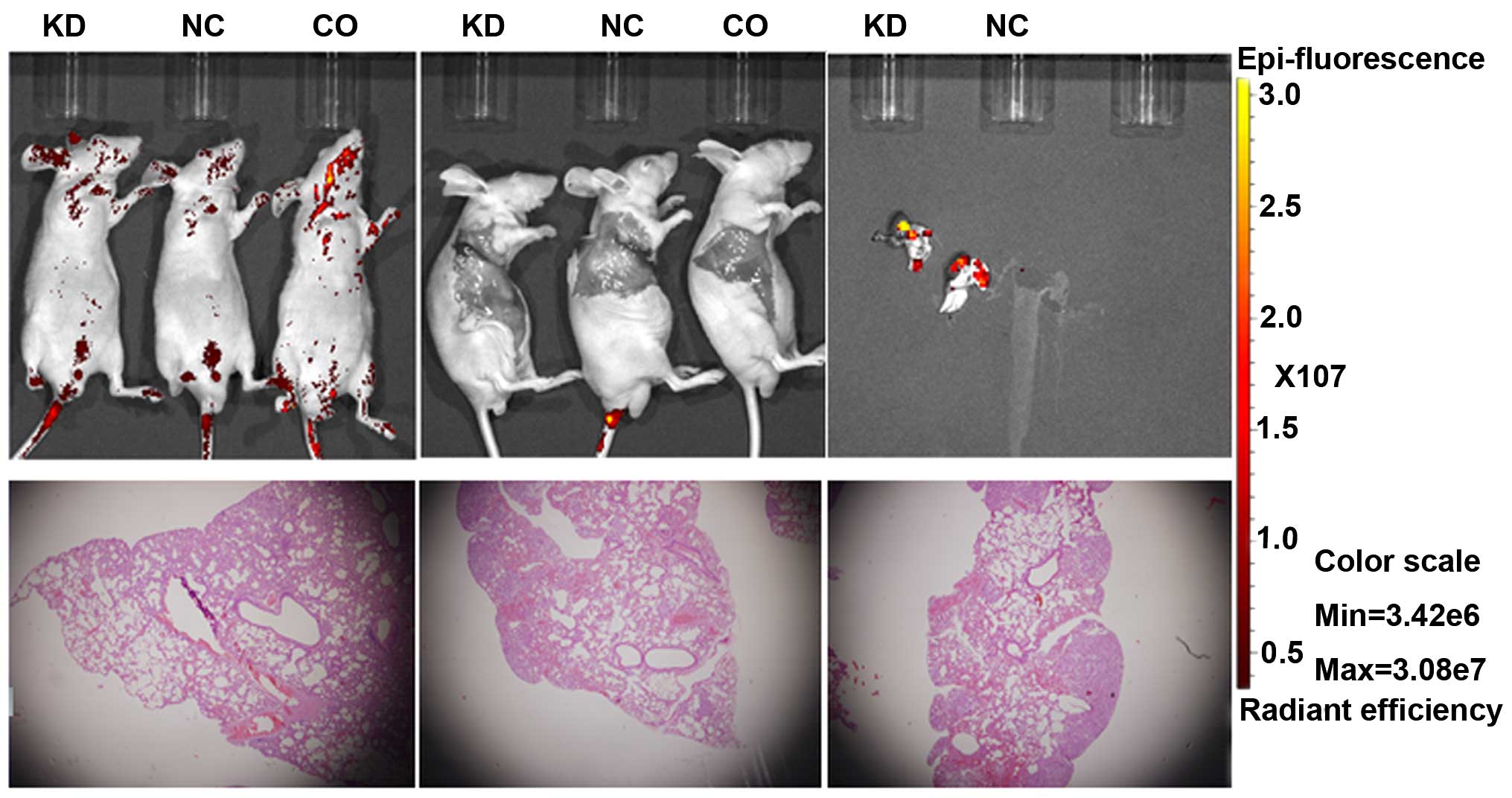
(P<0.001) (Fig. 5). In
vivo imaging after intravenous injection showed no fluorescence
in the lungs, although microscopic pathology confirmed the presence
of lung metastasis. There were only false positive areas of
increased uptake (Fig. 6). The
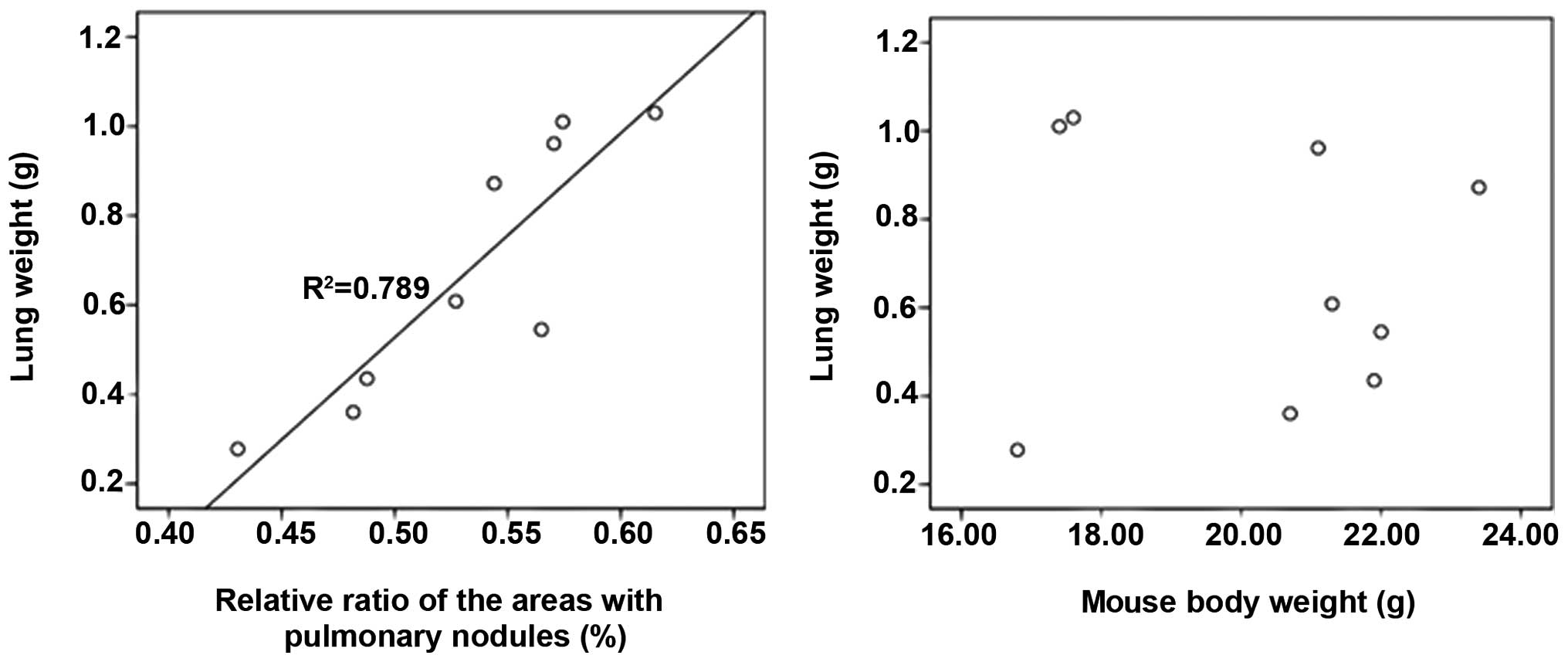
correlation of the lung weight, the mouse body weight and the
relative ratio of the areas with pulmonary nodules was analyzed to
figure out whether the weight of lungs could reflect the volume of
tumor nodules formed in the lungs, as seen in Fig. 7, there was a positive correlation
between the lung weight and the relative ratio of the areas with
pulmonary nodules (R2=0.789), however, no correlation
exists between the lung weight and the mouse body weight (nine mice
were randomly selected).
Discussion
Livin, a novel IAP family protein, plays crucial
roles in the regulation of cell proliferation, cell cycle and
apoptotic death (20,21,23).
Increasing number of studies reveal that Livin can inhibit
apoptosis induced by a variety of stimulus, while it is
specifically cleaved by caspases on a strong apoptotic stimulus to
produce a truncated protein which inversely harbors the
death-promoting activity, indicating a dual role of Livin in cell
biology (10,24–26).
The subcellular localization of Livin can be an alternative
mechanism that regulates the balance between the anti- and
pro-apoptotic activity of Livin (27). However, Livin is involved in
networks of closely related molecules and factors, and regulates
the cell function by numerous cross-communication methods, which
has made it a promising target for the development of potential
gene therapy. Thus, we developed a novel experimental animal model
to examine the effect of Livin in lung cancer.
The safety and efficiency of the lentiviral vector
transduction system, which did not alter the biological
characteristics of the cells or survival time of the animal models,
caused substantial expression of genes over 20 weeks (28–30).
In the current study, using the Livin specific shRNA expression
vector for gene knock-down, we successfully achieved stable
suppression of Livin expression in A549 cells. Downregulation of
Livin expression not only led to slower cell proliferation and
growth rate, but also regulated downstream gene expression
resulting in inhibition of cell invasion. In this study, it was
found out that genetic downregulation of Livin inhibited cell
proliferation and growth by causing a marked decrease in VEGF
(almost to 0% at the translation level). The mRNA expression which
was not in line with the protein expression suggested that
transcription accounted for only a minor fraction of gene
expression. VEGF, also known as VEGFA, has a number of functions in
tumor progression. According to the origin of VEGF, it is generally
classified as autocrine VEGF which participated in cell survival,
and was identified as a contributor to the size of the cancer stem
cell pool (31) and paracrine VEGF
which conveyed critical signals for cell biological behavior
including proliferation and permeability responses (32). In the present study consistent
findings were recorded by real-time PCR and western blotting, Livin
knock-down interfered with both forms of VEGF causing a sharp
reduction in VEGF secretion by A549 cells in vitro. The
MMPs, which are key nodal proteases and control the protease web,
consist of a large family with nearly 30 members (33). MMP2 and MMP9, known as gelatinases,
are considered to be involved in cell invasion. We showed that
total expression levels of MMPs were indistinguishable on the basis
of Livin silencing, in sharp contrast to the change in the
activated forms of MMPs. Although, the three main levels including
transcription, proenzyme activation and inhibition control MMP
proteolytic activity in the cells (34), the invasion ability of lung cancer
cells is attributed to the transformation of MMPs from pro-MMPs to
activated forms other than alteration in the total MMP expression.
Thus, specific cells have their own distinguishable mechanism in
the mediation of genome and proteome.
BALB-nude mice, which congenitally lack
immunological T cells have a leading role in anticancer immunity,
but are characterized by abundant B cells and inherited immune
components including cytokines and natural killer cells. In the
present study, we used BALB-nude mice as in vivo models for
a better evaluation of the interactions between xenotransplanted
tumors and host defense due to their immunodeficiency background.
There are many approaches for the analysis of metastases:
subcutaneous, intraperitoneal, orthotopic, intra-cardiac and
intravenous injection. The intravenous approach which was used in
our study was customized for the lung metastasis assay based on the
rationale that the pulmonary vascular bed acted as the first
station for tumor cells injected via the mouse lateral tail vein.
Lung cancer cells in the blood circulation were blocked in the
pulmonary capillary bed, and penetrated the blood vessel wall
mimicking the metastasis process after the intravasation step.
Subsequently, when the cancer cells arrested in the lung, the
tumorigenesis process began (2).
In this study, the lentivirus vector transfected cells were used
instead of the separate application of vectors and tumor cells,
because the extraneous vectors in the circulating blood might
motivate the animal defense to reversely neutralize the imminent
transfection (35).
The animals which were euthanized at different time
points, enabled a dynamic follow-up of the way Livin functioned
in vivo. Five weeks after tumor implantation, a significant
decrease in tumor burden in the experimental group strongly
indicated anti-metastasis and tumorigenesis inhibition. This was
ascribed to several reasons e.g., reduced MMPs expression in the
treatment group meant less metastatic ability, and resulted in
fewer cells settling in the lung (36). Following this, cells proliferated,
but the LV-shRNA transfected cells impeded tumor expansion because
of the impairment of tumor neoangiogenesis resulting from VEGF
inhibition. Further analysis of human specific GAPDH-normalized
Livin expression and the correlated genes in Livin modulated
signal-transduction pathways indicated reduced transfection
efficacy, i.e., transfected tumor cells which accounted for over
95% of the total cells injected into the mice were eliminated due
to decreased invasive ability, and the minority of untransfected
cells, which were arrested in the lung, expanded and led to
oncogenesis. Moreover, the gradual development of tumors from the
outer to the inner areas of the lungs seen on microscopic pathology
(Fig. 4A) was regarded as evidence
that tumor cells were arrested in the microvascular bed. In the
three groups, the lung cancer stage was not uniform, the stage of
the neoplastic process affected by Livin-targeted therapy in the
experimental group tended to be earlier. Overall, an important
conclusion drawn from our studies was that Livin was characterized
by stage-independent expression in lung adenocarcinoma cells
(37) and the growth of lung
cancer followed an S-shaped curve (Fig. 4A). With regard to vectors encoding
eGFP, the fluorescence microscopy for their real-time imaging was
feasible (38–40). In our work, we used lentiviral
vectors encoding eGFP as a dynamic fluorescent tracer but obtained
a negative outcome, although we further opened skin-flap window and
cut out the lungs for fluorescent imaging. One probable explanation
was that the low aggregation of tumor nodules accounted for the
negative results and the IVIS system was not fit for fluorescence
imaging of tumors inside the mouse body (41,42),
but rather the bioluminescent imaging (43). In the subcutaneous models, the
reduction in tumor weight and tumor volume followed Livin gene
silencing (9,21). In the intravenous models, the
xenografts grew larger as the tumor progressed, and the lung weight
was correlated with the relative ratio of the areas with tumor
nodules which represented the volume of the tumor in situ,
although the interaction between lung cancer and the host might
influence the lung weight. Taken together, we demonstrated that the
experimental models were reliable and reproducible in our work. In
addition, we postulate that prolonged survival should have been
elicited in the experimental group because our previous study
demonstrated that the local injection of lentivirus-delivered Livin
shRNA effectively suppressed lung carcinoma development with minor
adverse reactions. It is essential to elucidate whether eGFP, in
addition to other components of the heterogeneous tumor cell
lysates, are toxic to cells and amplify the arousal responses to
antigen in the anti-neoplastic inflammatory process during the
intravenous injection approach (44,45)
and vectors encoding Luciferase may be an alternative.
In conclusion, our findings indicate that knock-down
of Livin inhibits cell growth and invasion through blockade of the
VEGF and MMPs pathways in lung cancer cells in vitro, and
inhibits tumorigenesis and metastasis of lung cancer in
vivo, suggesting that Livin may be an appropriate anticancer
target for the treatment of lung cancer. This is the first
demonstration that tumorigenesis and metastasis of lung cancer can
be investigated in a reliable and reproducible experimental animal
model in a time-dependent manner.
Acknowledgements
This study was supported by a grant (no. 2011J01130)
from the Natural Science Foundation of Fujian Province, China and a
grant (no. WKJ-FJ-17) from the scientific research fund of the
National Board of health and family planning, China. The authors
would like to compliment the staff of the Pathology Department in
Fujian Provincial Hospital. The authors would also like to thank
Cardiovascular Key Laboratory of Fujian Province, Experimental
Animal Center of Fujian Medical University and the CDC of Fujian
Province, China, for providing us with necessary conditions and
facilities to complete this project.
Abbreviations:
|
RNAi
|
RNA interference
|
|
ShRNA
|
small hairpin RNA
|
|
VEGF
|
vascular endothelial growth factor
|
|
MMPs
|
matrix metalloproteinases
|
|
mTOR
|
mechanistic target of rapamycin
|
|
ERK1/2
|
extracellular signal-regulated kinase
1/2
|
|
MAPK
|
mitogen-activated protein kinase
|
References
|
1
|
Coleman MP, Forman D, Bryant H, Butler J,
Rachet B, Maringe C, Nur U, Tracey E, Coory M, Hatcher J, et al;
ICBP Module 1 Working Group. Cancer survival in Australia, Canada,
Denmark, Norway, Sweden, and the UK, 1995–2007 (the International
Cancer Benchmarking Partnership): An analysis of population-based
cancer registry data. Lancet. 377:127–138. 2011. View Article : Google Scholar :
|
|
2
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bissell MJ and Radisky D: Putting tumours
in context. Nat Rev Cancer. 1:46–54. 2001. View Article : Google Scholar
|
|
4
|
Yan B, Kong M, Chen S and Chen YH: VEGF
stimulation enhances Livin protein synthesis through mTOR
signaling. J Cell Biochem. 111:1114–1124. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Funahashi Y, Shawber CJ, Sharma A,
Kanamaru E, Choi YK and Kitajewski J: Notch modulates VEGF action
in endothelial cells by inducing Matrix Metalloprotease activity.
Vasc Cell. 3:22011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shan B, Li W, Yang SY and Li ZR: Estrogen
up-regulates MMP2/9 expression in endometrial epithelial cell via
VEGF-ERK1/2 pathway. Asian Pac J Trop Med. 6:826–830. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Enciso JM, Gratzinger D, Camenisch TD,
Canosa S, Pinter E and Madri JA: Elevated glucose inhibits
VEGF-A-mediated endocardial cushion formation: Modulation by
PECAM-1 and MMP-2. J Cell Biol. 160:605–615. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ebrahem Q, Chaurasia SS, Vasanji A, Qi JH,
Klenotic PA, Cutler A, Asosingh K, Erzurum S and Anand-Apte B:
Cross-talk between vascular endothelial growth factor and matrix
metalloproteinases in the induction of neovascularization in vivo.
Am J Pathol. 176:496–503. 2010. View Article : Google Scholar :
|
|
9
|
Ou JM, Ye B, Qiu MK, Dai YX, Dong Q, Shen
J, Dong P, Wang XF, Liu YB, Quan ZW, et al: Knockdown of Livin
inhibits growth and invasion of gastric cancer cells through
blockade of the MAPK pathway in vitro and in vivo. Int J Oncol.
44:276–284. 2014.
|
|
10
|
Lazar I, Perlman R, Lotem M, Peretz T,
Ben-Yehuda D and Kadouri L: The clinical effect of the inhibitor of
apopotosis protein livin in melanoma. Oncology. 82:197–204. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haferkamp A, Bedke J, Vetter C, Pritsch M,
Wagener N, Buse S, Crnkovic-Mertens I, Hoppe-Seyler K,
Macher-Goeppinger S, Hoppe-Seyler F, et al: High nuclear Livin
expression is a favourable prognostic indicator in renal cell
carcinoma. BJU Int. 102:1700–1706. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun JG, Liao RX, Chen ZT, Wang ZX, Zhang
Q, Hu YD and Wang DL: Gene transfection of Livin isoforms into A549
cell line and its effect on cell growth and sensitivity to
chemotherapy and radiotherapy. Zhonghua Jie He He Hu Xi Za Zhi.
28:836–840. 2005.(In Chinese).
|
|
13
|
Lee DH, Yoon TM, Kim SA, Park YL, Lee KH,
Lim SC, Lee JK and Joo YE: Relationship between expression of Livin
and the biological behavior of human oral squamous cell carcinoma.
Oncol Rep. 32:2453–2460. 2014.PubMed/NCBI
|
|
14
|
Yoon TM, Kim SA, Lee DH, Lee JK, Park YL,
Lee KH, Chung IJ, Joo YE and Lim SC: Expression of Livin and the
inhibition of tumor progression by Livin silencing in
laryngohypopharyngeal cancer. In Vivo. 28:751–759. 2014.PubMed/NCBI
|
|
15
|
Li F, Yin X, Luo X, Li HY, Su X, Wang XY,
Chen L, Zheng K and Ren GS: Livin promotes progression of breast
cancer through induction of epithelial-mesenchymal transition and
activation of AKT signaling. Cell Signal. 25:1413–1422. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li X, Fan S, Li L, Wang L, Fan G, Zhao Q
and Li Y: RNA interference-mediated knockdown of Livin suppresses
cell proliferation and invasion and enhances the chemosensitivity
to cisplatin in human osteosarcoma cells. Int J Oncol. 43:159–168.
2013.PubMed/NCBI
|
|
17
|
Chen F, Yang D, Che X, Wang J, Li X, Zhang
Z, Chen X and Song X: Livin mediates tumor cell invasion in the
DU-145 cell line via NF-κB. Oncol Rep. 27:2010–2016.
2012.PubMed/NCBI
|
|
18
|
Cho SB, Lee WS, Park YL, Kim N, Oh HH, Kim
MY, Oak CY, Chung CY, Park HC, Kim JS, et al: Livin is associated
with the invasive and oncogenic phenotypes of human hepatocellular
carcinoma cells. Hepatol Res. 45:448–457. 2015. View Article : Google Scholar
|
|
19
|
Myung DS, Park YL, Chung CY, Park HC, Kim
JS, Cho SB, Lee WS, Lee KH, Lee JH and Joo YE: Expression of Livin
in colorectal cancer and its relationship to tumor cell behavior
and prognosis. PLoS One. 8:e732622013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen YS, Li HR, Lin M, Chen G, Xie BS, Xu
NL and Lin LF: Livin abrogates apoptosis of SPC-A1 cell by
regulating JNKI signaling pathway. Mol Biol Rep. 37:2241–2247.
2010. View Article : Google Scholar
|
|
21
|
Chen YS, Li HR, Miao Y, Chen WY, Li YT,
Wang GQ and Wu ZC: Local injection of lentivirus-delivered livin
shRNA suppresses lung adenocarcinoma growth by inducing a G0/G1
phase cell cycle arrest. Int J Clin Exp Pathol. 5:796–805.
2012.
|
|
22
|
Liu B, Han M, Wen JK and Wang L:
Livin/ML-IAP as a new target for cancer treatment. Cancer Lett.
250:168–176. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao X, Yuan Y, Zhang Z, Feng X, Zhang J,
Yuan X and Li J: Effects of shRNA-silenced livin and survivin on
lung cancer cell proliferation and apoptosis. J BUON. 19:757–762.
2014.PubMed/NCBI
|
|
24
|
Nachmias B, Ashhab Y, Bucholtz V, Drize O,
Kadouri L, Lotem M, Peretz T, Mandelboim O and Ben-Yehuda D:
Caspase-mediated cleavage converts Livin from an antiapoptotic to a
proapoptotic factor: Implications for drug-resistant melanoma.
Cancer Res. 63:6340–6349. 2003.PubMed/NCBI
|
|
25
|
Abd-Elrahman I, Hershko K, Neuman T,
Nachmias B, Perlman R and Ben-Yehuda D: The inhibitor of apoptosis
protein Livin (ML-IAP) plays a dual role in tumorigenicity. Cancer
Res. 69:5475–5480. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shiloach T, Berens C, Danke C, Waiskopf O,
Perlman R and Ben-Yehuda D: tLivin displays flexibility by
promoting alternative cell death mechanisms. PLoS One.
9:e1010752014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nachmias B, Lazar I, Elmalech M,
Abed-El-Rahaman I, Asshab Y, Mandelboim O, Perlman R and Ben-Yehuda
D: Subcellular localization determines the delicate balance between
the anti- and pro-apoptotic activity of Livin. Apoptosis.
12:1129–1142. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kafri T, Blömer U, Peterson DA, Gage FH
and Verma IM: Sustained expression of genes delivered directly into
liver and muscle by lentiviral vectors. Nat Genet. 17:314–317.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Piacibello W, Bruno S, Sanavio F, Droetto
S, Gunetti M, Ailles L, Santoni de Sio F, Viale A, Gammaitoni L,
Lombardo A, et al: Lentiviral gene transfer and ex vivo expansion
of human primitive stem cells capable of primary, secondary, and
tertiary multi-lineage repopulation in NOD/SCID mice. Nonobese
diabetic/severe combined immunodeficient. Blood. 100:4391–4400.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tabatabai G, Hasenbach K, Herrmann C,
Maurer G, Möhle R, Marini P, Grez M, Wick W and Weller M: Glioma
tropism of lentivirally transduced hematopoietic progenitor cells.
Int J Oncol. 36:1409–1417. 2010.PubMed/NCBI
|
|
31
|
Goel HL and Mercurio AM: VEGF targets the
tumour cell. Nat Rev Cancer. 13:871–882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee S, Chen TT, Barber CL, Jordan MC,
Murdock J, Desai S, Ferrara N, Nagy A, Roos KP and Iruela-Arispe
ML: Autocrine VEGF signaling is required for vascular homeostasis.
Cell. 130:691–703. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Overall CM and Kleifeld O: Tumour
microenvironment -opinion: Validating matrix metalloproteinases as
drug targets and anti-targets for cancer therapy. Nat Rev Cancer.
6:227–239. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Overall CM and López-Otín C: Strategies
for MMP inhibition in cancer: Innovations for the post-trial era.
Nat Rev Cancer. 2:657–672. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shiau AL, Teo ML, Chen SY, Wang CR, Hsieh
JL, Chang MY, Chang CJ, Chao J, Chao L, Wu CL, et al: Inhibition of
experimental lung metastasis by systemic lentiviral delivery of
kallistatin. BMC Cancer. 10:2452010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liang Z, Yoon Y, Votaw J, Goodman MM,
Williams L and Shim H: Silencing of CXCR4 blocks breast cancer
metastasis. Cancer Res. 65:967–971. 2005.PubMed/NCBI
|
|
37
|
Hartman ML and Czyz M: Anti-apoptotic
proteins on guard of melanoma cell survival. Cancer Lett.
331:24–34. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rashidi B, Moossa AR and Hoffman RM:
Specific route mapping visualized with GFP of single-file streaming
contralateral and systemic metastasis of Lewis lung carcinoma cells
beginning within hours of orthotopic implantation [correction of
implantion]. J Cell Biochem. 114:1738–1743. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yamamoto N, Tsuchiya H and Hoffman RM:
Tumor imaging with multicolor fluorescent protein expression. Int J
Clin Oncol. 16:84–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li X, Wang J, An Z, Yang M, Baranov E,
Jiang P, Sun F, Moossa AR and Hoffman RM: Optically imageable
metastatic model of human breast cancer. Clin Exp Metastasis.
19:347–350. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li W, Li H, Li J, Wang H, Zhao H, Zhang L,
Xia Y, Ye Z, Gao J, Dai J, et al: Self-assembled supramolecular
nano vesicles for safe and highly efficient gene delivery to solid
tumors. Int J Nanomed. 7:4661–4677. 2012. View Article : Google Scholar
|
|
42
|
Cool SK, Breyne K, Meyer E, De Smedt SC
and Sanders NN: Comparison of in vivo optical systems for
bioluminescence and fluorescence imaging. J Fluoresc. 23:909–920.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Burrell-Saward H, Rodgers J, Bradley B,
Croft SL and Ward TH: A sensitive and reproducible in vivo imaging
mouse model for evaluation of drugs against late-stage human
African trypanosomiasis. J Antimicrob Chemother. 70:510–517. 2015.
View Article : Google Scholar
|
|
44
|
Liu HS, Jan MS, Chou CK, Chen PH and Ke
NJ: Is green fluorescent protein toxic to the living cells? Biochem
Biophys Res Commun. 260:712–717. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ejeskär K, Fransson S, Zaibak F and
Ioannou PA: Method for efficient transfection of in
vitro-transcribed mRNA into SK-N-AS and HEK293 cells: Difference in
the toxicity of nuclear EGFP compared to cytoplasmic EGFP. Int J
Mol Med. 17:1011–1016. 2006.PubMed/NCBI
|